





文摘
背景SARS-CoV-2呼吸系统感染可以进步multi-systemic疾病与异常炎症反应。细胞衰老促进慢性炎症,命名为senescence-associated分泌表型(SASP)。我们调查是否COVID-19疾病与细胞衰老和SASP相关联。
方法尸检肺组织样本11 COVID-19患者和43岁的同龄non-COVID控制SARS-CoV-2通过免疫组织化学方法分析类似的并发症,衰老的标志和关键SASP细胞因子。Virally-induced衰老功能完成在体外通过感染上皮Vero-E6肺泡ⅱ型细胞和一个三维的alveosphere系统(at₂)细胞SARS-CoV-2菌株隔绝COVID-19病人。
结果SARS-CoV-2检测主要通过免疫细胞化学和电子显微镜在at₂细胞。感染at₂细胞表达血管紧张素转换酶2 (ACE2)和衰老增加(p16展出INK4A和SenTraGorTM积极性)和IL-1β和il - 6表达。在体外、感染与SARS-CoV-2 Vero-E6细胞诱导衰老(SenTraGorTM),DNA损伤(γ-H2AX)和增加细胞因子(CXCL8 IL-1β,il - 6)和载脂蛋白B mRNA-editing (APOBEC)酶表达。新一代的基因测速性能分析中得到了感染/细胞衰老Vero-E6证明APOBEC-mediated SARS-CoV-2突变。传播SARS-CoV-2-infection和衰老是证实的肺外网站(肾脏和肝脏)COVID-19病人。
结论我们证明严重COVID-19 at₂细胞感染SARS-CoV-2展览衰老和炎性表型。在体外,SARS-CoV-2感染诱发衰老和炎症。重要的是,被感染的衰老细胞可能充当SARS-CoV-2诱变源由APOBEC酶。因此,SARS-CoV-2-induced衰老可能是一个重要的分子机制严重COVID-19疾病持久性和诱变。
介绍
严重急性呼吸系统综合症冠状病毒2 (SARS-CoV-2) 2019 (COVID-19)导致冠状病毒疾病,主要影响呼吸系统。病人的临床课程范围从无症状到危及生命的呼吸衰竭伴随着multi-systemic炎性疾病(1,2]。通过virally-mediated系统性疾病可能发生炎症反应,由多种促炎细胞因子和趋化因子,包括白介素(IL) 1β,IL - 6、IL - 12, IFN-γ,TNF-α,CXCL-8, CXCL-10 CCL-2 [3,4]。病毒感染的细胞之间的联系和发展严重肺部疾病和系统性表现仍知之甚少。病毒感染导致复杂的先天和适应性免疫反应的激活顺序编排,涉及多种细胞类型和炎症介质(5,6]。在细胞水平上,内在的防御机制被激活和结果从完全恢复到细胞死亡(7- - - - - -11]。一个“中间”被忽视和必要的细胞状态,由于缺乏有效的方法论工具,是细胞衰老12,13]。
细胞衰老是一种应激反应机制,保持体内平衡。衰老细胞的特点是长时间的,一般不可逆细胞循环逮捕和抗细胞凋亡(12,14]。此外,他们还具有分泌功能,集体称为senescence-associated分泌表型(SASP) [12]。SASP包括多种细胞因子,趋化因子,生长因子、蛋白酶和其他分子,这取决于类型的衰老和细胞(12,15]。这些炎症蛋白被释放的可溶性细胞外空间因素,跨膜蛋白ectodomain脱落后,或分子内吞噬细胞外囊泡(16- - - - - -18]。细胞衰老起着关键作用在一些肺部疾病,涉及几个细胞的衰老在肺19,20.]。在生理条件下,衰老和SASP期间短暂地激活介导免疫细胞衰老细胞间隙的招聘。此外,其他SASP因素促进组织再生和修复,全面确保细胞/组织内稳态。相反,持久性的衰老细胞产生有害属性促进组织功能障碍和慢性炎症的维护,通过旁分泌和系统性SASP [12,15]。
几乎没有证据表明病毒感染细胞衰老(发表21- - - - - -26]。鉴于炎症反应的含义COVID-19的恶化和SASP衰老细胞分泌,我们调查是否细胞衰老发生在COVID-19。因为单个标记不足够明确检测衰老,特别是在活的有机体内,因为他们也可能出现在非衰老状况,我们跟着在临床设置详细multi-marker算法,我们和其他人最近发表并被批准的衰老社区(12,27]。我们是第一个为衰老提供证据证明在COVID-19感染肺组织通过应用该算法之前,预印(bioRvix),当前版本的手稿(28),后来已经被其他人证实[29日]。这是最大的临床研究验证viral-induced衰老SARS-CoV-2感染与促炎的表型,这可能导致急性和慢性严重COVID-19临床表现。还重要的是,我们提供的新证据的生成病毒突变促进衰老细胞SARS-CoV-2持久性。
材料和方法
组织
Formalin-fixed和石蜡包埋尸检肺组织样本来自11个病人死于严重COVID-19(经RT-qPCR) (表1和表S1A)。肺实质显示类似病变(atelectasia、纤维化和由免疫细胞浸润)从43个年龄相似的并发症患者从以前公布的队列30.)和新情况下,切除COVID-19爆发前,被用于与COVID-19样本为所有实验详细低于(-控制)(表1和表印地)。non-COVID群60与肺样本被用作验证生物-控制anti-SARS-CoV-2 (G2)单克隆抗体(在线补充)。此外,肾和肝组织的十一COVID-19病例和五non-COVID-19患者可以和分析存在的病毒和细胞衰老。临床样本收集和实验使用委员会批准Cantonale D 'ethique de la矫揉造作的,洛桑大学、瑞士(2020 - 01257)和生命伦理委员会雅典大学医学院,希腊。
细胞和SARS-CoV-2文化
SARS-CoV-2(孤立30 - 287,B.1.222和(加入没有责任压力MT459880.1)是通过文化州立E6细胞(写明ATCC®crl - 1586),从两个不同的感染(COVID-19)患者。病毒从鼻咽拭子中恢复,冲洗1 mLsaline通过0.22纳米过滤器和过滤两次。股票是由病毒感染完全汇合的Vero-E6细胞在DMEM, 10%胎牛血清的边后卫用抗生素,在37°C和5%的公司2。接种病毒股票收集四天后,测序由门店(在线补充)和上层清液被冻结(−80°C),直到使用。感染进行了24-well盘子,使用SARS-CoV-2 0.01莫伊。细胞被固定用4%多聚甲醛或细胞溶解与NucleoZOL (Macherey-Nagel)感染后的17天。政府的具体ataxia-telangiectasia-mutated (ATM)蛋白激酶抑制剂ku - 55933进行了描述(其他地方31日]。三个独立的实验进行。所有操作都进行三级生物安全水平的实验室设施。
SARS-CoV-2主要人类肺alveosphere系统
福尔马林固定和石蜡包埋部分从三维alveosphere系统感染SARS-CoV-2和相应的控制(未受感染的)进行分析。alveospheres包括自我更新at₂细胞表达的at₂细胞标记,htii - 280和表面活性剂蛋白质C以及一些细胞表达AT1细胞标记HTI 56 [32]。Alveospheres轻轻打开使用移液允许感染,避免细胞应激(32]。
Anti-SARS-COV-2 (G2)抗体的一代
小鼠免疫和抗体集合,选择和特异性决心在网上所描述的补充。杂种细胞的转录组分析和氨基酸的测定选择克隆也提供。四个克隆(479 - s1, 480 - s2, 481 - 482 s3和s4)正在专利申请(Gorgoulis VG, Vassilakos D, Kastrinakis n GR-patent应用没有:22 - 0003846810)。
RNA提取和实时逆转录聚合酶链反应(RT-qPCR)检测进行如前所述(在线补充)33]。
下一代测序(门店)
如前所述执行门店(在线补充)34]。
蛋白分离和免疫印迹分析
蛋白质提取和免疫印迹分析所描述的其他地方(31日]。详细信息请参阅在线补充。辣根过氧化物酶抗体结合,anti-mouse anti-rabbit二级抗体(1:1000稀释)(细胞信号)。主要抗体:anti-APOBEC(载脂蛋白B信使rna编辑酶,催化polypeptide-like) 3 g / A3G (Abcam ab109727和ab172694), anti-APOBEC3H (LSBio LS-C151868)和anti-GAPDH(细胞信号)。
免疫细胞化学、免疫组织化学
免疫细胞化学、免疫组织化学进行根据出版协议(34]。一夜之间,以下主要抗体被应用在4°C: anti-SARS-CoV-2 (G2)单克隆抗体(稀释施用)anti-SARS-CoV-2单克隆抗体(1 a9克隆,Genetek) anti-ACE-2 (Abcam),甲状腺转录因子1 (TTF) (Dako) anti-Surfactant蛋白B (SP-B) (1 b9克隆,泽塔公司),anti-p16INK4A(Santa Cruz), anti-IL-1β(Abcam) anti-IL-6(研发系统),anti-phospho-histone (Ser139年)H2AX(γΗ2ΑΧ)(细胞信号),anti-Ki67 (Abcam)和anti-p21Waf1 / Cip1(细胞信号)。
Senescence-SenTraGorTM(GL13)方法
SenTraGorTMGL13化合物(商标)染色法和双染色实验和评估如前所述27,35]。SenTraGor积极的肺泡细胞的平均百分比至少10大功率领域(×400)每个病人是量化。
电子显微镜
代表地区的苏木精和eosin-stained石蜡部分肺解剖COVID-19患者和非COVID-19控制选择在光学显微镜下明显。石蜡包埋组织deparaffinized,水化和固定在2.5%戊二醛在PBS的24小时和你找找1%水锇。
生物信息学分析识别SARS-CoV-2基因组的突变特征
详细描述在线补充,参加APOBEC脱氨基作用C→U SARS-CoV-2基因组的突变替换菌株中可用GISAID数据库(36,37),在细胞培养后获得感染,是检查和验证实验验证APOBEC主题(图S7)。
统计分析
数据表示为±sd。与未配对比较进行非参数Mann-Whitney U测试(比较两组)或Kruskal沃利斯测试其次是邓恩的因果分析(三组间的比较)。Wilcoxon成对非参数测试是用来比较值受感染的细胞在体外。
结果
检测SARS-CoV-2肺细胞
为了检测肺组织中的SARS-CoV-2我们发达单克隆抗体反应与受体结合域(RBD)的突起蛋白SARS-CoV-2和确定了高亲和力抗体(G2),这是最近的有效性验证(38)(图S1,表S1)。SARS-CoV-2检测(使用我们的“房子”G2克隆和商用抗体从GeneTek)主要在2型(at₂)肺泡细胞,由TTF-1识别和表面活性剂蛋白B (SP-B)积极性,在稀疏的炎症细胞(肺泡巨噬细胞和组织)在所有COVID-19病人,从< 5细胞/ 4毫米2组织> 50细胞/ 4毫米2组织(图1一个,表S1A)。表面上皮细胞在小外围航空染色也积极的在某些情况下(图S2)。偶尔SARS-CoV-2 at₂感染细胞大而出现孤立(剥蚀或合胞体)或集群(增生),展示各种拓扑分布(图1一个)。这些细胞中的血管紧张素转换酶2 (ACE2)受体(图1 b),支持SARS-CoV-2感染ACE2受体介导的(39]。此外,电子显微镜分析代表COVID-19确诊病例at₂细胞内病毒的存在(图1 c)和高放大显示病毒粒子在内质网的距离,来显示他们可能装配和出芽,以及居住在胞质囊泡病毒粒子,这意味着他们的转移和释放到细胞外空间。
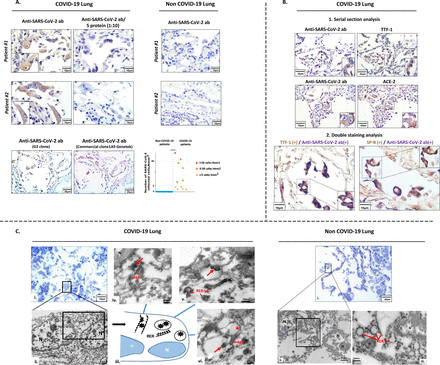
检测SARS-CoV-2从病人死于COVID-19肺细胞。SARS-CoV-2。代表图像(G2 mab)免疫组织化学(包含IHC)染色COVID-19尸检肺组织。与anti-peptide竞争(S蛋白)显示G2免疫染色的特异性。验证的G2 immunopositivity商业anti-S抗体(克隆1 a9克隆,Genetek)。代表消极控制免疫染色non-COVID-19肺组织。图显示了肺部SARS-CoV-2染色样品的量化。b。检测SARS-CoV-2 at₂细胞(证实了积极TTF-1和表面活性剂蛋白b (SP-B)染色)和ACE2-expressing细胞。双免疫染色显示SARS-CoV-2和TTF-1。c .检测SARS-CoV-2通过透射电子显微镜(EM)代表COVID-19病人。存在SARS-CoV-2 at₂细胞内的病毒粒子(i, ii)和邻近的内质网(iii, iv)以及胞质囊泡(三世,第5)。 Corresponding scale bars are depicted. AT2: alveolar type 2 cells; ACE2: Angiotensin-Converting Enzyme 2; ****Statistical significance: p<0.0001.
SARS-CoV-2感染细胞的衰老
SARS-CoV-2 at₂感染细胞的比例(8 - 21%范围)显示衰老表型,为阳性染色SenTraGor™和p16INK4A(图2一个- c,图S3)[12,29日,30.]。SenTraGor™的标志检测衰老细胞的衰老,有独特的属性在任何设置,包括档案材料[formalin-fixed石蜡包埋(FFPE)组织)][12,27,35]。相比之下,肺组织出现纤维化,atelectasia和炎症浸润的同龄non-COVID-19例类似的并发症(表1和表S1),包括一个单独的控制群患者急性肺炎(吸入性肺炎),显示显著降低衰老(范围1 - 2%,p < 0.0001) (图2一个- c,图S4),这表明SARS-CoV-2感染可能引起衰老。
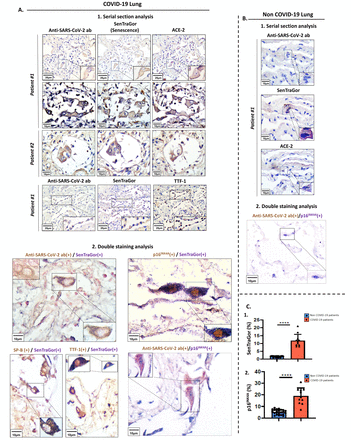
SARS-CoV-2感染细胞的衰老。SARS-CoV-2。代表图像,SenTraGor™(衰老)和ACE-2染色COVID-19串行部分的肺组织(1),SARS-CoV-2 Double-immunostaining分析SenTraGor™(衰老),p16INK4A,TTF-1和表面活性剂蛋白B (SP-B) COVID-19肺组织(2)B。代表SARS-CoV-2连续染色结果,SenTraGor™(衰老)和ACE-2 double-staining SARS-CoV-2和p16INK4A在non-COVID-19肺部组织。c .图形显示的表达增加SenTraGor™和p16INK4A在COVID-19肺部组织。相应规模的酒吧进行描述。* * * *统计学意义,p < 0.0001。
为了证明SARS-CoV-2可以诱导细胞衰老,我们维洛细胞感染病毒毒株COVID-19病人隔离。Vero-E6细胞病毒传播和研究是一个建立了蜂窝系统,除了他们的高传染性SARS-CoV-2他们为数不多的细胞系展示SARS-CoV-2-mediated细胞病变效应,在诊断的一个重要方面(40,41]。感染是在低感染复数(MOI)模仿自然的冠状病毒感染(42]。符合我们的假设,浪潮过去感染细胞后,细胞死亡达成平衡展示清楚衰老(p21 SenTraGor积极性增加,增加的证据WAF1 / CIP1和减少ki - 67),而未受控制细胞在感染后的17天(图3和图S5A)。维洛细胞p16缺乏INK4A(43),最可能引发衰老的DNA损伤,如前所报道(12,13]。DNA损伤衡量γ-H2AX疣状明显在SARS-CoV-2感染细胞(图3一4)。治疗感染细胞的选择性ATM抑制剂(ku - 55933)显著下降衰老评估SenTraGor染色(图S6)。ATM之前已经报道过的关键驱动因素NF-κB-dependent DNA-damage-induced衰老(44]。看来,基因毒性压力强加的结果从一个恶性循环病毒在宿主细胞劫持大多数细胞内蛋白质机械(11,45]。同样,细胞衰老被确认感染肺泡细胞/ alveospheres建立主肺肺泡的三维模型,证实了在活的有机体内观察和研究结果在维洛细胞(图3 b,图S5B)。
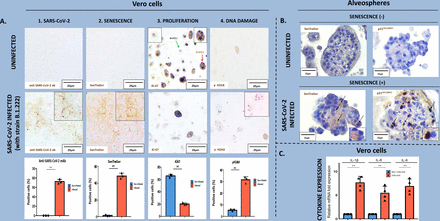
SARS-CoV-2在维洛细胞和alveospheres诱导衰老。衰老。SARS-CoV-2存在(1)感应(2)、细胞增殖(3)和DNA损伤激活(4),与相应的量化直方图,州立与没有SARS-CoV-2感染细胞(感染后的17天)。b。衰老感应(SenTraGor评估和p21WAF1 / CIP1肺泡细胞染色)(alveospheres)主肺肺泡SARS-CoV-2感染后三维模型(黑色箭头)相比,非衰老的(黄色箭头)。缺乏在非感染肺泡细胞衰老显然是明显。c图显示感应SASP相关细胞因子的州立SARS-CoV-2感染后细胞(mRNA expression-normalizedβ2-microglobulin mRNA表达)* *统计学意义:p < 0.01。
Senescence-associated分泌表型
我们发现很高IL-1β和il - 6的表达衰老at₂COVID-19患者的肺细胞,而在非COVID-19控制情况下,包括急性肺炎患者样本,表达很低的少数细胞衰老at₂检测肺组织纤维化,atelectasia和炎症浸润(p < 0.0001) (图4一- c,表S1)。炎症反应的细胞因子都是关键的组件SARS-CoV-2,我们的发现可能意味着这种模式可能是由于炎症细胞衰老的结果SASP COVID-19病人。同样,SARS-CoV-2衰老维洛细胞显示SASP-related细胞因子的表达,作为衰老的评估我们的算法评估,支持我们在活的有机体内发现(图3 c)。
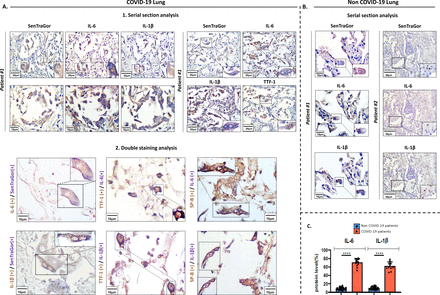
衰老相关的分泌表型(SASP) COVID-19肺组织。染色结果。代表SenTraGor(在低和高放大倍数)TM、il - 6、IL-1βTTF-1和/或表面活性剂蛋白B (SP-B)在相应的串行部分(1)和双免疫染色分析(2)COVID-19肺部组织。原来放大:400×。b。代表染色结果显示缺失或最低级别的SenTraGor™, il - 6和IL-1β年龄non-COVID-19控制样本。相应规模的酒吧进行描述。il - 1β:白介素- 1β;il - 6:白细胞介素- 6;* * * *统计学意义,p < 0.001。
非典CoV-2额外肺传播
SARS-CoV-2免疫反应性另外发现通过应用我们的“房子”G2克隆和商业从GeneTek串行部分可用的肾脏和肝脏组织从一个COVID-19病人(图5一个)。明显的胞质信号明显的肾小管以及肝实质的领域。相比之下,在组织non-COVID-19缺席情况下,信号,如预期(图5一个)。有趣的是,通过连续切片分析,区域窝藏病毒也表现出衰老(SenTraGor积极性),暗示发生SARS-CoV-2诱导衰老的肺网站(图5 b)。
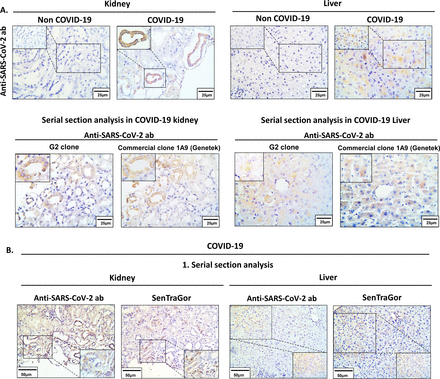
SARS-CoV-2传播的肺网站。染色的结果。代表SARS-CoV-2 COVID-19病人的肾脏和肝脏。缺乏相应的组织的免疫组织化学信号明显代表非COVID-19情况。b。连续切片分析代表COVID-19情况(参见面板)描述并发SARS-CoV-2积极性和衰老标记(SenTraGor™)在肝脏和肾脏的细胞。
受感染的衰老细胞和SARS-CoV-2诱变
最近,我们推测,衰老细胞可能代表SARS-CoV-2准物种来源的一代(46]。这种假设是基于事实,衰老细胞的凋亡宽容性质允许病毒被托管更长时间比其他细胞营业额较高的细胞(47,48),从而呈现SARS-CoV-2 virome更容易主机介导编辑。APOBEC酶是众所周知的参与病毒基因组编辑(12,33,49]。值得注意的是,我们最近显示这些酶在衰老细胞中表达的高度49]。
更新我们最初的生物信息学研究[46),我们筛选∼4500000 SARS-CoV-2菌株GISAID数据库(36,37]。这种分析主要表明SARS-CoV-2变异的主要特征是APOBEC-mediated (图6),符合最近的文献[50]。来验证我们的假设,我们进行细胞培养感染两种隔离病人和获得的后代从衰老细胞(图6 b)。下一代测序(上天)分析表明,收集到的病毒基因组获得突变。重要的是,主要的模式是APOBEC-driven,协议的生物信息学分析GISAID数据库。此外,符合我们的假设和先前的研究[49],SARS-CoV-2诱导衰老细胞表现出更高的信使RNA和蛋白质含量APOBEC酶,尤其是G和H (RNA编辑细胞质变种),据报道,发挥关键作用的病毒RNA编辑(12,33,49]。其他突变签名也发现在细胞培养分离virome基因组的菌株GISAID,也许氧化应激的结果出现在衰老细胞(51]。
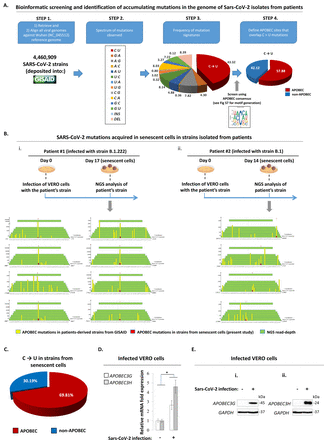
感染衰老细胞作为一个公认的来源SARS-CoV-2准种变体的一代。。原理图的布局呈现APOBEC-mediated突变的识别在4672296 SARS-CoV-2菌株的基因组GISAID数据库。算法评估看到图S7在材料与方法和相应的部分。b。SARS-CoV-2突变的识别在衰老细胞在两株(我:B.1.222和ii:责任)分离出两种不同的患者中,感染后细胞培养的时期。黄色栏显示C→U替换的频率在GISAID数据库捷读计数(绿色限速)和红酒吧描述C→你观察到的突变频率,感染后细胞在当前的研究中,在两个雇佣SARS-CoV-2菌株的基因组。这些网站是排名最高的相对C→U算得上观察到的GISAID数据库。每个C→U基因组序列侧翼也提出,而他们的坐标相对于叠加SARS-CoV-2基因组所示。门店(下一代测序)读取确认一式三份。
讨论
我们演示了存在SARS-CoV-2 at₂细胞的患者死于COVID-19使用我们的小说,一个商业anti-SARS-CoV-2抗体对其峰值蛋白质和电子显微镜。我们展示了第一次后,衰老检测算法在活的有机体内的比例SARS-CoV-2-infected at₂细胞衰老细胞表现出的特性(如通过显著增加染色与小说衰老标记SenTraGor™和p16INK4A)[12,27,35]。在这方面,我们的调查包括连续切片分析和co-staining首次展示在活的有机体内哪些细胞真正衰老和感染。发现在年龄non-COVID-19控制,包括肺组织样本一群患有急性肺炎,衰老细胞的比例要低得多(1 - 2%)比COVID-19病人(8 - 21%),强烈表明SARS-CoV-2触发衰老。确认SARS-CoV-2诱导细胞衰老每se,我们证明了SARS-CoV-2病毒感染上皮细胞(B.1.222应变)在体外增加SenTraGor™染色和诱导的DNA损伤,以增加γ-H2AX表达式。重要的是,我们发现抑制ATM (ataxia-telangiectasia-mutated蛋白激酶),DNA损伤反应途径的顶端协调器,大大减少了衰老在感染样本,表明SARS-CoV-2诱导衰老由DNA损伤和激活介导的DNA损伤反应通路。符合我们的在活的有机体内和在体外发现,SARS-CoV-2诱导衰老被确认在初选之前建立肺肺泡(三维模型32]。alveospheres这一事实进行了温和的机械操作不太可能是导致衰老的发生,鉴于此类干预措施经常应用于3 d瀑样系统和没有报告衰老表型也明显从感染后细胞增殖率和文化52]。
我们还表明,高表达的细胞感染SARS-CoV-2展览IL-1β和il - 6,这两个组件SASP和涉及严重的系统性特征COVID-19 [3,4]。因此,我们的在体外和在活的有机体内研究结果表明,SARS-CoV-2高度at₂细胞通过ACE2感染这些细胞和DNA损伤反应的激活信号,诱导细胞衰老和相关的促炎的表现型(SASP)。根据我们的观察,李et al。最近发现,SARS-CoV-2-induced衰老表现出增强γ-H2AX DNA损伤病灶废除的活性氧(ROS) (NAC)[清道夫防治作用29日),暗示暗示ROS和氧化应激(图S8)。此外,感染细胞显示SASP-positive衰老由p53和激活注册会计师/刺痛信号通路(29日]。他们也证明了senolytics减少COVID-19肺部疾病和炎症感染的仓鼠和老鼠29日),进一步表明SASP作为病毒诱导衰老的结果。Camellet al。还报导说,治疗senolytics老老鼠感染SARS-CoV-2-related鼠标β-coronavirus减少炎症(53]。基于我们的研究结果和最近的证据一个潜在的机械的场景可能是以下(图S8):在入口处,病毒劫持某些能源消费函数与RNA加工、翻译和ER (11]。因此,发生能量转移导致活性氧产量的增加,氧化应激和DNA损伤/ DNA损伤反应(DDR)途径激活。此外,DDR和随后的细胞周期阻滞可能由一个交互的冠状病毒nsp13蛋白质和DNA聚合酶δ(54)和SASP通过注册会计师/刺痛和其他DNA损伤相关的通路(图S8)。因此,我们以前的工作表明,DNA损伤/ DDR是衰老的有效的诱导物,表明其重要性建立病毒诱导衰老表型(31日]。
衰老细胞处于细胞周期阻滞,但保持新陈代谢活跃,分泌炎症蛋白质称为SASP的典型剖面。SASP组件包括促炎细胞因子IL-1β和il - 6,升高血浆的COVID-19病人急性呼吸窘迫综合征(ARDS)或系统性炎症特征。我们在COVID-19表明,衰老改变at₂的属性和功能的细胞,产生细胞因子,可以释放到体循环,放大和延续慢性炎症。当然,组织样本的可用性和更精致的COVID-19的缺失在体外和在活的有机体内模型很难得出可靠的结论。很可能SARS-CoV-2利差从上皮细胞下呼吸道感染at₂肺泡壁细胞,ACE2表达,导致当地的衰老和炎症通过SASP肺。符合这个概念,它之前已经报道过了在体外细胞衰老也作为一个天然抗病毒防御机制和SASP充当这个反应的主要因素,激活和招募免疫系统清除感染(21,22]。随后,其他细胞隔间(干/祖细胞,内皮细胞和炎症细胞)可能会受到影响通过不同的细胞和non-cell自治机制,导致异常的免疫反应,慢性炎症、组织功能障碍和/或纤维化,最终肺损伤和失败(26,55]。特别是,在干细胞/祖细胞衰老会损害肺再生能力。免疫衰老导致高架neutrophils-to-lymphocytes比率(NLR)和il - 6生产。此外,il - 1、il - 6、CXCL8 TNF-α可能是至关重要的维持慢性炎症和TGF-βPAI-1,基质金属蛋白酶可能支持纤维化通过纤维化的基因的表达(ACTA2, COL1A1、COL1A2 FN1)在周围的微环境。病毒可能会进入血液循环并衰老可能随后系统传播,影响其他器官(图5),导致多器官衰竭/ multi-morbidity和死亡在急性期(1,25),或导致急性后遗症COVID-19 (PASC或long-COVID),长期并发症的发展综合症(26,56]。通过这种方式,衰老也会使疾病严重程度根据SARS-CoV-2再次感染或由其他病毒感染,甚至产生负面影响疫苗效果,类似于岁所提出了个人(26]。
额外的含义与长期生存的衰老细胞感染病毒,因为衰老细胞抗凋亡和间隙efferocytosis [12,14]。这种情况下可以提供一段时间“窗口”病毒复制,因此暴露其基因组host-mediated编辑。沿着这个脉络我们表明,衰老细胞间充当“肥沃”环境突变病毒的进化,因为它是更容易APOBEC-3G和3 h RNA编辑(图6)[33,49]。当然,其他机制延长生存和复制SARS-CoV-2可能发生。冠状病毒代码的一种重要的多功能酶称为papain-like蛋白酶(PLP)产生内在deubiquitinating和deISGylating活动。后者与干扰素刺激基因(研究小组)上调和可以用来对抗宿主的免疫反应,否则阻碍感染(57]。以这种方式增加,长期病毒基因组的复制和接触宿主介导的编辑和准物种生成也可能发生。考虑到干扰素衰老抗病诱导剂,情况变得更加复杂,呈现难以区分IGS-mediated从涉及virally-induced衰老机制(58]。当然,还需要更多的研究和指导算法的方法在活的有机体内衰老评估,然后在当前工作,预计澄清在未来很多悬而未决的问题在临床设置(12,27]。值得注意的是,在最近确认买卖(B.1.1.529) SARS-CoV-2变体/应变APOBEC签名被确认为主要突变概要文件(图S9)。为了更好的理解这个角色的APOBEC代病毒突变,抑制APOBEC蛋白质的RNA干扰将是有价值的。然而,这是非常具有挑战性的,因为APOBEC抑制诱发DNA损伤和tslp细胞应激诱导死亡(59- - - - - -61年]。有趣的,并且符合我们的概念连接细胞衰老与病毒突变,通过进行生物信息学分析,我们发现病毒绕过细胞衰老(致癌)表现出显著降低突变负担相比,病毒诱导衰老(图S10)。鉴于突变会影响疫苗的有效性,我们的发现可能意味着病毒诱导衰老在疫苗接种战略的潜在含义。进一步的研究需要对这个根。
尽管到现在为止这是最大的临床研究证明virally-induced SARS-CoV-2衰老,限制我们的研究是相对比较小的样本的大小COVID-19肺尸检,由于难以访问这种稀有材料,鉴于尸检是有限的,只在初始阶段的进行COVID-19爆发为了确定这个新实体的病理基础。另一个限制由于疾病,病理的本质特性,如肺细胞衰老,只能在惨白的调查材料,代表最严重的结果COVID-19谱的临床表现。因此,评价衰老的严重状况目前不是可行的。此外,在我们的研究中,我们研究了18例吸入性肺炎,显示相似度衰老到其他未受控制的情况下。这将是有趣的扩展这些研究其他类型的肺炎。然而,材料从患者急性但不是COVID-19相关的炎症反应是罕见的,因为这些病人通常恢复后罕见病例的治疗和致命的尸检结果很少执行。
总的来说,SARS-CoV-2诱导衰老证明的治疗应用senotherapeutics COVID-19患者的治疗和可能long-COVID综合症(56]。最近,槲皮素测试与标准治疗两个随机临床试验(ClinicalTrials.gov标识符NCT04578158和NCT04861298)患者确诊SARS-CoV-2感染和轻度COVID-19-associated症状。在这两个试验,senolytic干预导致临床改善(62年,63年]。总的来说,这些数据暗示senolytics的有益作用在显著改善结果SARS-CoV-2感染。
确认
我们要感谢Konstantinos Ntostoglou宝贵的帮助在准备材料。我们承认在RNA序列支持希腊个性化医学研究基础设施(pMED-GR) (MIS 5002802),这是实现作用下研究和创新基础设施的“强化”,由运营计划“竞争力、创业和创新”(NSRF 2014 - 2020)和希腊和欧盟联合(欧洲区域发展基金)。这项工作是支持的:国家公共投资项目的发展和投资/总秘书处研究和技术框架的旗舰计划解决SARS-CoV-2ΣΕ01300001 (2020);地平线2020玛丽·斯卡洛多斯卡·居里培训项目没有。722729 (SYNTRAIN);福利社会和文化科学基础,雅典,希腊(KIKPE);h·帕帕斯捐赠;希腊的基础研究和创新(回报)资助。775年和3782年,格兰特NKUA-SARG 70/3/8916。KP是财政支持的国家心脏和肺研究所——伦敦帝国理工学院和英国心脏基金会项目资助(PG / 19/75/34686)。
脚注
作者的贡献:动向,D。V。:immunocytochemistry, SenTraGorTM染色;动向,P。G。F。,c。,L。Dl,V.G.G.:histopathological examination and evaluation of the stainings; M.D., I.K.: cell culture and viral manipulations; B.S.: 3D alveosphere system and viral manipulations; A.K., An.P.: RNA and protein extraction, RT-qPCR, NGS, western blot; N.L., S.H.: electron microscopy; O.H., Ai.P.: Statistical analysis, bioinformatic analysis and graphs generation; D.Va.: anti-SARS-CoV-2 antibody generation; K.E., A.G.T, K.P., S.T., V.G.G.: data analysis and interpretation, manuscript preparation; S.T., Ar. P., G.B., P.J.B., V.G.G.: experimental design, guidance, manuscript writing with input for all co-authors.
资助:这项工作是支持的:国家公共投资项目的发展和投资/总秘书处研究和技术框架的旗舰计划解决SARS-CoV-2ΣΕ01300001 (2020);地平线2020玛丽·斯卡洛多斯卡·居里培训项目没有。722729 (SYNTRAIN);福利社会和文化科学基础,雅典,希腊(KIKPE);h·帕帕斯捐赠;希腊的基础研究和创新(回报)资助。775年和3782年,格兰特NKUA-SARG 70/3/8916。国家公共投资项目的发展和投资/总秘书处研究和技术框架的地址SARS-CoV-2旗舰倡议;格兰特:2020ΣΕ01300001;H2020玛丽Skłodowska-Curie行动; DOI: http://dx.doi.org/10.13039/100010665; Grant: 722729 (SYNTRAIN); Welfare Foundation for Social & Cultural Sciences, Athens, Greece (KIKPE); Hellenic Foundation for Research and Innovation; DOI: http://dx.doi.org/10.13039/501100013209; Grant: 3782, 70/3/8916, 775, NKUA-SARG grant; H. Pappas.
利益冲突:PB获得研究经费从阿斯利康和勃林格殷格翰集团和阿斯利康的科学顾问,勃林格殷格翰的发言,Epi-Endo,诺华,地区和梯瓦。其他作者希望宣布无利益冲突。
- 收到了2021年8月27日。
- 接受2021年12月24日。
- 版权©2022年作者。
这个版本分布在创作共用署名非商业性许可证的条款4.0。商业生殖权利和权限接触权限在}{ersnet.org











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}