





摘要
肺癌,包括肺腺癌,是一种异质性疾病,由气道上皮分子改变演变而来。本研究探讨肺腺癌的一个亚型是否表达人气道基底细胞(british airway basal cells, BC)的独特分子特征,以及气道基底细胞特征的表达与肺腺癌的分子、病理和临床表型之间的关系。
分析了三个独立的肺腺癌数据集,以确定构成气道BC特征的基因的表达。肺腺癌中BC特征的表达与临床和生物学参数相关。
在肺腺癌中发现了气道BC特征基因的显著富集。一组肺腺癌(BC-高腺癌)表现出BC特征基因的高表达,与这些基因表达较低的腺癌相比,这些特征基因的表达与较差的肿瘤分级、较高的血管侵袭频率和较短的生存期相关。在分子水平上,BC-high腺癌显示出更高的频率喀斯特突变、与细胞周期、细胞外基质组织相关的转录网络和通路的激活,以及与纤毛和外分泌细支气管细胞(Clara细胞)相关基因抑制的明显分化模式。
气道BC程序的激活是一种独特的、侵袭性肺腺癌亚型的分子特征。
摘要
气道基底细胞基因的高表达与侵袭性肺腺癌的分子表型有关http://ow.ly/nIdJC
介绍
肺癌是全球癌症死亡率的主要原因,是一种异质性疾病,由环境致癌应激(主要是吸烟)介导的气道上皮分子改变演变而来[1- - - - - -3.]。人气道上皮是一种假层状细胞,主要由纤毛细胞、分泌细胞、中间细胞、基底细胞(bc)和罕见的神经内分泌细胞组成[4]。
这些单独的气道上皮细胞类型对肺癌异质性的具体贡献尚不清楚。气道bc,气道上皮的干/祖细胞群[5],被认为是肺鳞状细胞癌的候选“起源细胞”,部分原因是气道bc可能是鳞状细胞化生的来源,鳞状细胞化生是鳞状细胞癌相关的潜在肿瘤前病变[1]。相比之下,肺腺癌的细胞起源尚不清楚[1]。中心位置的腺癌被认为起源于支气管表面或腺上皮[6]。相比之下,外分泌细支气管细胞(Clara细胞)和II型肺细胞相关分化模式,也称为“终末呼吸单位”(TRU),已在周围腺癌中观察到[7,8]。外分泌细支气管细胞亚群,一种存在于小鼠气道上皮的分泌细胞类型[9],但仅限于人类的小气道[10],与小鼠肺腺癌的发展有关[11]。然而,其他细胞类型,如bc样祖细胞,对人类肺腺癌的贡献也被提出[1,12]。
在目前的研究中,使用我们最近对人类气道BC转录组的描述[13],我们分析了气道bc独特的分子特征对肺腺癌分子和临床表型的贡献。这些数据为肺腺癌亚型提供了证据,该亚型表达高水平的气道BC基因与侵袭性临床表型相关。
方法
所使用方法的更多细节可在在线补充材料中找到。
肺腺癌数据集
分析了三个独立的肺腺癌队列:一个主要队列(199名受试者中的182名最初由Chitale等[14],对其进行了组织学重新评估,并更新了最近的临床信息)和两个验证队列,其中一个由Bild等。[15](58名受试者),S海顿等。[16(442个科目中有327个,即。不包括在纪念斯隆-凯特琳癌症中心(MSKCC) (New York, NY, USA)分析的104例受试者,其中大多数存在于初级队列,以及11例基于病理重新评估确定的大细胞神经内分泌癌样本[17])。患者特征总结在在线补充表1中。补充图S1为本研究的实验流程示意图。
人肺腺癌气道BC特征表达分析
气道BC特征(862个基因)先前在我们的实验室中被表征[13]。为了分析基因富集,通过芯片对微阵列数据进行归一化,然后确定所有样本中所有基因的中位数表达水平。每个基因的中位数水平与所有862个气道BC特征基因的中位数水平进行比较,非BC特征基因在完整的大气道上皮中表达明显更高与基于全基因组微阵列比较纯化BC(高表达标准:fold-change≥5,benjamin - hochberg校正p<0.01)和50个随机862个基因集(使用Excel (Microsoft Corporation, Redmond, WA, USA) RAND函数从Affymetrix (Santa Clara, CA, USA) HG-U133A基因组中选择)。
比较不同亚型癌气道BC特征的表达[18- - - - - -24],使用genesspring version 7.3.1 (Agilent Technologies, Santa Clara, CA)对数据集进行主成分分析(PCA)。
A BC指数(I公元前)计算每个个体受试者的气道BC特征表达的累积测量,如前所述的完整气道上皮[25]。受试者的分类是基于I公元前使用四分位数法:底部四分位数内的个体被归类为“BC低”,顶部四分位数内的个体被归类为“BC高”。
为了确定bc -高和bc -低腺癌之间的转录组差异,我们进行了全基因组比较(差异表达基因的标准:fold-change bbb2,经Benjamini-Hochberg校正p<0.01)。利用DAVID Bioinformatics Resources 6.7分析工具(http://david.abcc.ncifcrf.gov/)。为了分析BC-高腺癌上调基因的网络,使用加权相关网络分析在上调基因中鉴定共表达基因,然后使用GNCPro分析工具(基于已知的物理蛋白质-蛋白质相互作用和转录调节,将鉴定的网络基因(标准:Spearman相关ρ>0.6, p<0.05)与BC-高肺腺癌中上调的气道BC特征基因联系起来(http://gncpro.sabiosciences.com/gncpro/gncpro.php)。
在原发性队列的肺腺癌亚型中,比较了与人气道上皮主要细胞类型(纤毛细胞、粘液分泌细胞、外分泌细支气管细胞和神经内分泌细胞)和上皮-间质转化(EMT)相关的基因表达。为了比较肺腺癌和鳞状细胞癌中气道BC特征的表达,该数据集包含58个腺癌和53个鳞状细胞癌ildet al。[15]进行了分析。
生存分析
为了评估气道BC信号表达与肺腺癌患者生存的关系,我们首先通过在总生存期<2年的腺癌患者(不良幸存者)之间进行全基因组比较,确定了不良生存相关基因。与原发性队列中5年总生存率为100的患者(差异表达基因标准:经Benjamini-Hochberg校正p<0.05)。所有生存分析均采用Kaplan-Meier法进行。使用log-rank检验比较腺癌亚型之间的生存。采用Cox比例风险模型进行多变量分析。
免疫组织化学分析
根据MSKCC机构审查委员会批准的方案和知情同意,从接受肺切除术的腺癌患者中独立采集活检样本。如前所述,基于TaqMan PCR分析(Applied Biosystems, Foster City, CA, USA),对BC-high腺癌(在所有三个独立的肺腺癌数据集中;基因列表见在线补充表II)。通过免疫组织化学分析验证bc -低和-高腺癌中所选蛋白的差异表达[13]。唯一的修改是用抗肿瘤蛋白63 (TP63)一抗(2 μg·mL)孵育−1;Santa Cruz Biotechnology, Santa Cruz, CA, USA)和甲状腺转录因子-1 (TTF-1) (3 μg·mL)−1;DAKO, Carpinteria, CA, USA)在37°C下放置2小时。市售的正常肺和肺鳞状细胞癌组织样本(US Biomax Inc., Rockville, MD, USA)用于比较分析。
统计分析
除芯片数据外,所有分析均使用SPSS统计软件包(SPSS Inc ., Chicago, IL, USA)进行。I之间的关系BC -和NKX2-1使用Pearson相关分析分析原发性腺癌队列中的基因表达。使用卡方检验或曼-惠特尼检验评估两组之间的关系。使用genesspring version 7.3.1对芯片数据进行分析。
结果
肺腺癌中气道BC信号丰富
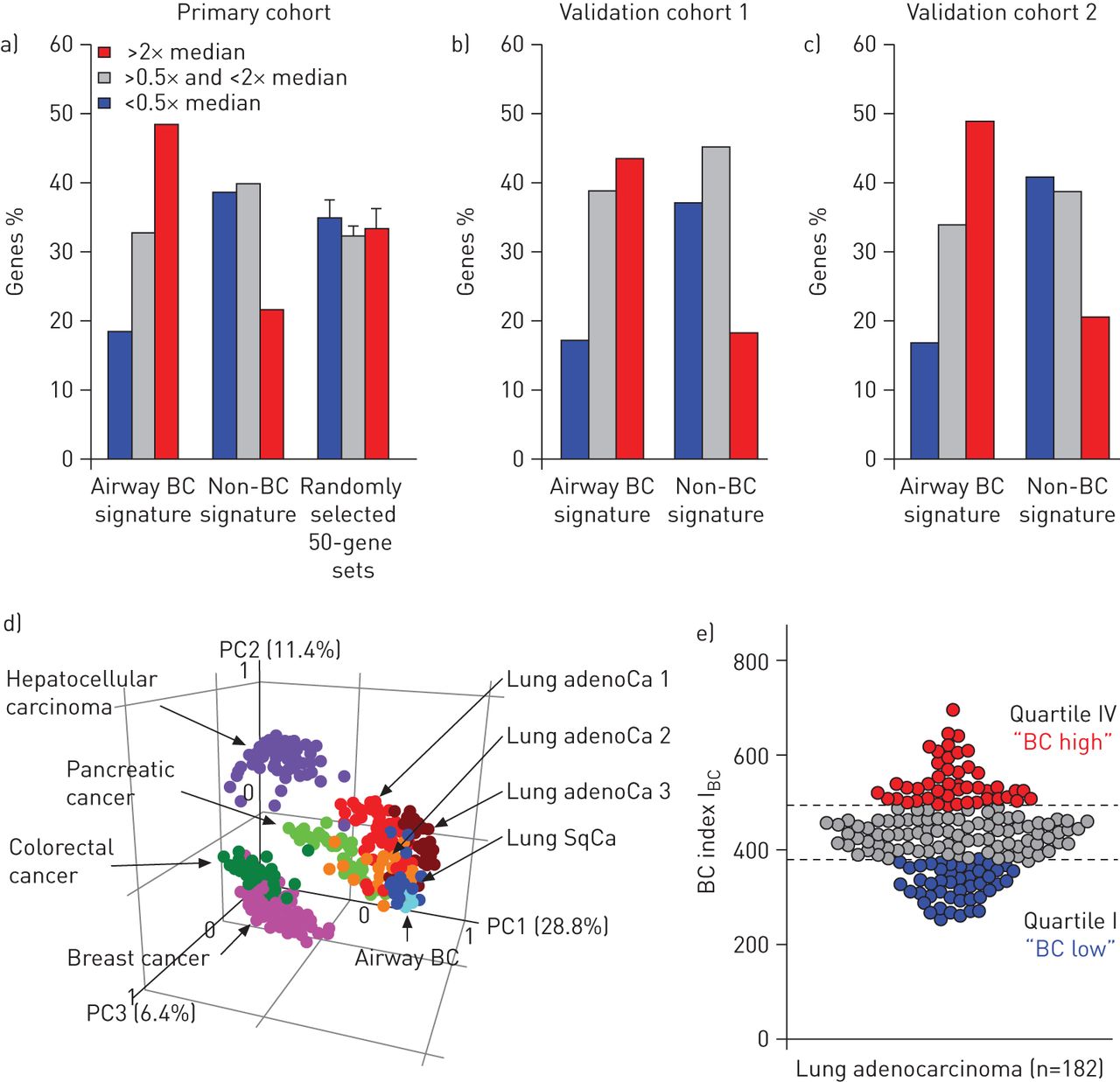
为了全面了解气道BC在肺腺癌中的表达分子特征,我们分析了862个气道BC特征基因(在线补充基因表I)的表达情况。在862个气道BC特征基因中,420个(48.7%)属于高表达的肺腺癌基因(表达水平超过所有基因中位数的两倍),而544个非BC特征基因(在线补充基因列表II)中只有118个(22%)和随机选择的基因集中<35%的基因属于高表达的肺腺癌基因(图1一个)。相比之下,在肺腺癌中,低于20%的bc特征基因导致了低表达的基因(表达水平低于所有基因中位数的一半),而非bc基因和随机选择的数据集中,这一比例为约35%。BC标记基因在肺腺癌中的富集通过两个独立队列(图1b和c)。对所有三个队列的综合分析显示,在高表达的肺腺癌基因中,气道BC特征基因的富集具有统计学意义与非bc基因(p<0.0006)与随机选择的基因集(p<0.02)(在线补充表III)。
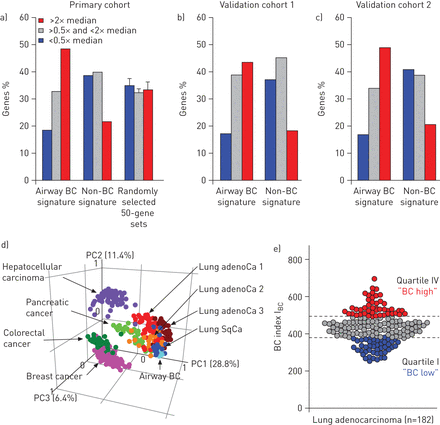
人肺腺癌(adenoCa)中气道基底细胞(BC)特征基因的表达。气道BC特征基因、非BC特征基因和50个随机862个基因组中导致肺腺癌高表达基因的频率(表达水平是所有表达基因中位数的两倍以上);红色),肺腺癌中低表达的基因(表达水平低于所有表达基因中位数的一半;蓝色)和在肺腺癌中中等表达的基因(其余肺腺癌表达基因;a)初级队列[14],验证队列b) 1 [15]及c) 2 [16]。详见在线补充表三。d)基于气道BC特征表达的主成分分析比较不同类型的人类癌和气道BC。分析的数据集包括肺腺癌1荷兰国际集团(ing)等。[22)(红色;n = 68);肺腺癌2来自H欧等。[21[深红色;n=40),腺oc3来自K味道等。[20.)(橙色;n=40),肺鳞状细胞癌(SqCa)味道等。[20.[深蓝色;n=18),结直肠癌来自Smith等。[18墨绿色;n=55),乳腺癌来自Lu等。[19)(粉色;n=129), C型肝细胞癌hiang等。[23)(紫色;n=91),胰腺癌来自Badea等。[24[浅绿色;n=39)和来自健康非吸烟者的气道BC样本(浅蓝色;n = 4)。每个圆圈代表一个单独的样本。指出了前三个主成分对观测到的变率的贡献百分比。e)肺腺癌分为高、低气道BC基因表达者。BC指数(I公元前)是基于在肺腺癌受试者(n=182)中表达高于中位数水平的气道BC特征基因的数量[14]。腺癌受试者分为bc -高(四分位IV;红色),bc -中间(四分位数II-III;灰色)和BC-low(四分位数I;蓝色)亚型。
在肺腺癌的一个亚群中,气道BC信号上调
接下来,我们询问肺腺癌中气道BC特征表达模式是否与其他癌症相同,还是相对独特于这种类型的肺癌。PCA显示肺鳞状细胞癌[20.]和所有三个肺腺癌数据集[20.- - - - - -22]表现出类似的模式,其聚类更接近气道BC样本而不是结肠直肠样本[18],乳房[19],肝细胞[23]和胰腺[24癌症(图1 d)。大多数肺鳞状细胞癌样本显示与气道BC基因表达模式相似,而肺腺癌则更具异质性。
为了进一步探讨肺腺癌在气道BC特征表达上的异质性公元前作为一个累积的基因表达参数。与上述PCA数据一致,基于气道BC特征表达的分析显示肺腺癌患者具有显著的异质性(图1 e)。基于I公元前bc -高(上四分位数)和bc -低(下四分位数)腺癌亚型被确定(图1 e)。
bc -高肺腺癌具有独特的生物学表型
为了确定BC-high腺癌中富集的生物学途径和模式,我们首先进行了BC-high的全基因组比较与BC-low腺癌(图2一个和在线补充基因表III)。在bc -高腺癌中上调的364个基因中,与细胞周期、细胞外基质(ECM) -受体相互作用和p53信号通路相关的生物通路(图2 b)。与通路分析一致,bc -高腺癌上调基因的网络分析显示,与ECM组织相关的转录网络元件富集(图2 c)。BC-高腺癌富集的共表达ECM网络组分是调节上皮-间质相互作用和肺组织稳态的BC特征基因的相互作用伙伴,包括转化生长因子-β1 (TGFB1)、金属蛋白酶(MMP)1和MMP2、金属蛋白酶组织抑制剂(TIMP)1、整合素-αV(ITGAV)及维生素D受体(VDR) (图2 c)。
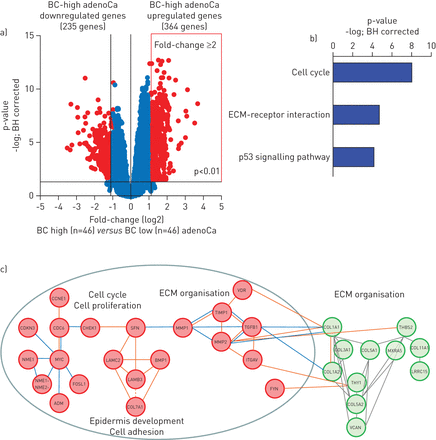
基底细胞(BC)高肺腺癌(adenoCa)与基底细胞(BC)低肺腺癌(adenoCa)基因的差异表达a)在原发性肺腺癌队列(n=182)中,bc -高腺癌(n=46)和bc -低腺癌(n=46)的全基因组比较火山图。红色:显著基因(折叠变化≥2;benjamin - hochberg (BH)校正p<0.01);蓝色:组间无显著差异的基因。b)京都基因百科和基因组通路分析显示,BC-high腺癌上调基因显著富集(经BH校正p<0.05)。c)通过加权相关网络分析鉴定的BC-高腺癌上调基因中富集的分子网络(绿色:在主要数据集中共表达BC-高肺腺癌基因),并使用GCNPro分析工具连接到BC-高腺癌上调的BC特征基因(红色)(橙色:转录调控;蓝色:物理蛋白质-蛋白质相互作用;详情请参见方法部分)。ECM:细胞外基质; VDR: vitamin D receptor; CCN: cyclin E1; TIMP1: tissue inhibitor of metalloproteases 1; CDKN3: cyclin-dependent kinase inhibitor 3; CDC6: cell division cycle 6; CHEK1: checkpoint kinase 1; SFN: stratifin; MMP1: matrix metalloprotease 1; TGFB1: transforming growth factor-β1; COL1A1: collagen Iα1; THBS2: thrombospondin 2; MMP2: matrix metalloprotease 2; COL3A1: collagen IIIα1; COL5A1: collagen Vα1; MXRA5: matrix remodelling-associated 5; COL11A1: collagen XIα1; NME1: NME/NM23 nucleoside diphosphate kinase 1; LAMC2: laminin-γ2; BMP1: bone morphogenetic protein 1; ITGAV: integrin-αV;COL1A2:胶原Iα2;LRRC15:富含亮氨酸的重复序列15;LAMB3:层粘连蛋白-β3;NME1-NME2:NME1- - - - - -NME2基因readthrough;FOSL1: fos样抗原1;COL5A2:胶原Vα2;ADM: adrenomedullin;COL7A1:胶原viii α1;VCAN: versican。
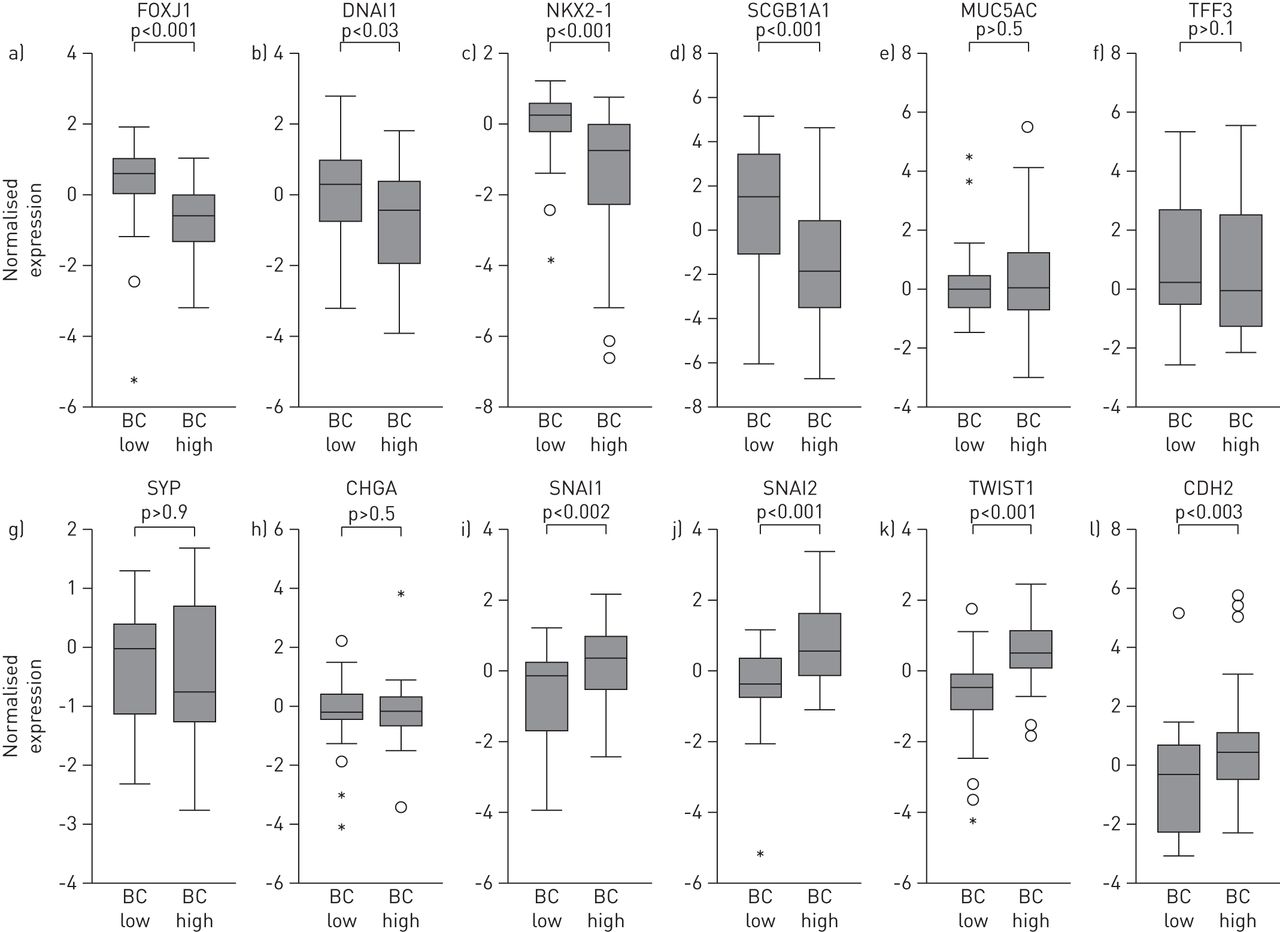
BC-high腺癌显示与小气道上皮主要细胞类型分化相关的基因显著下调,包括纤毛细胞(forkhead box J1 (FOXJ1)和动力蛋白轴突中间链1 (DNAI1))和外分泌细支气管细胞(NK2同源盒1 (NKX2-1)和分泌珠蛋白1A1 (SCGB1A))。粘液分泌细胞和神经内分泌细胞典型基因的表达在这两种亚型之间没有差异。I和I之间呈负相关公元前和NKX2-1基因表达(在线补充图S2)。与这一观察结果相一致的是,有一种较低表达的趋势NKX2-1与TTF-1阴性的鳞状细胞癌不同,它在两种腺癌亚型中均可检测到(在线补充图S3)。与分化基因相比,与EMT相关的基因,如SNAI1(蜗牛同源物1);SNAI2(蜗牛同源物2);TWIST1(twist basic helix-loop-helix转录因子1)和CDH2(N-cadherin) [26],与bc -低相比,bc -高的腺癌(图3)。与其他肺癌亚型相关的基因,包括小细胞肺癌,如编码p53、视网膜母细胞瘤-1和L-MYC的基因,在bc -高和bc -低腺癌亚型之间没有差异表达(在线补充图S4)。
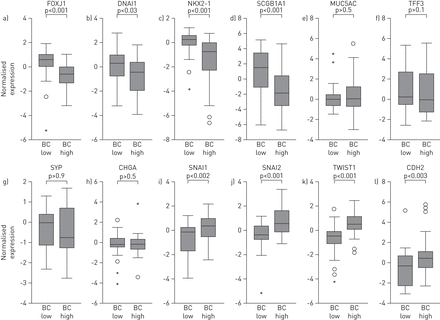
基底细胞(BC)高水平腺癌与基底细胞(BC)低水平腺癌中分化相关分子模式表达的比较纤毛细胞基因:叉头盒J1 (FOXJ1)和动力蛋白轴突中间链1 (DNAI1);外分泌细支气管细胞(Clara细胞)基因:NK2同源盒1 (NKX2-1)和分泌珠蛋白1A1 (SCGB1A1);产生黏液分泌细胞相关基因:黏液蛋白5AC (MUC5AC)及三叶因子3 (TFF3);神经内分泌细胞基因:突触素(SYP)及嗜铬粒蛋白A (CHGA);与上皮-间质转化相关的基因:蜗牛同源物1 (SNAI1)、蜗牛同源物2 (SNAI2),捻同系物1 (TWIST1)和n -钙粘蛋白(CDH2)。在所有的面板,原木2-基于微阵列分析的转化规范化基因表达水平显示;每组N =46。根据四分位数间距(IQR)表示异常值:圆圈,-1.5×IQR至3×IQR;星星:>3或<3×IQR。
BC-high腺癌具有侵袭性的临床表型
在BC-高水平腺癌中,细胞周期相关基因和EMT基因的上调以及分化相关基因表达程序的抑制表明,BC特征在肺腺癌中的高表达可能与更具侵袭性的肿瘤表型相关。分析显示,在139个与肺腺癌(图4),约20%的基因是BC标记基因(图4 b在线补充基因表IV)。
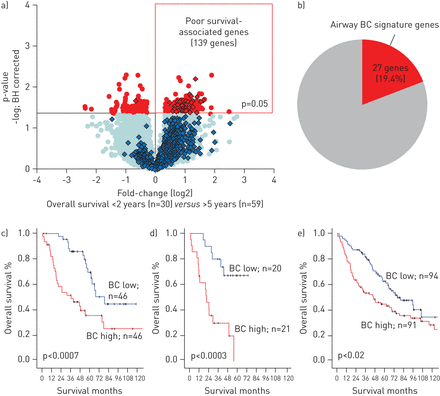
气道基底细胞(BC)特征表达与肺腺癌患者生存的关系。a)总生存期<2年的腺癌患者(生存差患者)的全基因组比较;在原发性肺腺癌队列(N =182)中,N =30)和5年总生存率为>的患者(N =59)。红色:显著基因(经Benjamini-Hochberg (BH)校正p<0.05);蓝色:组间无显著差异的基因;钻石:862气道BC特征基因。b) 139个生存不良相关基因(如上所述)与气道BC特征基因重叠。c-e)基于Kaplan-Meier分析的bc -高腺癌患者总生存率(红色)与bc -低腺癌患者(蓝色)来自c的主要队列hitaleet al。[14],验证队列为d) Bildet al。[15d)选d海顿et al。[16]。p值由log-rank检验确定;每一组的人数都被标示出来。
与bc -低腺癌相比,bc -高腺癌的特征是分化较差(p<0.001),具有鳞状模式的预后有利腺癌的频率较低(以前为支气管肺泡癌;P <0.001),血管侵犯频率较高(P <0.004)。在分子水平上,BC-high腺癌表现出明显更高的频率喀斯特突变和频率较低表皮生长因子受体(表皮生长因子受体)突变符合喀斯特突变状态,这被认为是吸烟相关腺癌的特征[1,3.],与bc -低的腺癌患者相比,bc -高的腺癌患者吸烟者明显更多(表1)。
与BC标记对生存相关基因组的显著贡献一致(图4 b), bc含量高的腺癌患者的总生存期比bc含量低的腺癌患者短(中位生存期为36与79个月;Log rank p<0.0007) (图4摄氏度)。多变量生存分析表明,气道BC特征的高表达是与较短生存期相关的独立预后因素(风险比1.59,95% CI 1.14-2.22;p < 0.008) (表2)。
通过两个独立的肺腺癌队列验证了气道BC特征表达的预后相关性[15,16]。bc -高腺癌病例的比例在36%至28%之间,而在初级队列中为25%。与初始队列相似,bc -高的腺癌患者的总生存期明显短于bc -低的腺癌患者(图4d和e)。与bc含量低的腺癌相比,bc含量高的腺癌也与更短的无病生存期相关(补充图5)。
bc -高肺腺癌不同于肺鳞状细胞癌
一些气道BC基因的异常激活先前与肺鳞状细胞癌有关[12]。与这些先前的观察结果一致,与腺癌队列相比,鳞状细胞癌中气道BC特征的总体表达明显更高(图5)。然而,BC-高和BC-低的鳞状细胞癌个体的总体生存期没有显著差异(在线补充图S6a和b)。值得注意的是,尽管与腺癌相比,BC基因在鳞状细胞癌中的总体表达更高,但在所分析的队列中,鳞状细胞癌患者的总体生存期比BC-高的腺癌患者更长(在线补充图S6c)。
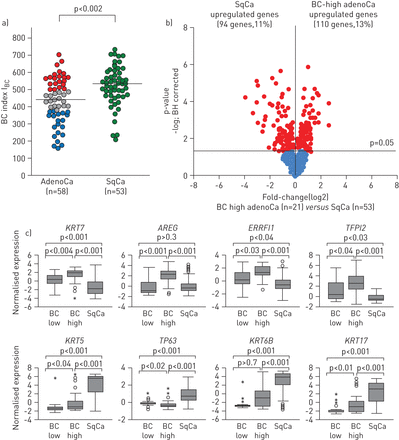
肺腺癌(adenoCa)和肺鳞状细胞癌(SqCa)气道基底细胞(BC)特征表达的比较分析。a) B区肺腺癌(58例)和SqCa(53例)ildet al。[15]数据集进行分析。BC指数(I公元前)根据每个基因的腺癌受试者的中位数水平计算。红色:bc -高腺癌;灰色:bc -中级腺癌;蓝色:bc -低腺癌;格林:SqCa。中值I公元前两种类型的癌症(腺癌436例,SqCa 529例)用水平线突出显示;p值由Mann-Whitney检验确定。b)比较气道BC特征基因表达的火山图[13[B]数据集中bc -高肺腺癌(n=21)到SqCa (n=53)的差异ild等。[15]。红色:显著基因(经Benjamini-Hochberg (BH)校正p<0.05);蓝色:组间无显著差异的基因。c)气道BC特征基因样本在BC高含量腺癌中差异表达(n=21)与从B的数据集中取SqCa (n=53)ildet al。[15]。在所有的面板,原木2-转化的规范化基因表达水平是基于微阵列分析。根据四分位间距(IQR)表示异常值:圆圈:−1.5×IQR至3×IQR;星星:>3或<3×IQR。KRT7: keratin-7;沙土荒漠: amphiregulin;ERRFI1: ErbB受体反馈抑制剂1;TFPI2:组织因子途径抑制剂2;KRT5: keratin-5;TP63:肿瘤蛋白63;KRT6B: keratin-6B;KRT17: keratin-17。
接下来,我们询问bc -高腺癌是否与鳞状细胞癌具有气道bc相关的分子特征。BC-高水平腺癌和鳞状细胞癌中气道BC特征基因表达的比较发现,13%的气道BC基因在BC-高水平腺癌中表达较高,11%在鳞状细胞癌中表达较高(图5 b和在线补充基因表V)。这表明BC-高腺癌的特征是气道BC基因的独特模式,将这种亚型的肺癌与鳞状细胞癌区分开来。在BC含量高的腺癌中,主要上调的气道BC基因包括角蛋白-7 (KRT7)、EGFR配体双调节蛋白(沙土荒漠)、ErbB受体反馈抑制剂1 (ERRFI1)及组织因子通路抑制因子2 (TFFPI2) (图5度)。相比之下,经典的BC标记物角蛋白-5 (KRT5),TP63,角蛋白5b (KRT6B)及角蛋白17 (KRT17)在鳞状细胞癌中的表达明显高于bc -高腺癌(图5度)。与这一观察结果一致,免疫组织化学分析显示,TP63蛋白通常在气道BC人群中表达,但在鳞状细胞癌中过度表达,而在两种腺癌亚型中均未表达(在线补充图S3)。
讨论
越来越多的证据表明,人类恶性肿瘤的生物学异质性是由特定的组织驻留细胞群决定的,这些细胞在一组不同的致癌改变的影响下,在每种癌症的个别组织学类型中产生特定的临床相关表型[28]。基于这一概念,我们利用我们最近对人气道bc(气道上皮的干/祖细胞群)的转录组特征评估了人肺腺癌的分子和临床异质性[5,13]。这项分析使我们确定了一种新的肺腺癌生物学亚型,称为BC-高腺癌,其特征是与肿瘤侵袭性的临床和病理特征相关的一组独特的气道BC特征基因上调。
肺癌起源于气道上皮的分子改变[1,3.,8],由纤毛细胞、中间细胞和分泌细胞(大气道中的杯状细胞和小气道中的外分泌细支气管细胞)、bc细胞和罕见的神经内分泌细胞组成的细胞群[4]。根据单个肺癌亚型的独特形态学特征,已经提出了每种组织学亚型起源的候选细胞类型。小细胞肺癌和大细胞神经内分泌癌被认为起源于神经内分泌细胞[1]。气道bc被认为是肺鳞状细胞癌的推定起源细胞,因为气道bc是鳞状化生的来源,鳞状化生是一种与肺鳞状细胞癌发展的早期阶段相关的组织学病变[1],以及某些BC标记物如KRT5和TP63在肺鳞状细胞癌中的过表达[29]。对于肺腺癌,起源细胞尚不清楚,尽管外分泌细支气管细胞和II型肺细胞被认为是周围腺癌的细胞起源,而表面和腺支气管上皮细胞是更多近端腺癌的来源[6]。在小鼠中,“细支气管肺泡干细胞”是一种独特的干细胞群,位于细支气管肺泡管交界处,具有外分泌细支气管和肺泡II型细胞的特征,与肺腺癌的发生和增殖有关[11]。然而,人气道上皮的细胞组成与小鼠不同。在人类中,bc存在于整个气道中,而在小鼠的小气道中几乎不存在[30.],阻碍了利用小鼠模型研究气道bc在肺腺癌发展中的作用。
在本研究中,我们通过假设肺腺癌亚型可能来源于气道bc,在转录水平上评估了肺腺癌的生物学异质性。基于气道BC特征基因的表达,数据表明肺腺癌可分为BC高亚型和BC低亚型,它们具有显著不同的生物学、病理和临床特征。这些数据通过证明人类肺腺癌的表型多样性可以(至少部分地)通过与气道bc相关的基因表达程序的或多或少的持续激活来解释,从而为肺腺癌的生物学提供了见解。
与BC-high相关的分子模式与BC-低腺癌也提供了可能导致肺腺癌子集中气道BC程序激活的机制的见解。首先,有更高的频率喀斯特BC-high腺癌的突变。的联系喀斯特干细胞/祖细胞突变已被报道与肺癌的发展有关。内胚层祖细胞喀斯特突变表现为增殖增加,不能分化和维持干细胞特征在体外[31]。这些数据支持bc -高腺癌可能源于气道bc携带的概念喀斯特与吸烟有关的突变[1,3.]。相比之下,BC-low腺癌的特征是更高频率的表皮生长因子受体突变。这与之前的观察一致表皮生长因子受体突变在非吸烟腺癌患者中更为常见[3.]并与被称为TRU的分化模式有关,TRU与外分泌细支气管和肺泡II型细胞典型基因的高表达有关,如编码TTF-1和表面活性蛋白的基因[7]。
其次,BC-高含量肺腺癌富含与ECM组织相关的转录通路和网络,这些通路和网络与各种BC特征基因相互作用,这些基因编码肺组织中稳态过程的重要调节因子,包括TGFB1,MMP1,MMP2,TIMP2(组织金属蛋白酶抑制剂2),ITGAV和论述以及与表皮发育、细胞粘附、细胞周期和增殖相关的网络。鉴于淋巴球位于基底膜上方的解剖位置,淋巴球可能通过调节ECM稳态和上皮-间质相互作用参与肺腺癌的发病机制。激活不同的生长因子信号机制,包括那些与TGFB1和/或介导的通过基质金属蛋白酶和整合素信号在上皮-间质界面的激活,可能负责细胞粘附和细胞周期相关网络的富集,并有助于在bc -高肺腺癌中观察到更具侵袭性的肿瘤特征。
第三,BC-high分化格局的比较分析与bc -低肺腺癌发现,在bc -高肺腺癌中,纤毛和外分泌细支气管细胞分化相关基因的下调伴随着EMT转录程序的激活,包括转录因子的诱导,如SNAI1/SNAIL、SNAI2/SLUG、TWIST1和CDH2 [qh]26]。EMT程序的激活被认为是促进癌症侵袭和转移的重要过程[32]。与这一概念相一致,BC-high腺癌表现出更高频率的血管侵袭和淋巴结转移。此外,有报道称EMT可产生具有致瘤特性的细胞[33]。本研究加强了EMT与肺腺癌发展背景下组织干细胞之间的关系。
最后,与BC基因也高表达的鳞状细胞癌相比,我们发现BC高含量的肺腺癌表现出一组不同的BC特征基因上调,包括与EGFR通路相关的基因,如沙土荒漠和ERRFI1。EGFR通路在气道bc中富集[13]和吸烟可以激活肺上皮细胞中没有EGFR突变的EGFR信号,从而诱导与肺癌发展相关的细胞过程[34]。
虽然导致bc -高腺癌富集分子通路的基因并不被认为是经典的致癌基因,但了解它们之间的相互作用对于开发旨在调节肿瘤细胞存活和生长的新治疗策略非常重要,例如,使用合成致死性方法。这种策略可能对bc -高肺腺癌特别有益,因为bc -高肺腺癌与高频率的喀斯特尚未开发出有效的特异性靶向方法的突变[3.]。靶向一些bc -高腺癌富集基因之间的相互作用,如癌基因MYC和周期蛋白依赖激酶,最近被证明在侵袭性bc样乳腺癌中诱导治疗相关的合成致死性[35]。此外,基于bc的生存、生长和分化取决于它们对ECM成分的粘附这一认识[36]和ECM相关基因在bc -高含量腺癌中富集,那么靶向ECM基因可能是在bc -高含量腺癌中诱导合成致死性相互作用的另一种方法。
总之,本研究确定了一种新的BC-高人类肺腺癌亚型,与一组独特的气道BC特征基因的激活有关,并提供了基于转录组的证据,支持这一侵袭性人类肺腺癌亚群可能来自气道BC人群的概念。
致谢
我们感谢M. Ladanyi (Memorial Sloan-Kettering Cancer Center, New York, NY, USA)提供MSKCC腺癌样本,J. Salit (Weill Cornell Medical College, New York, NY, USA)支持微阵列分析,N. Mohamed (Weill Cornell Medical College)帮助准备本文。
脚注
本文的补充资料来自www.www.qdcxjkg.com
支持声明:这些研究在一定程度上得到了美国国立卫生研究院授予P50 HL084936、T32 HL094284、UL1 RR02499斯塔尔基金会/斯塔尔癌症协会。R. Shaykhiev在一定程度上得到了帕克B.弗朗西斯基金会。
利益冲突:没有声明。
- 收到了2012年9月11日。
- 接受2013年1月21日。
- ©2013人队











![Expression of the airway basal cell (BC) signature genes in human lung adenocarcinoma (adenoCa). Frequency of the airway BC signature genes, non-BC signature genes and the genes of the 50 random 862-gene sets contributing to the genes with high expression in lung adenoCa (expression level more than twice the median for all expressed genes; red), genes with low expression in lung adenoCa (expression level less than half the median for all expressed genes; blue) and genes with intermediate expression in lung adenoCa (remaining lung adenoCa-expressed genes; grey) in the a) primary cohort [14], and validation cohorts b) 1 [15] and c) 2 [16]. See online supplementary table III for details. d) Principal component analysis comparing various types of human carcinomas and airway BCs based on expression of the airway BC signature. Analysed data sets include lung adenoCa 1 from Ding et al. [22] (red; n=68); lung adenoCa 2 from Hou et al. [21] (dark red; n=40), adenoCa 3 from Kuner et al. [20] (orange; n=40), lung squamous cell carcinoma (SqCa) from Kuner et al. [20] (dark blue; n=18), colorectal cancer from Smith et al. [18] (dark green; n=55), breast cancer from Lu et al. [19] (pink; n=129), hepatocellular carcinoma from Chiang et al. [23] (purple; n=91), pancreatic cancer from Badea et al. [24] (light green; n=39) and airway BC samples from healthy nonsmokers (light blue; n=4). Each circle represents an individual sample. The per cent contributions of the first three principal components (PCs) to the observed variability are indicated. e) Categorisation of lung adenoCa into high and low airway BC gene expressers. The BC index (IBC) is based on a number of airway BC signature genes expressed above the median level in lung adenoCa subjects (n=182) [14]. AdenoCa subjects were divided into BC-high (quartile IV; red), BC-intermediate (quartiles II–III; grey) and BC-low (quartile I; blue) subtypes.](http://www.qdcxjkg.com/content/erj/42/5/1332/F1.large.jpg?width=800&height=600&carousel=1){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
![Relationship between airway basal cell (BC) signature expression and lung adenocarcinoma patient survival. a) Genome-wide comparison between adenocarcinoma patients with <2-year overall survival (poor survivors; n=30) and those with >5-year overall survival (n=59) in primary lung adenocarcinoma cohort (n=182). Red: significant genes (p<0.05 with Benjamini–Hochberg (BH) correction); blue: genes with no significant difference between the groups; diamonds: 862 airway BC signature genes. b) Overlap between the 139 poor survival-associated genes (identified as described above) and airway BC signature genes. c–e) Kaplan–Meier analysis-based estimates of overall survival of BC-high adenocarcinoma patients (red) versus BC-low adenocarcinoma patients (blue) from c) the primary cohort of Chitale et al. [14], and the validation cohorts of d) Bild et al. [15] and e) Shedden et al. [16]. P-values were determined by the log-rank test; the number of individuals in each group is indicated.](http://www.qdcxjkg.com/content/erj/42/5/1332/F4.large.jpg?width=800&height=600&carousel=1){kind=link}
{kind=link}
![Comparative analysis of the airway basal cell (BC) signature expression in lung adenocarcinoma (adenoCa) and lung squamous cell carcinoma (SqCa). a) Lung adenoCa (n=58) and SqCa (n=53) cases from the Bild et al. [15] data set were analysed. The BC index (IBC) was calculated based on median levels of adenoCa subjects for each gene. Red: BC-high adenoCa; grey: BC-intermediate adenoCa; blue: BC-low adenoCa; green: SqCa. The median IBC for both types of cancer (436 for adenoCa and 529 for SqCa) are highlighted with horizontal lines; p-value indicated was determined by Mann–Whitney test. b) Volcano plot comparing expression of the airway BC signature genes [13] in BC-high lung adenoCa (n=21) to SqCa (n=53) from the dataset of Bild et al. [15]. Red: significant genes (p<0.05 with Benjamini–Hochberg (BH) correction); blue: genes with no significant difference between the groups. c) Airway BC signature gene examples differentially expressed in BC-high adenoCa (n=21) versus SqCa (n=53) from the data set of Bild et al. [15]. In all panels, log2-transformed normalised gene expression levels are based on the microarray analysis. Outliers were indicated on the basis of interquartile range (IQR): circles: −1.5×IQR to 3×IQR; stars: >3 or <3×IQR. KRT7: keratin-7; AREG: amphiregulin; ERRFI1: ErbB receptor feedback inhibitor 1; TFPI2: tissue factor pathway inhibitor 2; KRT5: keratin-5; TP63: tumour protein 63; KRT6B: keratin-6B; KRT17: keratin-17.](http://www.qdcxjkg.com/content/erj/42/5/1332/F5.large.jpg?width=800&height=600&carousel=1){kind=link}
{kind=link}