

文摘gydF4y2Ba
蛋白质酪氨酸磷酸化和内生的表达明显减少酪氨酸激酶的表达功能的电压门控钾(Kv)通道。蛋白质酪氨酸激酶蛋白和细胞基质的水平不受影响,符合减少酪氨酸磷酸化蛋白酪氨酸激酶抑制的结果的活动。蛋白质酪氨酸磷酸化的衰减与表达野生型和突变体的控制性能Kv频道。此外,细胞蛋白质酪氨酸磷酸化在几分钟内减少了与电致急性治疗钾离子载体缬氨霉素。因为酪氨酸磷酸化反过来影响Kv通道活动,这些结果表明,Kv通道发生了相互调节的相互作用和蛋白质酪氨酸磷酸化信号通路。gydF4y2Ba
- delayed-rectifier钾通道gydF4y2Ba
- 蛋白质酪氨酸激酶gydF4y2Ba
- srcgydF4y2Ba
- 表皮生长因子受体gydF4y2Ba
- 调制gydF4y2Ba
- 磷酸化gydF4y2Ba
蛋白质酪氨酸磷酸化调节多种细胞过程,包括增殖、分化、和生存。稳态细胞蛋白质phosphotyrosine水平受蛋白质酪氨酸激酶的反对活动(ptk)和蛋白质酪氨酸磷酸酶(PTPases)。大多数细胞表现出低蛋白质phosphotyrosine稳态水平,这反映了严格监管PTK活性和相对较高的总比活度比PTPases PTK (gydF4y2Ba亨特,1995gydF4y2Ba)。PTK活动被许多刺激细胞膜,调制和大量的工作被指向描述信号事件发生后绑定的生长因子,细胞因子、神经递质,和抗原到特定的膜受体(gydF4y2Ba1990年6月et al .,gydF4y2Ba;gydF4y2BaSchlessinger和乌尔里希,1992gydF4y2Ba;gydF4y2BaErpel Courtneidge, 1995gydF4y2Ba;gydF4y2Ba亨特,1995gydF4y2Ba;gydF4y2BaWan et al ., 1996gydF4y2Ba)。最近的研究表明,电气信号可以影响PTK活性和蛋白质酪氨酸磷酸化(gydF4y2Ba坏,格林伯格1991gydF4y2Ba;gydF4y2BaRusanescu et al ., 1995gydF4y2Ba;gydF4y2BaOlivotto et al ., 1996gydF4y2Ba;gydF4y2Ba西西里岛舞蹈et al ., 1996gydF4y2Ba)。gydF4y2Ba
电压门控钾(Kv)通道的活动是至关重要的电兴奋性(gydF4y2Ba1992年赴gydF4y2Ba)。Kv通道密度和向外钾电导的增加导致细胞膜电位超极化(gydF4y2Ba菲利普et al ., 1993gydF4y2Ba;gydF4y2Ba比彻et al ., 1996gydF4y2Ba;gydF4y2BaMarom et al ., 1996gydF4y2Ba),而钾电导下降造成去极化(gydF4y2Ba伦纳德et al ., 1992gydF4y2Ba;gydF4y2Ba比彻et al ., 1996gydF4y2Ba;gydF4y2BaPanyi et al ., 1996gydF4y2Ba)。调制的Kv频道活动改变细胞膜的特性,和蛋白质磷酸化是一种最合适的离子通道机制调制(gydF4y2Ba莱维坦,1994gydF4y2Ba;gydF4y2Ba乔纳斯和Kaczmarek, 1996gydF4y2Ba)。最近,有快速的进展描述酪氨酸磷酸化的影响在Kv频道包括哺乳动物gydF4y2Ba瓶gydF4y2Ba例如渠道Kv1.2, Kv1.3和Kv1.5 (gydF4y2Ba黄et al ., 1993gydF4y2Ba;gydF4y2Ba列弗et al ., 1995gydF4y2Ba;gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba,gydF4y2BabgydF4y2Ba;gydF4y2Ba萨博et al ., 1996gydF4y2Ba;gydF4y2BaFadool et al ., 1997gydF4y2Ba)。Kv渠道的活动往往是抑制酪氨酸磷酸化(gydF4y2Ba黄et al ., 1993gydF4y2Ba;gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba),通道动力学也观察到的变化(gydF4y2BaFadool et al ., 1997gydF4y2Ba);这些调节效应可以消除特定酪氨酸残基的突变。gydF4y2Ba
酪氨酸phosphorylation-induced调制的一个有趣的方面的Kv通道活动是Kv通道活动可能反过来影响PTK活性。先前的研究表明,PTK活性和蛋白质酪氨酸磷酸化是由离子电导和调制膜去极化(gydF4y2Ba坏,格林伯格1991gydF4y2Ba;gydF4y2BaVostal et al ., 1991gydF4y2Ba;gydF4y2Ba西西里岛舞蹈et al ., 1994gydF4y2Ba,gydF4y2Ba1996年gydF4y2Ba;gydF4y2Ba列弗et al ., 1995gydF4y2Ba;gydF4y2BaRusanescu et al ., 1995gydF4y2Ba;gydF4y2BaYu et al ., 1996gydF4y2Ba),离子电导会影响发育事件(gydF4y2Ba琼斯和Ribera, 1994年gydF4y2Ba;gydF4y2Ba斯皮策,1994gydF4y2Ba)。这就提出了一个有趣的可能性,相互调节Kv渠道和ptk之间可能发生的相互作用或PTPases。我们已经解决这个问题,不同的表达克隆Kv渠道在人类胚胎肾细胞(HEK 293),因为表达渠道确定膜电流响应和强烈主导转染细胞的电特性(gydF4y2Ba马歇尔et al ., 1995gydF4y2Ba;gydF4y2BaMarom et al ., 1996gydF4y2Ba)。gydF4y2Ba
材料和方法gydF4y2Ba
cDNA表达向量。gydF4y2Ba所有哺乳动物表达载体用于这些实验包含了巨细胞病毒(CMV)启动子上游的编码区。圣地亚哥(表达载体质粒pRc-CMV CA)被用来控制向量为所有实验。互补的v - src激酶(v - src)和人类表皮生长因子受体(EGFr)慷慨提供的理查德·哈甘耐尔教授博士(马里兰州巴尔的摩约翰斯·霍普金斯大学)。Kv1.4, Kv1.5 Kv1.3向量和路易博士提供的慷慨Philipson(芝加哥大学、芝加哥、IL),摩根博士盛(马萨诸塞州综合医院,波士顿,MA)和Richard Swanson博士(默克公司、夏普和Dohme研究实验室,西点军校,PA),分别。定点诱变用于构造五突变Kv1.3渠道酪氨酸或Trp残留物被突变检(三联体yyy111 - 113 fff、Y137F W386F, Y449F,和Y479F)。单一pcr使用诱变底漆和野生型引物被用来介绍突变。PCR孵化项目运行thermocycler(双块系统;圣地亚哥Eri-Comp CA)使用gydF4y2BaTaqgydF4y2Ba聚合酶(Promega,麦迪逊,WI)。PCR产品切割顺序在适当的限制性位点与苯酚和氯仿提取和乙醇沉淀之间的削减。一个相同的协议被用来减少野生型Kv1.3 pRc-CMV向量构造一个支柱。所需的部分凝胶纯化(Gene-Clean II;生物101年拉霍亚,CA)从2%琼脂糖凝胶,并插入诱变是绑定到Kv1.3-pRc-CMV支柱使用T4 DNA连接酶(Promega)。诱变插入经测序。gydF4y2Ba
细胞培养和转染程序。gydF4y2BaHEK 293细胞保持修改的鹰的介质(MEM)含有2%青霉素和链霉素和10%胎牛血清(GIBCO-BRL,马里兰州)。细胞种植confluency(1周),分离trypsin-EDTA和机械磨碎,稀释浓度在MEM∼600细胞/μl,和山肩康宁(纽约康宁)塑料盘子。互补脱氧核糖核酸向量引入HEK 293细胞(gydF4y2Ba格雷厄姆et al ., 1977gydF4y2Ba)lipofectamine转染(GIBCO-BRL)。简单地说,细胞转染3 - 5 d复苏后细胞通道,在20 - 30%(电生理学)或70 - 80%(生物化学)confluency。总cDNA用于转染的数量是相同的所有实验组和对照组。细胞转染DNA与10μg每60毫米盘生化或1μg DNA的每35毫米盘电生理实验。gydF4y2Ba
Kv通道和PTK coexpression实验,细胞cotransfected共有10μg DNA / 60毫米盘(5μg每个构建的DNA编码Kv通道和PTK;Kv通道或PTK-alone组10μg DNA的矢量控制cDNA 5μg)。Cotransfection控制细胞转染10μg向量的DNA / 60毫米。细胞培养与lipofectamine和DNA混合物稀释5人力资源serum-reduced介质(OptiMEM;GIBCO-BRL)。转染效率在平行板监控β-galactosidase反应产品的染色Lac-Z plasmid-transfected细胞表达。细胞染色效率(蓝色)通常范围从40到50%生物化学和电生理学实验的90%到70 (gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba)。gydF4y2Ba
Pervanadate准备和治疗。gydF4y2BaPervanadate准备2分钟孵化的100米gydF4y2Ba米gydF4y2BaNagydF4y2Ba3gydF4y2Ba签证官gydF4y2Ba4gydF4y2Ba和HgydF4y2Ba2gydF4y2BaOgydF4y2Ba2gydF4y2Ba,紧随其后的是稀释到适当的浓度(0 - 250μgydF4y2Ba米gydF4y2Ba在0.0008% HgydF4y2Ba2gydF4y2BaOgydF4y2Ba2gydF4y2Ba无血清MEM) (gydF4y2BaBourgoin格林,1992gydF4y2Ba;gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba)。在MEM细胞孵化与pervanadate *表示。gydF4y2Ba
缬氨霉素制备和治疗。gydF4y2Ba细胞被洗两次与血清MEM (2 ml /盘)。缬氨霉素制备5毫克/毫升乙醇和采用无血清稀释1:1000 MEM的股票。Valinomycin-treated细胞在无血清preincubated MEM含有缬氨霉素(5μg /毫升+ 0.1%乙醇)10分钟。控制细胞在无血清preincubated MEM含有0.1%乙醇汽车10分钟。所有组服用pervanadate 30分钟。pervanadate解决方案包含要么缬氨霉素(5μg /毫升)和乙醇汽车(0.1%)或乙醇汽车(0.1%)。gydF4y2Ba
细胞溶菌作用和免疫沉淀反应。gydF4y2Ba转染后细胞收获2 d细胞溶解在冰冷的Triton x - 100修改后的免疫沉淀反应缓冲区(1%gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba包含蛋白酶和磷酸酶抑制剂(25米)gydF4y2Ba米gydF4y2Ba三羟甲基氨基甲烷、液pH值7.5,150米gydF4y2Ba米gydF4y2Ba氯化钠、100米gydF4y2Ba米gydF4y2Ba氟化钠,5米gydF4y2Ba米gydF4y2BaEDTA, 1米gydF4y2Ba米gydF4y2BaNagydF4y2Ba3gydF4y2Ba签证官gydF4y2Ba4gydF4y2Ba1% Triton x - 100 1 mgydF4y2Ba米gydF4y2BaPMSF、1μg /毫升亮抑酶肽和抑肽酶2μg /毫升)。细胞溶解产物被离心澄清(15000×gydF4y2BaggydF4y2Ba;5分钟;4°C)。从上层清液对免疫沉淀物溶解产物蛋白质,我们事先批准的上层清液30μl蛋白质A / G(每毫升的皮尔斯,罗克福德,IL)细胞溶解产物在4°C和1小时之后与隔夜孵化5μl每毫升的抗体细胞溶解产物在4°C。抗体和蛋白复合物被孵化的30μl蛋白质A / G(皮尔斯)每毫升的细胞溶解产物1小时在4°C。免疫沉淀反应是洗了三次冰冷的0.1% Triton x - 100修改后的免疫沉淀反应缓冲缓冲洗(200卷每卷免疫沉淀物颗粒)。溶菌产物样品和洗免疫沉淀反应稀释在SDS-gel加载缓冲区(gydF4y2BaSambrook et al ., 1989gydF4y2Ba)。每个培养皿中总蛋白质含量(由皮尔斯BCA蛋白质化验)是为所有使用编码DNA转染条件等价。gydF4y2Ba
免疫印迹和放射自显影图程序。gydF4y2Ba蛋白质酪氨酸磷酸化测量通过免疫印迹分析,使用专门识别phosphotyrosine抗体。蛋白质(10μg / lane)对丙烯酰胺凝胶10% sds - page分离。凝胶与考马斯亮蓝染色加工,或银染色,或被electrotransferred硝基墨迹图(gydF4y2BaSambrook et al ., 1989gydF4y2Ba)。这些墨迹被封锁在5%脱脂牛奶和孵化隔夜主要抗体在4°C。然后他们被孵化与辣根过氧化物酶(合)共轭二次抗体(Amersham,阿灵顿高地,IL)在室温下2小时。增强化学发光(发射极耦合逻辑;Amersham)暴露在XAR-2电影(柯达,罗彻斯特,纽约)被用来可视化标记蛋白质。信号的大小直接关系到HRP-conjugated二级抗体的量。Coomassie蓝色和silver-stained凝胶和电影放射自显影图分析微使用Bio-Rad模型gs - 670成像密度计(大力神,CA)。相对微密度值测定由串行线性稀释的蛋白质样品用于西方的免疫印迹。gydF4y2Ba
酪氨酸磷酸化蛋白质被免疫印迹检测用鼠标单克隆抗体4 g10(纽约北部的生物技术,普莱西德湖)和PY20(肯塔基州列克星敦转导实验室)。Kv1.3表达式验证了通过使用兔多克隆抗血清免疫印迹慷慨提供的詹姆斯·道格拉斯(gydF4y2BaCai和道格拉斯,1993年gydF4y2Ba)。表皮生长因子受体表达检测人类小鼠单克隆抗体(E12020;转导实验室)。Src表达式(表示v - Src和内源性c - Src)检测小鼠单克隆抗体(MAb327;致癌基因科学,Cambridge, MA)和兔多克隆抗血清(北部生物技术)。所有其他化学物质用于免疫印迹、免疫沉淀反应,电生理学买来σ(圣路易斯,密苏里州)。gydF4y2Ba
片记录。gydF4y2Ba宏观趋势cell-attached膜补丁记录转染后24 - 72小时使用一个Axopatch-1B放大器(轴突仪器,福斯特城,CA)。细胞可视化40×放大使用相衬水油浸物镜(Thornwood蔡司,纽约)。细胞外的解决方案包括(mgydF4y2Ba米gydF4y2Ba):150氯化钾,10玫瑰,pH值7.5,1 EGTA, 0.5 MgClgydF4y2Ba2gydF4y2Ba;吸管的解决方案包括(mgydF4y2Ba米gydF4y2Ba70):30氯化钾、氯化钠、2 CaClgydF4y2Ba2gydF4y2Ba、1 EGTA和10玫瑰,pH值7.5。从Jencons玻璃电极制作(# M15/10;雷顿的秃鹰,贝德福德郡,英国),fire-polished∼1μm和涂层尖端附近蜂蜡减少电容的玻璃。9至14 MΩ吸管抗性。gydF4y2Ba
细胞−举行80 mV,走的40 + mV脉冲潜在1000毫秒的脉冲持续时间。电压信号生成和数据捕获使用微星DAP 800/2董事会(微星实验室,贝尔维尤,佤邦)。数据跟踪线性减去漏电导。gydF4y2Ba
免疫细胞化学。gydF4y2Ba表面标记的野生型和突变体Kv1.3测量如前所述(gydF4y2BaFadool et al ., 1997gydF4y2Ba)。HEK 293细胞被镀聚-gydF4y2BadgydF4y2Ba-lysine-coated玻璃盖玻片和转染与控制向量,Kv1.3或W386F Kv1.3互补脱氧核糖核酸。转染后两天,细胞被轻轻用1%多聚甲醛固定,用PBS洗净、孵化一夜之间在4°C anti-Kv1.3胞外表兔多克隆抗血清(gydF4y2BaCai和道格拉斯,1993年gydF4y2Ba)(1:50稀释10%正常山羊血清)。盖玻片上的细胞被洗PBS,孵化与二次抗体(anti-rabbit F (ab′)gydF4y2Ba2gydF4y2Ba-FITC;1:1000;30分钟在37°C) (Tago,伯林盖姆,CA)和PBS再次清洗。盖玻片是安装在玻璃幻灯片Gelmount (Biomeda,培育城市,CA)。间接immunofluorescent标签检测通过扫描共焦显微镜(mrc - 600;Bio-Rad)使用平等的获得和光圈设置对所有图像。平行实验使用cell-impermeant rhodamine-conjugated藻红蛋白证实Kv1.3标签是在细胞表面。没有观察到与另一个相同的固定条件下染色anti-Kv1.3抗血清针对胞内抗原决定基(gydF4y2BaFadool et al ., 1997gydF4y2Ba)。gydF4y2Ba
结果gydF4y2Ba
Pervanadate-induced细胞蛋白质酪氨酸磷酸化被Kv1.3表达减弱gydF4y2Ba
我们使用不同的表达克隆Kv1.3通道的影响研究Kv通道表达蛋白质酪氨酸磷酸化。Kv1.3表情的影响内源性PTK活性检查治疗控制和Kv1.3-transfected HEK 293细胞特定membrane-permeant酪氨酸磷酸酶抑制剂pervanadate。Pervanadate抑制PTPase活动是最大15μ的一半gydF4y2Ba米gydF4y2Ba和方法饱和100μgydF4y2Ba米gydF4y2Ba(gydF4y2BaBourgoin格林,1992gydF4y2Ba;gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba)。HEK 293细胞转染控制或Kv1.3向量和2 d转染后对待pervanadate(250μgydF4y2Ba米gydF4y2Ba在无血清培养基;0-60 min)单独或中等。基底低蛋白质phosphotyrosine水平HEK 293细胞溶解产物通过sds - page免疫印迹(无花果。gydF4y2Ba1gydF4y2Ba)使用单克隆抗体phosphotyrosine。微弱的乐队的分子量180 - 190年,130年,50 - 60 kDa长时间曝光后能被探测到。这微弱的信号可以通过免疫沉淀反应放大与一个anti-phosphotyrosine酪氨酸磷酸化蛋白抗体,其次是sds - page和免疫印迹分析immunoprecipitates anti-phosphotyrosine抗体(图。gydF4y2Ba1gydF4y2BaCgydF4y2Ba)。基底蛋白质phosphotyrosine水平减少免疫沉淀反应准备从Kv1.3-transfected细胞与免疫沉淀反应准备控制vector-transfected细胞(无花果。gydF4y2Ba1gydF4y2BaCgydF4y2Ba)。蛋白质phosphotyrosine信号不是检测碱性磷酸酶处理后的免疫沉淀反应(无花果。gydF4y2Ba1gydF4y2BaCgydF4y2Ba;注意,一个artifactual乐队出现在碱性phosphatase-treated样本)。Pervanadate治疗增加蛋白质phosphotyrosine水平时间的方式(gydF4y2Ba华莱士,1995gydF4y2Ba;gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba)。Kv1.3表达减弱pervanadate-induced蛋白质酪氨酸磷酸化后测量(图15 - 60分钟的pervanadate治疗。gydF4y2Ba1gydF4y2Ba一个gydF4y2Ba,gydF4y2BaBgydF4y2Ba)。蛋白酪氨酸磷酸化的相对衰减Kv1.3表达似乎是更短的时间点(无花果。gydF4y2Ba1gydF4y2BaBgydF4y2Ba)。这可能反映了磷酸化的饱和网站在稍后时间。或者,这可能反映出钾电导下降随着时间的推移,因为抑制酪氨酸phosphorylation-induced Kv频道活动随着时间的增加(gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba)。Kv1.3-mediated减少pervanadate-induced蛋白酪氨酸磷酸化似乎并未引起的总蛋白质含量的下降,以总蛋白测定和Coomassie蓝色染色的溶解产物蛋白质sds - page(数据没有显示)。Kv1.3表情的影响几个著名的内源性蛋白激酶的蛋白质含量,包括src家族激酶,表皮生长因子受体,和蛋白激酶C,是测量。Kv1.3表达并不影响这些激酶的蛋白质含量(例如,无花果。gydF4y2Ba2gydF4y2Ba一个gydF4y2Ba)。gydF4y2Ba

Kv1.3表达减少内源性蛋白酪氨酸磷酸化pervanadate-treated HEK 293个细胞。细胞转染与cDNA控制向量(−)或Kv1.3 (+)。转染两天后,细胞治疗membrane-permeant酪氨酸磷酸酶抑制剂pervanadate(250μgydF4y2Ba米gydF4y2Ba;0-60 min)。gydF4y2Ba一个gydF4y2Ba、细胞溶解产物准备和溶菌产物蛋白质分离通过sds - page和electrotransferred硝基。探讨了免疫印迹anti-phosphotyrosine抗体(gydF4y2Baanti-PYgydF4y2Ba)。主要抗体绑定被孵化与辣根peroxidase-conjugated可视化二次抗体和ECL放射自显影法。gydF4y2BaBgydF4y2Ba、蛋白质phosphotyrosine信号量化gydF4y2Ba车道gydF4y2Ba的污点gydF4y2Ba一个gydF4y2Ba微,值是表示相对于pervanadate-treatedgydF4y2Ba车道gydF4y2Ba,没有Kv1.3表达式,在60分钟(gydF4y2BangydF4y2Ba= 4;*gydF4y2BapgydF4y2Ba≤0.05和* *gydF4y2BapgydF4y2Ba≤0.01,学生的gydF4y2BatgydF4y2Ba测试)。gydF4y2BaCgydF4y2Ba酪氨酸磷酸化蛋白质免疫沉淀反应(gydF4y2Ba知识产权gydF4y2Ba使用anti-phosphotyrosine抗体)。一半的IP样本处理碱性磷酸酶隔夜(+)。所有IP样品分离通过sds - page和electrotransferred硝基。免疫印迹(gydF4y2Ba污点gydF4y2Ba与另一个anti-phosphotyrosine抗体)探测。一个artifactualgydF4y2Ba乐队gydF4y2Ba表观分子量略大于主要酪氨酸磷酸化gydF4y2Ba乐队gydF4y2Ba存在于碱性phosphatase-treated样本。gydF4y2Ba

Kv1.3 coexpression减少v-Src-induced蛋白质酪氨酸磷酸化。HEK 293细胞转染与控制、v - src Kv1.3或Kv1.3一起v - src (cotransfection)向量。gydF4y2Ba一个gydF4y2Ba,转染两天后,细胞溶解产物都准备好了,和溶菌产物蛋白sds - page分离,转移到硝基和探测anti-phosphotyrosine (gydF4y2Ba前gydF4y2Ba)或anti-Src (gydF4y2Ba底gydF4y2Ba)抗体。gydF4y2BaBgydF4y2Ba,蛋白质phosphotyrosine信号由微量化,和值是表示相对于v - srcgydF4y2Ba车道gydF4y2Ba(gydF4y2BangydF4y2Ba= 10;* * *gydF4y2BapgydF4y2Ba≤0.005,学生的gydF4y2BatgydF4y2Ba测试)。gydF4y2BaCgydF4y2Ba表示Src蛋白质被测密度术量化,和值是表示相对于v - SrcgydF4y2Ba车道gydF4y2Ba(gydF4y2BangydF4y2Ba= 10)。gydF4y2Ba
v-Src-induced细胞蛋白质酪氨酸磷酸化被Kv1.3表达减弱gydF4y2Ba
确定phosphotyrosine Kv渠道可以减少蛋白质水平的表达诱导表达强,我们测量蛋白质phosphotyrosine水平细胞溶解产物准备从细胞表达持续活跃nonreceptor PTK v - src,有或没有Kv1.3表达式。HEK 293细胞转染与控制,与Kv1.3 Kv1.3、v - src向量或v - src向量(cotransfection)。转染后细胞收获2 d,细胞溶解产物被用于sds - page使用anti-phosphotyrosine或anti-Src单克隆抗体免疫印迹。v - src表达诱发细胞蛋白质phosphotyrosine水平大幅增加(图gydF4y2Ba2gydF4y2Ba一个gydF4y2Ba,gydF4y2Ba前gydF4y2Ba),这v-Src-induced蛋白质酪氨酸磷酸化是大大减少了Kv1.3 cotransfection(无花果。gydF4y2Ba2gydF4y2Ba一个gydF4y2Ba,gydF4y2Ba前gydF4y2Ba,gydF4y2BaBgydF4y2Ba)。相比之下,内源性和Src蛋白质表达水平影响Kv1.3表达式(无花果。gydF4y2Ba2gydF4y2Ba一个gydF4y2Ba,gydF4y2Ba底gydF4y2Ba,gydF4y2BaCgydF4y2Ba)。gydF4y2Ba
的选通特性突变Kv1.3渠道决定的能力减弱v-Src-induced蛋白质酪氨酸磷酸化gydF4y2Ba
蛋白酪氨酸磷酸化的显著衰减Kv1.3表达提高的可能性,增加钾电导PTK活性和细胞超极化的影响。消除任何蛋白质的表达的可能性是非减弱蛋白质酪氨酸磷酸化,我们coexpressed v - src与突变Kv1.3不导电通道。如果Kv通道电导衰减蛋白酪氨酸磷酸化的一个关键变量,然后人会预测,不导电Kv的表达渠道会v-Src-induced蛋白质酪氨酸磷酸化的影响不大。相比之下,high-conducting Kv的表达渠道将大大减弱蛋白质酪氨酸磷酸化。Kv渠道的传导性能可以通过单点突变(发生了巨大的变化gydF4y2BaPerozo et al ., 1993gydF4y2Ba)。gydF4y2Ba
我们建造了一个不导电突变Kv1.3通道(W386F Kv1.3)。突变在这个网站相关的假定的孔隙gydF4y2Ba瓶gydF4y2Ba表达和通道产生一个不导电通道是细胞膜有针对性的有效(gydF4y2BaPerozo et al ., 1993gydF4y2Ba)。膜片箝分析显示没有检测到当前内生水平cell-attached膜补丁HEK 293细胞转染W386F Kv1.3向量(图。gydF4y2Ba3gydF4y2Ba一个gydF4y2Ba在补丁),而宏观电流明显HEK 293细胞转染野生型Kv1.3(无花果。gydF4y2Ba3gydF4y2Ba一个gydF4y2Ba)。缺乏W386F Kv1.3当前不是因为缺乏通道蛋白表达,因为Kv1.3 W386F Kv1.3蛋白水平在所有转染条件下(图是相似的。gydF4y2Ba3gydF4y2BaBgydF4y2Ba)。此外,扫描共焦采用免疫分析HEK 293个细胞转染Kv1.3或W386F Kv1.3和标记抗体针对这些渠道胞外表显示这两个通道是有效的表达在细胞表面(图。gydF4y2Ba3gydF4y2BaCgydF4y2Ba)。Kv1.3表达的影响和W386F Kv1.3细胞蛋白质酪氨酸磷酸化比较。与大衰减v-Src-induced Kv1.3蛋白质酪氨酸磷酸化,不导电W386F Kv1.3并不影响蛋白质phosphotyrosine水平(图。gydF4y2Ba3gydF4y2BaDgydF4y2Ba,gydF4y2BaEgydF4y2Ba)。在一组相关的实验中,HEK 293细胞转染与控制,Kv1.3, v - src,或Kv1.3和v - src向量(cotransfection)在brefeldin面前,逮捕新合成蛋白质的运输在高尔基早期栈,防止蛋白质运输到质膜(gydF4y2BaKlausner et al ., 1992gydF4y2Ba)。Kv1.3表达并不影响v-Src-induced brefeldin治疗后蛋白质phosphotyrosine水平(数据没有显示)。因此,蛋白质酪氨酸磷酸化衰减Kv频道表达需要的功能进行Kv渠道在质膜。gydF4y2Ba

不传导的突变Kv1.3通道表达并不影响v-Src-induced蛋白质酪氨酸磷酸化。HEK 293细胞转染与控制、v - src Kv1.3, W386F Kv1.3, Kv1.3加上v - src,或W386F Kv1.3一起v - src向量。gydF4y2Ba一个gydF4y2Ba,Cell-attached补丁录音转染后2 d。由去极化所引起的电流电压脉冲为野生型Kv1.3 40 + mV显示,不传导的突变W386F Kv1.3 (gydF4y2BangydF4y2Ba= 10)。gydF4y2BaBgydF4y2Ba,转染两天后,细胞溶解产物都准备好了,和溶菌产物蛋白sds - page分离,转移到硝基和探测anti-Kv1.3抗体。gydF4y2BaCgydF4y2Ba细胞表面标记Kv1.3和W386F Kv1.3被间接免疫荧光扫描共焦显微镜测量,nonpermeabilizing固定条件下(gydF4y2BangydF4y2Ba= 6)。gydF4y2BaDgydF4y2Ba,免疫印迹中描述gydF4y2BaBgydF4y2Ba探索了anti-phosphotyrosine抗体。gydF4y2BaEgydF4y2Ba,蛋白质phosphotyrosine信号gydF4y2BaDgydF4y2Ba被测密度术量化;值是表示相对于v - srcgydF4y2Ba车道gydF4y2Ba(gydF4y2BangydF4y2Ba= 7;* * *gydF4y2BapgydF4y2Ba≤0.005,学生的gydF4y2BatgydF4y2Ba测试)。gydF4y2Ba
Kv通道的活性抑制酪氨酸磷酸化(gydF4y2Ba黄et al ., 1993gydF4y2Ba;gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba,gydF4y2BabgydF4y2Ba;gydF4y2Ba萨博et al ., 1996gydF4y2Ba;gydF4y2BaFadool et al ., 1997gydF4y2Ba)。值得注意的是,突变通道缺乏特定的抗酪氨酸磷酸化网站phosphorylation-induced当前抑制(gydF4y2Ba黄et al ., 1993gydF4y2Ba;gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba;gydF4y2BaFadool et al ., 1997gydF4y2Ba)。我们有一组特征Y-to-F突变Kv1.3通道,抑制比野生型Kv1.3,当他们与v - src coexpressed HEK 293细胞(gydF4y2BaFadool et al ., 1997gydF4y2Ba)。这些突变通道表达野生型Kv1.3一样的程度,以及他们的基础生物物理属性区别野生型的通道(gydF4y2BaFadool et al ., 1997gydF4y2Ba)。HEK 293细胞转染控制向量;v - src向量;Kv1.3向量;向量编码的一个突变Kv1.3渠道yyy111 - 113 fff Kv1.3, Y137F Kv1.3, Y449F Kv1.3,或Y479F Kv1.3;或v - src向量与野生型或突变通道向量。转染后细胞收获2 d,细胞溶解产物被用于sds - page和anti-phosphotyrosine单克隆抗体免疫印迹分析探索。Y-to-F突变Kv1.3渠道减弱v-Src-induced蛋白质酪氨酸磷酸化在更大程度上比野生型Kv1.3通道(图。gydF4y2Ba4gydF4y2Ba一个gydF4y2Ba,gydF4y2BaBgydF4y2Ba)。gydF4y2Ba

Coexpression酪氨酸突变Kv1.3通道减少v-Src-induced蛋白质酪氨酸磷酸化。gydF4y2Ba一个gydF4y2Ba与控制向量,HEK 293细胞转染;v - src向量;Kv1.3向量;矢量编码的一个突变Kv1.3渠道yyy111 - 113 fff Kv1.3, Y137F Kv1.3, Y449F Kv1.3,或Y479F Kv1.3;或v - src向量与野生型(+)或突变Kv1.3(−)向量(cotransfection)。转染后细胞收获2 d,细胞溶解产物被用于sds - page使用anti-phosphotyrosine抗体免疫印迹。gydF4y2BaBgydF4y2Ba,蛋白质phosphotyrosine信号由微量化,和值是表示相对于v - srcgydF4y2Ba车道gydF4y2Ba(范围的gydF4y2BangydF4y2Ba第4 - 9;* *gydF4y2BapgydF4y2Ba≤0.01和* * *gydF4y2BapgydF4y2Ba≤0.005,学生的gydF4y2BatgydF4y2Ba测试)。之间的统计比较野生型通道和各种Y-to-F变异结构。gydF4y2Ba
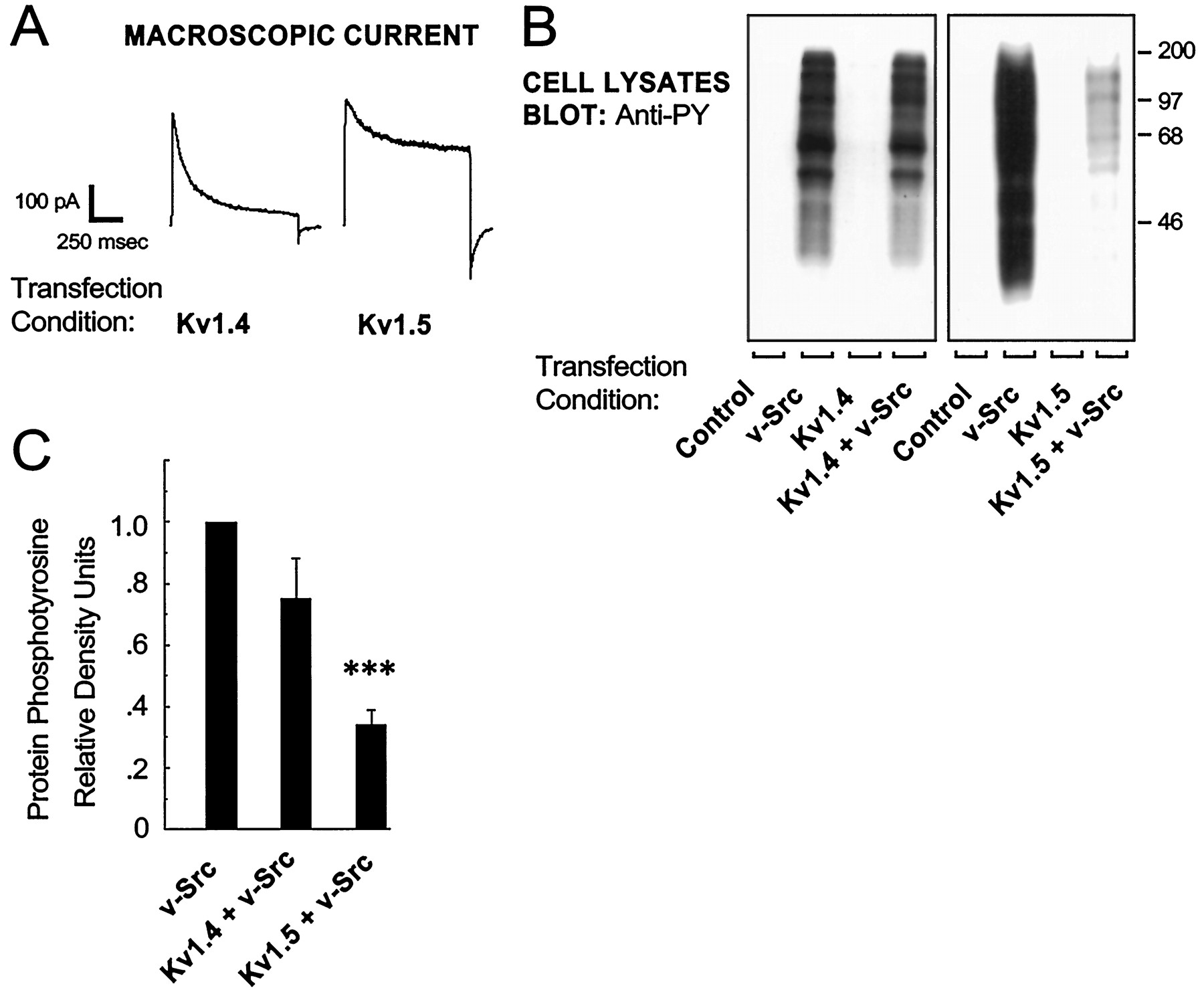
进一步测试的影响Kv通道表情v-Src-induced蛋白质酪氨酸磷酸化,我们coexpressed v - src与其他Kv渠道,表现出不同的控制特性。Kv1.4迅速灭活,通道打开后,由“锁链”机制(gydF4y2Ba钱迪和古特曼,1995gydF4y2Ba);相比之下,Kv1.5展品非常缓慢失活(无花果。gydF4y2Ba5gydF4y2Ba一个gydF4y2Ba)。快速灭活的coexpression Kv1.4通道对v-Src-induced蛋白酪氨酸磷酸化的影响不大(无花果。gydF4y2Ba5gydF4y2BaBgydF4y2Ba,gydF4y2BaCgydF4y2Ba)。相比之下,coexpression慢慢灭活Kv1.5通道的强烈抑制v-Src-induced蛋白质酪氨酸磷酸化(无花果。gydF4y2Ba5gydF4y2BaBgydF4y2Ba,gydF4y2BaCgydF4y2Ba)。gydF4y2Ba

Kv通道闸门影响v-Src-induced蛋白酪氨酸磷酸化的衰减。HEK 293细胞转染与控制、v - src Kv1.4, Kv1.5 Kv1.4加上v - src,或一起Kv1.5 v - src向量。gydF4y2Ba一个gydF4y2Ba,Cell-attached补丁录音转染后2 d。由去极化所引起的电流电压脉冲+ 40 mV Kv1.4和Kv1.5所示。gydF4y2BaBgydF4y2Ba,转染两天后,细胞溶解产物都准备好了,和溶菌产物蛋白sds - page分离,转移到硝基和探测anti-phosphotyrosine抗体。gydF4y2BaCgydF4y2Ba,蛋白质phosphotyrosine信号由微量化,和值是表示相对于v - srcgydF4y2Ba车道gydF4y2Ba(gydF4y2BangydF4y2Ba= 6;* * *gydF4y2BapgydF4y2Ba≤0.005,学生的gydF4y2BatgydF4y2Ba测试)。gydF4y2Ba
EGF受体的酪氨酸磷酸化是减毒Kv频道表达gydF4y2Ba
Kv通道表达减少pervanadate——或者v-Src-induced许多蛋白质酪氨酸磷酸化,包括著名的蛋白质的分子量185 - 190 kDa。这个尺寸范围包括表皮生长因子受体(gydF4y2BaCadena吉尔,1992gydF4y2Ba)。确定Kv通道的影响表情识别蛋白质,我们与Kv1.3 coexpressed人类表皮生长因子受体和测量蛋白质含量和不使用EGF和EGFr酪氨酸磷酸化的治疗。细胞EGF处理(10 ng / ml在无血清培养基;60分钟)或用无血清培养基和收获后EGF治疗。Kv1.3 coexpression并不影响表皮生长因子受体的蛋白质含量(无花果。gydF4y2Ba6gydF4y2Ba一个gydF4y2Ba,gydF4y2Ba前gydF4y2Ba)。仅EGFr表达了强烈酪氨酸磷酸化在缺乏外源性EGF(无花果。gydF4y2Ba6gydF4y2Ba一个gydF4y2Ba,gydF4y2Ba底gydF4y2Ba),观察到经常表达受体ptk (gydF4y2Ba陈et al ., 1997gydF4y2Ba)。表皮生长因子受体的酪氨酸磷酸化对基底的水平表示单独增加外源性EGF治疗。Kv1.3 coexpression显著降低表皮生长因子受体的酪氨酸磷酸化和没有EGF治疗(图。gydF4y2Ba6gydF4y2Ba一个gydF4y2Ba,gydF4y2Ba底gydF4y2Ba,gydF4y2BaBgydF4y2Ba)。gydF4y2Ba

表皮生长因子受体酪氨酸磷酸化是减少Kv1.3 coexpression。HEK 293细胞转染与控制、表皮生长因子受体或Kv1.3一起EGFr向量。gydF4y2Ba一个gydF4y2Ba转染两天后,细胞治疗与EGF(+)或无血清培养基(−)。细胞溶解产物都准备好,溶解产物蛋白质sds - page分离,转移到硝基和探测anti-EGFr (gydF4y2Ba前gydF4y2Ba)或anti-phosphotyrosine (gydF4y2Ba底gydF4y2Ba)抗体。gydF4y2BaBgydF4y2Ba、表皮生长因子受体表达水平和蛋白质phosphotyrosine信号在每个量化gydF4y2Ba车道gydF4y2Ba微。EGFr phosphotyrosine值归一化表皮生长因子受体蛋白水平(表皮生长因子受体phosphotyrosine /表皮生长因子受体蛋白),表示相对于表皮生长因子受体(gydF4y2Ba+ EGFgydF4y2Ba)车道(gydF4y2BangydF4y2Ba= 6;* * *gydF4y2BapgydF4y2Ba≤0.005,学生的gydF4y2BatgydF4y2Ba测试)。gydF4y2Ba
急性治疗缬氨霉素降低蛋白质酪氨酸磷酸化gydF4y2Ba
在实验报告,Kv channel-induced衰减测量的蛋白质酪氨酸磷酸化转染后2 d。确定的急性影响蛋白质phosphotyrosine水平增加钾电导,我们对待pervanadate-stimulated HEK 293个细胞的电致potassium-specific离子载体缬氨霉素(gydF4y2BaLindoy 1988gydF4y2Ba;gydF4y2Ba伍利et al ., 1995gydF4y2Ba)。缬氨霉素允许快速钾外流的细胞钾离子浓度梯度,因此敏锐地模仿Kv通道增加表达的影响。Pervanadate-induced蛋白质酪氨酸磷酸化是减毒的急性治疗缬氨霉素(无花果。gydF4y2Ba7gydF4y2Ba一个gydF4y2Ba,gydF4y2BaBgydF4y2Ba;5μg缬氨霉素/毫升的无血清培养基+ 250μgydF4y2Ba米gydF4y2Bapervanadate;30分钟)。因此,蛋白质酪氨酸磷酸化由内生ptk是降低急性治疗,增加钾电导。gydF4y2Ba

Pervanadate-induced phosphotyrosine蛋白质水平降低与电致急性治疗后钾离子载体缬氨霉素。gydF4y2Ba一个gydF4y2Ba,Valinomycin-treated (+) HEK 293个细胞在无血清培养基preincubated含有缬氨霉素(5μg /毫升+ 0.1%乙醇)10分钟,然后处理pervanadate(250μgydF4y2Ba米gydF4y2Ba;30分钟)缬氨霉素的持续存在。控制细胞(没有缬氨霉素(−)]是preincubated pervanadate-treated和无血清培养基含有0.1%的乙醇。pervanadate治疗后(+),细胞溶解产物都准备好了,和溶菌产物蛋白sds - page分离,转移到硝基和探测anti-phosphotyrosine抗体。gydF4y2BaBgydF4y2Ba、蛋白质phosphotyrosine信号在每个量化gydF4y2Ba车道gydF4y2Ba微,值是表示相对于pervanadate没有缬氨霉素条件(gydF4y2BangydF4y2Ba= 6;* * *gydF4y2BapgydF4y2Ba≤0.005,学生的gydF4y2BatgydF4y2Ba测试)。gydF4y2Ba
讨论gydF4y2Ba
先前的研究表明,膜去极化增加蛋白质酪氨酸磷酸化(gydF4y2BaStratton et al ., 1991gydF4y2Ba;gydF4y2Ba伍德罗et al ., 1992gydF4y2Ba;gydF4y2Ba西西里岛舞蹈et al ., 1994gydF4y2Ba)。我们这里显示出治疗引发膜超极化,如Kv的表达渠道或电致钾离子载体治疗,明显减弱蛋白质酪氨酸磷酸化催化内生和coexpressed ptk。蛋白质酪氨酸磷酸化的监管Kv频道活动特别感兴趣,因为它已被证明,Kv频道活动是由酪氨酸磷酸化调制(gydF4y2Ba黄et al ., 1993gydF4y2Ba;gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba,gydF4y2BabgydF4y2Ba;gydF4y2Ba萨博et al ., 1996gydF4y2Ba;gydF4y2BaFadool et al ., 1997gydF4y2Ba)。gydF4y2Ba
减少蛋白质phosphotyrosine水平可以反映ptk活性降低或增加PTPases的活动。区分这两种可能性,我们测量的影响Kv通道表达或急性电致钾离子载体治疗蛋白质phosphotyrosine水平条件下,强烈抑制PTPase活动。Kv通道表达减弱pervanadate-induced蛋白质酪氨酸磷酸化或急性缬氨霉素治疗。这些实验表明,Kv通道表达ptk活性的影响,尽管他们并没有完全消除的可能性Kv通道表达可能增加PTPase活动甚至在pervanadate的存在。观察到的Kv channel-induced减少蛋白质酪氨酸磷酸化并不能从减少PTK蛋白质或蛋白质底物水平,一致认为它反映的变化gydF4y2Ba在活的有机体内gydF4y2Baptk活性。虽然可能Kv频道表现可以降低细胞ATP水平,KgydF4y2Ba米gydF4y2BaATP的ptk几个数量级低于细胞ATP的水平。因此,调节蛋白质phosphotyrosine ATP水平的可用性似乎是不可能的。gydF4y2Ba
决定是否增加钾电导负责Kv channel-induced蛋白质酪氨酸磷酸化的衰减,我们检查了表达不导电突变Kv通道的影响。与蛋白质的健壮的衰减由野生型Kv1.3 phosphotyrosine水平的表达不导电W386F Kv1.3蛋白质phosphotyrosine水平没有影响。进一步确定是否需要Kv通道表达减弱的细胞膜蛋白质phosphotyrosine水平,我们coexpressed v - src和Kv1.3 brefeldin a Kv1.3表达式的存在并不减弱蛋白质phosphotyrosine水平在这些条件下,逮捕细胞膜的运输通道。这些结果表明,功能Kv的表达渠道在质膜需要减少蛋白质phosphotyrosine水平,表明钾电导对这个现象的重要性。gydF4y2Ba
Kv通道活动反过来降低了酪氨酸磷酸化(gydF4y2Ba黄et al ., 1993gydF4y2Ba;gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba,gydF4y2BabgydF4y2Ba;gydF4y2Ba萨博et al ., 1996gydF4y2Ba;gydF4y2BaFadool et al ., 1997gydF4y2Ba)。这酪氨酸phosphorylation-induced Kv渠道活动的减少变异Kv渠道少,缺乏一个或多个酪氨酸磷酸化位点(gydF4y2Ba黄et al ., 1993gydF4y2Ba;gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba;gydF4y2BaFadool et al ., 1997gydF4y2Ba)。因此细胞表达突变Kv渠道缺乏酪氨酸磷酸化位点表现出较大的电流比野生型Kv渠道,促进酪氨酸磷酸化的条件下(gydF4y2Ba黄et al ., 1993gydF4y2Ba;gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba;gydF4y2BaFadool et al ., 1997gydF4y2Ba)。如果Kv通道和强活动相互耦合,然后phosphorylation-resistant Kv的表达渠道应导致更大的衰减PTK活性。符合这样的预测,我们发现Kv channel-induced衰减的大小的蛋白质phosphotyrosine水平更有抵抗phosphorylation-induced抑制突变通道。进一步的证据支持的重要性Kv channel-gating蛋白酪氨酸磷酸化的属性来自实验比较迅速的影响,慢慢地灭活Kv渠道(Kv1.4和Kv1.5)。灭活Kv1.5慢慢的表达显著减弱蛋白质phosphotyrosine水平,与温和迅速灭活Kv1.4表达的影响。虽然我们不能排除这种可能性,一些其他区别Kv1.4 Kv1.5有助于这一发现,这些结果表明,该浇注的属性表示Kv渠道与能力减弱蛋白质phosphotyrosine水平。感兴趣的是表达Kv1.5但不是Kv1.4结果在一个大的细胞膜超极化(gydF4y2Ba菲利普et al ., 1993gydF4y2Ba),一致认为酪氨酸磷酸化水平耦合膜电位。但是我们不能排除其他机制的可能性,例如,细胞内钾离子浓度的变化,导致酪氨酸磷酸化的衰减。gydF4y2Ba
许多蛋白质的酪氨酸磷酸化水平降低Kv通道后的表情。表皮生长因子受体是一种蛋白质。这是特别感兴趣的,因为表皮生长因子受体的活动和其他ptk是细胞增殖密切相关(gydF4y2BaSchlessinger和乌尔里希,1992gydF4y2Ba),Kv通道超表达减少神经元的数量区分(gydF4y2Ba琼斯和Ribera, 1994年gydF4y2Ba)。值得注意的是,很难建立稳定细胞系表达Kv既定的渠道,也许是因为受损Kv channel-expressing细胞的增殖(gydF4y2Ba科伦et al ., 1990gydF4y2Ba;gydF4y2Ba铃木et al ., 1996gydF4y2Ba)。gydF4y2Ba
不等的Kv通道表达高峰转染后2 d (gydF4y2Ba施et al ., 1994gydF4y2Ba;gydF4y2Ba福尔摩斯et al ., 1996 agydF4y2Ba)。可想而知,蛋白质phosphotyrosine水平变化反映相对长期的细胞补充事件,包括基因表达的变化,可能发生在天。为了解决这个问题,我们检查了蛋白质与电致急性治疗后phosphotyrosine水平钾离子载体缬氨霉素,迅速增加钾电导(gydF4y2BaLindoy 1988gydF4y2Ba;gydF4y2Ba伍利et al ., 1995gydF4y2Ba)。急性缬氨霉素蛋白质酪氨酸磷酸化由内源性ptk治疗变弱。这些数据表明强烈的gydF4y2Ba在活的有机体内gydF4y2Ba一些ptk活性钾电导和敏感细胞休息的潜力。gydF4y2Ba
目前的结果添加到越来越多的证据表明,蛋白质酪氨酸磷酸化是由离子电导调制和细胞膜电位(gydF4y2Ba坏,格林伯格1991gydF4y2Ba;gydF4y2BaVostal et al ., 1991gydF4y2Ba;gydF4y2Ba西西里岛舞蹈et al ., 1994gydF4y2Ba,gydF4y2Ba1996年gydF4y2Ba;gydF4y2Ba列弗et al ., 1995gydF4y2Ba;gydF4y2BaRusanescu et al ., 1995gydF4y2Ba;gydF4y2BaYu et al ., 1996gydF4y2Ba)。之前的研究表明,去极化提升PTK活性和蛋白质phosphotyrosine水平。Src激酶在PC12细胞去极化激活(gydF4y2BaRusanescu et al ., 1995gydF4y2Ba),Ca Src激酶活性和表达增加gydF4y2Ba2 +gydF4y2Ba差异化的角化细胞(gydF4y2Ba赵et al ., 1992gydF4y2Ba)。脯氨酸酪氨酸激酶2 /细胞粘附激酶激活β钙流入和转染HEK 293细胞去极化,PC12细胞,海马神经元(gydF4y2Ba列弗et al ., 1995gydF4y2Ba;gydF4y2Ba西西里岛舞蹈et al ., 1996gydF4y2Ba)。相比这些早期的研究,使用治疗造成去极化和CagydF4y2Ba2 +gydF4y2Ba涌入,我们调查了影响蛋白质phosphotyrosine水平的治疗导致表达哺乳动物细胞超极化gydF4y2Ba瓶gydF4y2Ba例如Kv渠道Kv1.3, Kv1.4, Kv1.5。结果表明,不同的条件,促进向外钾离子通量减少细胞蛋白质酪氨酸磷酸化。gydF4y2Ba
之间存在的相互调制Kv渠道和蛋白质酪氨酸磷酸化建议的可能性之间的双重抑制千伏电气信号和强信号频道。有有趣的耦合影响离子通道调制和蛋白质酪氨酸磷酸化的监管。在目前的情况下,较低的条件下基底Kv通道磷酸化,Kv通道的开放会抑制弱蛋白酪氨酸磷酸化信号。相反,PTK激活增加的条件下,由外部刺激如神经递质或神经营养因子,增加酪氨酸的磷酸化Kv渠道将导致更少的通道活动和Kv的顺向救援通道抑制蛋白质酪氨酸磷酸化。这种类型的持续反馈可以迅速提高蛋白质酪氨酸磷酸化。千伏之间的这种dual-inhibition信号通道和蛋白质酪氨酸磷酸化可以调解快速开关类行为Kv通道和强的活动。gydF4y2Ba
这些结果挑衅的其他证据,链接离子通道调制和调节蛋白质酪氨酸磷酸化。酪氨酸磷酸化增加NMDA受体的活动(gydF4y2Ba王Salter, 1994gydF4y2Ba;gydF4y2BaYu et al ., 1997gydF4y2Ba);反过来,NMDA谷氨酸受体激活和治疗增加蛋白质酪氨酸磷酸化(gydF4y2Ba坏,格林伯格1991gydF4y2Ba;gydF4y2Ba西西里岛舞蹈et al ., 1996gydF4y2Ba)。电压门控l型钙gydF4y2Ba2 +gydF4y2Ba由ptk通道被激活(gydF4y2BaCataldi et al ., 1996gydF4y2Ba)。这些结果说明的可能性dual-activation NMDA受体/ Ca之间的信号gydF4y2Ba2 +gydF4y2Ba渠道和蛋白质酪氨酸磷酸化,而目前的结果表明dual-inhibition千伏之间的信号通道和蛋白质酪氨酸磷酸化。相似的离子通道之间可能存在相互耦合和丝氨酸和苏氨酸激酶和磷酸酶。候选人的例子包括II型Ca之间的夫妻gydF4y2Ba2 +gydF4y2Ba- - - calmodulin-dependent蛋白激酶和电压门控钙gydF4y2Ba2 +gydF4y2Ba频道活动(gydF4y2Ba地狱et al ., 1994gydF4y2Ba;gydF4y2Ba肖et al ., 1994gydF4y2Ba)和钙调磷酸酶磷酸酶和电压门控钙之间gydF4y2Ba2 +gydF4y2Ba频道活动(gydF4y2Ba阿姆斯特朗,1989gydF4y2Ba)。dual-modulatory电气和磷酸化信号可能是重要的调优生化和电气信号事件,影响突触可塑性等生物功能。它将确定感兴趣的动态相互作用的分子机制和细胞的后果离子channel-mediated电气信号和蛋白质酪氨酸磷酸化信号。gydF4y2Ba
脚注gydF4y2Ba
这项工作得到了国家卫生研究院的资助I.B.L.和由国家研究服务奖奖学金奖T.C.H.我们谢谢黛布拉Fadool,理查德•哈甘耐尔教授路易斯·Philipson摩根盛,詹姆斯•道格拉斯和Richard Swanson慷慨地提供抗体和互补结构,马克·鲍比不导电的电生理验证突变W386F Kv1.3, Manisha德赛Kv1.4和Kv1.5当前的痕迹,和克里斯·米勒有见地的建议。gydF4y2Ba
信件应该写给托德·c·霍姆斯Volen复杂系统中心布兰代斯大学,沃尔瑟姆,02254 MA。gydF4y2Ba
伯曼先生的礼物地址:德克萨斯大学达拉斯西南医学中心,哈里·海恩斯大道75235年达拉斯,TX。gydF4y2Ba
达冈博士的永久地址:伯纳德·卡兹密涅瓦中心,细胞生物物理学,邮政信箱9697,31096年海法,以色列。gydF4y2Ba
引用gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
在这个问题上gydF4y2Ba

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
![Pervanadate-induced protein phosphotyrosine levels decrease after acute treatment with the electrogenic potassium ionophore valinomycin. A, Valinomycin-treated (+) HEK 293 cells were preincubated in serum-free medium containing valinomycin (5 μg/ml + 0.1% ethanol) for 10 min and then treated with pervanadate (250 μm; 30 min) in the continued presence of valinomycin. Control cells [without valinomycin (−)] were preincubated and pervanadate-treated with serum-free medium containing 0.1% ethanol vehicle. After pervanadate treatment (+), cell lysates were prepared, and lysate proteins were separated by SDS-PAGE, transferred to nitrocellulose, and probed with anti-phosphotyrosine antibody. B, Protein phosphotyrosine signals were quantified in each lane by densitometry, and values are expressed relative to the pervanadate without valinomycin condition (n = 6; ***p ≤ 0.005, Student’s t test).](https://www.jneurosci.org/content/jneuro/17/23/8964/F7.large.jpg?width=800&height=600&carousel=1){kind=link}
{kind=link}







