





摘要gydF4y2Ba
蛋白转化酶(PCs)是丝氨酸蛋白酶,负责在鼻息肉病过程中参与上气道重塑的许多前体蛋白的蛋白水解成熟。我们之前已经发现PC1/3在人鼻黏膜中表达。gydF4y2Ba
但PC1/3是否与鼻息肉形成有关尚未研究。为了深入了解PC1/3在鼻息肉中的功能作用,我们通过免疫染色、Western blotting和酶学检测PC1/3在鼻息肉中的表达,并利用气道上皮细胞系NCI-H292生成稳定表达PC1/3的细胞。gydF4y2Ba
与正常鼻黏膜相比,鼻息肉PC1/3表达增加。PC1/3在正常鼻黏膜神经内分泌细胞中有表达,在鼻息肉杯状细胞和纤毛细胞中也有表达。稳定表达PC1/3的NCI-H292细胞表现出形态改变,细胞增殖和迁移增强,E-cadherin和细胞角蛋白下调,N-cadherin、波形蛋白、基质金属蛋白酶-2、胶原- i、snail和twist上调。重要的是,PC1/3表达与培养的人鼻上皮细胞和鼻息肉的上皮-间充质转化呈正相关。gydF4y2Ba
综上所述,我们的数据提示PC1/3过表达诱导气道上皮细胞形态和表型的上皮-间充质转化改变,这些改变可能参与鼻息肉的发病机制。gydF4y2Ba
摘要gydF4y2Ba
PC1/3过表达诱导气道上皮-间质转化的形态学和表型改变gydF4y2Bahttp://ow.ly/on9hZgydF4y2Ba
介绍gydF4y2Ba
慢性鼻窦炎伴鼻息肉(CRSwNP)是一种慢性炎症性鼻窦疾病,影响全球4%的人[gydF4y2Ba1gydF4y2Ba].其特征是持续性炎症和结构变化,称为气道重塑[gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2gydF4y2Ba].CRSwNP气道重塑由基底膜增厚、上皮下纤维化、细胞外基质(ECM)沉积增多和间质水肿引起[gydF4y2Ba3.gydF4y2Ba].CRSwNP与其他疾病相关,如哮喘、囊性纤维化、支气管扩张和阿司匹林敏感性[gydF4y2Ba1gydF4y2Ba].然而,我们对这些疾病的发病机制的认识仍然有限。gydF4y2Ba
上皮细胞可经历上皮-间充质转化(EMT)以应对组织损伤和炎症,导致器官纤维化和癌症[gydF4y2Ba2gydF4y2Ba,gydF4y2Ba4gydF4y2Ba].在EMT过程中,上皮细胞失去其定义特征,如E-cadherin和尖基底极性,而获得间质特征,包括α-平滑肌肌动蛋白(SMA)、波形蛋白、基质金属蛋白酶(MMPs)和转录因子snail、slug和twist的上调[gydF4y2Ba2gydF4y2Ba,gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba].越来越多的证据表明肺上皮损伤和异常上皮修复反应诱导上皮细胞持续激活,并通过EMT导致与组织纤维化相关的病理过程[gydF4y2Ba6gydF4y2Ba,gydF4y2Ba7gydF4y2Ba].最近的研究表明,α-SMA在鼻息肉和缺氧诱导的EMT的上皮细胞内层表达gydF4y2Ba通过gydF4y2Ba培养的人鼻上皮细胞中的SMAD (mother against decentapleichomolog)-3依赖机制[gydF4y2Ba8gydF4y2Ba],提示EMT在鼻息肉发病机制中具有重要作用。gydF4y2Ba
许多蛋白,如转录生长因子(TGF)-β、血管内皮生长因子(VEGF)、胰岛素样生长因子(IGF)-1、上皮生长因子、成纤维细胞生长因子-2和MMPs都能诱导EMT [gydF4y2Ba9gydF4y2Ba].有趣的是,这些蛋白质是作为非活性前体蛋白合成的,需要通过蛋白转换酶(PCs)去除前域才能成为生物活性形式[gydF4y2Ba10gydF4y2Ba].PCs是一个钙依赖的枯草菌素样内蛋白酶家族,包括furin、PC1/3、PC2、PC4、配对碱性氨基酸裂解酶(PACE)-4、PC5/6、PC7/淋巴瘤蛋白转化酶/PC8和哺乳动物枯草菌素/可心同工酶(SKI)-1/S1P [gydF4y2Ba10gydF4y2Ba].由于pc负责激活与癌症和相关转移过程有关的蛋白质前体,它们可能是治疗不同增生性疾病的有希望的靶点。事实上,早期的研究报告了pc在各种人类癌症(如肺癌)中的表达升高[gydF4y2Ba11gydF4y2Ba],乳腺癌[gydF4y2Ba12gydF4y2Ba及与内分泌有关的癌症[gydF4y2Ba13gydF4y2Ba].gydF4y2Ba
我们之前已经证明PC1/3在人类鼻腔粘膜中表达[gydF4y2Ba14gydF4y2Ba].但PC1/3是否与鼻息肉形成有关尚未研究。为了深入了解PC1/3在鼻息肉上皮中的功能作用,我们检测了PC1/3在鼻息肉中的表达,并使用NCI-H292细胞构建了稳定的表达PC1/3的细胞系。基于gydF4y2Ba在体外gydF4y2BaPC1/3可诱导气道上皮细胞emt样过程。gydF4y2Ba
材料和方法gydF4y2Ba
人类被试gydF4y2Ba
下鼻甲黏膜(正常鼻黏膜)标本取自6例鼻中隔成形术患者。6例鼻息肉标本取自无哮喘、阿司匹林敏感性或囊性纤维化的慢性鼻窦炎患者。所有患者在研究前3个月均未接受过鼻内药物、口服类固醇或抗生素治疗,所有患者的过敏性皮肤点刺试验均为阴性。gydF4y2Ba
细胞培养和转染gydF4y2Ba
延世大学医学院机构审查委员会批准了这项研究(IRB# 4-2012-0136)。正常人鼻上皮(NHNE)细胞取自鼻甲切除术后的患者。NHNE细胞采用气液界面培养系统(在线补充材料)。为了建立稳定表达PC1/3的细胞,NCI-H292细胞用FuGENE™6(罗氏,印第安纳波利斯,IN,美国)转染RcCMV编码的小鼠PC1/3。gydF4y2Ba
RNA分离、实时PCR和半定量逆转录酶-PCRgydF4y2Ba
根据制造商的方案,使用TRIzol试剂(Invitrogen公司,Gaithersburg, MD, USA)分离总RNA。合成cDNA,利用特异引物进行PCR扩增(gydF4y2Ba表1gydF4y2Ba以及在线补充材料)。gydF4y2Ba
西方墨点法gydF4y2Ba
标本用研钵和杵在液氮中研磨成细粉末。研磨组织和NCI-H292细胞在含有蛋白酶/磷酸酶抑制剂鸡尾酒的细胞裂解缓冲液(Invitrogen, Camarillo, CA, USA)中均质,置于冰上,经过8%十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE),然后使用适当的抗血清(在线补充材料)进行Western blotting。gydF4y2Ba
免疫组化和免疫荧光染色gydF4y2Ba
PC1/3在正常鼻黏膜和鼻息肉标本石蜡切片(4 μm)中进行免疫染色。免疫荧光染色,从正常鼻黏膜和鼻息肉标本的上皮细胞制备细胞自旋玻片(在线补充材料)。gydF4y2Ba
酶化验gydF4y2Ba
用100 mM醋酸钠、pH 5.5、1% Triton X-100和蛋白酶抑制剂混合溶液对正常鼻黏膜和鼻息肉进行超声治疗。12 000×离心2分钟gydF4y2BaggydF4y2Ba,上清液用于PC酶测定(在线补充材料)。gydF4y2Ba
在体外gydF4y2Ba伤口化验gydF4y2Ba
NCI-H292细胞生长在胶原蛋白涂层的12孔培养板上。用5 μg·mL孵育细胞gydF4y2Ba−1gydF4y2Ba丝裂霉素C (Sigma Chemicals, Saint Louis, MO, USA)阻断细胞增殖3小时gydF4y2Ba在体外gydF4y2Ba用无菌移液管尖端在孔的中心做圆形剥落区,然后用PBS洗涤去除游离细胞。在这个时间点(t=0 h),拍摄伤口边缘。然后,在不同时间点拍摄创面边缘相同的视野。使用显微镜上安装的摄像机拍摄细胞形态,并使用Adobe Photoshop软件进行处理。gydF4y2Ba
细胞增殖实验gydF4y2Ba
根据制造商的协议,使用标准比色法细胞计数试剂盒-8(日本熊本Dojindo Laboratories, Kumamoto, Japan)测定细胞生长。在450nm处每24小时测量一次吸光度,为期3天。gydF4y2Ba
明胶zymographygydF4y2Ba
酶谱法检测条件培养基(在线补充材料)中分泌的MMP-2活性。gydF4y2Ba
统计分析gydF4y2Ba
数据用平均值±表示gydF4y2BasdgydF4y2Ba三个实验。值分布的差异用单因素方差分析进行统计验证。使用GraphPad Prism 3程序(GraphPad Software, Inc., La Jolla, CA, USA)进行统计分析。在p<0.05时,差异被认为是显著的。gydF4y2Ba
结果gydF4y2Ba
PC1/3表达于鼻息肉上皮的杯状细胞和纤毛细胞以及神经内分泌细胞中gydF4y2Ba
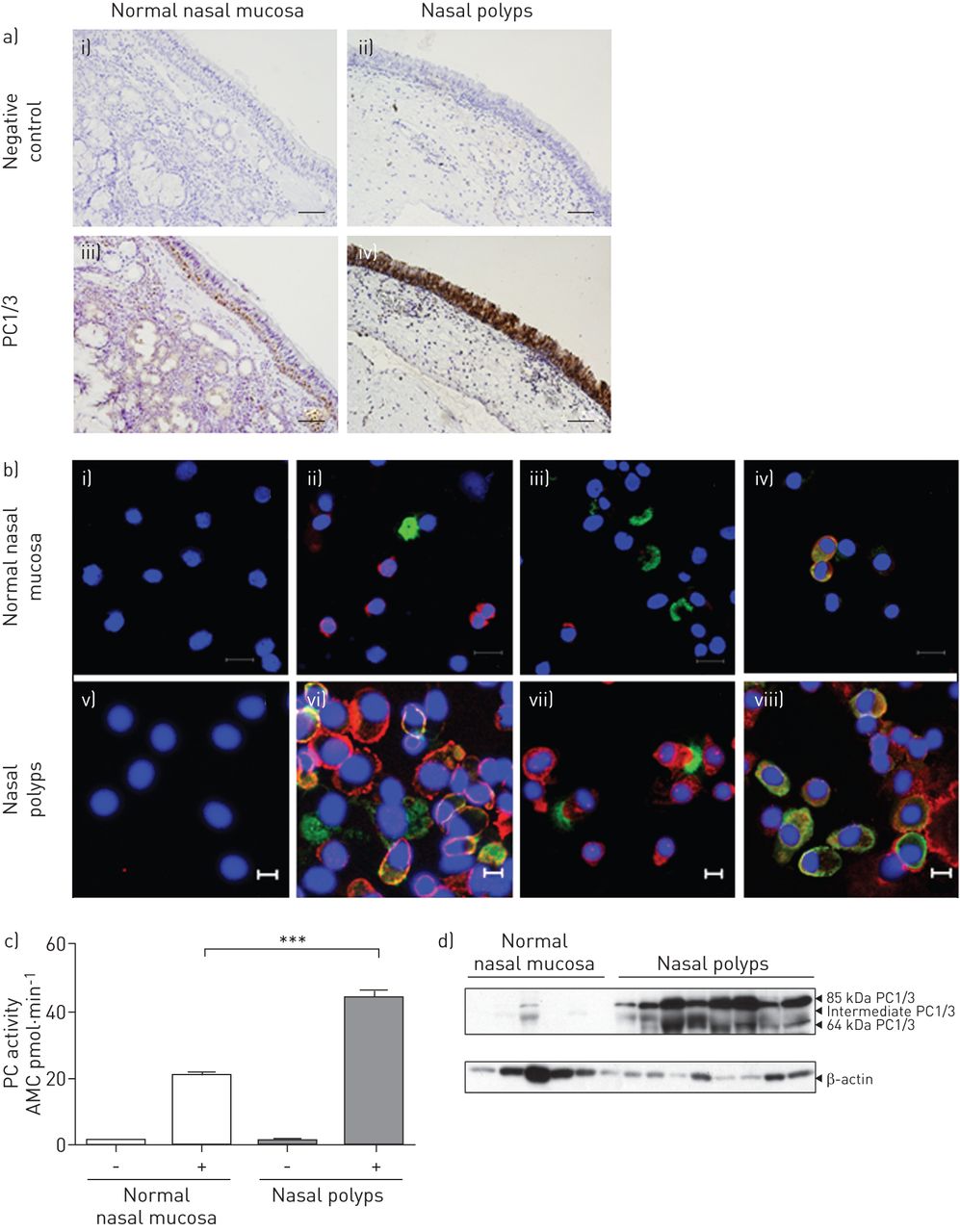
我们首先通过免疫组化染色检测PC1/3在正常鼻黏膜和鼻息肉中的分布。gydF4y2Ba图1一个gydF4y2Ba显示PC1/3的免疫反应局限于正常鼻黏膜上皮基底部的一些特化细胞(gydF4y2Ba图1一个gydF4y2Ba(iii)),而在鼻息肉中,PC1/3在整个上皮细胞层中被强烈检测到(gydF4y2Ba图1一个gydF4y2Ba(四))。IgG对照未检测到染色(gydF4y2Ba图1一个gydF4y2Ba(二),(我)。gydF4y2Ba
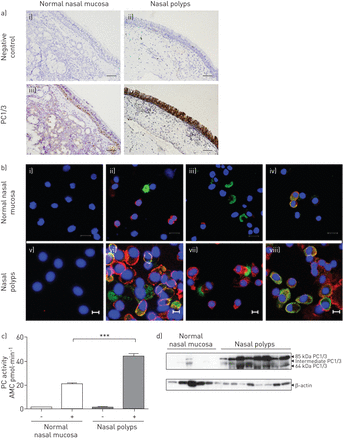
蛋白转化酶(PC)1/3在正常人鼻粘膜和鼻息肉中的定位。a)用抗PC1/3血清免疫组化染色分析PC1/3定位。PC1/3的积极信号是局限于一些专业基底细胞在正常鼻粘膜(iii),而鼻息肉显示强烈PC1/3免疫反应性上皮细胞(iv)。一个免疫球蛋白控制显示(i, ii)。实验重复使用四个正常鼻粘膜和四个鼻息肉标本。比例尺条= 100μm。b)使用抗PC1/3和粘蛋白(MUC)-5AC (ii, vi)、抗PC1/3和乙酰微管蛋白(iii, vii)或抗PC1/3和嗜铬粒蛋白A (ChrA) (iv, vi)的免疫染色分析PC1/3在正常鼻粘膜和鼻息肉中的定位。使用从正常鼻黏膜(i-iv)和鼻息肉(v-viii)的上皮细胞制备的细胞旋片。细胞核用4′,6-二氨基-2-苯基吲哚(DAPI)染色。IgG对照为(i, v)。i - iv标尺=100 μm;V-viii)比例尺=10 μm。c)从正常鼻黏膜和鼻息肉中提取的组织提取物(100 μg)测定细胞内PC的总活性。 Data represent n=6 tissues per group, each measured in triplicate, mean±sdgydF4y2Ba.AMC: 7-amido-4-methylcoumarin。***:单因素方差分析p<0.001。d)对正常鼻黏膜和鼻息肉的粗提物(50 μg)进行Western blot分析PC1/3的表达。β-肌动蛋白作为负载对照。gydF4y2Ba
为了检测正常鼻粘膜和鼻息肉中哪些细胞类型表达PC1/3,我们使用抗血清和mucin -5AC(杯状细胞标记物)对从正常鼻粘膜和鼻息肉上皮细胞制备的细胞旋片进行双重免疫细胞荧光染色,抗血清抗PC1/3和乙酰化微管蛋白(一种纤毛细胞标记物)或抗血清抗PC1/3和嗜铬粒蛋白a (ChrA,一种分泌颗粒标记物)[gydF4y2Ba15gydF4y2Ba].与免疫组化结果一致,PC1/3未与MUC5AC (gydF4y2Ba图1 bgydF4y2Ba(ii)和乙酰化微管蛋白(gydF4y2Ba图1 bgydF4y2Ba(iii)正常鼻黏膜。相比之下,在鼻息肉中,一些细胞PC1/3和MUC5AC均被阳性标记(gydF4y2Ba图1 bgydF4y2Ba(vi):黄色),其他则单独标记PC1/3 (gydF4y2Ba图1 bgydF4y2Ba(vi)红色)或MUC5AC (gydF4y2Ba图1 bgydF4y2Ba(vi):绿色),表明PC1/3在杯状细胞中表达。此外,PC1/3也存在于纤毛细胞中,在鼻息肉中与乙酰化微管蛋白共定位证实了这一点(gydF4y2Ba图1 bgydF4y2Ba(七))。在正常鼻粘膜和鼻息肉中,PC1/3位于神经内分泌细胞中含有chra的分泌颗粒中,合并图中可见黄色物体(gydF4y2Ba图1 bgydF4y2Ba(iv, viii)。IgG对照无染色(gydF4y2Ba图1 bgydF4y2Ba(我))。gydF4y2Ba
接下来,我们通过测量总PC酶活性来探讨活性PC在鼻息肉中的储存是否与正常鼻粘膜不同。鼻息肉中pc的总酶活性明显高于正常鼻粘膜(gydF4y2Ba图1 cgydF4y2Ba).此外,Western blotting显示PC1/3在鼻息肉中表达明显增加(gydF4y2Ba图1 dgydF4y2Ba).gydF4y2Ba图1 dgydF4y2Ba显示3种PC1/3物种,分子量分别为~ 64 kDa (c端截短型)、71 kDa(中间型,c端内部发生裂解)和85 kDa(成熟型),因为PC1/3在激活过程中发生蛋白水解裂解。这些数据提示PC1/3可能与鼻息肉形成有关。gydF4y2Ba
pc1 /3-过度产生的NCI-H292细胞出现形态学改变gydF4y2Ba
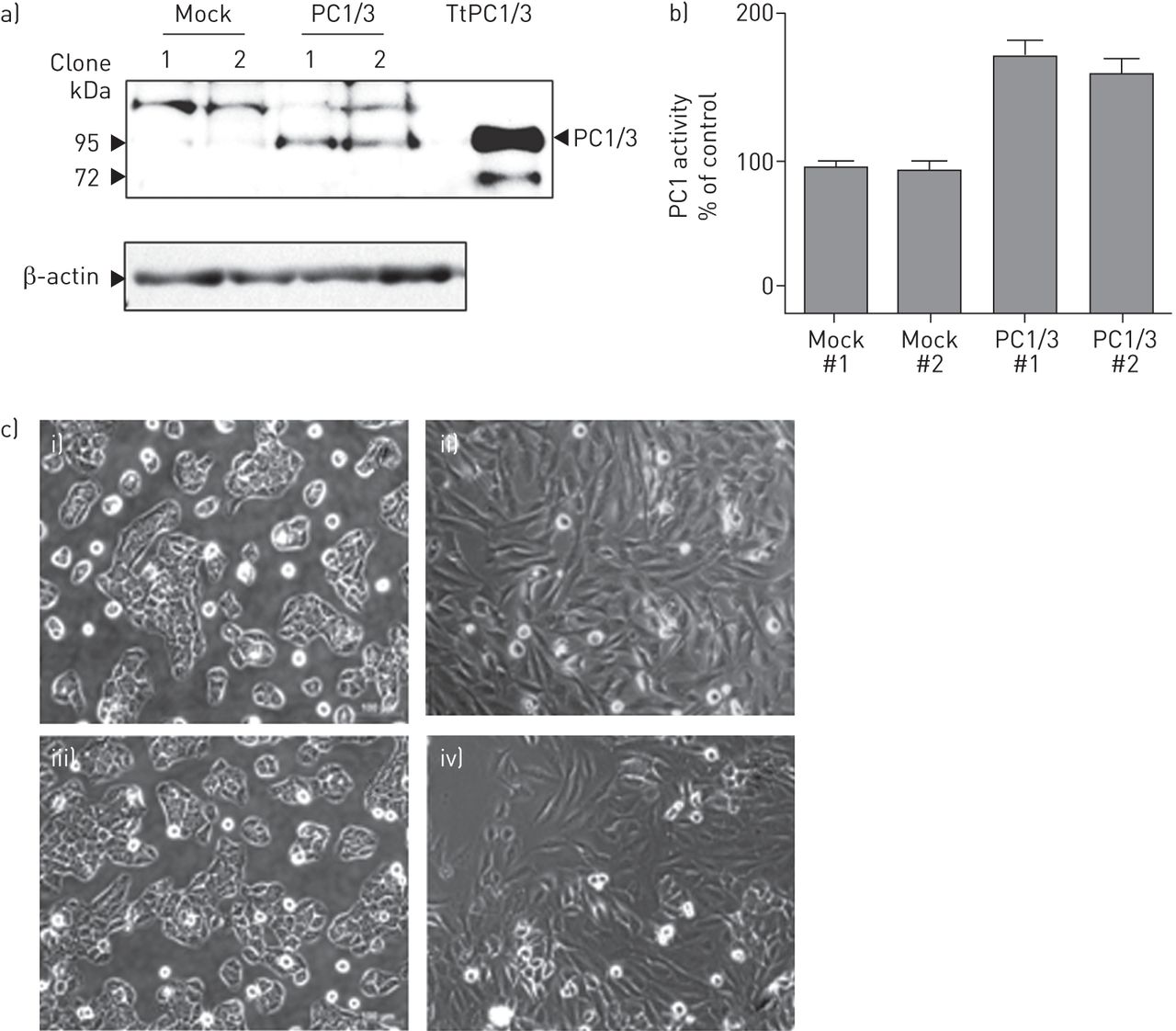
为了探讨PC1/3在气道上皮细胞中的功能作用,我们利用NCI-H292细胞构建了稳定的PC1/3细胞系。稳定转染含有PC1/3 cDNA的RcCMV载体(命名为PC1/3细胞;2个独立的细胞:1和2)或单独的RcCMV作为对照(模拟细胞:1和2)。Western blot分析显示分子量为85 kDa的免疫反应条带对应于PC1/3 (gydF4y2Ba图2一个gydF4y2Ba, PC1/3);瞬时转染PC1/3 cDNA的克隆作为阳性对照(TtPC1/3)。我们通过使用条件培养基(gydF4y2Ba图2 bgydF4y2Ba).有趣的是,模拟细胞呈鹅卵石状,细胞与细胞之间有连接(gydF4y2Ba图2 cgydF4y2Ba(i, iii)),而PC1/3细胞呈细长纺锤形,失去细胞与细胞的接触(gydF4y2Ba图2 cgydF4y2Ba(四),(ii)。gydF4y2Ba
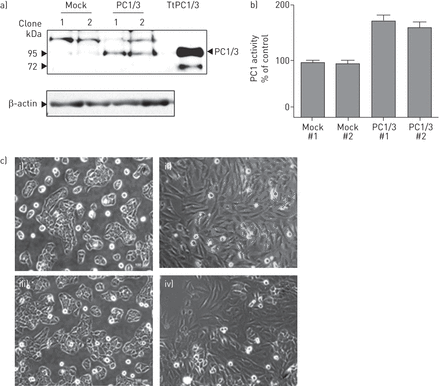
蛋白转化酶(PC)1/3过表达诱导NCI-H292细胞形态学改变。a)将含有mPC1/3 cDNA的表达质粒稳定转染NCI-H292细胞,并用Western blot分析PC1/3的产物。在PC1/3细胞中检测到分子量为87 kDa的免疫反应产物,与PC1/3的预测分子大小相对应。“Mock”为用空载体转染的稳定细胞系,“PC1/3”为表达PC1/3的稳定细胞系;使用两个独立的高表达细胞进行实验。以瞬时表达PC1/3的NCI-H292细胞(TtPC1/3)作为阳性对照。b) NCI-H292细胞在12孔板中培养过夜。培养基改为100 μg·mL的Opti-MEMgydF4y2Ba−1gydF4y2Ba用50 μ l培养基测定PC1/3活性。c) i)和iii)模拟细胞呈扁平的鹅卵石形状,连接紧密,而ii), iv) PC1/3细胞缺乏细胞间接触,呈更细长的形状。gydF4y2Ba
PC1/3过表达调节NCI-H292细胞的生长和迁移gydF4y2Ba
接下来,我们在模拟和PC1/3细胞中评估细胞增殖和迁移。在3天内,PC1/3细胞与模拟细胞相比,细胞增殖显著增强(gydF4y2Ba图3gydF4y2Ba).为了评估迁移活性,我们在5 μg·mL的浓度下进行迁移实验gydF4y2Ba−1gydF4y2Ba丝裂霉素C抑制细胞增殖,并证实伤口愈合归因于细胞迁移。伤后6、24 h测量创面面积。gydF4y2Ba图3 bgydF4y2Ba显示伤口面积随时间而减少。在24小时内,模拟细胞恢复了约44%的创面面积,而PC1/3细胞完全恢复了创面面积,表明PC1/3细胞获得了迁移表型。gydF4y2Ba
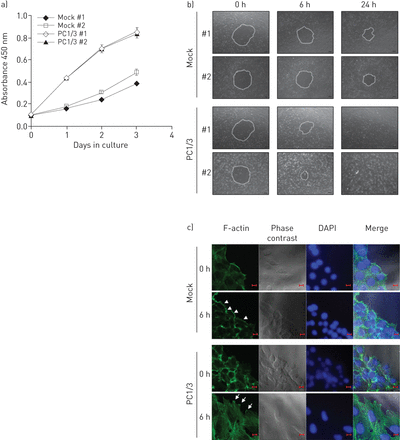
蛋白转化酶(PC)1/3过表达促进NCI-H292细胞增殖和迁移。a)使用细胞计数试剂盒-8试剂盒(Dojindo Laboratories, Kumamoto, Japan)在第1、2和3天对细胞增殖进行定量。数据用平均值±表示gydF4y2BasdgydF4y2Ba三个单独的实验,每个实验都有三份。b)细胞在i型胶原蛋白涂层的12孔培养板上融合。用丝裂霉素C (5 μg·mL)处理细胞gydF4y2Ba−1gydF4y2Ba)以阻止细胞增殖。伤后6、24 h测量创面面积。伤口边缘用白色虚线表示。c)使用异硫氰酸荧光素(FITC)-phalloidin对细胞进行丝状(F)-肌动蛋白染色。损伤后6小时,模拟细胞表现出典型的静止表型,边缘有肌动蛋白束(箭头),而PC1/3细胞则在整个细胞中形成广泛的肌动蛋白应力纤维,并在损伤前缘有丝足伸长(箭头)。DAPI: 4 ', 6-diamidino-2-phenylindole。规模酒吧= 10μm。gydF4y2Ba
迁移细胞通常在前缘表现出与肌动蛋白重组相关的质膜突出[gydF4y2Ba16gydF4y2Ba].我们通过异硫氰酸荧光素-phalloidin染色进一步分析了伤口边缘的细胞形状和丝状肌动蛋白的分布(gydF4y2Ba图3 cgydF4y2Ba).6 h后,模拟细胞在细胞边缘形成边缘肌动蛋白束,这是典型的静止表型。相比之下,PC1/3细胞呈拉长铺展状,在整个细胞中形成广泛的肌动蛋白应力纤维,并在创面前缘形成片状和丝状的延伸。综上所述,我们的结果表明PC1/3过表达可以促进细胞增殖,并诱导NCI-H292细胞的迁移表型。gydF4y2Ba
PC1/3过表达增强NCI-H292细胞中EMT标记物、MMPs和ECM蛋白的表达gydF4y2Ba
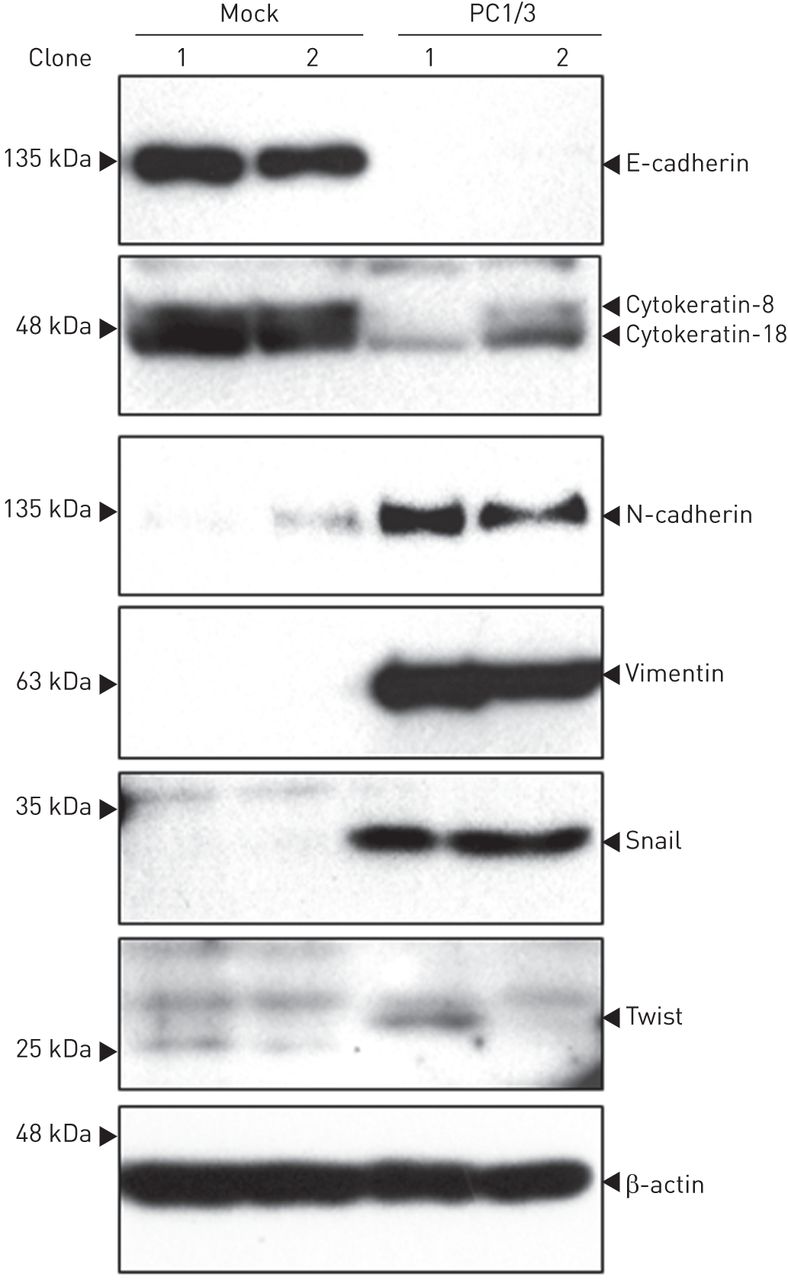
Western blotting分析EMT标记物的表达。PC1/3细胞上皮标记物E-cadherin、细胞角蛋白-8和-18表达下调,间质标记物N-cadherin、波形蛋白、snail和twist表达上调(gydF4y2Ba图4gydF4y2Ba).gydF4y2Ba
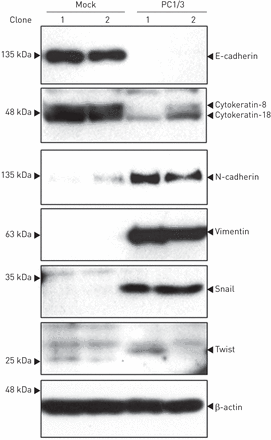
蛋白转化酶(PC)1/3过表达诱导NCI-H292细胞上皮-间充质转化相关蛋白生产的上调。PC1/3细胞E-cadherin、cytokeratin-8和-18表达显著降低,N-cadherin、vimentin、snail和twist表达显著升高。gydF4y2Ba
由于MMPs、α5整合素和ECM蛋白胶原和纤维连接蛋白的上调是EMT的共同特征[gydF4y2Ba17gydF4y2Ba- - - - - -gydF4y2Ba20.gydF4y2Ba],我们利用实时荧光定量PCR检测这些基因在模拟细胞和PC1/3细胞中的表达水平。gydF4y2Ba图5一个gydF4y2Ba结果表明,PC1/3过表达导致NCI-H292细胞中MMP-2、MMP-3和α5-整合素表达明显增强,而MMP-1、-7、-9和-10表达水平未发生变化;Western blotting和酶谱分析证实MMP-2表达增加(gydF4y2Ba图5 bgydF4y2Ba).我们还观察到I型胶原显著上调,IV型胶原和纤维连接蛋白表达增加(gydF4y2Ba图5度gydF4y2Ba);Western blotting也证实了I型胶原蛋白上调(gydF4y2Ba图5 dgydF4y2Ba).综上所述,这些数据表明PC1/3在NCI-H292细胞中可诱导emt样表型。gydF4y2Ba
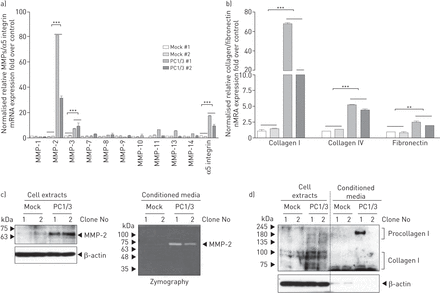
蛋白转化酶(PC)1/3过表达上调NCI-H292细胞基质金属蛋白酶(MMPs)和细胞外基质蛋白的表达。a) Real-time quantitative reverse transcripase (RT)-PCR分析显示,PC1/3细胞中MMPs和α5-integrin的基因表达量显著高于模拟细胞。数据以平均值±表示gydF4y2BasdgydF4y2Ba三个单独的实验,每个实验都有三份。***:单因素方差分析p<0.001。b)实时定量RT-PCR检测细胞外基质基因I型胶原、IV型胶原、纤连蛋白的表达情况。数据以平均值±表示gydF4y2BasdgydF4y2Ba三个单独的实验,每个实验都有三份。* *: p < 0.01;***: p<0.001,均通过单因素方差分析。c) PC1/3细胞中MMP-2的表达和活性上调,通过细胞提取物的Western blot分析(左图)和条件培养基的酶谱分析(右图)证实。d)细胞提取物和条件培养基的Western blot分析显示I型胶原蛋白表达增加。gydF4y2Ba
PC1/3在培养的NHNE细胞和鼻息肉中表达均与EMT呈正相关gydF4y2Ba
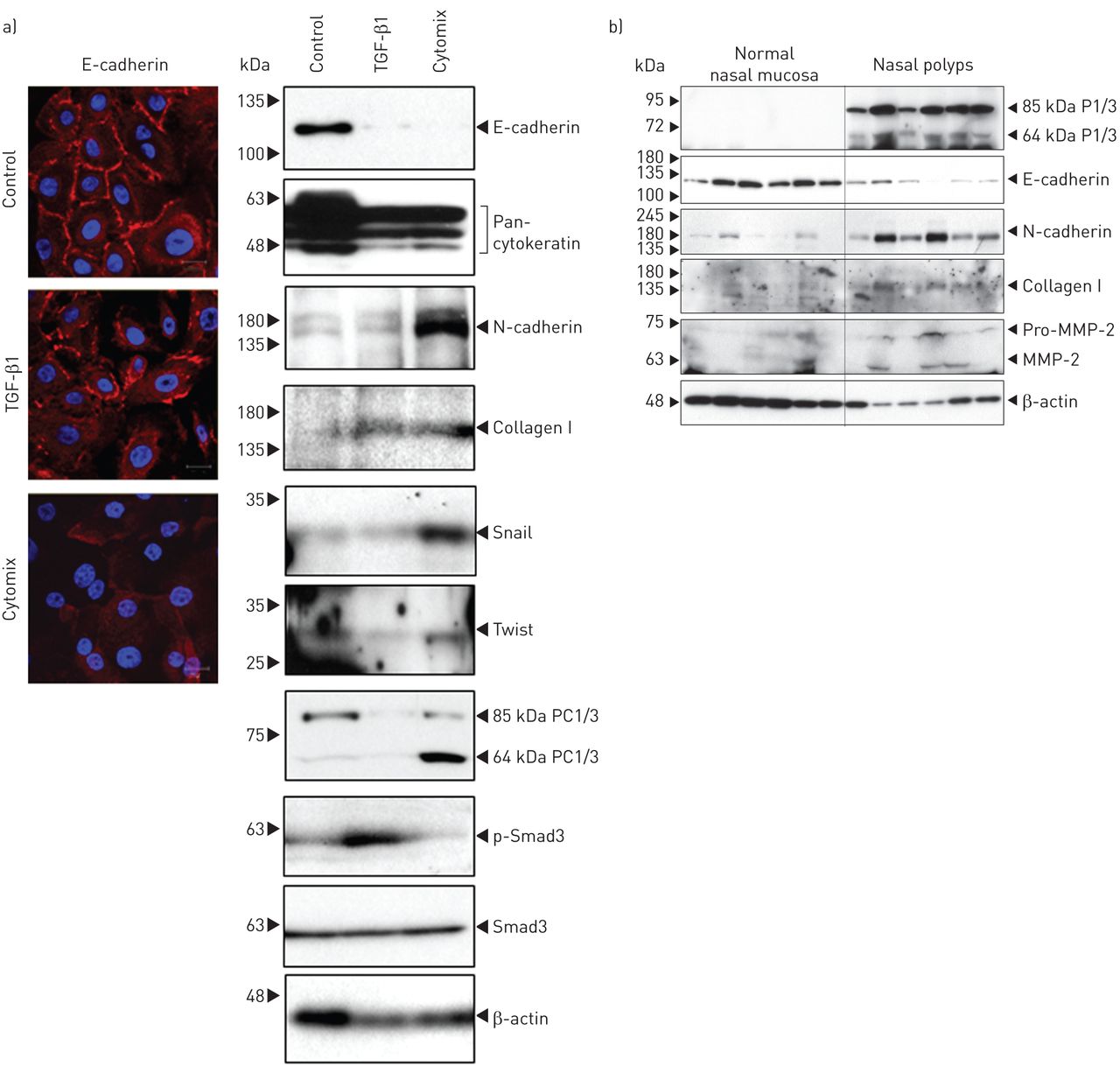
接下来,我们研究了TGF-β1和cytomix(肿瘤坏死因子(TNF)-α和白细胞介素(IL)-1β的组合)在NHNE细胞中诱导EMT时PC1/3的表达是否发生改变;TGF-β1、TNF-α和IL-1β可诱导EMTgydF4y2Ba在体外gydF4y2Ba[gydF4y2Ba6gydF4y2Ba,gydF4y2Ba7gydF4y2Ba].gydF4y2Ba图6gydF4y2BaTGF-β1和cytomix抑制E-cadherin和泛细胞角蛋白的表达,增加了多种间质标志物N-cadherin、collagen I、snail和twist的表达。有趣的是,与TGF-β1处理的细胞相比,NHNE细胞中N-cadherin、snail和twist的表达水平显著升高。PC1/3的表达是由cytomix诱导的,而不是由TGF-β1诱导的,说明PC1/3参与了cytomix介导的EMT,而不是由TGF-β1诱导的EMT;通过检测磷酸化smad3 (p-SMAD3)的水平来检测TGF-β1信号[gydF4y2Ba8gydF4y2Ba].我们进一步研究了鼻息肉中PC1/3与EMT标记物的关系,发现PC1/3表达与E-cadherin表达的缺失和N-cadherin、collagen I和MMP-2表达的增加呈正相关(gydF4y2Ba图6 bgydF4y2Ba).综上所述,这些结果提示PC1/3可能在鼻息肉EMT中起重要作用。gydF4y2Ba

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
蛋白转化酶(PC)1/3在正常培养的人鼻上皮细胞和鼻息肉中均上调。a)培养的正常人鼻上皮细胞(NHNE)用10 ng·mL孵育gydF4y2Ba−1gydF4y2Ba转化生长因子(TGF)-β1或5 ng·mL组合gydF4y2Ba−1gydF4y2Ba肿瘤坏死因子-α和5 ng·mLgydF4y2Ba−1gydF4y2Ba左图固定NHNE细胞进行E-cadherin免疫细胞荧光染色,右图取细胞提取物进行Western blotting。TGF-β1和cytomix均可改变上皮间质转化(epithelial-mesenchymal transition, EMT)标志物和I型胶原的表达,而PC1/3仅在cytomix组表达升高。p:磷;SMAD:母亲对抗四肢瘫痪的同系物。规模酒吧= 20μm。b)取正常鼻黏膜和鼻息肉组织提取液(50 μg),检测EMT标记物、I型胶原和PC1/3的表达变化。MMP:基质金属蛋白酶。β-肌动蛋白在这两种情况下均作为负载对照。gydF4y2Ba
讨论gydF4y2Ba
上气道黏膜的炎症和重塑过程被认为在鼻息肉的发生中起重要作用[gydF4y2Ba3.gydF4y2Ba].许多涉及重构的蛋白质,如生长因子及其受体和基质金属蛋白酶[gydF4y2Ba9gydF4y2Ba]应该被pc水解成具有生物活性的形式[gydF4y2Ba10gydF4y2Ba].与正常鼻黏膜相比,PC1/3在鼻息肉中表达强烈增加,并诱导气道上皮细胞发生emt样过程。我们还注意到,与正常鼻粘膜相比,鼻息肉中总PC活性显著升高;转化酶furin, PC5/6和PACE4具有较宽的pH最适值,在中性至弱碱性pH值(furin为6.0-8.5,PC5/6为7.0-8.0,PACE4为7.0-8.5)时具有峰值活性,而PC1/3活性范围较窄(5.0-6.5)[gydF4y2Ba21gydF4y2Ba].因此,我们推测,在我们的PC酶测定条件(pH 5.5)下,该酶的大部分活性来自内源性PC1/3。gydF4y2Ba
我们发现PC1/3在位于鼻上皮基底部的神经内分泌细胞中含有chra的分泌颗粒中表达。PC1/3已知主要表达于神经和内分泌细胞,并定位于分泌颗粒[gydF4y2Ba22gydF4y2Ba].已有报道PC1/3及其生理底物,如降钙素基因相关肽、亮氨酸脑啡肽、蚕豆素,在散布于呼吸上皮的神经内分泌细胞中表达[gydF4y2Ba23gydF4y2Ba].此外,JgydF4y2Ba不租gydF4y2Baet al。gydF4y2Ba[gydF4y2Ba15gydF4y2Ba]报道了人鼻部上皮细胞含有脑源性神经营养因子,一种PC1/3底物[gydF4y2Ba24gydF4y2Ba,在神经内分泌细胞中含有chra的分泌颗粒中。令人惊讶的是,PC1/3也在鼻息肉上皮的杯状细胞和纤毛细胞中表达。CgydF4y2BaarolangydF4y2Baet al。gydF4y2Ba[gydF4y2Ba25gydF4y2Ba]的研究表明,神经内分泌细胞的标记物——泛素羧基末端水解酶(UCH)-L1仅在不吸烟者气道上皮的神经内分泌细胞中表达,而吸烟者的UCH-L1在纤毛上皮细胞中也表达上调。根据这些研究,我们认为杯状细胞和纤毛细胞都可以转分化为其他上皮细胞。综上所述,我们推测PC1/3可能通过其产生多种活性肽的能力在鼻息肉的发病机制中发挥重要作用。gydF4y2Ba
在本研究中,我们研究了PC1/3过表达在NCI-H292细胞(PC1/3细胞)中的生物学后果;NCI-H292细胞在培养过程中保持粘液表皮样结构,易于接受基因转移。PC1/3细胞的形态由典型的上皮样转变为细长纺锤形。这一结果与之前的一项发现相吻合,即人类乳腺癌细胞系——密歇根癌症基金会(MCF)的PC1/3过表达细胞(MCF)-7细胞,表现出细胞过程拉长的形态变化,可能是由于PC1/3改变了细胞粘附分子的过程,如整合素、纤维连接素和胶原蛋白[gydF4y2Ba26gydF4y2Ba].我们还证明PC1/3过表达显著增加细胞增殖。由于生长因子如IGF-1、VEGF和血小板源性生长因子在肿瘤细胞生长中起着关键作用[gydF4y2Ba27gydF4y2Ba],有可能这些底物在NCI-H292细胞中被PC1/3加工,导致细胞增殖增强。gydF4y2Ba
细胞迁移在许多生理和病理过程中都很重要,如正常发育、肿瘤侵袭、炎症反应和伤口修复[gydF4y2Ba28gydF4y2Ba].细胞迁移是由改变的局灶黏附动力学与前缘板状足的突出共同介导的,从而产生细胞向前运动[gydF4y2Ba29gydF4y2Ba].我们观察到PC1/3细胞损伤后迁移速度明显加快,与模拟细胞相比,PC1/3细胞表现出更多伸长和突出的垂直肌动蛋白应力纤维以及片状和丝状的伸长。此外,PC1/3细胞上皮标记物下调,间质标记物上调,称为EMT [gydF4y2Ba2gydF4y2Ba,gydF4y2Ba4gydF4y2Ba].最近,年代gydF4y2Ba欣gydF4y2Baet al。gydF4y2Ba[gydF4y2Ba8gydF4y2Ba显示缺氧诱导的EMTgydF4y2Ba通过gydF4y2Basmad3依赖于培养的人鼻上皮细胞和鼻息肉内壁上皮细胞的机制,表明EMT在鼻息肉的发病机制中发挥重要作用。我们的蛋白质组学分析表明,PC1/3细胞和模拟细胞之间产生的差异蛋白涉及碳水化合物代谢、细胞增殖、细胞骨架重组、DNA代谢和信号转导(在线补充图S1和表S2)。有趣的是,在PC1/3细胞中UCH-L1表达上调(在线补充图S1c和表S2)。如前所述,UCH-L1是一种神经内分泌细胞特异性蛋白,用于评估肺癌[gydF4y2Ba25gydF4y2Ba].gydF4y2Ba
在气道上皮损伤后的修复过程中,细胞在细胞前缘积极合成α5整合素[gydF4y2Ba19gydF4y2Ba,gydF4y2Ba20.gydF4y2Ba],需要整合素与MMP-14的功能合作进行细胞迁移[gydF4y2Ba17gydF4y2Ba].这些蛋白是PC底物,激活的MMP-14可以促进前mmp -2的成熟[gydF4y2Ba17gydF4y2Ba].年代gydF4y2BaeomungydF4y2Baet al。gydF4y2Ba[gydF4y2Ba30.gydF4y2Ba]报道过表达MMP-2的人晶状体上皮细胞表现出典型的肌成纤维细胞样表型特征,提示MMP-2在病理性纤维化中发挥作用。在这里,我们发现PC1/3过表达诱导NCI-H292细胞中MMP-2、MMP-14和α5-整合素的表达上调。PC1/3细胞中MMP-3水平升高;乳腺上皮细胞中MMP-3的产生诱导EMT [gydF4y2Ba18gydF4y2Ba].gydF4y2Ba
我们证实,在接受EMT的NHNE细胞中,TNF-α/IL-1β的作用下,PC1/3和twist的表达上调。lgydF4y2Ba我gydF4y2Baet al。gydF4y2Ba[gydF4y2Ba5gydF4y2Ba]报道TNF-α和IL-1β在正常乳腺上皮细胞和乳腺癌细胞中均可诱导EMTgydF4y2Ba通过gydF4y2Ba坏死因子-κ b介导的扭转上调。我们还发现PC1/3的表达与鼻息肉中的EMT密切相关。gydF4y2Ba
总之,我们的数据通过诱导鼻息肉发病机制中emt样过程,为了解鼻息肉上皮细胞表型的可塑性和PC1/3可能的新功能提供了见解。需要进一步研究PC1/3的生理底物,进一步了解PC1/3在鼻息肉发病中的作用。临床上,PC1/3特异性抑制剂或调节剂的开发可能在控制这种病理中找到相关应用。gydF4y2Ba
致谢gydF4y2Ba
我们感谢Iris Lindberg(马里兰大学,巴尔的摩,MD,美国)提供了含有小鼠PC1/3 cDNA和抗小鼠PC1/3抗体的RcCMV表达载体。gydF4y2Ba
脚注gydF4y2Ba
这篇文章有补充资料可从gydF4y2Bawww.www.qdcxjkg.comgydF4y2Ba
支持声明:韩国国家研究基金会基础科学研究计划,由gydF4y2Ba教育部gydF4y2Ba(基金资助:2012-0000803 (J-H.)2011-0026256 (S-N.)李))。gydF4y2Ba
利益冲突:无声明。gydF4y2Ba
- 收到了gydF4y2Ba2012年6月29日。gydF4y2Ba
- 接受gydF4y2Ba2013年1月3日。gydF4y2Ba
- ©2013人队gydF4y2Ba
参考文献gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba










