





文摘
钾通道TWIK-related酸敏感的钾(任务)1通道,连同其他的钾离子通道,控制着低肺动脉的语气。Src家族酪氨酸激酶(SrcTK)可能控制钾通道功能在人类肺动脉平滑肌细胞(hPASMCs)氧张力的变化和临床使用SrcTK抑制剂导致部分可逆的肺动脉高压。
本研究旨在确定SrcTK的角色在hPASMCs低氧诱导的抑制钾离子通道。
我们表明,SrcTK co-localised task 1频道。抑制SrcTK减少钾电流密度,导致相当大的两极化,而激活SrcTK膜片箝记录增加钾电流。中度缺氧和SrcTK抑制剂减少酪氨酸磷酸化状态的任务1频道。缺氧也能减少phospho-SrcTK水平(tyr419)和减少任务1的co-localisation通道和phospho-SrcTK。相应,缺氧减少任务1电流SrcTK抑制之前而不是之后,包括鼠肺,SrcTK抑制剂增加肺动脉压力。
我们建议SrcTK控制钾离子通道是一个关键因素,作为辅因子设置hPASMCs消极静止膜电位和低静息肺血管的基调。
在肺动脉血管张力调节部分的静止膜电位的肺动脉平滑肌细胞(PASMCs)。在所有的细胞中,钾离子通道是静止膜电位的主要候选人监管通过保持静止膜电位接近钾平衡电势。钾离子通道的激活或抑制导致hyperpolarisation或细胞膜的两极化,分别导致血管舒张的起始和血管收缩。至少有四个类型的钾离子通道已确定在PASMCs [1]。背景或泄漏potassium-selective通道异常在钾离子通道,作为他们的活动不是由电压控制,他们可能会被视为静止膜电位的主要因素和输入电阻主要人类肺动脉平滑肌细胞(hPASMCs) [2,3]。
Src家庭酪氨酸激酶(SrcTK) nonreceptor酪氨酸激酶家族成员参与多种细胞信号和功能(4]。c-SrcTK是针对等离子体膜由于myristylated n端地区。SH1 SrcTK领域包含Tyr419磷酸化的网站,这是需要全c-SrcTK激活(5]。这种机制允许c-SrcTK与离子通道调节它们的属性(6,7]。失活的人类c-SrcTK发生在它的c端Tyr530磷酸化;然后结合SH2域。晶体的研究表明,糖和SH2域之间的相互作用,和激酶域和SH3域之间,导致c-SrcTK分子认为关闭配置覆盖衬底的激酶结构域,减少其潜在交互作用(5]。糖基的背景TWIK-related酸敏感钾(任务)1通道包含可能网站酪氨酸、丝氨酸磷酸化激酶(图S1a)。如果通常由SrcTK激活钾离子通道,逆转这种机制可以解释观察到的肺血管张力增加SrcTK抑制的达沙替尼治疗后(8]。
在目前的研究中,我们调查的功能角色SrcTK钾通道功能和膜电位。我们研究了缺氧抑制任务1频道SrcTK hPASMCs定义特定的角色。我们证明特定的抑制SrcTK结果在任务1的抑制和其他膜钾离子通道,导致两极化。此外,缺氧减少活跃phospho-SrcTK co-localisation task 1通道的膜,同时激活SrcTK增加任务1电流这激活电流由中度缺氧抑制。这表明,钾通道功能极度依赖SrcTK,从而解释了为什么SrcTK抑制可能导致危及生命的肺部血管收缩。这是第一个报告演示的功能角色SrcTK在静止膜电位的规定,在缺氧抑制任务1频道在初级hPASMCs,最后,在肺血管的基调。
方法
研究协议组织捐赠的机构审查委员会批准的格拉茨医科大学的格拉茨,奥地利,依照国家法律和良好的临床实践指南/国际协调会议(www.ich.org/)。书面知情同意从每个病人是否合适。所有动物实验机构批准的动物保健和使用委员会(BMWF,奥地利格拉茨医科大学的,格拉茨,奥地利)。
的制备主要hPASMCs和细胞培养
主要从人类抵抗肺动脉平滑肌细胞来自患者(n = 30)肺癌肺手术没有肺血管疾病史或动脉低氧血,或未使用的供体肺肺移植的收获。在获得肺癌患者的肺动脉,只有动脉是一个距离≥5厘米的肿瘤组织。详情,请参阅在线补充材料。
电生理学
采用全细胞膜片箝技术在hPASMCs如前所述测量静止膜电位在当前夹和宏观钾电流电压钳(下3]。协议的详细描述和解决方案给出了在线补充材料。
数据存储和分析商用pCLAMP 9.0软件(轴突仪器,福斯特城、钙、美国)。
钙的测量
荧光染料fluo-4-AM用于检测hPAMSCs检测细胞内钙的变化。协议的详细描述和解决方案给出了在线补充材料。
随后获得的图像存储和离线处理与TillVision软件(直到光子学,德国慕尼黑)。
小干扰RNA转染对c - src,菲英岛和任务1
小干扰rna (siRNAs)与c - src (siC-Src),菲英岛(siFyn)和任务1 (siTASK-1)商业合成(Eurogentec、血清、比利时)。详情,请参阅在线补充材料。作为一个消极的控制,nonsilencing RNA (nsRNA),不目标任何人类基因产品,使用。hPASMCs被种植在盖玻片或six-well盘子,和小干扰rna转染使用退火Effectene转染试剂(试剂盒、希尔登,德国)。基因混战被定量rt - pcr检查使用RNA提取(RNeasy;试剂盒)转染细胞。
RNA水平,live-cell钙和电生理测量进行48-56 h post-transfection。评估小干扰rna转染的效率,异硫氰酸荧光素(FITC)结合使用了核。只用于FITC-positive细胞电生理学的研究。此外,siTASK-1转染被溢出功能控制的细胞有浴室的解决方案调整pH值为8.3。
rt - pcr
详细描述是网上提供的补充材料。
定量rt - pcr
定量rt - pcr进行检查c - src的表达,菲英岛和hPASMCs任务1。更多细节,请参阅在线补充材料。
Co-immunoprecipitation,免疫印迹和immuno-co-localisation
详情,请参阅在线补充材料。
缺氧的治疗hPASMCs
缺氧的作用在膜片箝和calcium-imaging研究研究了在常氧和缺氧灌流液水库之间切换。详细描述是网上提供的补充材料。
孤立的,肺灌注和通风的鼠标
从成人C57BL / 6小鼠肺(哈伦实验室,Inc .,印第安纳波利斯,美国)被移除的胸部下深anaesthaesia和人工通风,并灌注与克雷布斯Henseleit缓冲区(氯化钠120更易·L−14.3、氯化钾更易·L−1,KH2阿宝41.1更易·L−1,CaCl22.4更易·L−1,MgCl21.3更易·L−1和葡萄糖13.3更易·L−1,以及5%(重量/体积)hydroxyethylamylopectin(分子量200000 Da))。肺被安装在一个热室,允许负压通风气体含有5.3%股份有限公司2和21.0%啊2,用N和平衡2。最初的稳态阶段的15分钟(克雷布斯′Henseleit缓冲流量1毫升·敏−1)作为基线。详细描述,请参阅在线补充材料。4-amino-5——(4-chlorophenyl) 7 (t丁)pyrazolo [3,4 - d]嘧啶(PP2;30μM), 4-amino-7-phenylpyrazol [3,4 - d]嘧啶(很少出现;30μM)或达沙替尼(100μM)添加到缓冲一段Δ15分钟P巴勒斯坦权力机构显示肺动脉压力的变化(P巴勒斯坦权力机构应用PP2)后,项目或达沙替尼。
解决方案和化学物质
PP2和项目从σ化学公司购买(圣路易斯,密苏里州,美国)。Src激活肽来自圣克鲁斯生物技术有限公司(圣克鲁斯、钙、美国)。所有的药物都溶解在实验(浴)解决方案,除了PP2,项目,达沙替尼。他们是溶解在二甲亚砜(DMSO)。在这个浓度,车辆本身没有对离子电流的影响,静止膜电位P巴勒斯坦权力机构。pH值的解决方案包含药物测试和修正,消除潜在的pH-induced影响使用克雷布斯Henseleit缓冲区。NaHCO3作为一个缓冲区,调整导致一个常数pH值7.37 - -7.40。
统计分析
给出了数值意味着±sen的细胞或测量。组间差异被析因方差分析与评估事后分析与图基的测试,或未配对,配对t。假定值< 0.05被认为是重要的。
结果
的表达在人类肺癌和PASMCs SrcTK
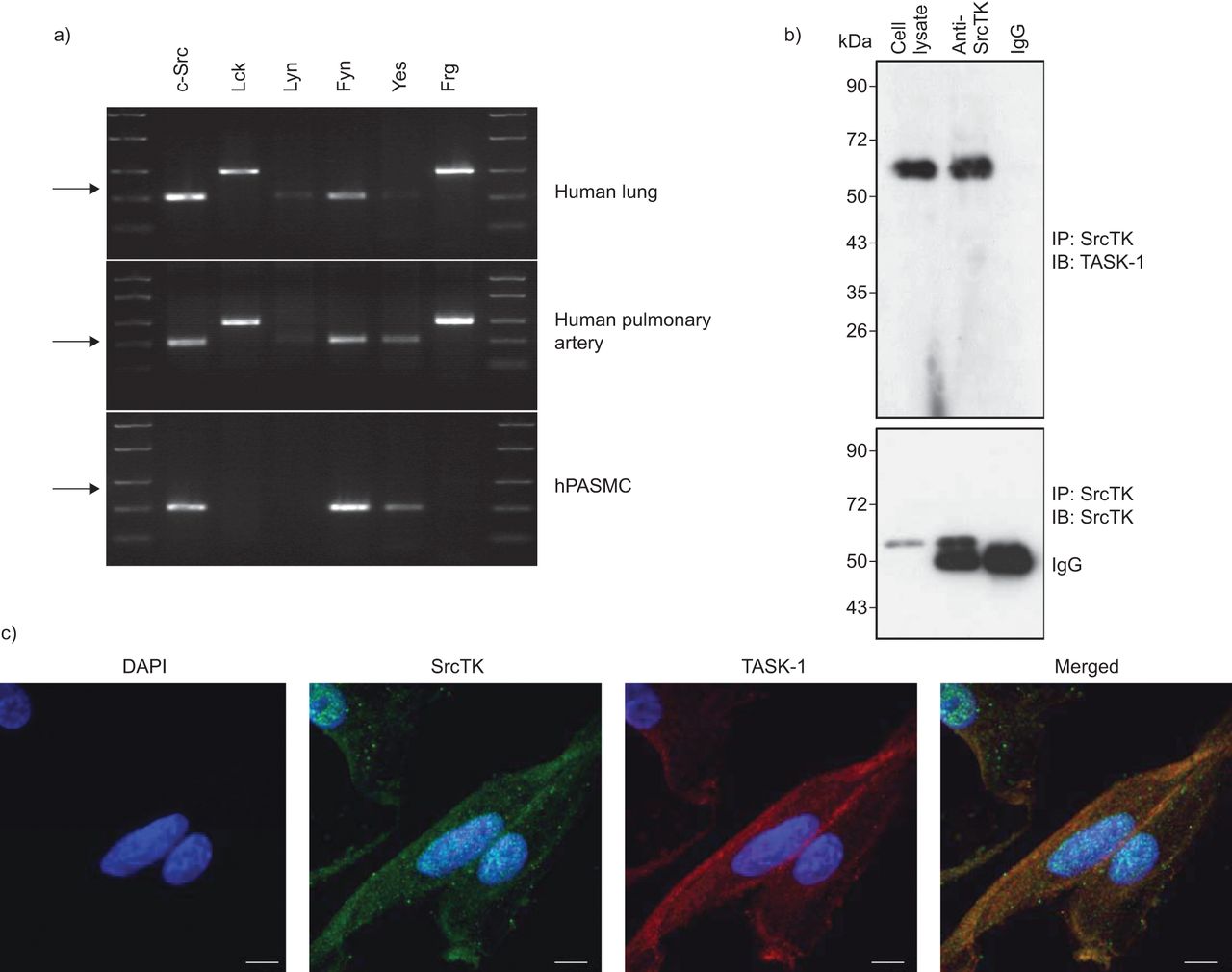
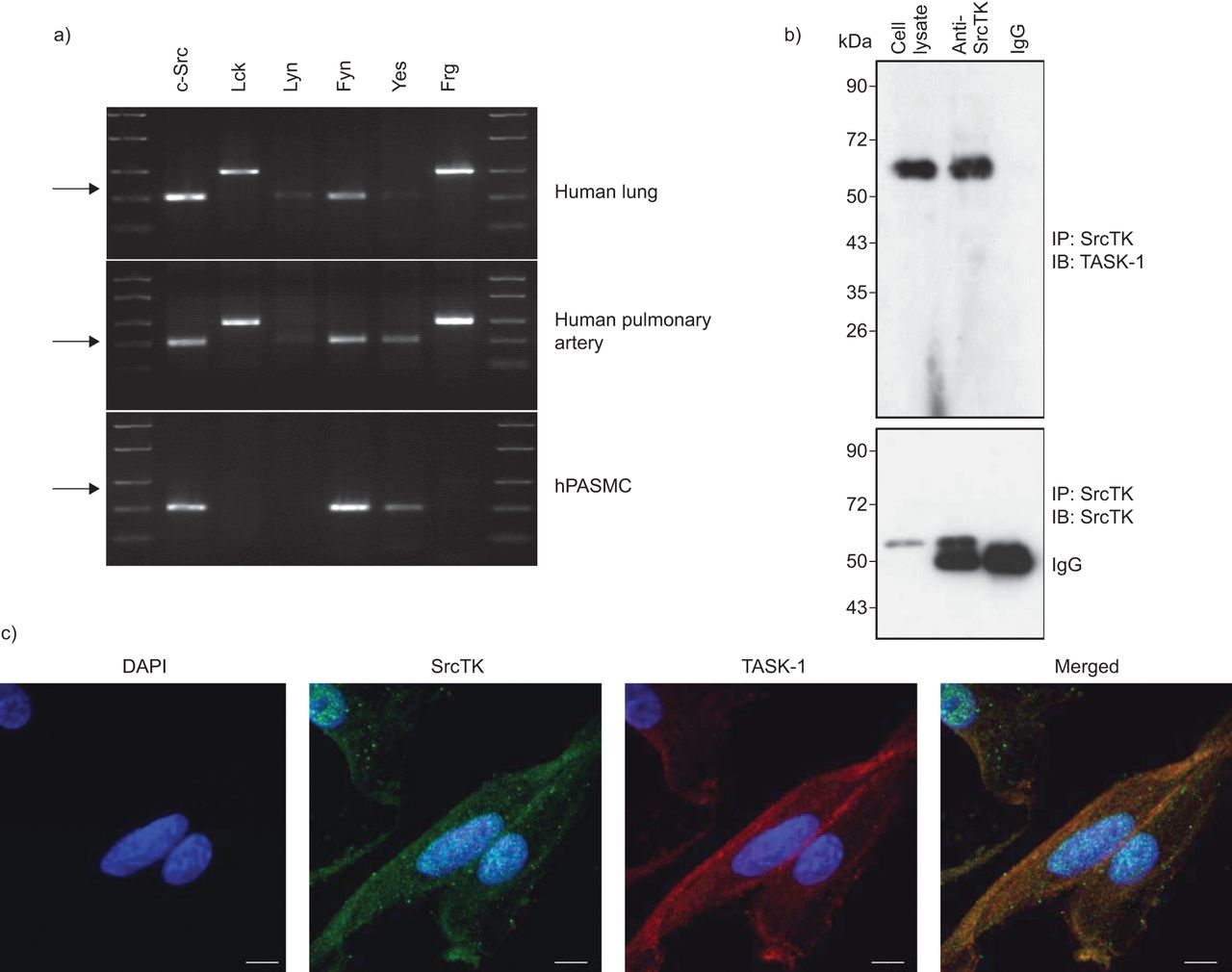
分析的表达SrcTK亚型、c - src Lck,林恩,菲英岛,是的,德意志联邦共和国,rt - pcr进行人类的肺组织,人类肺动脉和hPASMCs (图1一个)。我们的结果表明存在信使rna编码所有亚型在肺组织调查。主要只hPASMCs mRNA表达亚型c - src,菲英岛和肯定的。
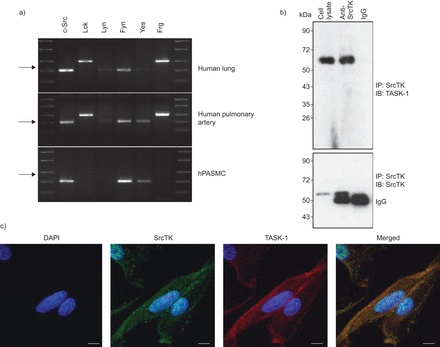
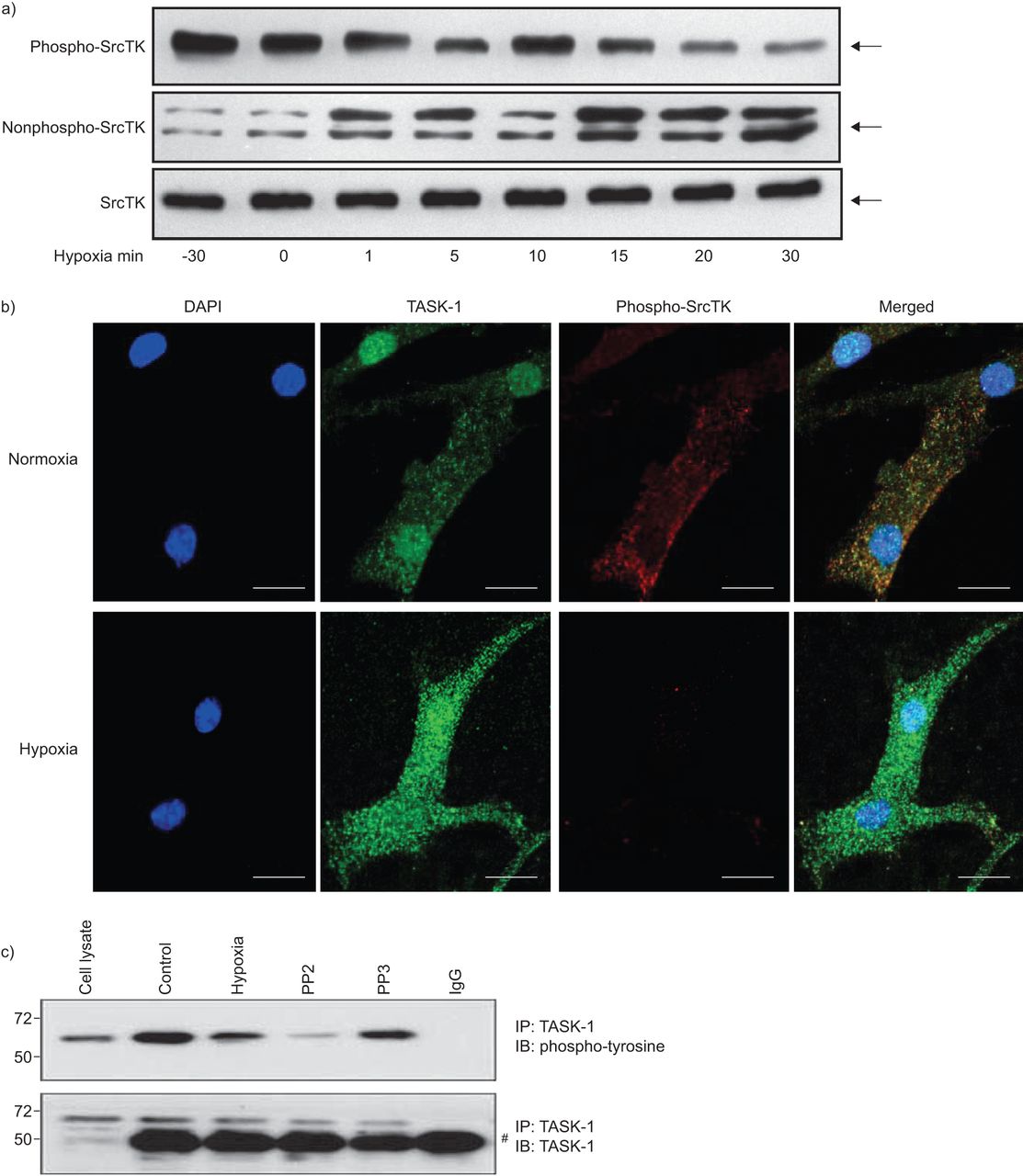
Src家庭酪氨酸激酶(SrcTK)是co-localised TWIK-related酸敏感钾(任务)1频道在人类肺动脉平滑肌细胞(hPASMCs)。RNA提取的)rt - pcr筛选SrcTK人类肺组织和初级hPASMCs单一化。代表凝胶说明mRNA的表达c - src(204个基点),Lck(398个基点),林恩(213个基点),菲英岛(206个基点),是的(202个基点)和德意志联邦共和国(402个基点)。箭头表示200个基点。相同的结果至少有三个准备工作从不同的供体肺和初级hPASMCs RNA。b)污点代表co-immunoprecipitation anti-SrcTK (IP),同形像控制免疫球蛋白(Ig) G和细胞溶解产物,然后免疫印迹(IB)任务1(上半部分)和SrcTK(下图)。c)荧光免疫染色显示DAPI核染色(蓝色),Src家族激酶(红色),任务1频道(绿色)和一个显示图像(合并)单排hPASMCs共焦图像。酒吧= 20μm规模。
SrcTK是co-localised hPASMC task 1频道
Co-immunoprecipitation SrcTK的任务1
免疫沉淀反应进行anti-SrcTK抗体或同形像控制(免疫球蛋白(Ig) G) hPASMC溶解产物。后来通过anti-TASK-1执行免疫印迹(图1 b、上面板)或anti-ScrTK抗体(图1 b,下半部分),显示的直接绑定SrcTK任务1。
背景的本地化task 1频道和SrcTK也想起在hPASMCs共焦激光扫描显微镜(图1 c)。Immunofluorescent染色task 1和SrcTK显示task 1和SrcTK co-localised细胞膜。染色没有负控制当细胞没有接触主要抗体被成像。任务1和SrcTK抗体进行免疫印迹(图印地)。
在hPASMCs SrcTK调节任务1频道活动
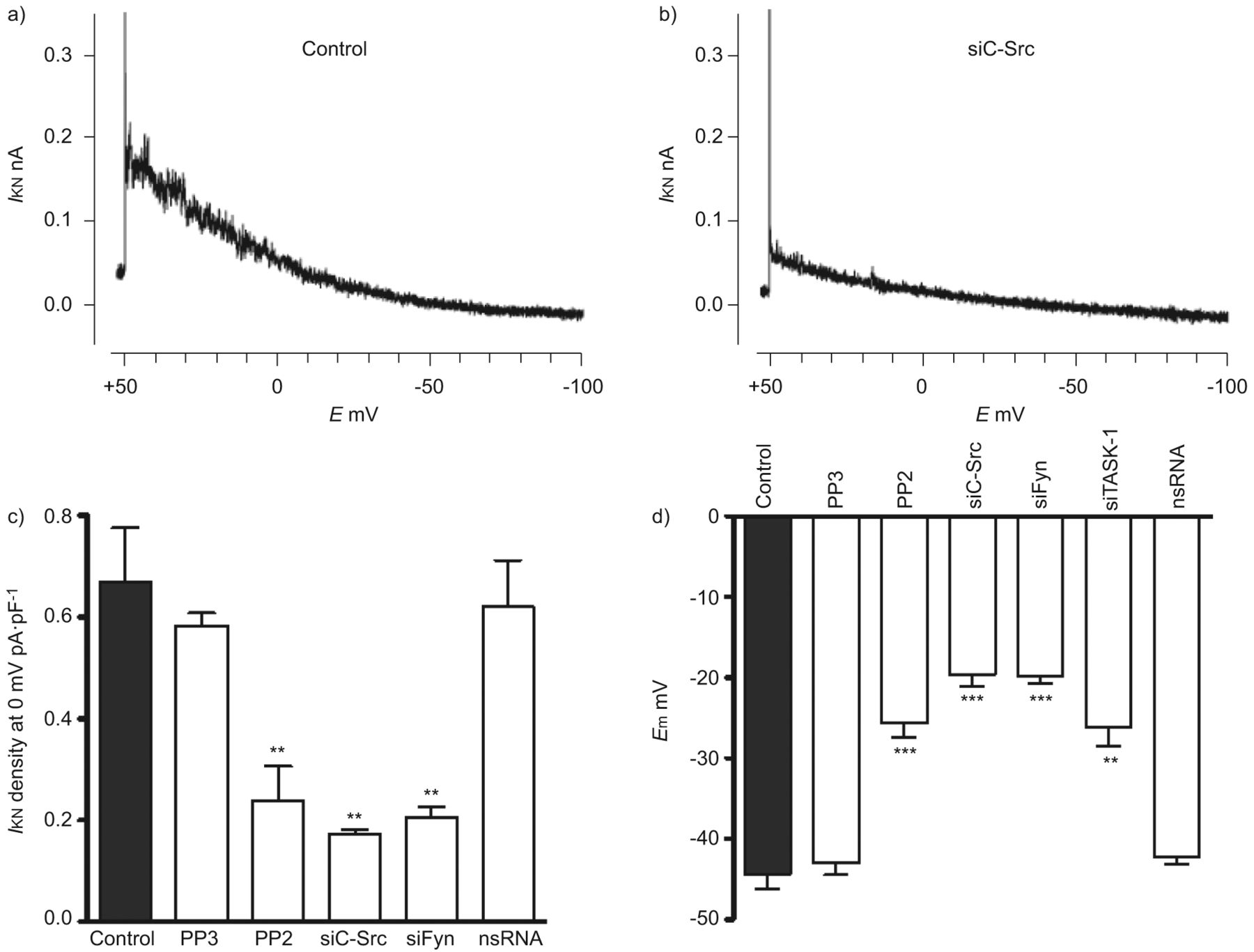
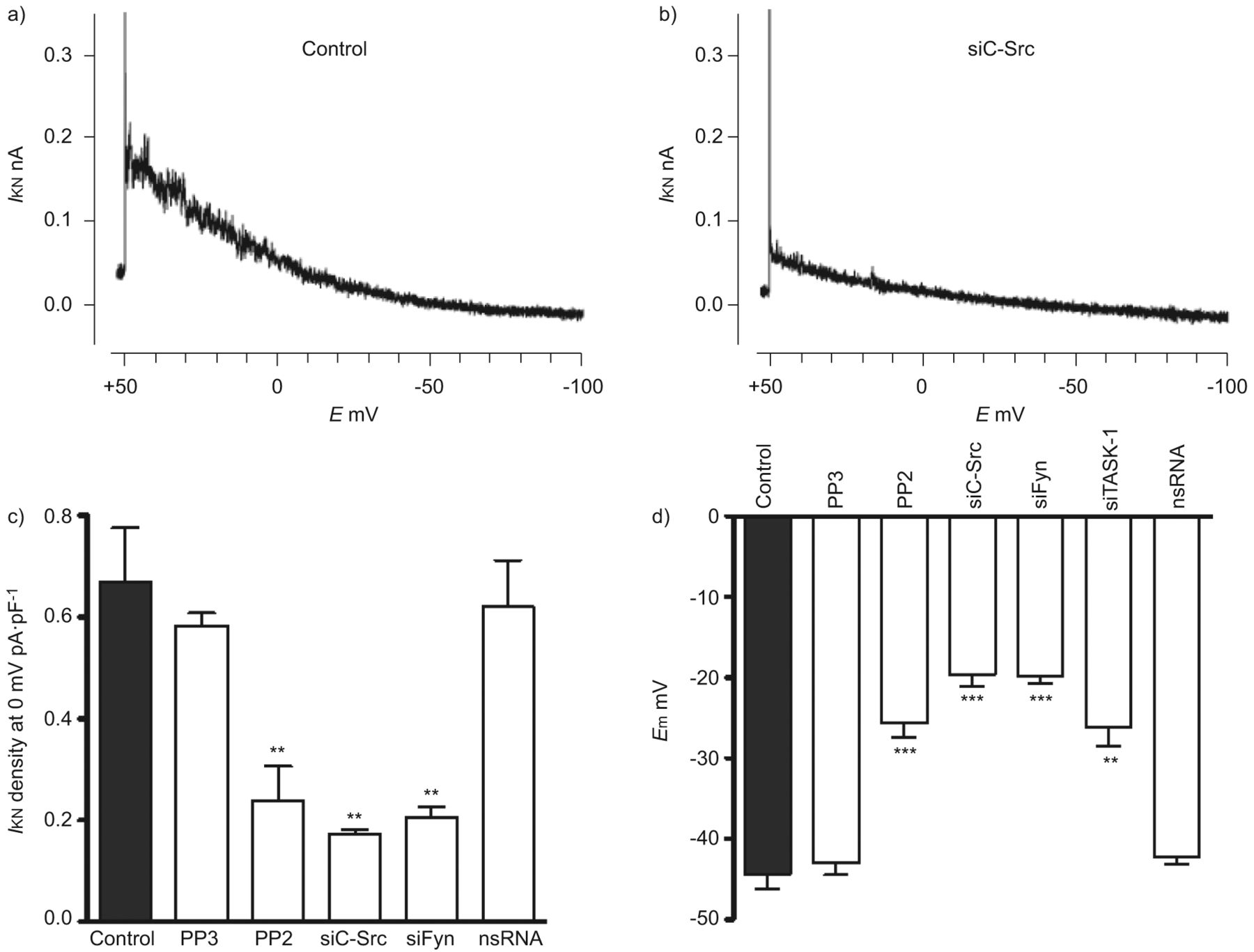
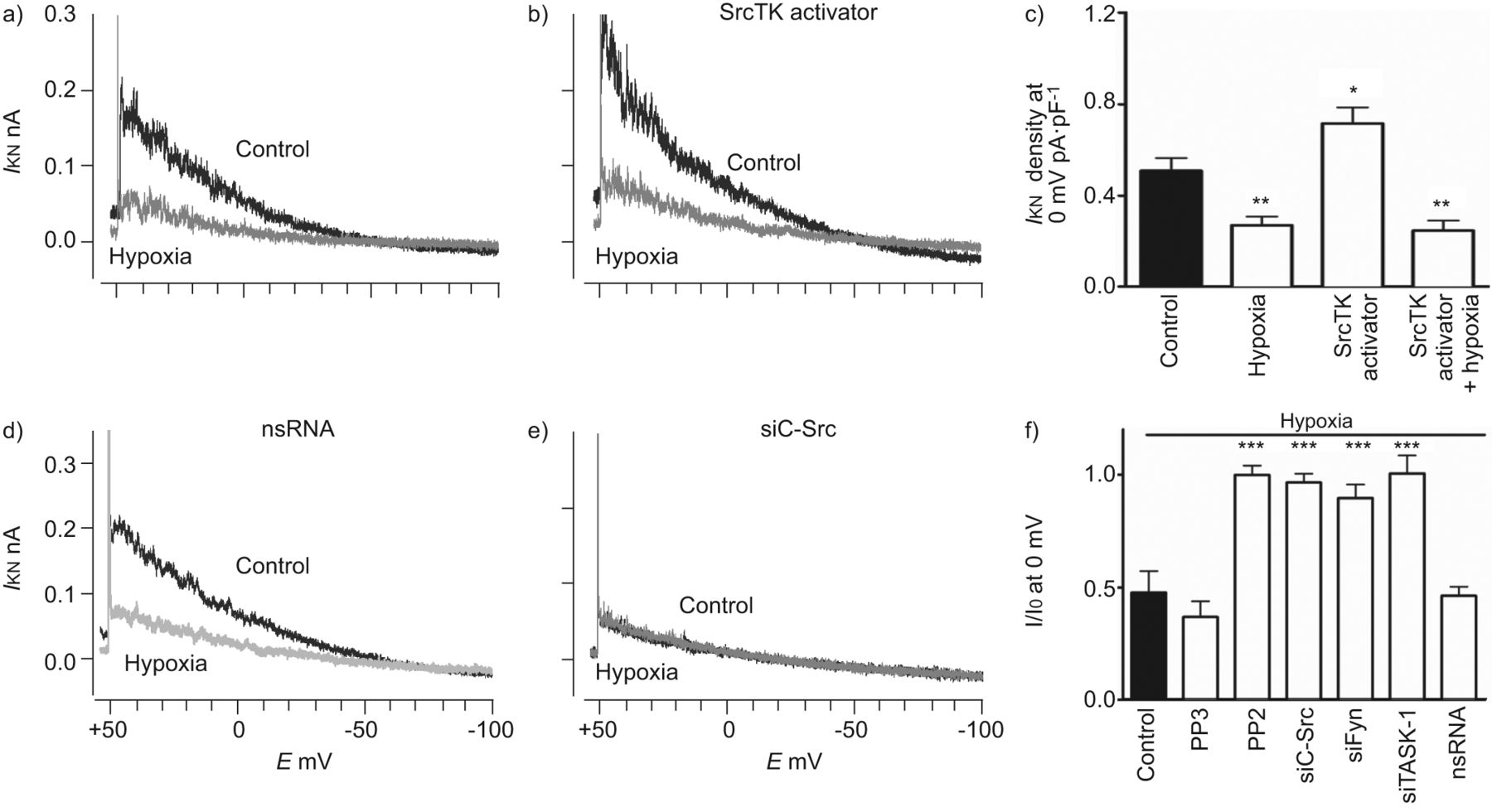
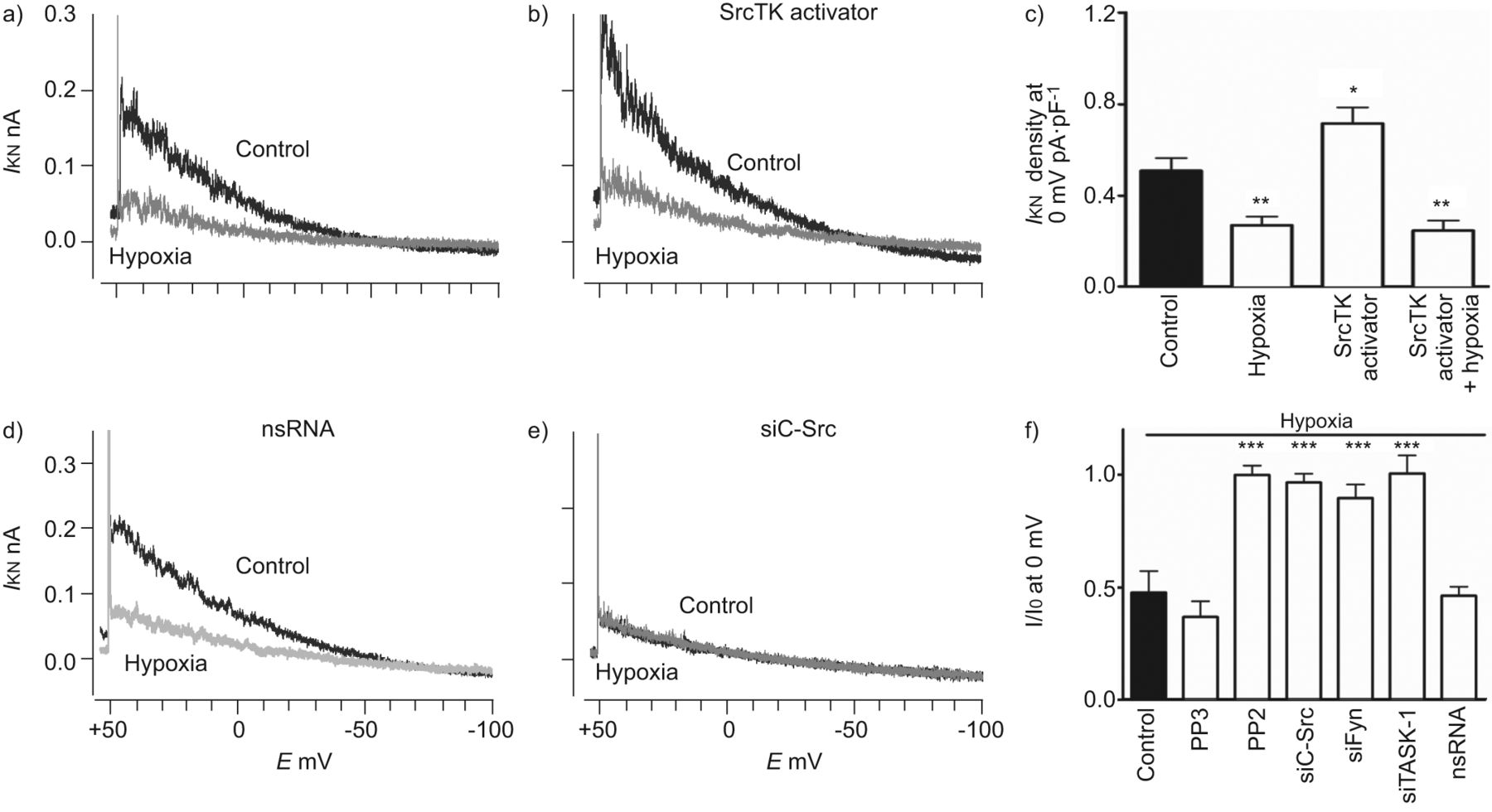
进一步确认的功能相关性SrcTK交互任务1频道,我们使用药理工具和核技巧。膜片箝记录的noninactivating task 1电流hPASMCs是如前所述[3]。SrcTK增加任务1频道活动证实了沉默的c - src和菲英岛主hPASMCs,导致降低电流与控制(图2 a和b)。任务1电流密度显著抑制通过治疗SrcTK抑制剂PP2 (0.23±0.06 pA·pF−1,n = 12)。尽管PP2 Src的有力和选择性抑制剂家族的蛋白酪氨酸激酶,为了研究Src子类型的作用,可以阻止对c - Src和使用菲英岛。转染与siC-Src (0.17±0.01 pA·pF−1,n = 4)或siFyn (0.20±0.02 pA·pF−1,n = 8)显著抑制任务1电流密度与控制(0.67±0.1 pA·pF−1,n = 13)。作为一个控制,很少出现的不活跃的模拟c - src抑制剂PP2和转染nsRNA应用。治疗项目(0.58±0.02 pA·pF−1,n = 4)或转染nsRNA (0.62±1 pA·pF−1,n = 8)不明显影响任务1电流(图2 c)。c - src的相对程度的降低和菲英岛各自siRNA证实了定量rt - pcr(图S2)。
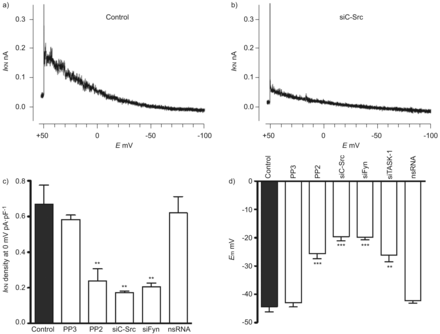
Src家庭酪氨酸激酶(SrcTK)是至关重要的TWIK-related酸敏感的钾(任务)1通道活动主要人类肺动脉平滑肌细胞(hPASMCs)。代表的录音任务1电流(我KN在控制细胞和b)))细胞转染携带RNA针对c - src (siC-Src)。c)直方图总结主要hPASMCs task 1电流密度。治疗后与Src的家人激酶抑制剂PP2(1μM)或转染siC-Src siFyn,电流密度(我KN)是显著降低。没有减少电流密度与nonsilencing转染后(ns) RNA或治疗后项目(1μM;不活跃的模拟PP2)。d)直方图总结SrcTK抑制的影响在静止膜电位(E米hPASMCs)。明显两极化的观察,当细胞接受PP2(1μM)或与siC-Src转染,siFyn或siTASK-1,但不与nsRNA转染后和治疗项目。E:膜电位。* *:p < 0.01;* * *:与控制相比p < 0.001。c和d)中的数据均值±sen的细胞或测量。
SrcTK抑制消除hPASMCs
任务1通道是活跃在静止膜电位和设置在PASMCs消极静止膜电位,正如我们先前已证明了主hPASMCs siRNA治疗。随着SrcTK抑制也降低了任务1电流(图2 c)的生理作用SrcTK静止膜电位的主要hPASMCs进一步调查。电生理测量进行hPASMCs显示治疗后明显两极化PP2 (-24.1±1.3 mV, n = 39)或转染siC-Src (-19.6±1.5 mV, n = 14)或siFyn (-20±1.1 mV, n = 14)相比与控制(-44.3±1.6 mV, n = 36)或与细胞治疗的不活跃的模拟PP2,很少出现(-43.4±1.5 mV, n = 4)或转染nsRNA (-43.6±1.5 mV, n = 18) (图2 d)。综上所述,这些数据进一步加强任务1的作用和SrcTK的重要性设置主PASMCs消极静止膜电位。
缺氧的监管SrcTK hPASMCs
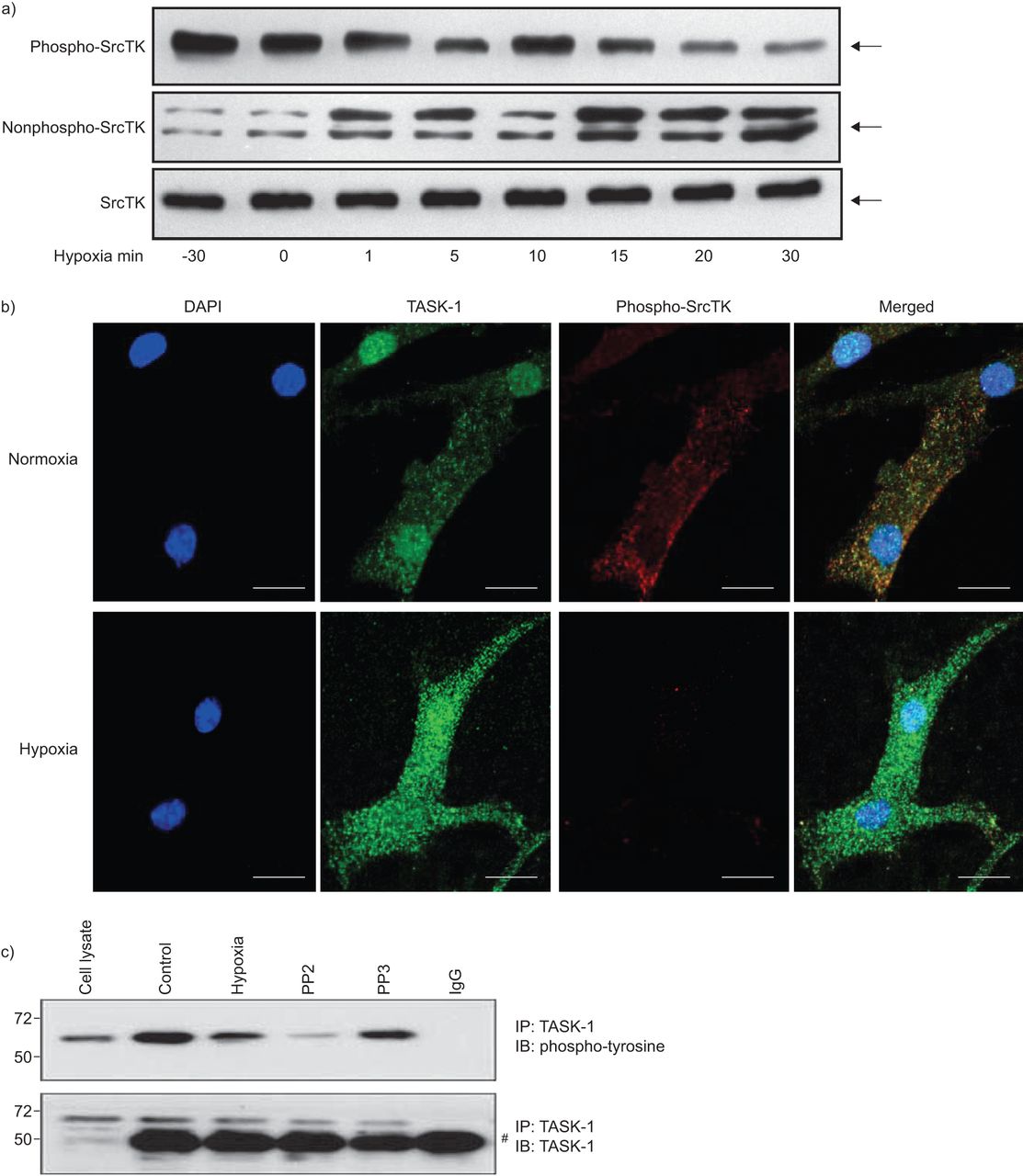
缺氧导致hPASMCs膜两极化,部分原因是任务1通道抑制。沉默SrcTK减少任务1的电流导致膜两极化。SrcTK活动是由在Tyr419 SrcTK的磷酸化状态。因此,我们调查了磷酸化(活跃;图3、上面板)和nonphosphorylated(不活跃;图3、中间面板)的状态在hPASMCs SrcTK不同跨度为缺氧(30分钟normoxia或0,1,5、10、15、20到30分钟的缺氧)。接下来,co-localisation后台任务1频道和phospho-SrcTK检查在hPASMCs normoxia 15分钟缺氧后,和呈现结果共焦激光扫描显微镜(图3 b)。较低的面板的图3 b清楚地显示了减少phospho-SrcTK染色在缺氧。图3 c表明,缺氧,以及应用SrcTK-inhibitor PP2,减少tyrosine-phosphorylation task 1通道的状态,而很少出现PP2的不活跃的模拟,不会改变任务1的磷酸化。图3 c显示了不变的总体水平在不同的实验条件下任务1。
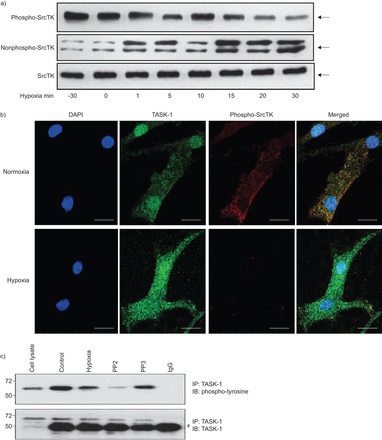
缺氧对Src家庭酪氨酸激酶(SrcTK)磷酸化在初级人类肺动脉平滑肌细胞(hPASMCs)。)缺氧phospho-SrcTK下降(Tyr419)免疫反应性(60 kDa),而增强免疫反应性检测在nonphospho-SrcTK (60 - 65 kDa)时间的方式。蛋白质载荷等效总SrcTK所示(n = 4)(总SrcTK由phospho-SrcTK和nonphospho-SrcTK)。b)的明显降低phospho-SrcTK Tyr419(红色)15分钟后显示缺氧而normoxia hPASMC单排共焦图像。酒吧= 20μm规模。c)应用缺氧或SrcTK抑制剂PP2 TWIK-related tyrosine-phosphorylation状态降低酸敏感的钾(任务)1通道。较低的面板显示了总水平在不同实验条件下的任务1不变。知识产权:免疫沉淀反应;IB:免疫印迹。#:免疫球蛋白G (Ig)。
接下来,功能作用的SrcTK缺氧抑制任务1通道检查。胞内透析与Src激活肽(EPQYEEIPIYL)显著增加任务1电流密度(0.72±0.07 pA·pF−1,n = 8)与控制(0.51±0.05 pA·pF−1,n = 10;图4和b)。缺氧能够抑制控制和激活电流。图4 c总结SrcTK活化剂肽和缺氧的影响。作为初始任务1电流下降抑制SrcTK拦截器或小干扰RNA (si) (图4 e),缺氧对细胞没有任何进一步的影响处理PP2或转染核目标c - src (siC-Src)或siFyn。然而,缺氧还能抑制任务1电流控制,PP3-treated或nonsilencing RNA转染hPASMCs (图4 d)。相对任务1电流在hPASMCs缺氧了图4 f。
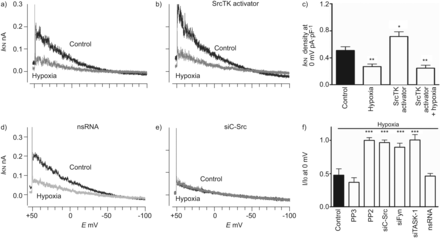
Src家庭酪氨酸激酶(SrcTK)激活增加TWIK-related acid-sensitive钾(任务)1通道电流,这种效果是完全抑制缺氧的主要人类肺动脉平滑肌细胞(hPASMCs)。代表的录音任务1电流(我KN)控制和在缺氧)没有或b)与SrcTK激活肽(EPQYEEIPIYL;1毫米)补丁吸管。c) SrcTK活化剂肽的影响,大大增加当前任务1的电流密度比控制。这个任务1电流显著地抑制增加了缺氧。任务1当前录音从细胞转染与d) nonsilencing (ns) RNA或e)携带RNA针对c - src (siC-Src)。在后一种情况下,缺氧不能进一步抑制当前任务1。f)的缺乏进一步治疗后低氧诱导任务1电流抑制PP2, siC-Src、siFyn siTASK-1。g)我KN任务1通道密度的ro - 31 - 8220(蛋白激酶C抑制剂(PK)), Go6983 (PKC抑制剂),KT5720 (PKA抑制剂)和复合C(活化激酶抑制剂)normoxia主hPASMCs(控制)和缺氧。我0:当前normoxia在每一个细胞;E:膜电位。*:p < 0.05;* *:p < 0.01;* * *:与控制相比p < 0.001。
任务1频道可以通过不同的途径,调制等活化蛋白激酶(AMPK) [9),蛋白激酶(PK) (3],PKC [10)和磷脂酶C (PL) (11),以应对不同的受体激动剂刺激。为了排除这些途径的参与,我们调查了缺氧抑制任务1当前使用不同的抑制剂。这些治疗影响控制任务1电流或其缺氧抑制(无花果S3和S4g)表明,AMPK, PKA, PKC和PLC无关。
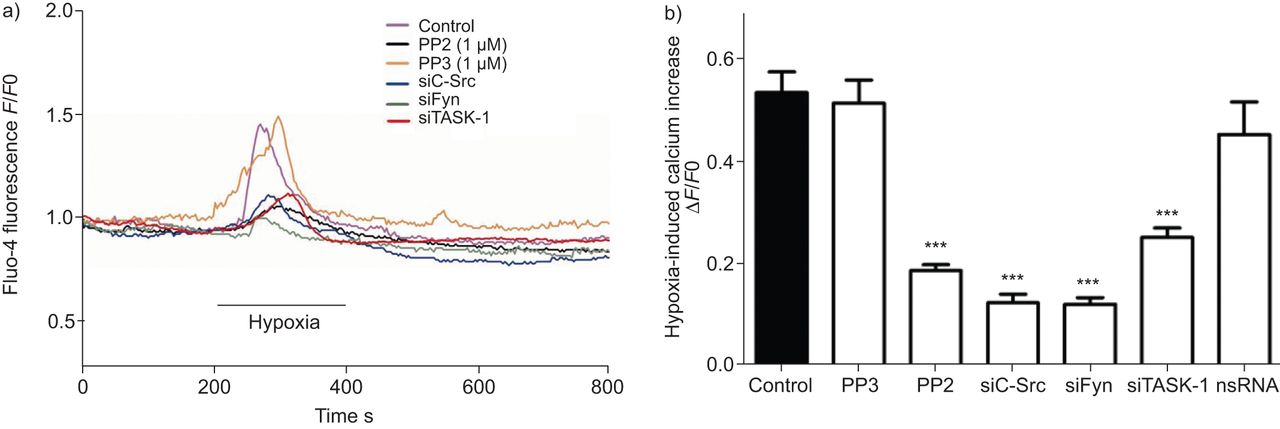
影响SrcTK hPASMCs低氧诱导的胞内钙增加
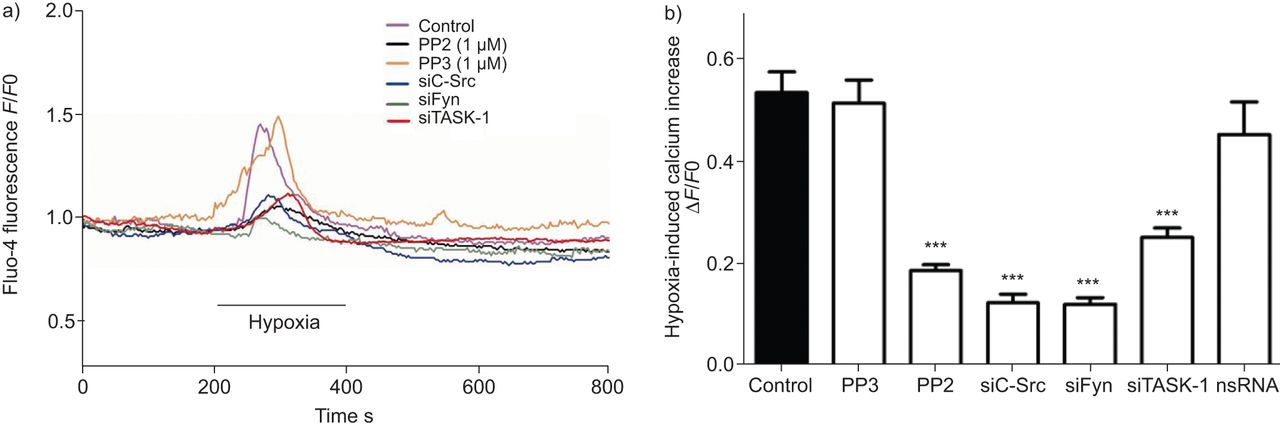
低氧诱导细胞内自由钙浓度上升(Ca2 +]我是一个积分和的hPASMCs特性直接关系到下游信号导致血管收缩。在低氧诱导SrcTK (Ca的角色2 +]我在hPASMCs上升进一步分析。Fluo4-loaded hPASMCs是连续监测(Ca的变化2 +]我在缺氧(图5)。缺氧(Ca显著增加2 +]我在控制细胞(0.54±0.04,n = 63)、和在细胞治疗项目(0.52±0.04,n = 17)或转染nsRNA (0.45±0.06, n = 16)。相比之下,低氧诱导增加0.54±0.04要么PP2治疗后明显减弱(0.18±0.01,n = 68)或转染siC-Src (0.12±0.02, n = 18), siFyn (0.12±0.02, n = 14)或siTASK-1(分别为0.25±0.02,n = 22)。总结研究的结果发表在图5 b。
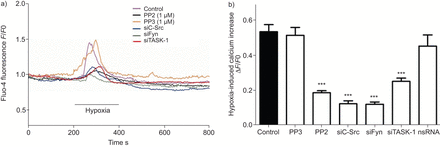
Src家族的抑制酪氨酸激酶变弱诱导细胞内钙离子浓度增加主要是人类肺动脉平滑肌细胞(hPASMCs)。)代表的录音fluo-4荧光(F)在缺氧挑战控制hPASMCs和细胞治疗PP2,很少出现小干扰RNA针对c - src (siC-Src)或siFyn。b)低氧诱导细胞内钙增加与PP2治疗后下降,siC-Src或siFyn,但不是nonsilencing (ns) RNA或很少出现。F0:荧光的挑战。任务1:TWIK-related potassium-1酸敏感。* * *:与控制相比p < 0.001。
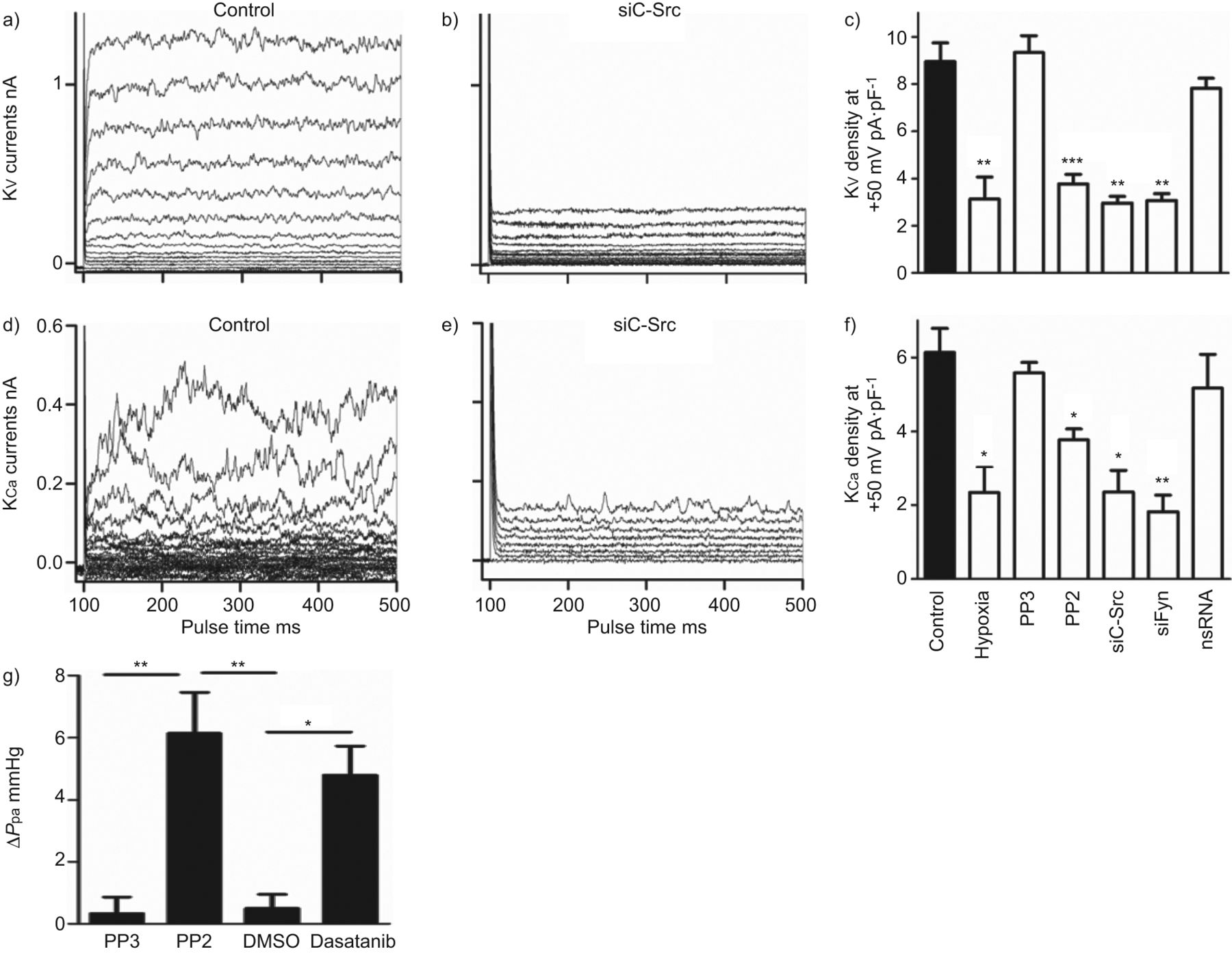
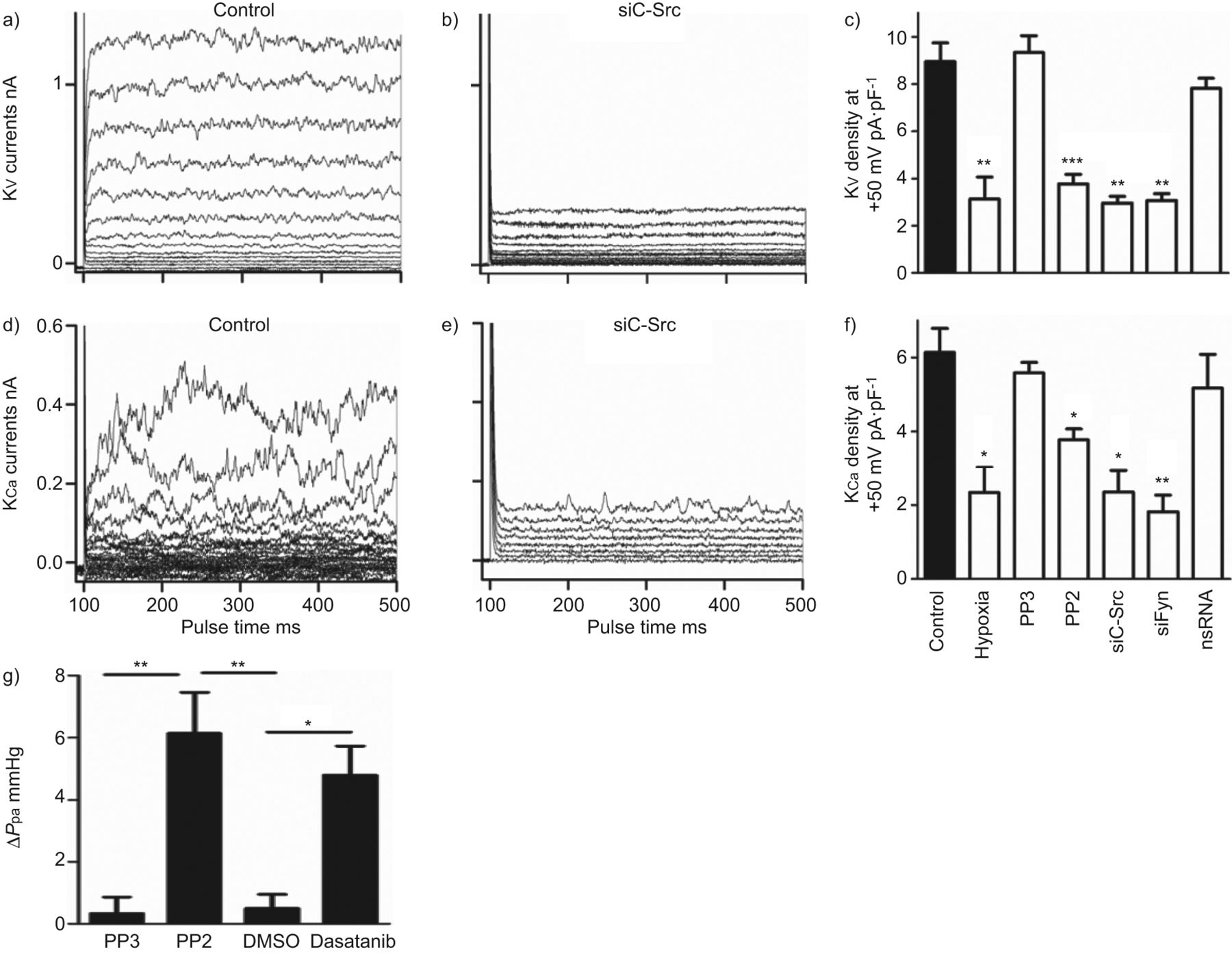
抑制SrcTK变弱电压门控和hPASMCs calcium-dependent钾电流
SrcTK抑制的影响在全细胞钾电流(电压门控(Kv)和calcium-dependent (KCa钾电流)是人类PASMCs调查在初级。K代表v录音控制hPASMCs (图6)和治疗后siC-Src介绍图6 b,表明沉默的c - src显著降低Kv电流。类似的结果从实验记录KCa电流控制(图6 d)和c - src沉默后(图6 e主hPASMCs)。
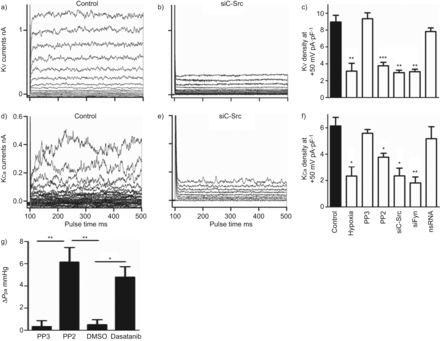
Src家庭酪氨酸激酶抑制减少全细胞电压门控(Kv)和calcium-dependent (KCa)钾电流主要人类肺动脉平滑肌细胞,增加肺动脉压(P巴勒斯坦权力机构)的孤立,灌注小鼠肺癌模型。a, d)代表K的痕迹v和KCa控制细胞,b, e)电流治疗后用小干扰RNA针对c - src (siC-Src)。c、f)直方图总结缺氧的影响,很少出现PP2, siC-Src siFyn或nonsilencing K (ns) RNA)v和KCa电流密度。g)直方图总结改变(Δ)P巴勒斯坦权力机构从模型包括鼠标肺在PP2面前,很少出现二甲亚砜(DMSO)和达沙替尼。*:p < 0.05;* *:p < 0.01;* * *:与控制相比p < 0.001。
进一步评估的角色SrcTK K的激活v和KCa渠道,PP2的影响,项目和siC-Src治疗,siFyn或nsRNA调查并与缺氧的影响(图6 c)。治疗的hPASMCs PP2 (3.8±0.4 pA·pF−1、n = 16)或siC-Src (2.9±0.3 pA·pF−1(n = 5)或siFyn (3.1±0.3 pA·pF−1,n = 5)显著减少了Kv当前与控制(8.9±0.8 pA·pF−1分别为,n = 24)。缺氧表现出相似的效果(图6 c)。治疗项目(9.3±0.7 pA·pF−1,n = 4)或转染nsRNA (7.8±0.4 pA·pF−1,n = 9)没有改变当前(图6 c)。K时观察到类似的结果Ca当前记录(图6 e和f)。只有缺氧(2.3±0.5 pA·pF−1n = 16),治疗PP2 (4±0.3 pA·pF−1n = 17), siC-Src (2.35±0.6 pA·pF−1(n = 5)或siFyn (81±0.4 pA·pF−1,n = 5)减少当前与控制(5.7±0.6 pA·pF−1,n = 12),而项目(5.58±0.3 pA·pF−1(n = 5)或nsRNA (5.17±0.9 pA·pF−1,n = 8)没有显著的影响(图6 f)。
肺血管收缩,以应对SrcTK抑制剂
描述SrcTK在肺血管张力的作用,我们用模型包括老鼠肺部,PP2有显著提高P巴勒斯坦权力机构(6.3±1.3毫米汞柱,n = 4)较不活跃的模拟项目(0.6±0.2毫米汞柱,n = 4),有趣的是,慢性粒细胞白血病的二线治疗,达沙替尼,这也是一个强有力的抑制剂SrcTK,显示一个类似的增加(5.5±0.2毫米汞柱,n = 3)P巴勒斯坦权力机构与溶剂DMSO(0.4±0.4毫米汞柱,n = 4)。图6克显示了总结ΔP巴勒斯坦权力机构PP2前后,很少出现DMSO和达沙替尼。
讨论
本研究的主要发现是:1)两个SrcTK的家人,c - src和菲英岛,是高度表达主要hPASMCs;2)任务1频道和SrcTK co-localised hPASMCs的质膜;3)需要SrcTK task 1频道的活动;4)的抑制SrcTK动摇hPASMCs;5)缺氧降低任务1的酪氨酸磷酸化水平;6)缺氧减少SrcTK的活跃的磷酸化状态,抑制任务1电流通过SrcTK活化剂;7)SrcTK抑制低氧诱导细胞内钙升高明显变弱;8)Kv和KCa电流减少SrcTK抑制;和9)抑制SrcTK PP2或达沙替尼导致大量增加P巴勒斯坦权力机构灌注分离小鼠肺。
平滑肌细胞的膜电位控制肺血管张力的一个重要因素。静止膜电位的PASMCs大约是-50 mV2,3,12]。这是由钾从这些细胞通过钾离子通道流出。药物,抑制或激活肺血管平滑肌细胞钾离子通道导致两极化或hyperpolarisation,分别。函数、控制和钾离子通道的表达在肺动脉持续的兴趣,因为这可能减少钾离子通道表达,如。电压门控钾通道,shaker-related亚科,5 (Kv1.5)(13,14),或钾离子通道功能障碍引起膜两极化的(15),(Ca一起增加2 +]我,导致增加增殖和细胞凋亡的减少,最终导致肺动脉高压的发病机制(16]。
hPASMCs两极化,诱导的抑制钾离子通道,紧随其后的是大量钙通过电压门控钙通道(l型)和导致肺血管阻力增加。通过几种机制急性缺氧引起血管收缩。它消除PASMCs通过抑制钾离子通道,导致钙流入如上所述[17),导致从肌浆网钙的释放和随后的饱满通过门店渠道(18,19),增加钙流入PASMCs通过l型钙通道,独立的膜电位20.和也促进钙敏化作用21),从而增加肺血管阻力(22]。大部分的钙负责缺氧引起的胞质钙的增加来自PASMC的外面,但有些是释放内部商店,如肌浆网(1]。很可能减少低氧诱导钙的增加是由于PP2, siC-Src或siFyn (图5)是次要的缺乏缺氧抑制钾电流,因为抑制钾离子通道已经由SrcTK抑制引起的。缺氧的反应是废除了在减少SrcTK活动(无花果4和6)。
几项研究已经表明,酪氨酸激酶可能作用于钾离子通道。然而,这些研究进行了在细胞系或异源表达系统使用广泛抑制剂但没有显示结果的生理作用(7,23,24]。之前我们已经表明,背景两个孔隙域task 1频道集人类PASMCs主要的膜电位,它可以被PKA调制,PLC, PKC和AMPK途径通过serine-threonine磷酸化不同的受体激动剂,如内皮素(10],treprostinil [3),等。此外,缺氧消除膜电位的可逆抑制任务1频道在hPASMCs [3]。此外,KCa和Kv渠道也可能导致膜电位,特别是如果他们是刺激代理环腺苷酸、环磷鸟苷。因此,我们的研究结果表明,SrcTK活动对于低生理PASMCs的语气是至关重要的,因此,对于低肺血管阻力。
缺氧的机制抑制这些渠道目前未知。然而,在目前的研究中,特定的抑制内源性SrcTK减少hPASMC task 1电流。同样地,当hPASMC透析SrcTK活化剂,观察任务1电流显著增加,表明当前需要SrcTK活动任务1,降低当前和磷酸化增加脱磷酸化。其他几个钾离子通道,包括KCa和Kv,曾被SrcTK-mediated酪氨酸磷酸化调制(25]。我们的观察证实了这一点但演示ScrTK活动的关键作用这些通道和显示任务1的功能活动需要SrcTK活动。
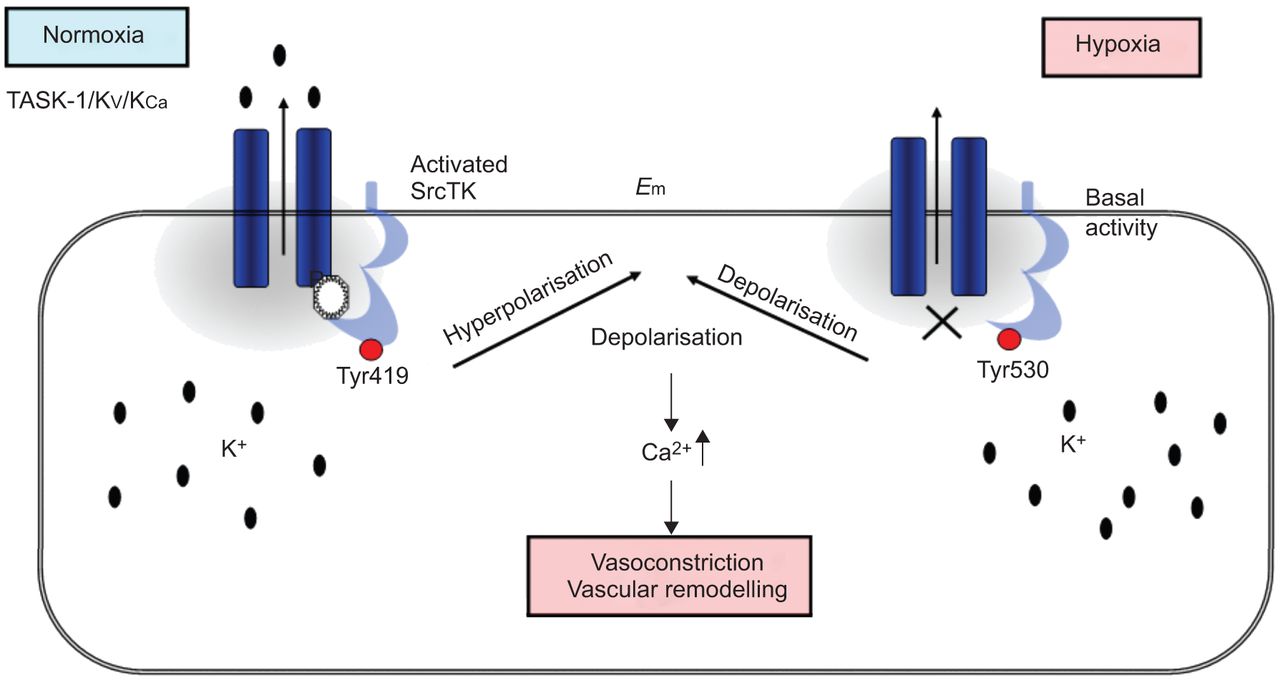
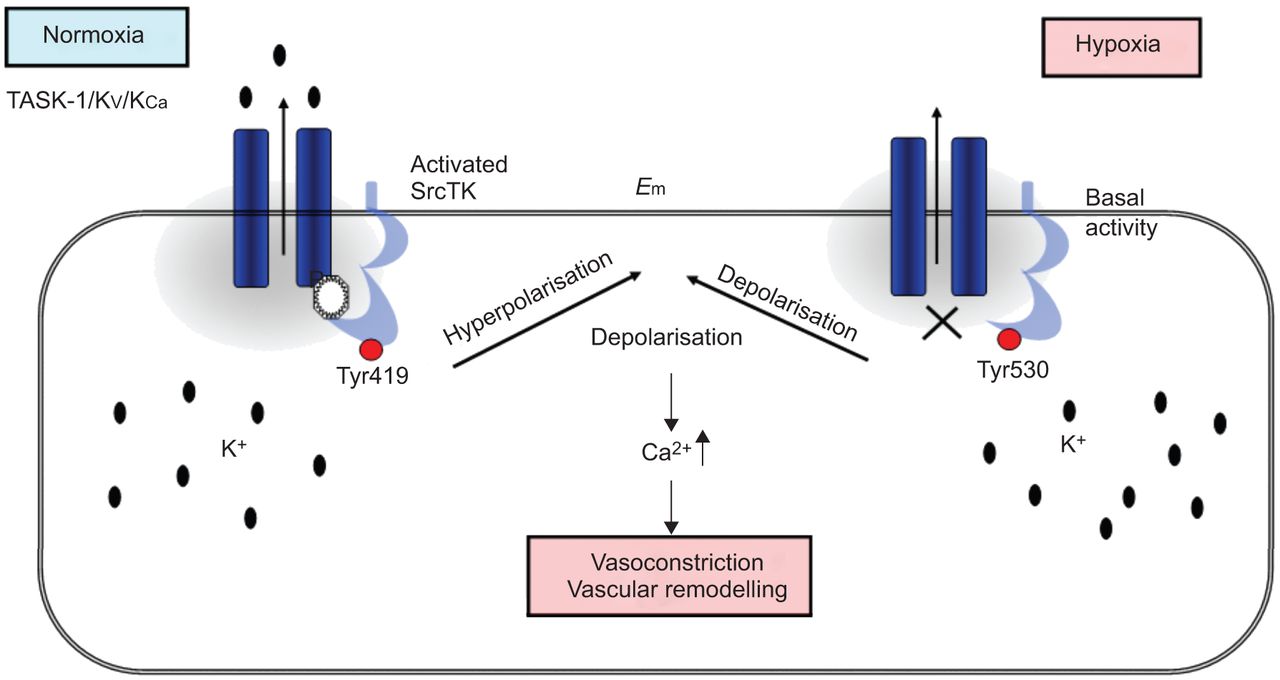
在目前的研究中,我们表明,SrcTK抑制减少当前任务1和任务1的缺氧抑制中扮演着关键角色渠道,可能通过减少磷酸化SrcTK Tyr419和离解task 1和SrcTK,虽然这个链接的分子调控尚未确定(图7)。
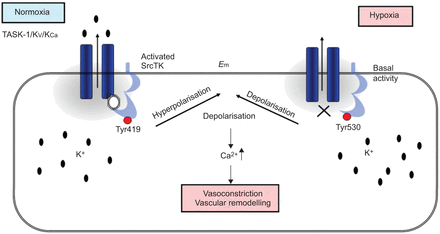
方案提出的相互作用TWIK-related酸敏感钾通道(任务)1和c - src在人类肺动脉平滑肌细胞(hPASMCs)。normoxia下,phospho-Src(活跃的Src,磷酸化Tyr419)结合任务1通道导致功能性任务1频道。在hPASMCs活跃任务1通道保持消极休息的潜力。在缺氧,phospho-Src (Src)却降低了。关闭任务1导致两极化和增加细胞内钙水平。E米:静止膜电位。SrcTk: Src家庭酪氨酸激酶;Kv:电压门控钾电流;KCa:calcium-dependent钾电流。
最近的研究由K诺和同事(26,27]描述SrcTK抑制由PP2缓解低氧诱导的肺血管收缩和抑制ρPASMCs激酶在大鼠肺动脉和孤立。他们发现PP2减少了低氧诱导增加PASMC (Ca2 +]我与我们的观察是一致的。鉴于PP2的抑制效果和siRNAs task 1, Kv和KCa电流,我们在这里描述,类似缺氧对这些水流的影响,是有意义的序列中的SrcTK磷酸化的抑制导致钾电流抑制。然而,在他们的实验中,K诺和同事(26,27)指出,在Tyr419缺氧SrcTK磷酸化的增加而不是减少,正如我们也报告在这里。这些结果的差异可能是由于不同的物种,细胞培养条件下,周期的数量和严重程度、持续时间或时间点研究缺氧。
观察到的增加P巴勒斯坦权力机构SrcTK PP2和抑制后的达沙替尼表明功能角色SrcTK调节肺血管的基调。这意味着严重肺动脉高压患者尚不清楚。T条件下等。(28c - src)描述水平的下降在16个原发性肺动脉高压(特发性肺动脉高压(PAH)根据当前分类)的病人。这将对应于钾通道活性下降,可能导致血管收缩和其他病理机制导致肺动脉高压。在此,我们报告一个减少任务1,全细胞钾电流和KCa当前在hPASMCs c - src抑制后。这是符合观测表明,降低Kv1.5活动导致肺动脉高压(13]。相比之下,最近的一项研究Courboulin等。(29日]报道水平的提高phosphor-Src和总Src肺样本三PAH患者。
最近的临床观察强调潜在的病理生理学的酪氨酸激酶PAH的重要性。慢性粒细胞白血病(CML)是由一个既定的活跃的bcr - abl酪氨酸激酶。伊马替尼,抑制这个激酶,是一线治疗CML [30.]。伊马替尼也是一个有效的血小板源生长因子受体抑制剂(31日),这被认为是为什么它可能改善一些PAH患者的血液动力学(32]。达沙替尼是酪氨酸和serine-threonine 325倍的伊马替尼的激酶抑制剂的抑制bcr - abl激酶在体外,导致较高的CML cytogenic反应(33]。然而,达沙替尼强有力地抑制了Src家庭,这可能是最著名的dasatinib-targeted家族的蛋白质激酶,包括c - Src和菲英岛,据报道导致多环芳烃(8,34,35]。多环芳烃倾向于停药后迅速解决的达沙替尼(34),这表明它可能不是主要是由于标记细胞增殖,但慢性血管收缩。人们可以推测dasatinib-initiated肺动脉高压可能与我们发现核与c - src和菲英岛减少hPASMCs钾通道电流,导致两极化。很明显,许多工作有待完成澄清不同的激酶的作用多环芳烃的病因学和治疗。
研究的局限性
尽管目前的研究结果表明,任务1频道和SrcTK co-localised hPASMCs的质膜,由co-immunoprecipitation研究进一步支持,我们不能排除,Src也可以间接地对钾离子通道通过其他Src下游分子。此外,机制(s)参与急性低氧诱导的抑制钾离子通道可能不同于那些参与慢性缺氧和/或达沙替尼。最后,改变P巴勒斯坦权力机构在PP2和/或达沙替尼可能不会建议与此相关的精确机制。
结论
总之,我们证明SrcTK有重要作用的控制任务1和其他钾离子通道,它集hPASMCs消极静止膜电位。SrcTK的生理意义和任务1频道协会强调,当前任务1的低氧诱导抑制和细胞内钙依赖SrcTK上升。很可能是一种更好的方式来描述的多功能作用SrcTK关于钾通道监管将促进我们对肺动脉高压的病理生理学的理解。
确认
m . Schloffer优秀的技术援助和p . Blumel(实验麻醉,麻醉和重症监护医学部门,格拉茨医科大学的格拉茨,奥地利)我将非常感谢。
脚注
可以从本文的补充材料www.www.qdcxjkg.com
社论评论看第3页。
支持声明
这项研究的部分资金由格拉茨医科大学的(c . Nagaraj在分子医学博士项目)。E.K.堰VA医学研究资助和支持RO1 HL 65322(美国国家卫生研究院,马里兰州贝塞斯达)。
感兴趣的语句
语句对h . Olschewski和a . Olschewski可以找到www.www.qdcxjkg.com/site/misc/statements.xhtml
- 收到了2011年12月6日。
- 接受2012年4月3日。
- ©2013人队











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}