





文摘
结节病是一种全身性炎性疾病的原因不明,受遗传和环境因素的影响。然而,到目前为止的位点确定结节病解释认为遗传的一部分。
进一步识别易感性位点,我们进行了全基因组关联分析使用Affymetrix 6.0人类GeneChip其次是验证和复制阶段。
质量控制后,637例,1233控制和677 619单核苷酸多态性(snp)是用于初步筛选。选择99个snp进行验证在一个独立的研究小组(1664名患者中,2932名对照)。SNP rs1050045与结节病显著相关(p = 0.0215)纠正验证小组和产生9.22×10的假定值−8(或1.24)的风险的荟萃分析筛选和验证阶段。一个荟萃分析的三种群来自德国、捷克共和国和瑞典证实这个发现(p = 0.024;或1.14)。精细定位和mRNA表达研究指出骨肉瘤9 (OS9放大),最可能的候选人潜在的风险因素。
OS9蛋白起着重要的作用在内质的reticulum-associated蛋白质降解和行为在toll样受体诱导髓系细胞的激活。表达分析OS9信使rna检测协会功能机制提供依据信号。
结节病(MIM: 181000)是一种原因不明的全身性炎性疾病,特点是non-caseating上皮样细胞肉芽肿。虽然可以影响任何器官系统,肉芽肿是经常发现在肺和淋巴结。发病机理的特点是高的巨噬细胞和CD4的活动+辅助t细胞接触后的监管影响下至今难以捉摸的抗原细胞因子由当地的单核吞噬细胞,t细胞、树突细胞和成纤维细胞(1,2]。根据疾病的过程中,病人可以分为受到影响急性或慢性结节病(3]。总之,急性结节病的特点是突然抱怨和恢复2年内。它包括洛夫格伦综合征,特点是结节性红斑,双侧肺门淋巴结病和多发性关节炎。相比之下,慢性结节病病人表现出微妙的早期症状加剧,紧随其后的是持久的疾病活动2年或更长时间。
结节病是一种罕见疾病的患病率约40每100 000居民在德国,和主要影响年轻人(20 - 40年)是女性中更普遍4]。它被认为是由环境和遗传因素的复杂组合,估计遗传66%5]。疾病的遗传基础支持的风险的识别基因,例如:BTNL2(6- - - - - -8),ANXA11(9,10),TNF-α(11)和几个HLA位点(12]。几个候选人等待进一步的支持例如Rab23(13和趋化因子受体CCR2, CCR5和IL23R(14]。与目前的研究中,我们旨在进一步为结节病易感性位点的确定使用Affymetrix SNP数组6.0,包括近一百万个单核苷酸多态性(SNP),因此收益率更高,而且部分不同,覆盖范围比以前公布的全基因组关联研究(GWAS) [9,13]。因为我们预期,而小尺度效应,因此,没有结果的全基因组的意义在最初的全基因组筛查,包括一个独立的验证步骤和一个复制步骤后多级设计(在线补充图E1)。
材料和方法
患者和对照组
结节病病人分为慢性或急性结节病,如前所述[9,15,16),根据疾病的课程和演示基于所有可用的信息(调查问卷完成病人和医生,医院记录和访谈信息)。短暂,巧妙地加剧早期症状之后,持久的疾病活动2年或更长时间定义了慢性结节病样本(称为“慢性”)。患者在急性结节病样本(“急性”)遭受突如其来的投诉和2年内恢复。只有病人可以明确归类为急性或慢性subphenotypes被招募。因此,对于分类为急性和慢性,疾病过程中至少2年进行了分析。所有的病人显示胸腔疾病介入的证据。
质量控制之前,检查小组由640名患者,其中包括191 401年急性和慢性病人,和1256名对照组。面板与面板几乎完全重叠(也称为面板)之前的结节病协会中使用屏幕使用Affymetrix 5.0 GeneChip [9]。此外,部分筛选面板前协会已经被用于分析(6,17]。1664年验证、结节病病人,包括563人急性和慢性结节病,947人和2932名健康的人在质量控制之前,一起形成面板b复制面板我由303年德国结节病病人和281个对照组和没有重叠与其他面板。subphenotype状态信息仅用于有限数量的患者(急性:n = 40;慢性:n = 61)。复制面板C-II包括267例结节病病例和330例对照样本来自捷克共和国和大幅重叠(其它地方描述的10]。没有可用于此示例subphenotype信息。瑞典样本(复制面板C-III)由招募在门诊1066例肺的卡罗琳斯卡大学医院,桑纳,瑞典,其中333名患者被诊断出患有洛夫格伦综合征,急性的结节病。其余病例洛夫格伦综合征是排除或没有subphenotype信息是可用的。940年瑞典控制由风湿性关节炎的瑞典流行病学调查(EIRA)研究[18]。精细定位(D)是1829年德国结节病病人,包括所有的病人从面板B和部分面板,包括597敏锐和1055长期影响患者,1465年,德国控制面板B .结节病的患者急性课程总共有123个人显示古典洛夫格伦综合征的症状。招聘和诊断的病人在电池板A, B, c和D都完成如上所述。所有的研究参与者在面板,B, c和D都起源于德国。在线诊断和招聘详情,请参见补充材料方法部分。45结节病病人和45个人控制面板(E)用于测序的OS9基因区域选择从精细定位板D为了丰富rs1050045风险等位基因携带者的,因为这些人有更高的机会(s)携带致病变种,认为是在连锁不平衡与rs1050045 (LD)。
基因分型和质量控制
A和B基因型的面板都使用了Affymetrix人类全基因组SNP数组6.0(美国Affymetrix,圣克拉拉,CA)和SNPlex™技术(美国应用生物系统公司,培育城市,CA),分别。额外的板我做基因分型,C-II C-III, D和E进行了使用Taqman技术(应用生物系统公司)。保守和建立质量过滤器使用惯例(后19];对每个小组的质量控制和基因分型的细节看到在线补充材料。简而言之,所有人都必须小于10%丢失的基因型。样品显示证据的关联性(相同的状态值> 0.8;看到网上补充图E2)其他样本从数据集中删除。为每一个面板(筛选、验证和复制),单核苷酸多态性是检查缺失基因型(排除阈值:< 95%病人或对照组),轻微的等位基因频率(< 2%的病人或对照组),偏离哈迪温伯格平衡控制样本(p≤0.01),导致257 349标记的排斥从GWAS数据集(27.5%)。任何SNP选择验证进行目视检查集群的情节(参见图E3在线补充集群铅SNP rs1050045的情节)。
SNP选择和统计分析
那些排名最高的假定值的标记的全基因组关联分析的面板和至少一个附加相关SNP (r2> 0.5 p < 10−3)是当前被选中进行验证。标记的HLA区域(6 p21.1-6p21.3),从ANXA11基因区域(chr10、位置81 850 - 82 000 kb)不包含在验证阶段,因为一个强大的疾病相关的位点已经建立了基于相同的研究人群(6,9]。统计分析的基因型数据进行了使用叮铃声v.1.06 [20.除非另有规定。在整个实验中,single-marker allele-based协会分析使用卡方测试(1自由度)。LD的可视化进行了用金(21]。逻辑回归,向后模型选择使用AIC (22使用哈普罗)和单体型分析。统计包(23)进行了R v2.10.1 v2.15.0 [24]。人类基因组单体型图标记snp被选为精细定位使用Haploview [25]。人群归因危险度(PAR) GWAS铅SNP是使用以下公式计算:PAR = (f * (rr-1)) / (f * (rr-1) + 1), f表示等位基因频率在人口和rr =风险等位基因的相对风险,相应的估计的优势比(26]。详细描述请参阅在线补充材料方法部分。人口分层校正板进行了A和B使用估计基因组通货膨胀因素为1.149。荟萃分析的小组进行了A和B使用逆常规方法(27),而我复制的荟萃分析面板,——和iii是使用固定效应模型中实现叮铃声v.1.07。
交互分析使用上位选项的叮铃声v.1.07。一个重要在这个测试结果表明偏差相关的snp的乘法模型。
此外,标记的关联分析是调整之前报道的影响标记使用逻辑回归模型(见在线补充表E1)。所有标记被认为是在一个基因型的风险模型。向后统计模型选择使用R和阶跃函数的执行是基于默认Akaike信息准则(AIC)。只有样品,没有缺失基因型在考虑任何标记都包含在这一分析(637例,1233控制)。意义是由一个似然比评估测试。
桑格测序
其实,exon-flanking和监管的区域OS9使用标准桑格测序DNA测序技术ABI棱镜3700分析器(应用生物系统公司)。引物设计使用Primer3 [28]。引物序列给出E2在网上补充表。
通过PCR分析组织的表达
候选基因的组织表达模式的调查我们使用商用组织和免疫细胞小组从Clontech(美国帕洛阿尔托,加州)和半定量PCR。表情是正常GAPDH表达式。各自的引物序列见表E3在线补充。
拜尔港准备,信使rna隔离和存在
支气管肺泡灌洗(BAL)细胞BAL面板样品我是匹配的部分肺泡巨噬细胞(有关详细信息,请参阅在线补充材料)。从snap-frozen BAL细胞总RNA分离使用商业套装(RNeasy、试剂盒、希尔登,德国)和使用优势互补脱氧核糖核酸是合成RT-for-PCR工具包(Clontech实验室)根据制造商的协议。给出有针对性的引物的序列中存在在网上补充表E3。记录被正常化GAPDH信使rna水平。目标基因的相对表达水平检测结节病病人和影响个体之间的显著差异(n = 4)使用非参数Mann-Whitney紫外线测试中实现Graphpad统计软件(Graphpad, Inc .,拉霍亚,CA,美国)。
第二个BAL一系列BAL-samples获得46名患者的活跃结节病和八个控制面板(BAL II)。BAL细胞涂片干并使用May-Grunwald-Gimsa染色法染色。细胞差异是由至少200个细胞计数。分析人类白细胞antigen-DR (HLA-DR)对淋巴细胞表达细胞被固定在保利-l赖氨酸涂布幻灯片,孵化与单克隆抗体针对HLA-DR浓度由供应商和开发建议peroxidase-antiperoxidase技术。引物的序列用于E4中存在有在线补充表。引物不区分的三个已知亚型OS9信使rna。周期的数量GAPDH和OS9都是平等的(35周期)。阈值周期值计算(Ct)和用于计算相对表达(重新)mRNA水平为每个示例使用以下公式:= 2(CtGAPDH-Ct-Target)×10 000。相对表达式作为dimension-free比率。统计分析的执行队列使用StatView (SAS研究所卡里、数控、美国)使用Mann-Whitney紫外线测试。从血液中提取DNA, pcr进行如上所述。球样品和更多细节的描述参见在线补充材料方法部分。
免疫组织化学
部分肺组织固定与HEPES-glutamic酸buffer-mediated有机溶剂从匿名保护效应正常对照组(n = 4)和活跃的结节病(n = 8)沾兔多克隆抗体OS9 (nb100 - 519 b;罗福斯biotechnicals,利特尔顿,美国公司)使用标准协议的主要抗体稀释1:10 0 (29日]。省略使用无关的主要抗体和抗体主要担任正面和负面的控制,分别。显微照片拍摄在蔡司Axio成像仪Z1(蔡司、从、德国)。
结果
GWAS分析
后将保守和建立质量过滤器应用到数据集,1870个样本(面板:637例中,有1233控制)和677年619个snp包含在最初的全基因组筛查。评估人口异质性在面板一个温和,基因组通货膨胀系数λGC基于卡方分布中值(= 1.1530.),λGC= 1.0对应没有通货膨胀。QQ-plot和关联信号给出了已知的结节病风险位点图E4在线补充材料和补充。
除了snp进入验证阶段(见下文),多个snpBTNL2基因(rs2076533、rs2076530 rs9268480 rs3806156)和在几个HLA位点(rs7194 rs7195、rs3177928 HLA-DRA;rs9277550、rs1431403 rs2856816 HLA-DPB1;rs2071475、rs2071473 HLA-DOB) 6 . 3号染色体上(在线补充图E5),该地区的特点是高LD的模式,被发现在GWAS明显与结节病有关。各自的单核苷酸多态性密切相关,名义上的假定值介于1.01×10−15(rs2076533)和9.17×10−5(rs2856816)。
验证铅变体
通过预定义的选择标准是99个snp基因分型在一个独立的验证样本(面板B)。质量控制后,2770年德国控制和1572年德国结节病病人,其中包括894 530年慢性和急性病人,组成99人洛夫格伦综合征,是包括在分析中。21标记显示一个名义上的重要与结节病在验证阶段(表1)。一个变种,rs1050045,位于12 q13.3-q14.1与结节病与未修正的假定值为7.38×10−5。因为筛选面板和验证小组B源自相同的德国人口,我们假定一个通货膨胀的测试统计由于人口分层λ= 1.15的板相结合,作为小组的结果估计这种影响校正后,rs1050045依然显著Bonferroni调整后为多个测试(假定值的修正(p相关系数)= 99×2.18×10−4= 0.0215;或1.20,95%可信区间1.10 - -1.31)。苏格兰民族党授予票面的8% - -12%(基于获得的频率在验证和筛选面板中,分别)。a和B的荟萃分析板与p与结节病有关相关系数9.22×10−8(或1.24)[31日]。没有明显SNP-SNP交互的SNP与结节病的易感性变异ANXA11,BTNL2, Rab23和IL23R位点观察面板(数据未显示)。
为了确定rs1050045施加一个统计独立对结节病之前报道敏感性变异的影响BTNL2,ANXA11,Rab23,IL23R和HLA,我们执行一个逆向模型选择基于AIC的逻辑回归模型。除了标记rs1050045之外,另一个21标记进入前的模型选择,即:rs6664119 (IL23R);rs644045 (chr6 p21.33);rs9268402、rs9391858 rs2076533 rs2076530(所有BTNL2);rs3177928、rs7194 rs7195(所有HLA-DRA);rs502771 (HLA-DRB1/5);rs4530903 (HLA-DRB1 / DQA1地区);rs9275371、rs9275418 rs2856717、rs9275522 rs9275523(所有hla dq地区);rs9277550、rs3117242 rs3128923(所有HLA-DPB);rs3957366 (BEND6);和rs1953600 (ANXA11)。所有标记被认为是在一个基因型的风险模型。最终的模型中,除了rs1050045,标记rs6664119 rs644045, rs2076533, rs3177928, rs502771, rs9275371, rs9275418, rs2856717, rs3128923, rs3957366 rs1953600。调整后的其他11个标记,rs1050045仍在与结节病显著相关(p = 6.2×10−4)。
Subphenotype-specific分析显示一个更强大的协会rs1050045急性结节病人口分层(校正后纠正p急性= 6.75×10−4;或1.30,95% CI 1.14 - -1.49)相比,慢性subphenotype(纠正p慢性= 0.021;或1.16,95%可信区间1.04 - -1.30)。洛夫格伦综合征患者(纠正p显示更强的影响洛夫格伦= 0.044;或1.41,95%可信区间1.05 - -1.88)。基因型是验证使用TaqMan SNP基因分型作为一个独立的技术基因型一致(98.74%)。风险控制C等位基因频率为42%,46%的情况下,48%患者急性结节病病例和50%洛夫格伦综合征。以下所有snp基因分型结果详细的结果包括计数研究验证阶段提出了E5在网上补充表。
独立样本中复制不同的欧洲人
为了复制检测协会领导SNP rs1050045结节病,我们执行一个荟萃分析的SNP独立病例对照样本来自德国、捷克共和国和瑞典(分别面板我C-II和C-III)。变异显示重要的协会与结节病荟萃分析这三个样本集(p = 0.023;或1.14)。风险等位基因频率(和相应的优势比)之间的不同人群,从41.9%的控制与43.1%的情况下(或1.05,95% CI 0.83 - -1.32)在面板我37.7%与40.1%(或1.11,95% CI 0.97 - -1.26)在面板C-III高达42.7%与48.6%(或1.27,95% CI 1.00 - -1.61)在面板C-II。subphenotype分析面板我没有承诺由于小样本大小(< 15%),和没有可供subphenotype信息面板C-II。在面板C-III尽管,我们发现一个重要的协会与洛夫格伦综合征,急性结节病的子表单(p = 0.015;或1.26,95%可信区间1.05 - -1.52)。
精细定位在rs1050045(染色体12 q13.3-q14.1)
除了领导SNP rs1050045,我们选择从人类基因组单体型图57标记SNP CEU∼500 kb的精细定位12 q13.3-q14.1 rs1050045周边地区在细绘图面板d质量控制后,53个单核苷酸多态性的基因型1753例,包括570 1016年急性和长期影响患者和1429控制个人分析。22标记了协会的名义p值< 0.05的分析。再次,最强的协会观察信号与铅SNP rs1050045 (p = 1.10×10−4;或1.22,95%可信区间1.10 - -1.35)。完整的分析结果显示在在线补充表E6。图1概述协会的信号,基因,重组率和LD结构12 q13.3-q14.1轨迹,表明协会信号的强度随遗传和物理距离的增加而逐渐减小的SNP rs1050045,位于3′utr的骨肉瘤9 (OS9放大)基因。在面板D,这标志与rs11172300强大的连锁不平衡(r2在控制)= 0.96,位于12 kb的上游OS9和与结节病也显著相关(p = 1.5×10−3;或1.18,95%可信区间1.06 - -1.30)。
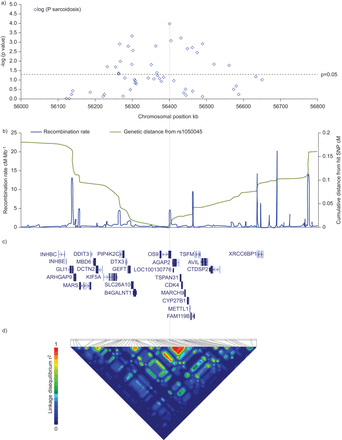
从之前的页面。区域的情节在12 q13.3-q14.1结节病协会进行验证。一)Nominal-log10假定值从一个等位基因的获得卡方测试一个自由度的精细定位板53 d标签单核苷酸多态性(SNP)基因分型在500 kb的地区周围的GWAS铅SNP rs1050045(突出了橙色的垂直线)。黑色虚线对应于一个阈值为0.05。b)复合强度的情节厘摩每megabase铅的累积遗传距离在厘摩SNP根据人类基因组单体型图(CEU三人小组,第一阶段+ 2)。c)位置和intron-exon结构记录根据美国国家生物技术信息中心(NCBI)的参考序列(RefSeq)集合。(r d) rPairwise连锁不平衡值2)在控制个体。作为构建36 NCBI给出坐标位置。
五个单核苷酸多态性,即rs1689585、rs1628552、rs4760168 rs7979246 rs10783844,位于rs1050045和rs11172300之间OS9基因区域,彼此强烈的LD (r2= 0.91 - -0.99),但无论是与铅SNP rs1050045还是rs11172300。这五个snp之一,即rs1689585,显示一个名义上的重大协会与普通结节病表型(p = 4.0×10−2;或0.90,95% CI 0.81 - -1.00)和一个非常强大的协会在急性子样品(p急性= 5.0×10−4;或0.77,95%可信区间0.66 - -0.89)。剩下的四个snp只急性结节病显著相关(见在线补充图E6和表E6)。单体型分析这七个snp (rs1050045、rs11172300 rs1689585, rs1628552, rs4760168, rs7979246和rs10783844)之间的单体型频率分布显示显著差异结节病病人和控制(p = 9.1×10−3)和急性患者subphenotype相比控制(p = 4.7×10−3)。看到网上补充表E7完整的结果。然而,逆向模型选择一个逻辑回归模型,运用AIC给不确定的结果是否rs1050045代表该地区表型关联的唯一来源(数据没有显示)。预测功能的后果使用国家环境卫生科学研究所(NIEHS) SNPinfo web服务器(32)透露可能的影响这七个单核苷酸多态性有关,和单核苷酸多态性强LD (r2> 0.9),在转录因子结合位点以及microrna的结合位点。看到网上补充表E8完整的结果。
测序OS9编码区域
为了验证现有的变异和小说来确定突变位点有关,我们测序OS9监管和其实地区45个结节病病人和45个人控制面板(E)利用Sanger测序技术。我们发现七个已知和两个新颖的单核苷酸多态性,包括外显子2,非同义SNP命名OS9-SNP1 (表2和在线补充图E7)。的外显子测序10失败由于技术原因。两个九检测单核苷酸多态性(OS9-SNP1和rs74368191)基因型和调查经细绘图面板D,都使用Taqman技术。OS9-SNP1是极其罕见的变异,与一个控制个人是唯一的杂合的载体。rs74368191,只发现的情况下在测序面板E,略更频繁的情况下(小等位基因频率(加)= 0.014)比在控制面板D(加= 0.011),但表现出较弱的效果比铅SNP rs1050045 (OR = 1.22;p > 0.05)。
候选基因的表达分析染色体12 q13.3-q14.1地区
基于精细定位实验的结果,我们选择八个基因附近的SNP作为候选人的易感性基因驾驶协会信号(图1)。我们提出,致病变种(s)可能会影响对结节病通过改变一个或多个这些基因的表达水平,即OS9,ArfGAP GTPase域,锚蛋白重复和PH值域2(AGAP2),tetraspanin 31(TSPAN31),细胞周期蛋白依赖性激酶4(到),膜相关无名指(C3HC4) 9(MARCH9),亚科B细胞色素P450,家庭27日,多肽1(CYP27B1),甲基转移酶1(METTL1),家庭使用序列相似性119,成员B(FAM119B)。来证明这一假设,我们首先评估表达式的八个记录在健康组织和免疫细胞类型相关的半定量PCR结节病的发病机制。所有记录显示中度到高表达在健康的肺部组织。一些记录,即OS9,AGAP2, TSPAN31和MARCH9激活单核细胞被抑制,CD4细胞+和CD8+t细胞休息同行相比(在线补充图E8)。
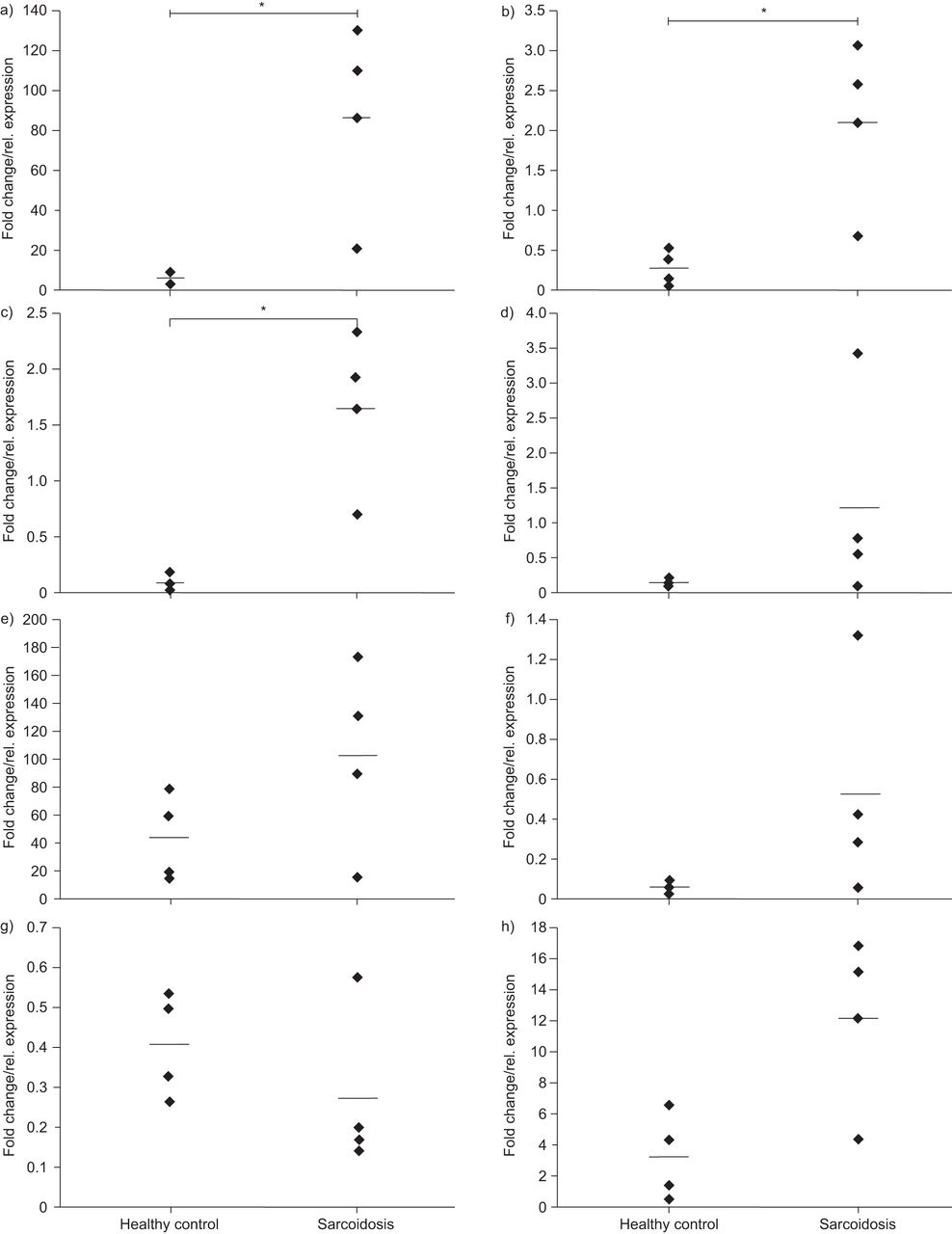
我们进一步假设,致病变种(s)可能带来风险增加结节病通过改变一个或多个这些基因的表达水平。因此我们分析候选基因的表达水平在BAL细胞结节病病人和影响人(BAL面板,n = 4 /集团)通过执行中存在的互补。所有八个候选基因的表达在BAL细胞检测,同时表达OS9,TSPAN31和FAM119B在结节病BAL显著增加样本相对于控件(每个名义p = 0.029,图2)。
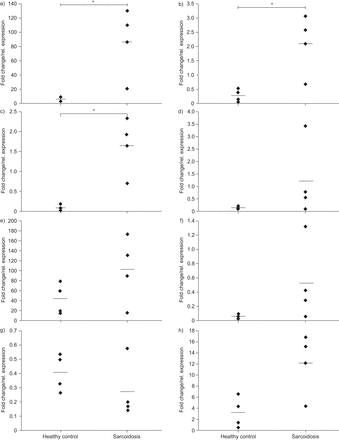
从之前的页面。八个候选基因的表达在支气管肺泡灌洗(巴尔斯)结节病病人(n = 4)相比,影响个人(n = 4, BAL面板)。转录水平的)OS9,b)TSPAN31c)FAM119Bd)AGAP2,e)到f)MARCH9,g)CYP27B1和h)METTL1在正常GAPDH信使rna水平。*:意义在百分之十五水平基于非参数Mann-Whitney紫外线测试。
Allele-specific OS9的表达和免疫组织化学
从精细定位实验的结果和表达的研究,OS9似乎最有希望的候选基因在相关地区。因此,我们研究了OS9表现在独立BAL面板。在这里,表达OS9信使rna中检测出8个8名健康对照组但只有38 47个结节病的患者使用中存在。更详细的分析表明,在病人表达OS9,被称为OS9艾滋病患者从现在开始,OS9表达显著高于控制(3774±2794 (n = 38)与1587±857 (n = 8)、在线补充图E9;p = 7.0×10−3)。自从BAL的结节病病人的特点是低比例的肺泡巨噬细胞(33),有趣的是,BAL细胞成分之间的不同OS9艾滋病患者和患者不表达OS9(称为OS9-negative患者)和控制对肺泡巨噬细胞的百分比(在线补充图E10汽油)。两组不同也明显对HLA-DR的百分比+t细胞的比例最高OS9负例(在线补充图E11)。分层球结节病病人样本根据基因型在铅SNP rs1050045显示显著负相关OS9信使rna表达水平与CC基因型(AA: 2879;交流:3078;答:824;CC与AA p = 0.019;CC与AC p = 0.016;图3)。
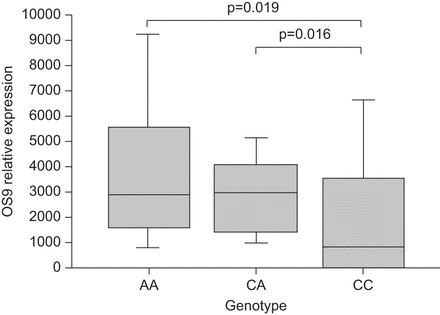
Allele-specific OS9的表情。转录水平的OS9测量使用定量rt - pcr的患者支气管肺泡灌洗面板II (n = 46)。信使rna表达模式的改变的基因型纯合子风险显著相关rs1050045“CC”(p = 0.016)。
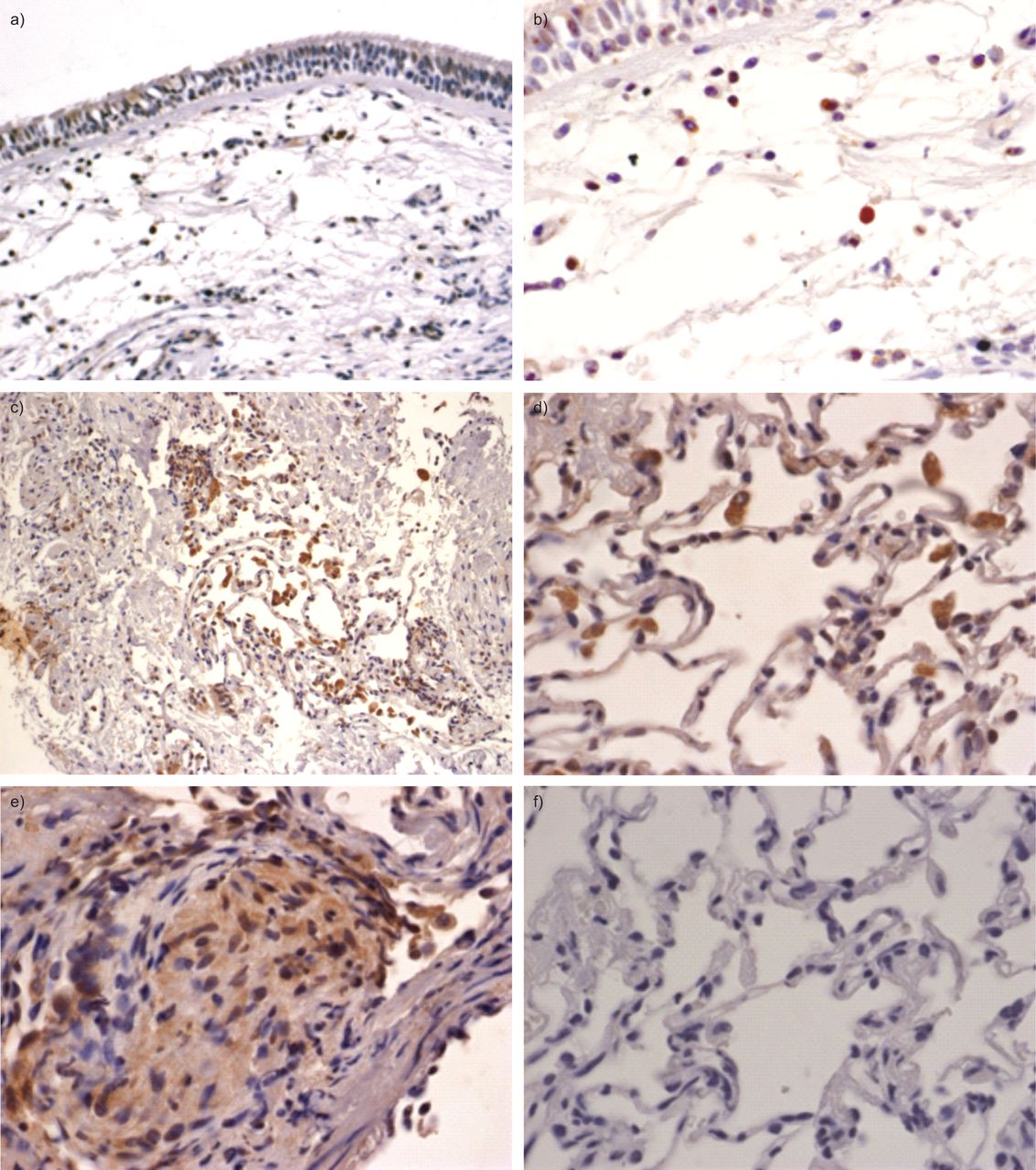
接下来,我们验证了原位本地化OS9蛋白免疫组织化学表达的肺活检的第三组(n = 4正常对照组;n = 8活跃的结节病病人)。无处不在的OS9蛋白表达可以观察到明显细胞核周围的和颗粒胞质染色模式并行的报道endoplasmatic网(ER) OS9本地化。强烈的免疫反应性可以观察到在肺泡巨噬细胞和淋巴细胞细胞病变样本和正常对照组。结节病肉芽肿结构的活检也OS9呈阳性(图4)。
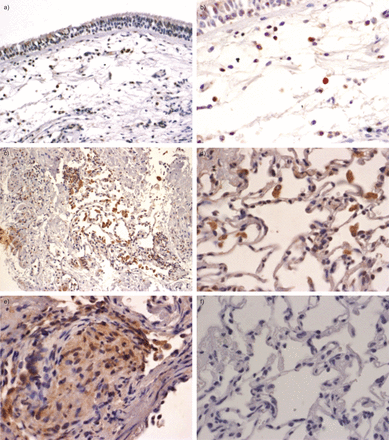
OS9免疫组织化学染色。染色的OS9肺活检材料)呼吸道上皮细胞和b)黏膜下层的健康话题。注意强OS9单核细胞的染色(棕色)和淋巴细胞的细胞。c和d)从结节病病人标本展示积极的肺泡巨噬细胞的染色。e)在结节病肉芽肿性结构的分散颗粒染色标本。f)负控制相邻部分的d)。图片显示代表结果n = 4健康受试者和n = 8结节病病人。核与苏木精复染色。
讨论
我们进行了全基因组关联分析的677年的637 619个snp德国结节病病例和1233控制和确定一个新的染色体在结节病易感性位点12 q13.3-q14.1,德国病例对照人口在一个独立的验证。协会领导SNP rs1050045荟萃分析的三个独立的群体间的是来自德国、捷克共和国和瑞典(面板我,C-II和C-III)。Subphenotype-specific分析发现强关联的SNP与结节病的急性子表单与结节病表型。
轨迹被忽视了在前面的全基因组的研究由于缺乏覆盖的地区和不同的分析策略尽管使用几乎相同的主要研究人群(9,13]。
该地区12 q13.3-q14.1报道之前与类风湿性关节炎(34- - - - - -36),SNP rs1678542显示类似的效应对于结节病(-0.94或0.88),以及1型糖尿病(37- - - - - -39),多发性硬化(40和腹腔疾病41]。因此代表第一个临床遗传风险共享的轨迹截然不同的疾病。然而,由于相关地区港口的潜在风险基因,没有任何提到的疾病进行了精细定位除了结节病,没有结论可以得出不同的疾病是否共享单个风险因素或在该地区受到不同因素的影响。
结节病、地区的精细定位和表达研究表明OS9作为最有可能的候选人的潜在风险基因。其实和exon-flanking区域的测序OS9显示两个新颖的snp,但没有明显的候选致病变种(年代)。除了rs1050045,产生最强协会调查了SNP的精细定位,只有一个额外的SNP在3′utr OS9被发现。由于16个snp注释在这个区域,根据dbSNP建立132年,14岁的没有发现,这个结果会显著降低潜在的数量监管3′utr SNPs和强调rs1050045的潜在功能的相关性。
在网上最密切相关的功能分析snp OS9地区建议对基因表达产生影响通过改变转录因子以及microrna的绑定和剪接事件。SNP rs799265与rs1050045完美LD (r2= 1根据人类基因组单体型图27)显示高监管潜力以及最大保护在17脊椎动物做这个SNP一个有趣的候选人进行进一步的功能分析。分析OS9信使rna从BAL面板,只包含与高比例的巨噬细胞样本,发现升高OS9表达水平在结节病病人相比,控制。这一发现被证实在患者的一组中落下帷幕小组第二,特点是高百分比的巨噬细胞。此外,我们发现没有OS9信使rna伴有CD4的比例降低+/ CD8+t细胞。虽然不明显,这一发现可能表明OS9参与结节病的活动状态,它的特点是CD4很高+/ CD8+比(33),而缺乏OS9信使rna可能与结节病的慢性和更少的活性形式。初步调查的潜在allele-specific表达式OS9在拜尔(面板II)显示出显著的减少OS9表达rs1050045风险等位基因的纯合子运营商。可能的机械解释这个微分表达式包括更短的半衰期OS9信使rna或一个不同的绑定微rna (microRNA) -145的行为。这个监管microrna的结合3′utrOS9。它是表达人类气管和肺组织根据UCSC基因组浏览器(42),已被证明是参与气道炎症小鼠(43]。最近发布的结果显示高2.8倍的microrna的表达完整的血液结节病的患者与健康对照组(p = 3.9×10−3)[44]。进一步功能实验现在才能阐明功能合理的候选致病变种的后果,如。rs1050045或rs799265。
假定的风险基因OS9由15个外显子和编码四个不同的剪接变体(45,46),所有这些都含有甘露糖6-phosphate受体同源域(47]。ER-associated OS9蛋白起着重要作用的蛋白质错误折叠或未装配的退化(48,49]。这可能是重要的,因为只有肽来源于蛋白质在人体中发现了常见的槽HLA-DR的肺泡巨噬细胞结节病病人在最近的一项研究[50]。此外,它表明,这些分子诱导抗原t细胞反应(51]。因此OS9可能作为一种自身免疫性组件在结节病的免疫发病机理。此外,它最近表明OS9与树突特异性跨膜蛋白的胞质尾在toll样receptor-induced成熟的树突细胞在骨髓分化OS9暗示作用,细胞融合(52]。讨论了树突状细胞为结节病免疫学的重要介质53)和广泛调查关于结节病的各个方面(53- - - - - -56]。
但是,根据我们的研究结果,我们不能排除基因以外的12个q13.3-q14.1功能变化OS9影响对结节病的易感性。从临床的观点来看,CYP27B1代表了一个进一步的竞逐结节病易感性为多发性硬化症(类似的讨论40]。这个基因编码细胞色素P450超家族的一员。酶激活维生素D3,一般的免疫调节功能(57,58),也在肺部免疫过程中发挥作用,在结节病59,60]。此外,这种基因最近一直报道差异表达在结节病progressive-fibrotic,患者相比,具有自限性,病程(61年]。自整个12 q13.3-q14.1轨迹显示subphenotype-specific遗传协会模式在我们的研究中,可以想见,在相关地区特定的基因变异可能影响CYP27B1表达式的监管限制在某些阶段或疾病的亚型。到目前为止没有明显的其余的基因产物功能的含义,即AGAP2, TSPAN31,到,MARCH9, METTL1 FAM119B,结节病发病机理可以从文学。
总之,这是第一次报告的染色体区域协会12 q13.3-q14.1结节病。精细定位的地区和初步研究显示表达式OS9作为底层的易感性基因最有可能的候选人和自体免疫反应可能支持的概念在结节病的免疫发病机理。根据本文提供的数据,更详细的研究报告基因协会和OS9函数需要在结节病现在的背景下,为了定义致病变种(s)和分子机制我们的观察。此外,类似的研究需要完成不同的种族背景,以确定患者的祖先与新结节病易感性位点。
确认
作者要感谢所有的患者,家属和医生合作。德国结节病病人的支持组织(德意志Sarkoidose-Vereinigung汽车集团),PopGen生物库和贡献位肺脏。我们感激地承认技术援助人员的临床分子生物学研究所(德国基尔)。狮子座Padyukov(风湿病单位,医学部门,卡罗林斯卡研究所,斯德哥尔摩,瑞典),贡献与控件的选择瑞典队列和作者感谢瑞典EIRA研究成员控制样本的集合。
作者的从属关系如下。s·霍夫曼:临床分子生物学研究所Christian-Albrechts大学、基尔,德国;费舍尔:临床分子生物学研究所Christian-Albrechts大学、基尔,德国;m .测量:医学信息学研究所和统计,Christian-Albrechts大学;德国基尔;g·雅各布斯:临床分子生物学研究所Christian-Albrechts大学、基尔和Popgen生物,大学医院石勒苏益格-荷尔斯泰因州;德国基尔;b·施密德:临床分子生物学研究所Christian-Albrechts大学、基尔,德国;m·维蒂希:临床分子生物学研究所Christian-Albrechts大学、基尔,德国;因特网:临床分子生物学研究所Christian-Albrechts大学、基尔,德国; K.I. Gaede: Dept of Pneumology, Research Center Borstel, Borstel, Germany; M. Schürmann: Institute of Human Genetics, University of Lübeck, Lübeck, Germany; M. Petrek: Laboratory of Immunogenomics and Immunoproteomics, Faculty of Medicine and Dentistry, Palacky University, Olomouc, Czech Republic; F. Mrazek: Laboratory of Immunogenomics and Immunoproteomics, Faculty of Medicine and Dentistry, Palacky University, Olomouc, Czech Republic; S. Pabst: Medical Clinic II, Dept of Pneumology, University of Bonn, Germany; C.Grohé: Dept of Respiratory Medicine, Evangelische Lungenklinik Berlin-Buch, Germany; J. Grunewald: Respiratory Medicine Unit, Dept of Medicine, Karolinska Institutet, Stockholm, Sweden; M. Ronninger: Dept of Medicine, Rheumatology Unit, Karolinska Institute, Stockholm, Sweden; A. Eklund: Respiratory Medicine Unit, Dept of Medicine, Karolinska Institutet, Stockholm, Sweden; P. Rosenstiel: Institute of Clinical Molecular Biology, Christian-Albrechts University, Kiel, Germany; K. Höhne: Dept of Pneumology, University of Freiburg, Freiburg, Germany; G. Zissel: Dept of Pneumology, University of Freiburg, Freiburg, Germany; J. Müller-Quernheim: Dept of Pneumology, University of Freiburg, Freiburg, Germany; S. Schreiber: Institute of Clinical Molecular Biology, Christian-Albrechts University, Kiel and Popgen Biobank, University Hospital Schleswig-Holstein; Kiel and Dept of General Internal Medicine, University Hospital Schleswig-Holstein, Kiel, Germany.
脚注
可以从本文的补充材料www.www.qdcxjkg.com
社论评论看778页。
支持声明
这项工作被授予的支持德国联邦教育和研究(BMBF)通过国家基因组研究网络(NGFN)的集群卓越”炎症在接口”和GenPhenReSa MU692/8-1,两者兼而有之德国研究基金会(DFG)的网络Diffus实质肺病(GOLD.net),在2010年_08深造大学IGA PU低频的一部分,由瑞典心肺基础,瑞典医学研究委员会,并通过区域医疗培训和临床研究协议(ALF)斯德哥尔摩郡议会和卡罗林斯卡医学院。
感兴趣的语句
没有宣布。
- 收到了2012年2月25日。
- 接受2012年7月19日。
- ©2013人队











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}