





摘要
几丁质酶样蛋白YKL-40介导气道炎症,血清水平与哮喘严重程度相关。然而,与YKL-40水平相关的哮喘表型尚未被精确定义。
我们对在耶鲁哮喘和气道疾病中心治疗的哮喘患者(n=156)进行了无监督聚类分析,根据YKL-40水平确定亚组。得到的YKL-40聚类在严重哮喘研究项目(n=167)和纽约大学/贝尔维尤哮喘库(n=341)的队列中进行交叉验证。痰转录组分析发现了与YKL-40亚群相关的分子途径。
鉴定出4个YKL-40簇(C1-C4)。C3和C4血清YKL-40水平高于C1和C2。C3与发病早、病程长、严重气流阻塞和接近致命的哮喘加重相关。C4患者血清YKL-40水平最高,成年发病,气流阻塞较少,但频繁加重。C3和C4的气道转录组分析显示非2型炎症通路激活。
血清YKL-40水平升高与两种不同的临床哮喘表型相关:一种是不可逆气道阻塞,另一种是严重加重。YKL-40基因簇对鉴别患有严重或易加重哮喘的个体有潜在的帮助。
摘要
血清YKL-40水平高的哮喘与严重肺功能损害和严重加重有关http://ow.ly/wyly30elajo
简介
在患有哮喘的受试者中,血清中几丁质酶样蛋白YKL-40的水平较高,这种蛋白是由CHI3L1基因与严重哮喘、肺功能损害和疾病加重有关[1].然而,更精确地定义与YKL-40相关的表型的无监督学习模型尚未得到测试[2].
我们之前已经证明,哮喘风险、疾病严重程度和血清YKL-40高水平之间的关系部分是由单核苷酸多态性(SNP)的遗传效应介导的CHI3L1基因启动子和内含子SNP [3.,4].虽然人类和机制研究已经证明YKL-40参与气道重塑[5,6],他们没有证明2型(T2)炎症标志物和YKL-40之间的明确关联[1,2,7].这提示YKL-40可能影响或受哮喘非t2炎症反应的影响。
近年来,基于疾病的临床和分子特征,聚类分析已成功用于识别具有相似哮喘特征的患者亚组[8- - - - - -11].在严重哮喘研究计划(SARP), Mooreet al。[9]使用哮喘的11个临床和生理特征来确定5组具有明显临床特征的患者。这些特征已在独立队列中得到重复,表明这些临床和生理特征是哮喘异质性的重要鉴别器[12,13].
由于已知YKL-40与严重哮喘相关,且有证据表明YKL-40与非t2炎症通路之间存在联系,我们试图使用无监督聚类方法来了解疾病的临床和生理特征,以及存在于血清YKL-40高水平哮喘受试者气道中的独特分子机制。这些实验的目的有两个。首先,我们在无监督聚类分析中测试了是否将血清YKL-40水平添加到之前验证的哮喘严重程度的临床特征中[9]将导致确定以YKL-40表达增加为特征的特定临床亚组。其次,我们试图确定痰中与YKL-40集群相关的特定基因表达谱。
方法
主题
位于美国康涅狄格州纽黑文的耶鲁大学哮喘和呼吸道疾病中心(YCAAD)队列研究参与者完成了全面的表型研究访问。纳入和排除标准以及YCAAD表型分型协议已在前面描述过[1].补充材料中包含了对这些方法的附加说明。SARP和纽约大学/贝尔维尤哮喘库(NYUBAR)队列的研究参与者使用既定的标准操作程序完成了研究访问,如前所述[9,14].这些主题的特点已在以往的刊物中报道过[9,12,14,15].这些研究是在机构审查委员会批准所有相关机构后进行的,所有参与者都提供了知情同意。
痰诱导和基因表达测定
痰诱导仅在YCAAD队列中进行;补充材料中描述了其他细节。
统计分析
所有的统计、聚类和分类器分析都使用R软件(R统计计算基金会,维也纳,奥地利)进行。除非另有说明,报告结果为中位数和四分位间范围(25-75%)。连续变量采用非参数检验,包括两组比较用Wilcoxon检验,两组以上比较用Kruskal-Wallis检验。分类变量用卡方检验进行分析。p值<0.05被认为显著。关于聚类和基因表达分析的具体细节在补充材料中描述。
结果
YKL-40簇的发现和验证
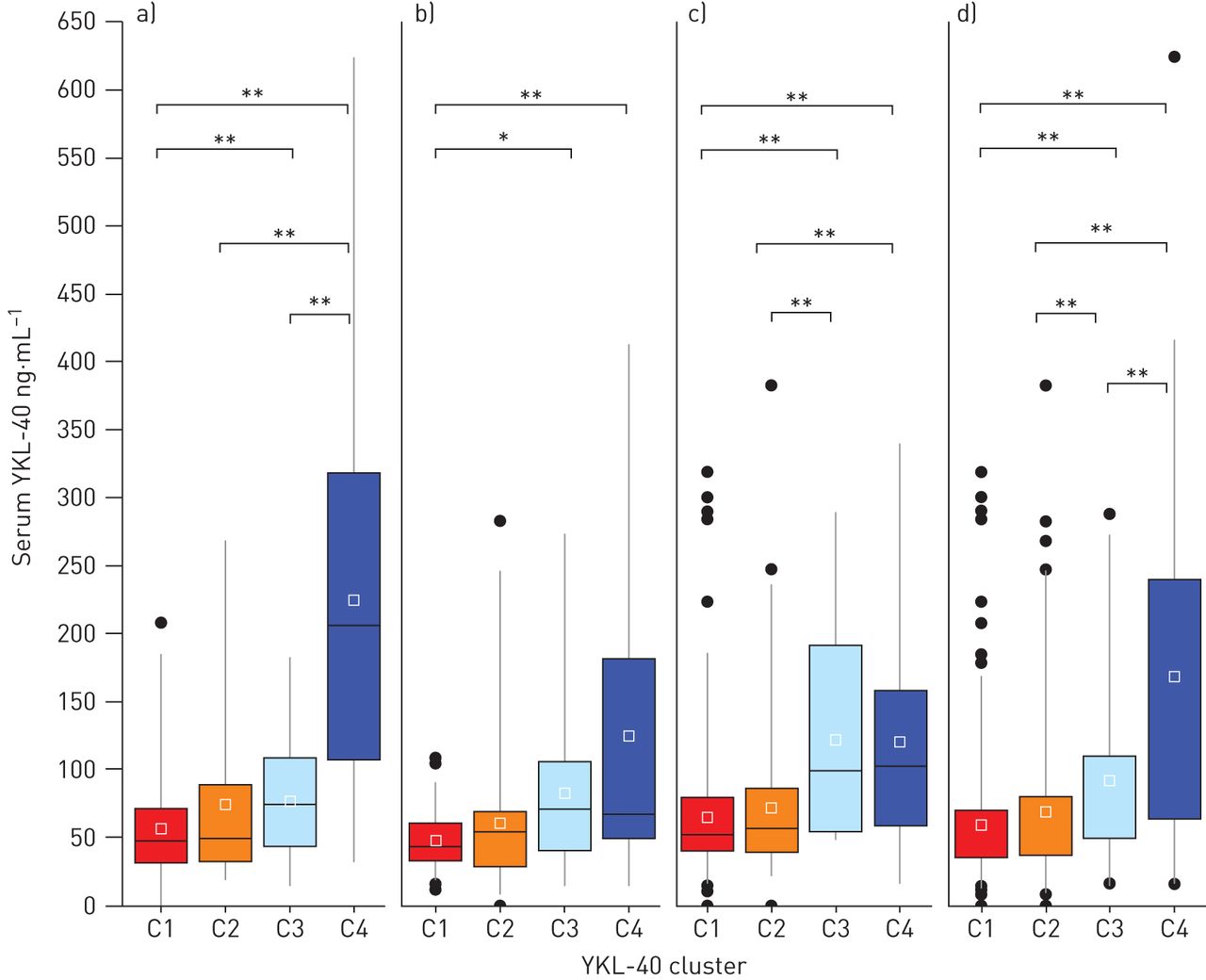
为了确定与血清YKL-40水平升高相关的哮喘患者亚组,我们对来自YCAAD队列的156名个体进行了无监督聚类分析,使用血清YKL-40水平和哮喘的11个临床和生理特征,如方法[9].图1说明了研究工作流程。4个亚组的疾病患者具有不同的血清YKL-40水平和不同的临床和生理特征的疾病(群集C1-C4)被确定(图2一个).C1的YKL-40中位值最低(48 (32-71)ng·mL−1),与C2的中位数(50 (33-88)ng·mL相近−1).这些中值也类似于YKL-40值(43 (20-184)ng·mL)−1)在健康的受试者中[16].血清YKL-40簇C3中位YKL-40水平高于C1和C2的中位YKL-40水平(74 (6-108))与49 (32 - 74) ng·毫升−1;p = 0.05)。C4的中位水平也高于C1和C2的总和(206 (110-311))与49 (32 - 74) ng·毫升−1;p < 0.01)。C1、C2组YKL-40中位水平低于C3、C4组(49 (32-74))与110 (68 - 222) ng·毫升−1;p < 0.01) (表1).
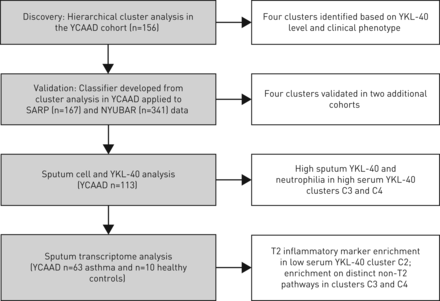
研究工作流程。YCAAD:耶鲁哮喘和呼吸道疾病中心;严重哮喘研究计划;NYUBAR:纽约大学/贝尔维尤哮喘登记处;T2: 2型。
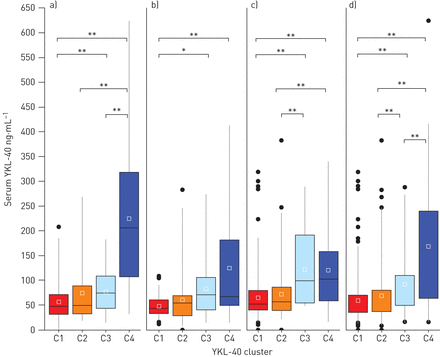
4个YKL-40哮喘分簇(C1-C4)的血清YKL-40水平。a)耶鲁哮喘和呼吸道疾病中心(YCAAD)队列中C1-C4的血清YKL-40水平。与C1和C2相比,C3和C4血清YKL-40水平升高。b, c)血清YKL-40水平在b)严重哮喘研究计划和c)纽约大学/贝尔维尤哮喘登记处队列与YCAAD队列的集群中相似。d)所有队列的综合值。*: p < 0.05;* *: p < 0.01。
为了验证YKL-40聚类及其相关的临床、生理和生物学特征,在YCAAD队列上使用递归划分开发了一个分类器算法(补充图E1)。该分类器应用于SARP (n=167)和NYUBAR (n=341)队列(图2 b和c,表1).使用该算法对SARP和NYUBAR队列进行分组,结果显示YKL-40水平的集群与YCAAD集群相似(图2 b和c,表1).在两个SARP队列中,C1和C2的血清YKL-40水平低于C3和C4 (46 (32-65))与68 (46 - 126) ng·毫升−1;p=0.002)和NYUBAR队列(53 (40-81))与99 (59 - 162) ng·毫升−1;p < 0.001) (表2).结合三个队列的数据显示,与C1和C2合并相比,C3组血清YKL-40水平高出58%,C4组高出133%(补充表E2)。总的来说,对三个哮喘队列的分析显示,血清YKL-40水平正常(C1和C2)或升高(C3和C4)的个体亚组是一致的。
YKL-40簇的临床和生理特征
为了确定每个YKL-40簇(C1-C4)的表型,我们比较了它们的临床特征(表1).在所有三个队列中,C1组(血清YKL-40水平最低的群集)的个体最普遍(53%),最年轻(中位年龄42岁),主要为女性(97%)。与血清YKL-40含量较高的C3和C4组相比,C1组的疾病严重程度较低,需要住院或机械通气的加重率较低,肺功能正常(图3 a - c)和较低剂量的药物。
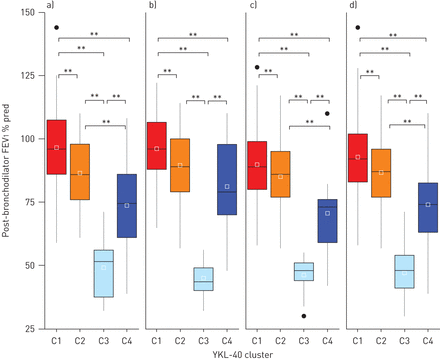
YKL-40群(C1-C4)在三个队列中的肺功能差异:a)耶鲁大学哮喘和气道疾病中心,b)严重哮喘研究项目和c)纽约大学/贝尔维尤哮喘登记处。使用支气管扩张剂后1 s用力呼气量(FEV)1)在所有三个队列的YKL-40聚类中均有显著差异。C3严重降低了支气管扩张剂后FEV1在所有三个队列中。d)所有队列的综合值。* *: p < 0.01。
YKL-40簇C2,也以低血清YKL-40水平为特征,是第二常见的簇(27%),主要是男性(70%)。C2是所有三个队列中使用短效β受体激动剂后出现支气管扩张的唯一集群(表1).尽管IgE水平在当地存在差异(表2), C2组的IgE水平高于C1组(172 (66-392))与96 (29 - 252) IU·毫升−1;p<0.01)(补充表E2)和与C3和C4相近的水平。因此,C1和C2的YKL-40水平相似,但支气管扩张反应和IgE水平不同。
高血清YKL-40簇C3包括年龄较大(中位年龄49岁)且比C1和C2有更严重疾病的个体。与其他三个分组相比,C3组哮喘发病最早(中位数6年),病程最长(中位数36年)(补充表E2),需要插管的近致命性哮喘(NFA)加重率最高(平均44%)(图4)和不可逆气流阻塞(支气管扩张后1 s的中位用力呼气量(FEV)1) 48%预测)(补充表E2)。此外,96%的C3患者使用吸入性和/或全身皮质类固醇(图3).尽管皮质类固醇的使用率很高,C3也需要额外的控制药物(表1).
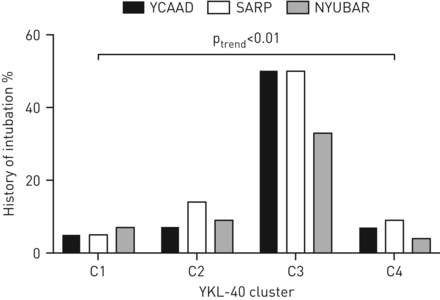
三个队列中YKL-40簇(C1-C4)中需要机械通气的近致命性哮喘加重的发生率。YCAAD:耶鲁哮喘和呼吸道疾病中心;严重哮喘研究计划;纽巴:纽约大学/贝尔维尤哮喘登记处。在所有三个队列中,C3的发生率都明显更高。
血清YKL-40水平最高的簇为C4 (图2).C4与C3在其他几个关键特征上有所不同,包括哮喘发病年龄较大(中位数43岁)、病程较短(中位数11年)(补充表E2),以及NFA发生率较低(9%)和持续性气流阻塞较轻(支气管扩张后FEV中位数)174%的预测)(图3而且4,补充表E2)。有趣的是,尽管仅在YCAAD中具有统计学意义,但C4在YCAAD群中肥胖率最高(67%)与41%;p = 0.007)。对所有三个队列的综合分析表明C3和C4组的肥胖率高于C1和C2组(57%)与35%;p < 0.01)。与C1相似,C4以女性为主(73%),但发病年龄不同(43岁)与15年;p < 0.01)。C4组的支气管扩张剂反应较C2组低(6%)与9%;p=0.04)(补充表E2)。
该分析在三个哮喘队列的YKL-40聚类中显示了一致的表型特征,包括一些在原始SARP聚类中没有观察到的特征,因此没有包括在聚类算法中(C2中较高的IgE和C3中较高的NFA率)。将YKL-40聚类与原始SARP聚类进行卡方检验,结果显示两种聚类方法的结果有统计学差异(p<0.001)(补充表E3)。总的来说,最显著的临床发现是C2以IgE升高为特征,而C3和C4以病情严重为特征,并以高血清YKL-40水平、严重加重和更严重的气流阻塞为区别。
痰细胞特征及与YKL-40簇相关的YKL-40蛋白表达
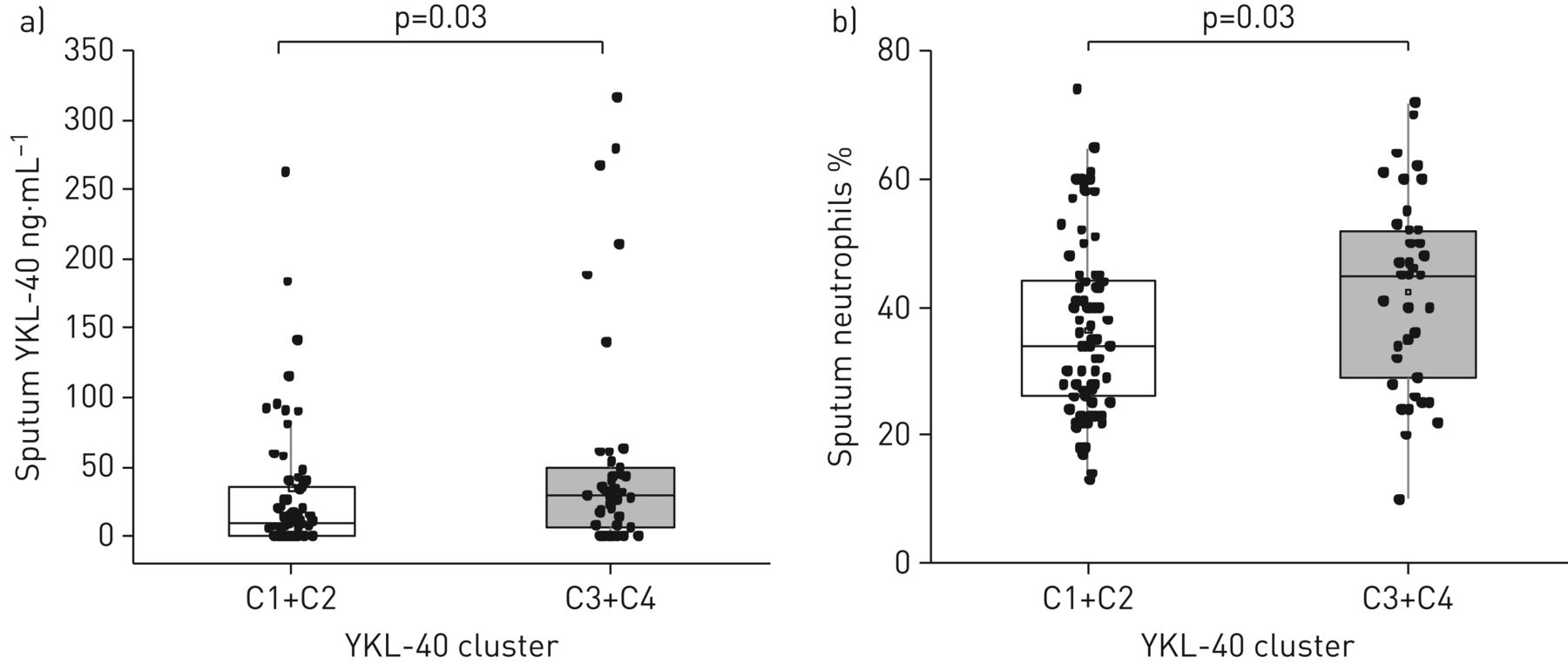
接下来,我们评估了YCAAD亚群诱导痰中YKL-40簇与细胞和蛋白质数据之间的相关性(n=113: C1, n=54;C2, n = 17;C3, n = 18;C4, n = 24) (表2).虽然跨组的两两比较没有发现痰细胞计数或YKL-40水平的特殊差异(表2),我们研究了血清YKL-40水平高的组簇(C3和C4)与血清YKL-40水平低的组簇(C1和C2)相比,是否有明显的痰液特征。血清YKL-40水平高的组(C3和C4)痰YKL-40蛋白水平较高(28.8与9.6 ng·毫升−1;p = 0.03) (图5一个)和较高的中性粒细胞水平(45%与34%;p = 0.03) (图5 b)与低血清YKL-40簇(C1和C2)比较。在这个YCAAD组的所有受试者中,痰YKL-40水平也与血清YKL-40水平相关(ρ=0.23;P =0.02)(数据未显示)。C1和C2的痰巨噬细胞百分比高于C3和C4 (58与44%;p < 0.01)。痰细胞特征分析显示,四组YKL-40的痰细胞数量或细胞活力无差异(数据未显示)。
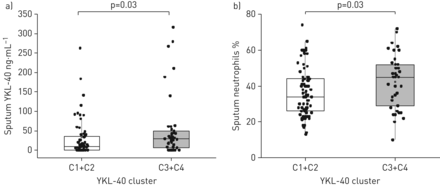
a) YKL-40痰水平和b)耶鲁哮喘和呼吸道疾病中心队列患者的痰中性粒细胞水平。与C1和C2相比,C3和C4簇的这两个值都有所增加。
痰T2炎症基因表达与YKL-40簇相关
根据观察到的YKL-40组间的临床、生理和痰液差异,我们假设气道中的基因表达谱也会在组间有所不同。因此,我们对发现YKL-40聚集的YCAAD患者的痰中mRNA表达进行了分析(n=63: C1, n=30;C2, n = 8;C3, n = 13;C4, n=12)和健康对照(n=10)(补充表E4)。痰CHI3L1在63例YCAAD患者中,或在组与健康对照组的两两比较(数据未显示)中,mRNA表达没有显著差异。
根据上述YKL-40低表达和C2中IgE水平升高,预测该簇以T2炎症为特征。与这一假设一致的是,与其他YKL-40组相比,C2组的T2表达特征(白细胞介素(IL)-4、IL-5和IL-13的平均mRNA表达量)最高(p=0.04) (图6).更广泛的转录组分析显示C2中IL-5信号通路有额外的转录富集,为该簇中T2富集提供了额外的证据(补充表E7和E8)。在该簇中存在最高的T2 mRNA表达,而YKL-40蛋白水平较低,这表明YKL-40主要与非T2炎症途径相关[11,17].相反,血清YKL-40高簇(C3和C4)的特征是痰中YKL-40蛋白水平高和气道中性粒细胞增多,而与痰中T2炎症标志物无关。综上所述,这些结果表明YKL-40可能是非t2哮喘的生物标志物。
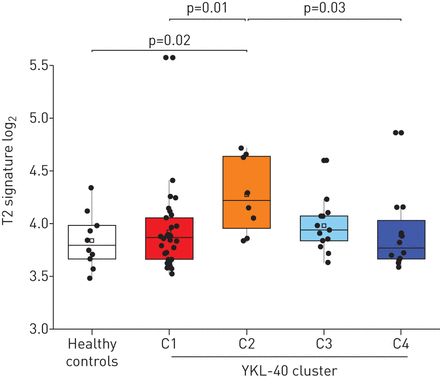
耶鲁哮喘和呼吸道疾病中心队列中YKL-40患者群和健康对照组2型(T2)痰转录组分析YKL-40簇中T2基因的平均表达水平(白细胞介素(IL)-4、IL-5和IL-13)显示C1和C4中T2基因表达水平较低。
痰非t2免疫基因表达与YKL-40簇相关
为了进一步阐明非t2炎症机制与YKL-40之间的关系,我们将63例YCAAD患者的痰表达谱与健康对照组的痰表达谱进行了比较。C1富集的基因涉及呼吸道免疫和对病毒感染网络的先天免疫反应(补充表E5和E6)。
高血清YKL-40簇(C3和C4)显示出类似的免疫反应基因本体富集。C3和C4均有11个基因具有相同的失调和相似的折叠变化幅度(补充表E9)。补充图E2显示了先前与气道生物学和哮喘相关的两个集群共有的5个基因的表达:TNFAIP3[18- - - - - -20.),MIR21[21,22),HLA-DQA1[23,24),IL1RAP[25),CCL18[26- - - - - -28].
C3特异的转录改变出现在图7补充表E10。鉴定的转录本包括与先天和适应性免疫反应有关的几个基因,如防御素DEFA1,DEFA1B而且DEFA3,TLR2,IFITM1,LY86(补充图E3)。C3转录本的途径富集分析发现了几种调控异常的途径,包括中性粒细胞胞外陷阱(NET)形成(NETosis),对气道细菌感染的免疫反应,以及通过促溶解脂质介质抑制中性粒细胞迁移。富集分析表明,在C3的个体中,有激活的先天免疫和NETosis通路,涉及无菌炎症和自身免疫(补充表E10和E11)。
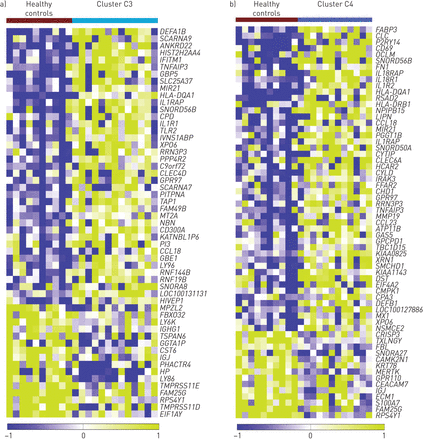
耶鲁哮喘和呼吸道疾病中心队列中YKL-40患者群和健康对照的非2型痰转录组分析a)与健康对照相比,C3中痰基因表达的热图显示先天免疫通路和NETosis通路的激活。b)与健康对照相比,C4患者痰基因表达的热图显示,C4患者白细胞介素(IL)-1和IL-18通路被激活。有关探针集和基因符号的详细信息,请参见补充表E10和E12。
与C3相反,与C4相关的转录谱显示了参与IL-1和IL-18通路的几个基因的激活(图7 b,补充图E4,补充表E12和E13),没有激活NETosis。
讨论
在本研究中,我们基于血清YKL-40水平和哮喘临床特征的结合,采用无监督聚类方法确定了4个YKL-40哮喘聚类。四个YKL-40集群中的两个(C3和C4)与其他两个集群(C1和C2)相比具有较高的血清YKL-40水平。该簇具有明显的临床、生理和痰炎细胞特征。在YCAAD队列中发现后,YKL-40集群在另外两个队列中得到验证,即。SARP NYUBAR。重要的是,YKL-40聚类具有明显的特征,将它们与原始的SARP聚类区分开,这是由于在聚类算法中包含了血清YKL-40水平,将这些临床特征与YKL-40蛋白的循环水平联系起来。这种聚类由非聚类特征支持,包括插管史和痰转录组的分子差异。已知的YKL-40和非T2炎症之间的联系是由发现低血清YKL-40簇C2中T2炎症基因的富集以及高血清YKL-40簇C3和C4中非T2炎症基因的富集所支持的。此外,C3中观察到的非t2网络与C4中不同。
在所有三个队列中,C3的受试者均与较高的NFA发生率和严重的气流阻塞有关。这些共同的特性可能是导致该群体YKL-40产量增加的机制基础。一些分子机制可以解释YKL-40与这些临床和生理特征之间的联系。第一种机制提出了NFA患者中机械应力与YKL-40之间的联系。鉴于已知对气道上皮细胞的机械应激可导致YKL-40的产生[29], nfa相关呼吸衰竭或严重气流阻塞反应中出现的气道拉伸可能是导致C3中YKL-40增加的原因,至少是部分原因。
第二种机制是YKL-40激活平滑肌增殖,导致气道重塑和严重的气流阻塞。这一机制基于以下观察结果:1)在气道上皮细胞中,YKL-40暴露导致IL-8合成增加和平滑肌增殖在体外2) YKL-40增加蛋白酶激活受体-2活性,促进支气管平滑肌增殖[5,6].
我们研究的一个新发现是,在C3中痰中性粒细胞和参与NETs形成的基因表达均增加。NET作为由中性粒细胞和嗜酸性粒细胞形成的细胞外DNA陷阱,nettosis是与NET形成相关的高度免疫原性细胞死亡过程。nettosis与无菌性炎症、自身免疫和与炎症相关的慢性疾病有关[30.].先前的报道发现过敏性哮喘患者的气道中存在NETs,而我们的研究为NETosis在以高YKL-40为特征的严重哮喘亚组中的作用提供了支持。31].
考虑到C3与严重的固定气流阻塞和NFA相关,以及通过吸入重组dna酶(类似于囊性纤维化)靶向气道内NET生成的可能性,对哮喘集群C3的识别是相关的[32].
与C3相比,YKL-40簇C4血清YKL-40水平最高,发病年龄较大(成人),气流阻塞较少。C4的特征还包括哮喘控制不良、频繁的严重加重需要住院治疗和高肥胖率(尽管仅在YCAAD队列中具有统计学意义)。与肥胖的联系可以解释,至少部分解释了C4中YKL-40水平升高的原因。已知内脏脂肪组织是YKL-40的来源[33].更重要的是,肥胖和哮喘之间的联系可能是通过哮喘中内脏脂肪积累和T2炎症引起的YKL-40升高的组合介导的[34].尽管C3和C4在痰中性粒细胞计数和气道相关基因激活方面有相似之处TNFAIP3,HLA-DQA1,IL1RAP,CCL18而且MIR21C4痰基因表达谱显示IL-1和IL-18通路明显激活。C3和C4之间的差异,包括不同的基因表达谱和血清YKL-40水平的不同倍性差异,表明它们与YKL-40的分子联系可能由至少两种不同的非t2炎症机制介导。我们发现了痰CHI3L1YCAAD队列中各组间mRNA表达无差异,提示C3和C4中较高的血清YKL-40水平可能与独立于升高的机制有关CHI3L1痰细胞mRNA表达(即。由支气管上皮细胞、平滑肌和内脏脂肪产生)。
聚类分析还发现了两个低血清YKL-40聚类。C1为女性亚组,以非严重哮喘和保留肺功能为特征,而C2为高T2亚组,IL-5信号通路富集,类似于先前发现的高T2基因表达的哮喘集群[17,35].这些临床差异和血清YKL-40水平的显著差异证实了我们使用这种方法区分哮喘亚组的能力。根据临床特征和血清YKL-40对不同哮喘患者亚组进行特征描述的新框架将使进一步的研究能够揭示这些关联背后的独特机制。
我们的研究确实有一些局限性,包括它的横断面性质。我们无法确定YKL-40簇是否随着时间的推移保持稳定,还是受到环境或药物变化的影响,包括对后者的依从性。因此,有必要对这些簇进行前瞻性研究,以了解这些簇与时间和哮喘治疗的相互作用。并非所有组群的成对比较都显示出相同的差异。尽管两个验证队列的纳入标准、治疗效果、样本量和分类器性能的差异相似,但研究队列的局部差异可能部分解释了这一点。然而,我们的方法揭示了三个队列中具有几乎相同的生理和临床特征的集群和相似的YKL-40水平。选择其他哮喘特征和模型可能会产生不同的聚类结果;然而,我们试图平衡集群发现和在独立队列中验证这些新集群的能力。随着我们对YKL-40生物学的理解的提高和其他哮喘队列在其患者特征中纳入痰转录组,我们的模型可能会随着时间的推移而发展。
总之,基于YKL-40水平和疾病的临床和生理特征对哮喘受试者进行聚类分析,我们确定了四个不同的受试者群。两组由血清YKL-40水平升高连接,但由YKL-40水平升高的幅度和特定的临床/生理异常进行区分。本研究表明,结合哮喘的临床和生理特征,使用YKL-40水平可以识别出具有明显临床特征和严重气流阻塞、NFA和频繁严重加重风险的特定亚组患者。高血清YKL-40簇(C3和C4)可能与鉴别伴有非t2疾病的严重哮喘患者具有临床和生物学意义。
补充材料
披露的信息
补充材料
基准线Chupperj - 00800 - 2017 - _chupp
l·科恩erj - 00800 - 2017 - _cohn
通用Crisafierj - 00800 - 2017 - _crisafi
J.L.戈麦斯erj - 00800 - 2017 - _gomez
N.N. Jarjourerj - 00800 - 2017 - _jarjour
j . Reibmanerj - 00800 - 2017 - _reibman
确认
纽约大学/Bellevue哮喘登记处:作者感谢Colton家族的持续支持和Maria Elena Fernandez Beros(纽约大学医学院,纽约,纽约,美国)。耶鲁哮喘和呼吸道疾病中心:作者感谢Sarah Marone和Eleni Kapagiannidou(耶鲁医学院,纽黑文,CT, USA)。
脚注
本文的补充资料可从www.qdcxjkg.com
支持声明:NIH T32HL007778-18和T15LM007056-26。R01HL095390-03;1K01HL125474-01, R01HL069116和GCRC RR03186, CTSA UL1 TR000038;FAMRI青年临床科学家奖113393;航空同行奖2010。本文的资助信息已存入Crossref资助者注册表.
利益冲突:可以在本文旁边找到披露www.qdcxjkg.com
- 收到了2017年4月17日。
- 接受2017年7月16日。
- 版权©2017人队











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}