





文摘gydF4y2Ba
Haemorrhage-induced免疫抑制与院内感染。我们评估的影响monophosphoryl脂质,人数/ interleukin-1 receptor-domain-containing适配器蛋白质诱导interferon-biased Toll样receptor-4受体激动剂目前在人类作为疫苗佐剂,post-haemorrhage对感染的易感性。gydF4y2Ba
我们使用由methicillin-susceptible post-haemorrhage肺炎诱发的小鼠模型gydF4y2Ba金黄色葡萄球菌gydF4y2Ba。Monophosphoryl脂质是管理静脉注射后出血之前,肺炎发病。gydF4y2Ba
出血改变成活率,增加肺损伤(中性粒细胞聚集,水肿和细胞因子释放)和树突和自然杀伤细胞的功能改变。在这里,我们表明,monophosphoryl脂质减少系统性传播gydF4y2Ba金黄色葡萄球菌gydF4y2Ba和抑制炎症性肺部病变。Monophosphoryl脂质部分恢复的能力在树突状细胞抗原表达和转录活动。Monophosphoryl脂质没有恢复interferon-γmRNA但防止白细胞介素- 10”mRNA超表达与未经处理的小鼠相比,自然杀伤细胞。gydF4y2Ba体外gydF4y2Bamonophosphoryl脂质模拟树突细胞和自然杀伤细胞是从出血动物收养转移到老鼠接受post-haemorrhage肺炎。刺激树突细胞(但不是刺激自然杀伤细胞)提高了人们的存活率与老鼠不及时治疗。gydF4y2Ba在活的有机体内gydF4y2Ba消耗的自然杀伤细胞存活率降低monophosphoryl脂质经处理的老鼠。gydF4y2Ba
树突和自然杀伤细胞极度参与的有利影响monophosphoryl脂质在post-haemorrhage肺炎。gydF4y2Ba
文摘gydF4y2Ba
树突细胞和NK细胞极度参与内MPLA的有利影响post-haemorrhage肺炎gydF4y2Bahttp://ow.ly/oc4ZpgydF4y2Ba
介绍gydF4y2Ba
创伤是一个主要的危重患者院内肺炎的危险因素。院内肺炎发展在30 - 50%的创伤患者,主要在第一周,并增加在重症监护室的长度和死亡的风险gydF4y2Ba1gydF4y2Ba]。Methicillin-susceptiblegydF4y2Ba金黄色葡萄球菌gydF4y2Ba(MSSA)创伤后所涉及的主要病原体肺炎(gydF4y2Ba2gydF4y2Ba]。这种创伤后对脓毒症相关的免疫抑制状态持续7 - 10天之后的免疫恢复(gydF4y2Ba3gydF4y2Ba]。树突状细胞(dc)扮演了重要的角色在连接先天和适应性免疫反应的细胞因子的生产,以及抗原的幼稚淋巴细胞(gydF4y2Ba4gydF4y2Ba]。自然杀伤(NK)细胞促进抗菌反应,特别是通过细胞毒性活动对感染细胞和细胞因子的生产(gydF4y2Ba5gydF4y2Ba]。在早期激活NK细胞功能缺陷与老鼠和脑损伤患者受损的抗菌反应(gydF4y2Ba6gydF4y2Ba]。协作的DCs和NK细胞对病原体的控制至关重要的负担完全激活适应性免疫之前(gydF4y2Ba7gydF4y2Ba]。改变DCs之间的串音和NK细胞可能是参与了创伤后对感染的易感性。gydF4y2Ba
toll样受体(通常)是进化的best-characterised受体识别细菌和启动免疫反应。toll样受体激动剂是有前途的治疗预防传染病在人类gydF4y2Ba8gydF4y2Ba]。Monophosphoryl脂质(MPLA)、截脂多糖,是一个人数/ interleukin-1 receptor-domain-containing适配器蛋白质诱导干扰素(TRIF)偏见TLR4受体激动剂是常用的作为疫苗佐剂在人类gydF4y2Ba9gydF4y2Ba]。TLR4 / TRIF途径刺激转录的I型干扰素(IFN),诱导DCs和NK细胞的成熟,白介素(IL) -12,使DCs的交错活化和NK细胞(gydF4y2Ba10gydF4y2Ba]。gydF4y2Ba
我们之前已经证明MPLA恢复IL12p40 CD11c +细胞转录后出血(gydF4y2Ba11gydF4y2Ba]。DCs和NK细胞是两个主要的子集CD11c +细胞,在当前机械的研究中,我们提出的有利影响MPLA可以由DCs或NK细胞的直接刺激。为了验证这个假设,我们评估了MPLA对肺损伤的影响和传统DCs的功能(疾控中心),血浆DCs(髓)和NK细胞。最后,我们进行直流和NK细胞过继转移描绘各自的角色MPLA DCs和NK细胞反应。我们使用的模型,模拟临床场景严重创伤病人呈现出血性休克和后续gydF4y2Ba金黄色葡萄球菌gydF4y2Ba院内肺炎。出血就得以证实复制人类创伤后免疫抑制的主要特点(gydF4y2Ba12gydF4y2Ba]。这个模型允许我们治疗后小鼠出血和肺炎发病和之前复制时照顾创伤患者。gydF4y2Ba
材料和方法gydF4y2Ba
动物保健gydF4y2Ba
实验按照实验动物保健原则(gydF4y2Ba13gydF4y2Ba]。南特大学动物伦理委员会的批准所有动物实验研究。雄性BALB / cJ小鼠(20 - 24 g)从Janvier购买实验室(拉瓦尔、法国)。老鼠维持在12 h光/暗周期与免费的食物和水。gydF4y2Ba
出血过程gydF4y2Ba
全身麻醉后异氟烷(巴克斯特,Maurepas,法国),进行经胸廓的心脏穿刺取三分之一的血液volaemia每10克体重(0.3毫升)(gydF4y2Ba11gydF4y2Ba]。流血量恢复了retro-orbital丛注入90分钟之后。这种交织出血以前已被证明产生可逆的低血压(gydF4y2Ba14gydF4y2Ba]。收缩期动脉压力正常后立即归还的流血量和肺炎发病。心脏穿刺引起的死亡率,如果有的话,发生在第一个30分钟。gydF4y2Ba
肺炎过程gydF4y2Ba
MSSA应变(写明ATCC 29213,溶血素积极和潘顿情人杀白细胞素负),生长了18 h在大豆胰蛋白酶的肉汤培养基37°C,洗两次(1000×gydF4y2BaggydF4y2Ba10分钟37°C),在无菌稀释等渗盐水通过浊度测定法和校准。肺炎过程,transtracheal 24-gauge喂养的插入针用于注射致命剂量的MSSA (7×10gydF4y2Ba6gydF4y2Ba犀牛小鼠CFU)(生存范围肺炎组10 - 30%天7)。Non-necrotising肺炎开发的第一个24小时后气管滴注法(在线补充图S1;肺组织学分析)。gydF4y2Ba
学习小组gydF4y2Ba
小鼠随机分为5组:虚假的(S组),出血(H组),gydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(P组),出血后24 h后gydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(HP组)gydF4y2Ba注射。gydF4y2Ba每个动物注射MPLA(50μg)后立即出血之后24小时gydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(HP-MPLA组)。gydF4y2Ba
评估细菌生长和传播gydF4y2Ba
肺和脾是在无菌条件下机械单一化。器官匀浆受到连续10倍稀释和镀trypticase大豆琼脂和选择性的媒体。gydF4y2Ba
髓过氧物酶测定gydF4y2Ba
肺机械单一化的磷酸钾(50毫米)gydF4y2BaNgydF4y2Ba-ethylmaleimide(10毫米)。匀浆洗两次(12 000×gydF4y2BaggydF4y2Ba30分钟在4°C),悬浮在1毫升的磷酸钾缓冲(50毫米)包含0.5%的十六烷基trimethylammonium,和180年代的用。热休克进行2 h 60°C,和样本离心机在000×12gydF4y2BaggydF4y2Ba10分钟。HgydF4y2Ba2gydF4y2BaOgydF4y2Ba2gydF4y2Ba端依赖氧化gydF4y2BaogydF4y2Ba邻联茴香胺决心通过测量吸光度在460海里。浮在表面的髓过氧化酶(MPO)活动正常化肺重量(gydF4y2Ba11gydF4y2Ba]。gydF4y2Ba
肺血管内皮通透性gydF4y2Ba
老鼠接受了2 mg的腹腔内注射异硫氰酸荧光素(FITC)共轭白蛋白(σ,里昂,法国)。2 h后,肺是收获,机械单一化等渗盐水然后离心机(4000×gydF4y2BaggydF4y2Ba10分钟)。血液收集gydF4y2Ba通过gydF4y2Ba右心室穿刺和离心机(4000×gydF4y2BaggydF4y2Ba10分钟)。FITC-albumin测量在100 -μl上层的整除从肺匀浆和血液荧光测定术在480海里。肺血管内皮通透性计算(如前所述gydF4y2Ba11gydF4y2Ba]。gydF4y2Ba
ELISA肺匀浆的准备gydF4y2Ba
肺样本机械单一化在寒冷的裂解缓冲(PBS, 0.1% Triton x - 100)包含1毫米蛋白酶抑制剂鸡尾酒(σ,巴黎,法国)。匀浆是离心机在000×12gydF4y2BaggydF4y2Ba在4°C为20分钟。测定样品中的蛋白质浓度用BCA蛋白质化验设备根据制造商的指示(美国皮尔斯,罗克福德,IL)。肿瘤坏死因子(TNF) -αIL-1β和巨噬细胞炎症蛋白(MIP) 2浓度与ELISA试剂盒量化根据制造商的指示(研发系统、里尔、法国)。gydF4y2Ba
支气管肺泡灌洗gydF4y2Ba
小鼠安乐死并受支气管肺泡灌洗(BAL) 2毫升无菌PBS。球液体立即被处理量化的上皮细胞,DCs (CD11bCD11c肺DCs)和NK细胞。gydF4y2Ba
脾细胞悬液gydF4y2Ba
脾脏在2 mg·毫升消化gydF4y2Ba−1gydF4y2Ba胶原酶D(罗氏诊断、Meylan、法国)RPMI 1640年补充1%胎牛血清(FCS) 25分钟37°C。添加EDTA(10毫米)的最后5分钟消化。细胞悬液40-μm过滤器过滤,用PBS(离心机在000×12gydF4y2BaggydF4y2Ba10分钟在4°C)。gydF4y2Ba
试剂和抗体gydF4y2Ba
从Invivogen MPLA购买(图卢兹,法国)。单克隆抗体(mAb)用于血细胞计数和/或细胞分选获得eBiosciences(美国圣地亚哥,CA): anti-CD3 (a2) 17日,anti-CD8α(53.6 7),anti-CD11b (M1/70) anti-CD11c (N418) anti-CD45R (B220 RA3-6B2) anti-CD49beta (DX5) anti-CD69 (H1.2F3) anti-CD324(钙粘蛋白,DECMA-1) anti-F4/80 (BM8) anti-siglec-H (eBio440c)和anti-TLR4 (UT41)。Anti-CD40 (3123), anti-CD80 (16-10A1) anti-CD86 (GL1) anti-IAd (II类主要组织相容性复合体(MHC)和ams - 32.1)是获得BD生物科学(Le Pont de Claix、法国)。gydF4y2Ba
表型分析gydF4y2Ba
脾DCs子集被定义为特定的膜标记:CD11c +疾控中心和CD8区分CD8 +和CD8 -疾病预防控制中心;B220 siglec H的髓。肺DCs被定义为CD11c + CD11c + F4/80 -。NK细胞被定性为CD3-CD49beta + CD122 +细胞。直流状态评估使用下面的表面标记:TLR4、CD40, CD80、CD86和I-Ad (MHC II级),而CD69用于NK细胞。细胞内染色后TLR4进行固定和permeabilisation Cytofix / Cytoperm +细胞的(BD生物科学)根据制造商的指示。总共3×10gydF4y2Ba4gydF4y2Ba细胞成熟标志进行分析。数据分析使用FlowJo软件(美国Treestar、亚什兰或)。gydF4y2Ba
实时定量聚合酶链反应gydF4y2Ba
CD8gydF4y2Ba+gydF4y2Ba疾病预防控制中心,CD8gydF4y2Ba- - - - - -gydF4y2Ba疾病预防控制中心和髓样细胞排序进行流式细胞仪咏叹调(BD生物科学)。NK细胞隔离设备II (Miltenyi研究,巴黎,法国)是根据制造商的说明用于没有NK细胞的选择。这些程序通常产生细胞群与纯度高达97%和85%,分别。从排序细胞总RNA分离与试剂盒试剂(表达载体,Cergy Pontoise,法国)和治疗的45分钟37°C 2 U的RQ1 DNase (Promega,里昂,法国)。RNA(1μg)与上标reverse-transcribed三世逆转录酶(表达载体)。的cDNA(1μL)受到RT-qPCR BioRad iCycler智商系统使用QuantiTect SYBR绿色PCR试剂盒(试剂盒、Courtaboeuf、法国)。老鼠引物序列TNF-αIL-12p40, il - 10, IFN-α,IFN-γglyceraldehyde-3-phosphate脱氢酶(GAPDH)给出了在线补充表S1。GAPDH基因表达正常化。2相对基因表达进行了计算gydF4y2Ba−ΔΔgydF4y2BaCt方法使用虚假的组织样本作为校准器样本。gydF4y2Ba
DCs和NK细胞的过继转移gydF4y2Ba
脾脏DCs (CD3、CD49beta CD122 -和CD11c +细胞)和NK细胞(CD3 - CD49beta +和CD122 +细胞)分离得到8到10池作为老鼠的细胞分类进行流式细胞仪咏叹调(BD生物科学)。排序脾DCs的由疾病预防控制中心和髓(在线补充图S2)。孤立的细胞群的纯度> 96%。一夜之间,DCs和NK细胞被刺激与MPLA(10μg·毫升gydF4y2Ba−1gydF4y2Ba),在等渗盐水洗两次解决方案(300×gydF4y2BaggydF4y2Ba10分钟)和静脉注射管理(3×10gydF4y2Ba5gydF4y2Ba细胞/鼠标)。gydF4y2Ba
统计分析gydF4y2Ba
GraphPad棱镜(La Jolla、钙、美国)软件是用于统计分析。连续非参数变量表示为中位数(四分位范围),而利用克鲁斯卡尔-沃利斯测试多个比较;邓恩的测试(gydF4y2Ba事后gydF4y2Ba测试)被用于组间比较和允许我们控制family-wise I型错误率。过继转移实验的生存曲线比较生存率较。Bonferroni调整是用于多个比较。一个假定值< 0.05被认为是具有统计学意义。gydF4y2Ba
结果gydF4y2Ba
MPLA减少MSSA系统性传播和肺部炎性病变gydF4y2Ba
在老鼠身上进行post-haemorrhage肺炎,MPLA发现减少MSSA系统性传播和肺部炎性病变。之前报道的死亡率要高于HP组比P(小鼠接受post-haemorrhage肺炎)组和治疗小鼠(HP-MPLA集团)gydF4y2Ba11gydF4y2Ba]。我们先评估是否MPLA注入改变肺反应post-haemorrhage肺炎。肺和脾收获后24 hgydF4y2Ba金黄色葡萄球菌gydF4y2Ba气管内的滴注法。出血没有修改肺部细菌负荷(gydF4y2Ba图1一个gydF4y2Ba),但增加了系统性传播MSSA (gydF4y2Ba图1 bgydF4y2Ba)。出血加剧了pneumonia-induced中性粒细胞聚集,pneumonia-induced肺内皮损伤和增加IL-1β生产(gydF4y2Ba图1中gydF4y2Ba)。MPLA (HP-MPLA组)治疗显著降低脾脏细菌负荷,MPO活性和肺血管内皮损伤在不改变生产的促炎细胞因子在肺部与小鼠相比不及时治疗(惠普集团)(gydF4y2Ba图1 b-ggydF4y2Ba)。肺炎肺落下帷幕的比例增加了DCs和NK细胞(gydF4y2Ba图1 h和igydF4y2Ba)。既不出血也MPLA改变了pneumonia-induced直流和NK细胞积累。gydF4y2Ba
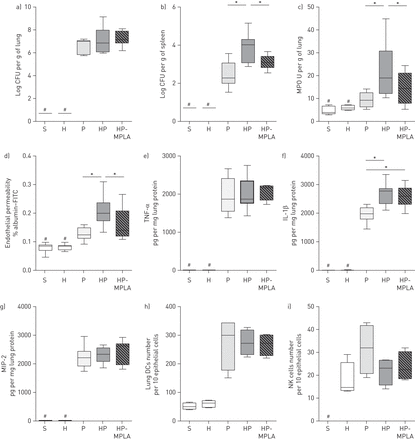
Monophosphoryl脂质(MPLA)下降gydF4y2Ba金黄色葡萄球菌gydF4y2Ba系统性传播和肺部炎性病变小鼠接受post-haemorrhage肺炎。五组的老鼠进行了研究:虚假的(S),出血(H),gydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(P), post-haemorrhagegydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(HP)和接受post-haemorrhage MPLA-treated老鼠gydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(HP-MPLA)。肺和脾收获后24 h气管滴注法。本地和系统性细菌负担在肺部和b)评估脾脏这个方法检出限较低1 CFU / 50μL未稀释的组织匀浆。c)中性粒细胞聚集被测量评估myeloperoxydase活动(MPO)和d)内皮渗透性测量使用albumin-fluorescein异硫氰酸酯(FITC)。浓度的e)肿瘤坏死因子(TNF) -α,f)白介素(IL) 1β和g)巨噬细胞炎性蛋白(MIP) 2评估肺匀浆。h的比例)肺部树突状细胞(dc)和我)自然杀伤(NK)细胞是评估在支气管肺泡灌洗。数据提出了中位数(四分位范围),这一问题从两个独立的实验(n≥5老鼠每组)。gydF4y2Ba#gydF4y2Ba:p < 0.05与p相比,惠普和HP-MPLA组;*:p < 0.05。gydF4y2Ba
MPLA部分恢复TNF-αIFN-α脾DCs和白介素mRNA水平gydF4y2Ba
在老鼠身上进行post-haemorrhage肺炎,MPLA部分恢复TNF-αIFN-α脾DCs和白介素mRNA水平。肺部炎症和白细胞招聘是通过控制调节疾病预防控制中心和pDCs在脓毒症小鼠模型gydF4y2Ba15gydF4y2Ba]或出血[gydF4y2Ba16gydF4y2Ba]。描述了三个主要的子集脾DCs在老鼠身上:CD8 +疾控中心定位自适应免疫反应对炎症反应,生成大量的il - 12;CD8 - cdc展览通过il - 10生产抗炎作用;和髓专攻IFN-α分泌(gydF4y2Ba4gydF4y2Ba]。我们之前显示出血改变生产和co-stimulatory分子细胞因子表达在脾DCs肺炎(gydF4y2Ba11gydF4y2Ba]。我们因此调查MPLA是否恢复细胞因子的转录脾内DCs post-haemorrhage肺炎。脾脏是收获后6 hgydF4y2BaS.aureusgydF4y2Ba气管滴注法(在线补充图S3)。在CD8 +疾控中心(gydF4y2Ba图2 a - cgydF4y2Ba),MPLA治疗(HP-MPLA集团)部分恢复TNF-α和恢复IL-12p40 mRNA水平相比,老鼠不及时治疗(HP组)。MPLA IFN-αmRNA表达增加与其他团体。CD8 -疾病预防控制中心(gydF4y2Ba图2 d-fgydF4y2Ba)、TNF-αil - 10和IFN-αmRNA表达并没有因为MPLA治疗而改变(HP-MPLA组)与post-haemorrhage肺炎(HP组)。在髓(gydF4y2Ba图2 g和hgydF4y2Ba),MPLA治疗并没有改变TNF-α表达但部分恢复IFN-α相比post-haemorrhage感染小鼠不及时治疗(HP组)。gydF4y2Ba
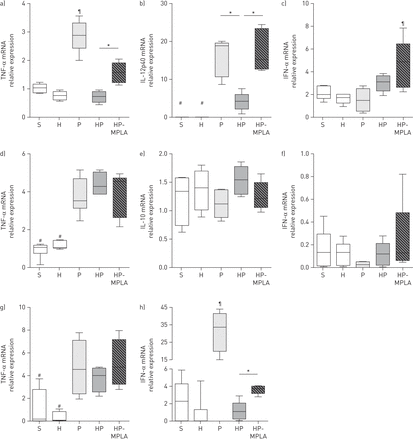
Monophosphoryl脂质(MPLA)部分恢复肿瘤坏死因子(TNF) -α,白介素(IL) -12和干扰素(IFN) -αmRNA水平树突状细胞(dc)在小鼠接受post-haemorrhagegydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎。五组的老鼠进行了研究:虚假的(S),出血(H),gydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(P)、post-haemorrhagegydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(HP)和接受post-haemorrhage MPLA-treated老鼠gydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(HP-MPLA)。脾脏是收获后6 h气管滴注法。)的mRNA水平TNF-α,b) IL-12p40和c) IFN-α评估CD8 +传统DCs(疾控中心);d) TNF-αe) il - 10和f) IFN-α评估CD8 -疾病预防控制中心;和g) TNF-α和h) IFN-α血浆DCs进行评估。数据作为中位数(四分位范围),这一问题从两个独立的实验(n≥6小鼠每组)。gydF4y2Ba#gydF4y2Ba:p < 0.05与p相比,惠普和HP-MPLA组;gydF4y2Ba¶gydF4y2Ba:p < 0.05gydF4y2Ba与gydF4y2Ba其他所有人;*:p < 0.05。gydF4y2Ba
MPLA部分恢复常规和血浆DCs的成熟状态gydF4y2Ba
在老鼠身上进行post-haemorrhage肺炎,MPLA部分恢复常规和血浆DCs的成熟状态。MPLA恢复TNF-αIFN-α和可能的表达,因此,加快dc的成熟。为了获得更多的知识,我们评估的影响MPLA成熟的膜表达标记(CD40、CD86和MHC II级)在DCs。值得注意的是MPLA治疗没有修改的数量脾直流子集与小鼠相比不及时治疗(惠普集团gydF4y2Ba图3 lgydF4y2Ba)。在CD8 +疾控中心,MPLA治疗增加CD86表达和没有影响的表达CD40和MHC II级与小鼠相比不及时治疗(惠普集团)(gydF4y2Ba图3模拟gydF4y2Ba)。CD8 - cdc MPLA治疗增加MHC II级表达和没有影响的表达CD40和CD86 (gydF4y2Ba图3情况gydF4y2Ba)。在髓,MPLA治疗(HP-MPLA组)增加CD86表达但不是CD40和MHC II级表达与小鼠相比不及时治疗(惠普集团)(gydF4y2Ba图3我gydF4y2Ba)。gydF4y2Ba
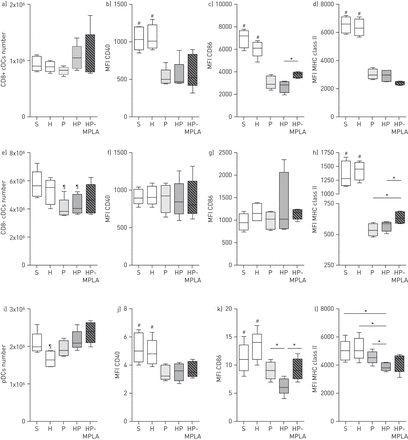
Monophosphoryl脂质(MPLA)改变了骨髓的成熟状态,血浆树突细胞(dc)在小鼠接受post-haemorrhagegydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎。五组的老鼠进行了研究:虚假的(S),出血(H),gydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(P), post-haemorrhagegydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(HP)和接受post-haemorrhage MPLA-treated老鼠gydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(HP-MPLA)。老鼠牺牲后6 h气管滴注法。细胞数量和成熟的标记(CD40、CD86和主要组织相容性复合体(MHC)类II)评估在脾模拟)CD8 +传统DCs(疾控中心),超高频)CD8 - cdc和我)血浆DCs(髓样)。数据作为中位数(四分位范围),这一问题从两个独立的实验(n≥6小鼠每组)。小额信贷机构:平均荧光强度。gydF4y2Ba#gydF4y2Ba:p < 0.05与p相比,惠普和HP-MPLA组;gydF4y2Ba¶gydF4y2Ba:p < 0.05gydF4y2Ba与gydF4y2Ba其他所有人;*:p < 0.05。gydF4y2Ba
MPLA阻止il - 10 mRNA在NK细胞过度gydF4y2Ba
在老鼠身上进行post-haemorrhage肺炎,MPLA阻止il - 10 mRNA在NK细胞过度。NK cell-to-DC相声是非常成熟的DCs在细菌感染(gydF4y2Ba7gydF4y2Ba,gydF4y2Ba10gydF4y2Ba]。我们因此调查NK细胞的功能状态和MPLA是否改变了出血感染小鼠NK细胞的反应。脾脏是收获后24 hgydF4y2Ba金黄色葡萄球菌gydF4y2Ba气管滴注法(在线补充图S3)。脾NK细胞的数量减少小鼠接受post-haemorrhage肺炎(HP组)与受感染的小鼠(P组)相比,没有任何的影响MPLA (HP-MPLA集团gydF4y2Ba图4gydF4y2Ba)。NK细胞表现出更活化表型HP组相比P组由表面CD69的表达(如图所示gydF4y2Ba图4 bgydF4y2Ba)。MPLA治疗没有修改CD69的百分比gydF4y2Ba+gydF4y2BaNK细胞与小鼠相比不及时治疗(HP组,gydF4y2Ba图4 bgydF4y2Ba)。IFN-γ,这主要是由NK细胞(gydF4y2Ba5gydF4y2Ba),是非常参与宿主对病原体的防御。IFN-γmRNA表达强烈降低小鼠接受post-haemorrhage肺炎肺炎相比(P组),而不是恢复MPLA治疗(HP-MPLA集团gydF4y2Ba图4摄氏度gydF4y2Ba)。主要抗炎细胞因子il - 10在描述haemorrhage-induced immunosupression [gydF4y2Ba17gydF4y2Ba]。在这种背景下,主要细胞来源的细胞因子仍然是有争议的。NK细胞是我们发现,il - 10 mRNA水平大幅度增加HP组相比P组和MPLA治疗降低il - 10 mRNA水平与惠普相比集团(gydF4y2Ba图4 dgydF4y2Ba)。gydF4y2Ba
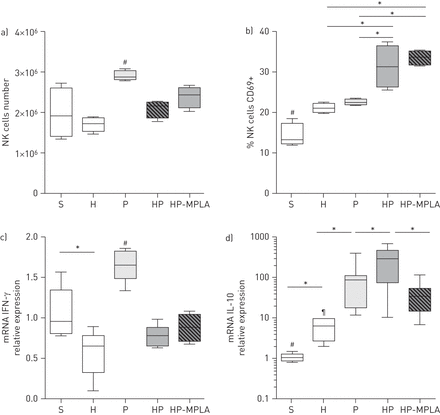
Monophosphoryl脂质(MPLA)减少白介素(IL) -10 mRNA水平自然杀伤(NK)细胞的老鼠接受post-haemorrhagegydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎。五组的老鼠进行了研究:虚假的(S),出血(H),gydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(P)、post-haemorrhagegydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(HP)和接受post-haemorrhage MPLA-treated老鼠gydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(HP-MPLA)。脾脏是收获后24 h气管滴注法。NK细胞数量,b) CD69 +细胞的比例和mRNA水平的c)干扰素(IFN) -γ和d) il - 10在脾NK细胞进行评估。数据作为中位数(四分位范围),这一问题从两个独立的实验(n≥6小鼠每组)。gydF4y2Ba#gydF4y2Ba:p < 0.05相比,所有其他群体;gydF4y2Ba¶gydF4y2Ba:p < 0.05相比,感染组;*:p < 0.05。gydF4y2Ba
MPLA-stimulated DCs增加小鼠的生存经历post-haemorrhage肺炎gydF4y2Ba
我们旨在确定哪些细胞与MPLA直接回应。因此,我们评估了在DCs和NK细胞TLR4的表达。pDCs限制性TLR剧目没有TLR4表达(gydF4y2Ba4gydF4y2Ba]。的膜表达TLR4在疾控中心并没有改变在我们的实验(gydF4y2Ba图5 a和bgydF4y2Ba)。如前所述在NK细胞(gydF4y2Ba18gydF4y2Ba),我们未能发现表面表达TLR4的NK细胞(数据未显示),但细胞TLR4水平降低H和HP组与控制(集团,gydF4y2Ba图5度gydF4y2Ba)。尽管这在NK细胞TLR4表达减少,MPLA提高DCs和NK细胞的功能gydF4y2Ba在活的有机体内gydF4y2Ba。我们因此评估如果MPLA的影响仅限于每个细胞类型的直接刺激这些细胞之间或相声。为了解决这个问题,脾DCs和NK细胞从老鼠资本(gydF4y2Ba图6gydF4y2Ba)。gydF4y2Ba图6罪犯gydF4y2Ba是代表脾DCs和NK细胞的排序策略。DCs和NK细胞培养过夜gydF4y2Ba体外gydF4y2Ba要么有或没有MPLA(10μg·毫升gydF4y2Ba−1gydF4y2Ba)。细胞培养或单独在一起之前gydF4y2Ba注射。gydF4y2Ba管理作为老鼠与此同时MSSA气管滴注法(gydF4y2Ba图6gydF4y2Ba)。存活率是评估超过168 h。与HP组相比,存活率从8%上升到31%,出血老鼠MPLA-stimulated DCs的收件人(HP-stimulated DCs组;p = 0.03)和42%作为老鼠的MPLA-stimulated DCs / NK细胞一起培养(HP-stimulated DCs / NK组;p = 0.02gydF4y2Ba与gydF4y2Ba惠普集团)(gydF4y2Ba图6 egydF4y2Ba)。如果细胞的过继转移和管理MPLA-stimulated NK细胞(HP-stimulated NK组)改变了存活率(gydF4y2Ba图6 egydF4y2Ba)。一起,这些结果表明,DCs参与的直接MPLA-stimulation MPLA治疗的保护作用,而NK细胞的直接刺激TLR4可有可无的出现在这个模型。gydF4y2Ba
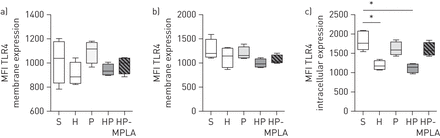
toll样受体(TLR) 4膜表达在传统树突状细胞(cdc)不变,但胞内表达降低自然杀伤(NK)细胞的小鼠出血(H)或post-haemorrhagegydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(HP)。五组的老鼠进行了研究:虚假的(S), H,仅gydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(P),惠普和monophosphoryl脂质与惠普(MPLA)治疗的老鼠。TLR4水平评估的膜)CD8 +疾控中心和b) CD8 -疾病预防控制中心,和c)细胞内TLR4在NK细胞水平进行评估。小额信贷机构:平均荧光强度。*:p < 0.05gydF4y2Ba与gydF4y2BaS组。gydF4y2Ba
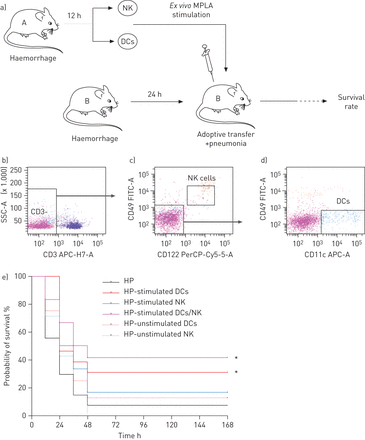
Monophosphoryl脂质(MPLA)刺激树突细胞(dc)小鼠的存活率增加接受post-haemorrhage肺炎。一)实验的示意图。小鼠脾脏收获12 h后出血。脾脏自然杀伤(NK)细胞(CD3 -, CD49β+和CD122 +细胞)和DCs (CD3、CD49β和CD11c +细胞)是排序(罪犯),然后培养有或没有MPLA(10μg·毫升gydF4y2Ba−1gydF4y2Ba一夜之间)。刺激,如果细胞(3×10gydF4y2Ba5gydF4y2Ba每只老鼠)细胞gydF4y2Ba注射。gydF4y2Ba管理作为老鼠,与此同时收到气管内的滴注法gydF4y2Ba金黄色葡萄球菌。gydF4y2Bae)六组的生存率是评估超过168 h: post-haemorrhage肺炎没有过继转移(HP组;n = 18),出血老鼠接收如果DCs(惠普−如果DCs组;n = 8);如果NK细胞(HP−如果NK组;n = 8);MPLA-stimulated DCs(惠普−刺激DCs组;n = 13);MPLA-stimulated NK细胞(HP−刺激NK组;n = 12);MPLA-stimulated DCs / NK细胞培养(HP−刺激DCs / NK组; n=12). Survival rates are expressed as percentage and are representative of two independent experiments. SSC-A: side-scatter analogue; FITC: fluorescein isothiocyanate. * p<0.05与gydF4y2Ba惠普集团。gydF4y2Ba
NK细胞所需的保护作用的MPLA post-haemorrhage肺炎gydF4y2Ba
探讨NK细胞是否可有可无的MPLA的保护作用gydF4y2Ba在活的有机体内gydF4y2Ba期间,我们评估了NK细胞耗竭的影响post-haemorrhage肺炎。NK细胞的损耗,通过执行gydF4y2Ba注射。gydF4y2Ba注入anti-asialo GM1 (anti-ASGM1)(25μL;Wako纯化学物质,日本大阪)24小时前出血过程(gydF4y2Ba图7gydF4y2Ba)。我们下一个评估的存活率MPLA-treated出血老鼠有或没有NK细胞耗竭。存活率下降40% MPLA-treated出血老鼠(HP-MPLA组)在小鼠NK细胞衰竭(0% HP(耗尽NK) mpla组;p = 0.03) (gydF4y2Ba图7 bgydF4y2Ba)。这些结果强调关键作用对肺炎和NK细胞的免疫反应表明,NK细胞参与MPLA的保护作用在这个模型haemorrhage-induced免疫功能障碍。gydF4y2Ba
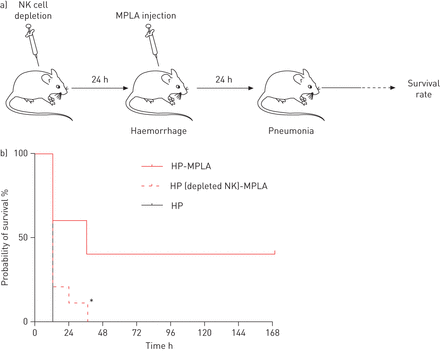
自然杀伤(NK)细胞的损耗的存活率降低monophosphoryl脂质(MPLA)治疗的老鼠接受post-haemorrhagegydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎。NK细胞耗竭,老鼠有一个gydF4y2Ba注射。gydF4y2Ba注入anti-asialo GM1(25μL /鼠标)出血休克前24小时。MPLA是gydF4y2Ba注射。gydF4y2Ba注射后立即出血和24小时后的气管滴注法gydF4y2Ba金黄色葡萄球菌gydF4y2Ba。b)我们评估小鼠的存活率经历post-haemorrhaged methicillin-sensitivegydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎(HP组;n = 10),作为老鼠的对待MPLA (HP-MPLA组;n = 10)或出血的老鼠接受MPLA耗尽后NK细胞(惠普(耗尽NK) MPLA组;n = 10)。数据表示为百分数,代表两个独立的实验。*:p < 0.05gydF4y2Ba与gydF4y2BaHP-MPLA组。gydF4y2Ba
讨论gydF4y2Ba
MPLA的影响在post-haemorrhage肺炎可概括如下:1)MPLA减少系统性传播gydF4y2Ba金黄色葡萄球菌gydF4y2Ba和肺部病变;2)MPLA直流部分恢复功能,防止在NK细胞il - 10 mRNA过度;和3)的直接MPLA-activation DCs减少死亡率,而NK细胞免疫反应并不是可有可无的肺炎。gydF4y2Ba
小鼠的出血休克模型常用于伤害引起免疫抑制,因为它们繁殖的研究其主要特点:TLR刺激后细胞因子的生产降低了循环白细胞(gydF4y2Ba14gydF4y2Ba[],降低HLA-DR表情抗原呈递细胞gydF4y2Ba12gydF4y2Ba在适应性免疫应答[]和改变gydF4y2Ba19gydF4y2Ba]。在不同的小鼠出血模型,Wiggers模型(gydF4y2Ba20.gydF4y2Ba由不可逆休克诱导长期低血压。然而,人类创伤患者死亡的常见原因是快速放血,而不是长期低血压。在当前模型与复苏,醒着的老鼠可能会限制通过激素低血压反应,这是一个主要决定因素对感染的免疫反应。此外,心脏穿刺直接改变了免疫反应(gydF4y2Ba14gydF4y2Ba),忠实地复制伤害引起出血的临床场景比没有组织损伤出血。创伤后肺炎感染(模型),影响多达40%的患者,主要是由MSSA [gydF4y2Ba2gydF4y2Ba]。因此,当前的post-haemorrhage模式gydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎密切模仿临床情况和似乎是有用的临床前评价创伤后免疫调节治疗。gydF4y2Ba
DCs与呼吸道上皮细胞和扩展过程密切相关的腔气道。这些扩展使得DCs的激活细菌通过承认其为使用模式识别受体分子模式。DCs产生细胞因子和趋化因子,调节刺激吞噬细胞的大量涌入,提高肺部细菌清除率。成熟dc迁移到淋巴组织和呈现抗原的淋巴细胞。几天后适应性免疫细胞是招募;他们通过抗体减少系统性细菌负担生产和补体级联激活(gydF4y2Ba21gydF4y2Ba]。我们有报道说,出血并没有改变肺部细菌间隙(白细胞细胞),但增加脾脏细菌负担(自适应免疫反应)。这种矛盾也是sepsis-induced免疫功能紊乱的小鼠模型中描述(gydF4y2Ba15gydF4y2Ba]。几个假说可以解释这种差异地方和系统性细菌清除率。首先,在一个小鼠模型gydF4y2Ba刚地弓形虫gydF4y2Ba从事一个IL-10-mediated感染,NK细胞免疫调节电路降低全身炎症,但增加传播的病原体gydF4y2Ba22gydF4y2Ba]。在当前的模型中,出血诱导il - 10在NK细胞转录的削弱可能增加的系统性传播gydF4y2Ba金黄色葡萄球菌gydF4y2Ba。其次,减少细菌抗原肺b细胞功能(gydF4y2Ba19gydF4y2Ba)或细胞毒性t细胞功能(gydF4y2Ba21gydF4y2Ba创伤后)可以参与细菌传播。最后,post-haemorrhage的系统演化gydF4y2Ba金黄色葡萄球菌gydF4y2Ba肺炎突显出一个适当的协调至关重要的先天和适应性免疫反应。gydF4y2Ba
有效对抗感染,通过膜成熟dc激活适应性免疫分子表达两个家庭:第一个是由分子MHC II级(人类HLA-DR)和第二个由co-stimulatory分子(CD80和CD86分子)。表示没有co-stimulatory分子的抗原导致公差通过t细胞无力或凋亡。在这种背景下,炎症过程,如出血或肺炎,改变dc的抗原呈递能力(gydF4y2Ba23gydF4y2Ba),减少HLA-DR (MHC II级)表达单核细胞和DCs的描述特性的创伤后免疫抑制人类。I型干扰素已成为中央协调员先天和适应性免疫反应的诱导成熟的抗原呈递细胞,参与肺反应细胞外的细菌。TLR4新兵两种不同人数/ interleukin-1 receptor-domain-containing适配器:首先,TLR4 / MyD88通路激活转录因子核转录因子(NF) -κB和促炎细胞因子的转录;其次,TLR4 / TRIF途径导致磷酸化的转录因子干扰素调节因子(IRF) 3,导致I型干扰素的转录gydF4y2Ba24gydF4y2Ba]。NF-κB弱核易位,来响应一个gydF4y2Ba体外gydF4y2BaTLR4 / MyD88通路的活化脂多糖,据报道在创伤患者的循环单核细胞(gydF4y2Ba25gydF4y2Ba]。迄今为止,TLR4的功能/ TRIF通路trauma-haemorrhage后没有被记录。有趣的是,招聘的TRIF TLR4激活参与了肺界限在肺炎的细菌(gydF4y2Ba26gydF4y2Ba]。在目前的模型haemorrhage-induced免疫功能障碍,MPLA, TRIF-biased TLR4受体激动剂(gydF4y2Ba27gydF4y2Ba),增加的转录IRF - (IFN-α)和NF -gydF4y2BaκgydF4y2BaB在DCs (TNF-α和IL-12p40)端依赖细胞因子。gydF4y2Ba
成熟的集群DCs与NK细胞对启动T-cell-mediated至关重要的免疫反应在细菌感染(gydF4y2Ba28gydF4y2Ba]。NK细胞的两个子集描述:1)IFN-γ分泌NK细胞(NK1)也有细胞毒性的活动,使得破坏感染细胞;和2)il - 10分泌NK细胞(NK2)显示监管职能通过antigen-induced t细胞增殖的抑制作用gydF4y2Ba29日gydF4y2Ba]。改变NK细胞的描述在创伤患者,并与肺炎小鼠的脑损伤(gydF4y2Ba6gydF4y2Ba]。MPLA减少il - 10的能力生产这款鼠标模型可能相当重要,il - 10在不断证明有关创伤后免疫抑制是一个关键因素。在危重患者中,il - 10负责单核细胞的脑损伤患者的失活(gydF4y2Ba30.gydF4y2Ba]。在受伤的患者中,il - 10生产和HLA-DR抑制先于院内肺炎(gydF4y2Ba31日gydF4y2Ba]。NK细胞的转录从IFN-γ转向il - 10可能有助于减少DCs成熟期间post-haemorrhage肺炎。gydF4y2Ba
DCs之间的串音和NK细胞的成熟都是至关重要的细胞谱系在细菌感染。DCs的成熟对炎性或耐受性特征取决于直接TLR4活化和细胞间接触的集成IFN-γ分泌NK细胞(gydF4y2Ba10gydF4y2Ba]。NK细胞的激活需要细胞因子(il - 12和IFN-α等)和细胞接触激活DCs (gydF4y2Ba28gydF4y2Ba]。因此,我们报道,MPLA的保护作用需要DCs和NK细胞的作用恢复免疫内稳态出血老鼠。有趣的是,NK细胞的直接刺激TLR4没有改善死亡率在我们的模型中。这些数据符合最近的两项研究显示,NK细胞活性TLR4受体激动剂,减少在sepsis-induced免疫抑制小鼠模型(gydF4y2Ba18gydF4y2Ba)和人类患有败血症或无菌系统性炎症(gydF4y2Ba32gydF4y2Ba]。最后,NK细胞hyporeactivity MPLA可能与haemorrhage-induced胞内TLR4下降。gydF4y2Ba
我们的研究也有一些局限性。首先,我们使用一个小鼠模型,限制了结果的外推给人类。然而,这个模型再现了创伤后免疫抑制的主要免疫功能在人类和报道gydF4y2Ba金黄色葡萄球菌gydF4y2Ba创伤患者的肺炎是最常见的并发症。其次,我们报道了DCs和NK细胞TLR4表达,但我们并没有证明MPLA特定受体。过继转移的gydF4y2Ba体外gydF4y2Ba刺激细胞表明DCs MPLA直接改变,尽管这一事实gydF4y2Ba体外gydF4y2BaNK细胞治疗并没有改变生存,我们不能排除,NK细胞响应MPLA。最后,其他细胞,肺泡巨噬细胞等也可以参与伤害引起的免疫抑制。gydF4y2Ba
结论gydF4y2Ba
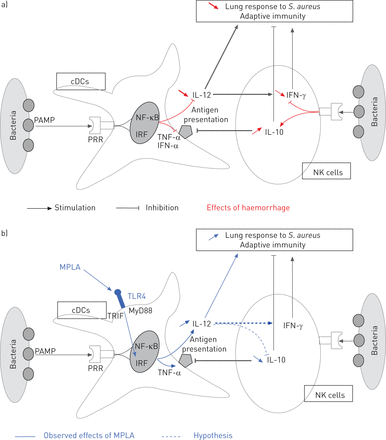
总之,MPLA治疗改善结果的老鼠接受post-haemorrhage肺炎、和疾病预防控制中心的主要功能部分恢复,pDCs和NK细胞。gydF4y2Ba体外gydF4y2Ba刺激的DCs(但不是NK细胞)MPLA改善模型小鼠的存活率。损耗的NK细胞的存活率显著下降MPLA-treated老鼠。总之,这些结果表明,直接MPLA-stimulation DCs诱发NK细胞的活化状态是至关重要的免疫功能不全的主机对抗细菌细胞外(gydF4y2Ba图8gydF4y2Ba)。gydF4y2Ba
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
示意图说明)的出血和b) monophosphoryl脂质在肺炎(MPLA)对免疫反应的影响。承认其为分子模式(PAMP时)模式识别受体(PRR)生成信号,激活树突状细胞(dc)和自然杀伤(NK)细胞。肿瘤坏死因子(TNF) -α和干扰素(IFN) -α提高dc的抗原呈递能力。关于NK细胞,PRR接触刺激抗菌反应(IFN-γ)白介素(IL) -10直流功能下降。一)出血减少抗原表达和细胞因子由DCs生产,和驱动NK细胞对il - 10免疫抑制反应。b) MPLA, TRIF-biased toll样受体(TLR) 4,恢复白介素生产由DCs和防止NK细胞il - 10生产的削弱。最后,MPLA提高肺反应post-haemorrhage肺炎。gydF4y2Ba金黄色葡萄球菌gydF4y2Ba:gydF4y2Ba金黄色葡萄球菌gydF4y2Ba;疾病预防控制中心:传统树突状细胞;NF-κB:核factor-κB;IRF:干扰素调节因子。gydF4y2Ba
脚注gydF4y2Ba
可以从本文的补充材料gydF4y2Bawww.www.qdcxjkg.comgydF4y2Ba
支持声明:这项工作只能由机构基金支持。gydF4y2Ba
利益冲突:没有宣布。gydF4y2Ba
- 收到了gydF4y2Ba2012年9月25日。gydF4y2Ba
- 接受gydF4y2Ba2012年12月20日。gydF4y2Ba
- ©2013人队gydF4y2Ba
引用gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba










