





文摘gydF4y2Ba
背景gydF4y2Ba阻塞性睡眠呼吸暂停(OSA)导致间歇性缺氧,进而诱发内皮功能障碍和动脉粥样硬化进展。我们提出VE-cadherin乳沟,血液中检测到其细胞外释放片段水溶性(sVE),可能是一个紧急内皮通透性异常的早期指标。我们的目的是评估VE-cadherin乳沟在阻塞性睡眠呼吸暂停综合症的病人gydF4y2Ba在活的有机体内gydF4y2Ba和gydF4y2Ba在体外gydF4y2Ba间歇性缺氧模型来解释细胞机制和后果。gydF4y2Ba
方法gydF4y2Ba血清来自7个健康志愿者暴露于14晚的间歇性缺氧,43阻塞性睡眠呼吸暂停综合症患者和健康对照组31日sVE内容进行分析。人类主动脉内皮细胞(HAECs)暴露在6 h间歇性缺氧gydF4y2Ba在体外gydF4y2Ba,有或没有抗氧化剂或抑制剂的低氧诱导因子(HIF) 1,酪氨酸激酶或血管内皮生长因子(VEGF)的途径。内皮渗透性VE-cadherin乳沟和磷酸化进行评估,评估通过测量transendothelial电阻(te)和异硫氰酸荧光素(FITC)右旋糖酐通量。gydF4y2Ba
结果gydF4y2BasVE显著升高血清中健康志愿者提交间歇性缺氧和阻塞性睡眠呼吸暂停综合症患者在治疗之前,但反过来降低阻塞性睡眠呼吸暂停患者的6个月后持续正压通气治疗。阻塞性睡眠呼吸暂停综合症的主要因素是占sVE多元分析的变化。在gydF4y2Ba在体外gydF4y2Ba实验,乳沟和表达VE-cadherin增加HAEC间歇性缺氧。三通和FITC-dextran通量的增加而减少。这些影响被所有的药理抑制剂逆转测试。gydF4y2Ba
结论gydF4y2Ba我们建议在阻塞性睡眠呼吸暂停综合症,间歇性缺氧阻塞性睡眠呼吸暂停通过诱导VE-cadherin乳沟的内皮通透性增加活性氧的生产,和激活HIF-1, VEGF和酪氨酸激酶通路。gydF4y2Ba
文摘gydF4y2Ba
这项研究首次表明,VE-cadherin裂解在睡眠呼吸暂停患者,在志愿者暴露于14晚上间歇性缺氧和内皮细胞接触gydF4y2Ba在体外gydF4y2Ba间歇性缺氧,导致内皮通透性增加gydF4y2Bahttps://bit.ly/3sAy5scgydF4y2Ba
介绍gydF4y2Ba
阻塞性睡眠呼吸暂停(OSA)是最常见的慢性疾病之一,影响了全球将近10亿人(gydF4y2Ba1gydF4y2Ba),特点是重复在睡眠中出现呼吸暂停和hypopnoeas [gydF4y2Ba2gydF4y2Ba]。反复发作的关键特性hypoxia-reoxygenation,阻塞性睡眠呼吸暂停综合症的有害后果的主要中介机制是间歇性缺氧。gydF4y2Ba
阻塞性睡眠呼吸暂停与心血管发病率和死亡率升高由于风险增加高血压、动脉粥样硬化、冠心病和脑血管疾病(gydF4y2Ba2gydF4y2Ba]。间歇性缺氧是公认的关键中介机制导致心血管发病率和死亡率。的确,间歇性缺氧诱发各种过程,如氧化应激、激活低氧诱导因子(HIF) 1,轻度慢性炎症和交感神经激活,进而负责内皮功能障碍、高血压和动脉粥样硬化gydF4y2Ba2gydF4y2Ba- - - - - -gydF4y2Ba4gydF4y2Ba]。重要的是,阻塞性睡眠呼吸暂停综合症之间的联系/间歇性缺氧和血管内皮细胞的结构和功能障碍已被解决在患者和动物模型,提高氧化应激,更高的炎性细胞因子的生产和pro-atherogenic粘附分子(已报告gydF4y2Ba3gydF4y2Ba- - - - - -gydF4y2Ba7gydF4y2Ba]。然而,尽管内皮通透性的调节主要在早期动脉粥样化形成的起始和发展重要性,这方面到目前为止一直在稀疏研究阻塞性睡眠呼吸暂停综合症。细胞和分子机制的调节内皮通透性阻塞性睡眠呼吸暂停综合症的研究在有限数量的研究无论是在阻塞性睡眠呼吸暂停综合症患者和啮齿动物等实验模型(gydF4y2Ba8gydF4y2Ba)或细胞培养(gydF4y2Ba9gydF4y2Ba,gydF4y2Ba10gydF4y2Ba暴露于间歇性缺氧。gydF4y2Ba
内皮屏障通透性是由VE-cadherin,粘合连接处并且通过其亲同种抗原的粘附能力,发挥了关键作用[gydF4y2Ba11gydF4y2Ba]。针对VE-cadheringydF4y2Ba在体外gydF4y2Ba使用特定针对抗原表位的抗体在细胞外领域阻碍的形成和内皮通透性增加(粘合连接处并且gydF4y2Ba12gydF4y2Ba]。两种机制解释了减少VE-cadherin描述了亲同种抗原的附着力和已知增加内皮通透性:第一个是掩饰的整个分子(gydF4y2Ba13gydF4y2Ba),第二,最近描述,乳沟的胞外蛋白酶如基质金属蛋白酶域。这乳沟释放90 kda可溶性片段(sVE)检测在血液gydF4y2Ba14gydF4y2Ba,gydF4y2Ba15gydF4y2Ba)和血清水平升高sVE已经记录在几个人类疾病的特点是内皮功能障碍,如动脉粥样硬化、系统性血管炎和癌症转移(gydF4y2Ba16gydF4y2Ba,gydF4y2Ba17gydF4y2Ba]。gydF4y2Ba
VE-cadherin的胞质域的磷酸化酪氨酸站点(Y685)酪氨酸激酶的Src的早期事件的重大机制VE-cadherin乳沟和调节内皮渗透性gydF4y2Ba15gydF4y2Ba,gydF4y2Ba18gydF4y2Ba- - - - - -gydF4y2Ba20.gydF4y2Ba]。这些磷酸化过程等细胞因子诱导的血管内皮生长因子(VEGF) [gydF4y2Ba18gydF4y2Ba)和肿瘤坏死因子(TNF) -α[gydF4y2Ba15gydF4y2Ba]。gydF4y2Ba
我们所知,在阻塞性睡眠呼吸暂停综合症VE-cadherin监管从未被研究过。几个信号通路参与VE-cadherin乳沟被激活在患有阻塞性睡眠呼吸暂停综合症或间歇性缺氧模型,在升高VEGF (gydF4y2Ba6gydF4y2Ba,gydF4y2Ba21gydF4y2Ba,gydF4y2Ba22gydF4y2Ba]和TNF-α[gydF4y2Ba23gydF4y2Ba)水平。因此,我们假设,间歇性缺氧可以激活信号通路导致VE-cadherin解理,从而增加内皮渗透性。这些间歇性低氧诱导通路可能包括代氧化应激激活的HIF-1活性氧(ROS) (gydF4y2Ba2gydF4y2Ba,gydF4y2Ba24gydF4y2Ba),VEGF的表达作为目标基因HIF-1 [gydF4y2Ba25gydF4y2Ba,gydF4y2Ba26gydF4y2Ba),和激活的Src激酶VEGF信号(gydF4y2Ba18gydF4y2BaROS(直接)和/或gydF4y2Ba27gydF4y2Ba]。gydF4y2Ba
我们的目标是1)阻塞性睡眠呼吸暂停综合症患者的血清研究sVE水平治疗前后与持续气道正压(CPAP)以及健康志愿者暴露于14晚的间歇性缺氧,和2)解剖gydF4y2Ba在体外gydF4y2Ba的分子机制参与VE-cadherin乳沟和渗透率增加内皮细胞模型间歇性缺氧。gydF4y2Ba
一个视觉的研究可以发现的概述gydF4y2Ba补充材料gydF4y2Ba。gydF4y2Ba
方法gydF4y2Ba
研究人群gydF4y2Ba
健康志愿者暴露于14日晚上间歇性缺氧gydF4y2Ba
我们使用血清从双盲随机交叉研究(gydF4y2BaClinicalTrials.govgydF4y2Ba:gydF4y2BaNCT02058823gydF4y2Ba),12名身体健康的志愿者接触间歇性缺氧(吸入氧气分数(gydF4y2BaFgydF4y2BaIOgydF4y2Ba2gydF4y2Ba骑自行车在0.21和0.13之间;120年代hypoxia-reoxygenation周期),每晚8 - 9小时,14日晚上或间歇空气(gydF4y2BaFgydF4y2BaIOgydF4y2Ba2gydF4y2Ba0.21)与类似的气流控制(gydF4y2Ba28gydF4y2Ba]。其中,7个志愿者完成的两臂交叉试验。收集血液样本从这七个早晨禁食志愿者,之前和之后的2周的隔夜敞口间歇空气或间歇性缺氧,并存储在−80°C。可以发现更多细节gydF4y2Ba补充材料gydF4y2Ba。gydF4y2Ba
阻塞性睡眠呼吸暂停综合症的病人之前和之后CPAP治疗gydF4y2Ba
生物样本病例对照研究,包括43阻塞性睡眠呼吸暂停综合症的病人没有任何已知的心血管疾病和31个对照组没有阻塞性睡眠呼吸暂停综合症或心血管疾病(gydF4y2BaClinicalTrials.govgydF4y2Ba:gydF4y2BaNCT00764218gydF4y2Ba)进行分析gydF4y2Ba29日gydF4y2Ba]。血清样本被早晨禁食后一夜之间多导睡眠图6个月之前和之后执行CPAP治疗,或在6个月随访干预对照组。血清是储存在−80°C到使用。VEGF在血清ELISA测定(Quantikine;研发系统,明尼阿波利斯,美国)(gydF4y2Ba6gydF4y2Ba]。gydF4y2Ba
道德gydF4y2Ba
两个研究伦理从我们的机构审查委员会批准了(拉西德保护des人Sud-Est V,格勒诺布尔,法国),所有科目或病人给书面知情同意。协议符合赫尔辛基宣言的原则。gydF4y2Ba
细胞培养gydF4y2Ba
细胞gydF4y2Ba
低温贮藏的主要人类主动脉内皮细胞(HAECs)(美国马热费希尔科学、沃尔瑟姆)被使用之前通过8。细胞被播种密度为1.6×10gydF4y2Ba4gydF4y2Ba细胞·厘米gydF4y2Ba−2gydF4y2Ba与大血管内皮补充(M200中补充母体;热费希尔科学)和青霉素和链霉素根据供应商的建议,对细胞培养使用传统孵化器有限公司(37°C / 5%gydF4y2Ba2gydF4y2Ba)。细胞培养试剂购买从Gibco热费希尔科学(Illkirch、法国)。gydF4y2Ba
间歇性低氧暴露gydF4y2Ba
细胞暴露于快速间歇性缺氧周期使用最近开发的设备,模仿的hypoxia-reoxygenation周期阻塞性睡眠呼吸暂停综合症的病人,如前所述[gydF4y2Ba24gydF4y2Ba]。一个间歇性缺氧周期包括一个5分钟normoxia阶段(16%啊gydF4y2Ba2gydF4y2Ba),后跟一个5分钟缺氧阶段(2%啊gydF4y2Ba2gydF4y2Ba)。可以发现更多细节gydF4y2Ba补充材料gydF4y2Ba。gydF4y2Ba
化学品和抑制剂gydF4y2Ba
可以找到的细节gydF4y2Ba补充材料gydF4y2Ba。gydF4y2Ba
transendothelial电阻的测量gydF4y2Ba
支流HAECs播种transwell插入(gydF4y2Ba补充材料gydF4y2Ba)被放置在M200没有母体,用二甲基亚砜(DMSO)或抑制剂治疗和暴露于间歇性缺氧或normoxia 6 h。内皮通透性是评估通过测量transendothelial电阻(te)在0和6 h的接触使用EVOM电阻计(美国佛罗里达州萨拉索塔世界精密仪器)。三个值的te记录每插入一个空白的插入(缺乏细胞)被用来减去空白t值。数据表示为te的百分比值的插入包含支流常氧细胞。gydF4y2Ba
测量荧光素isothiocyanate-dextran通量gydF4y2Ba
支流HAECs播种transwell插入(gydF4y2Ba补充材料gydF4y2Ba)被放置在M200没有母体和间歇性缺氧或normoxia公开。4 h后曝光,40-kDa异硫氰酸荧光素(FITC)右旋糖酐(Sigma-Aldrich,达姆施塔特,德国)添加µg·200毫升gydF4y2Ba−1gydF4y2Batranswell的上院。细胞被放置在间歇低氧设备2 h。FITC-dextran通量穿过内皮单层的量化监控通过测量荧光在众议院在文化。数据表示为百分比的荧光常氧插入。gydF4y2Ba
免疫荧光gydF4y2Ba
细胞(先前与4%多聚甲醛固定,PBS 10 min) permeabilised和沾鼠单克隆anti-VE-cadherin主要抗体针对细胞外域(BV9克隆;默克密理博,达姆施塔特,德国)其次是Alexa萤石488 anti-mouse二级抗体(热费希尔科学)。肌动蛋白是沾phalloidin和细胞核染色和4′,6-diamidino-2-phenylindole。细胞是由荧光显微镜检查(Axio成像仪;蔡司、Oberkocheņ德国)和荧光膜面积使用ImageJ量化软件(斐济;gydF4y2Baimagej.net/FijigydF4y2Ba)减去从全细胞染色细胞质和核染色。gydF4y2Ba
VE-cadherin磷酸化检测gydF4y2Ba
细胞被置于M200没有母体1 h,然后处理抑制剂和暴露于间歇性缺氧或normoxia 6 h。曝光后,细胞被取消在冰上用PBS和离心机250×gydF4y2BaggydF4y2Ba在4°C, 5分钟。的存在磷酸化VE-cadherin在细胞提取物被西方墨点法评估与一只兔子anti-phospho-Y685-VE-cadherin抗体(1µg·毫升gydF4y2Ba−1gydF4y2Ba)。可用的更多细节gydF4y2Ba补充材料gydF4y2Ba。gydF4y2Ba
sVE检测gydF4y2Ba
参与者的血清和集中HAEC上层清液为sVE内容进行分析。血清在PBS稀释1:50补充特里同为0.5%。HAEC上层清液收集后间歇性低氧暴露和集中使用Amicon Ultra-4离心过滤列与30-kDa分子量截止(Sigma-Aldrich,达姆施塔特,德国)。sVE的存在是由西方墨点法评估用鼠标单克隆anti-VE-cadherin主要抗体(BV9克隆;1µg·毫升gydF4y2Ba−1gydF4y2Ba)。调查人员对血清的来源也不清楚。更详细的免疫印迹协议中是可用的gydF4y2Ba补充材料gydF4y2Ba。gydF4y2Ba
统计分析gydF4y2Ba
未配对比较两组都用t检验或Mann-Whitney测试和成对比较进行使用Wilcoxon符号秩检验。对比几组是由双向方差分析的紧随其后gydF4y2Ba事后gydF4y2Ba图基测试。参数或非参数测试的选择取决于值的正态分布和方差的平等。当适用(执行重复测量方差分析gydF4y2Ba即。gydF4y2Ba受试者在基线和月6)。一个单变量线性模型被用来确定sVE的相关因素。自从sVE不是正态分布,进行对数变换。变量与p < 0.2的单变量被认为是在多变量线性回归分析,我们调整了混杂参数(年龄、性别、心率、动脉压,腰围/臀围,胆固醇,glycaemia和肌酐)。最后,评估下血清sVE CPAP的进化,一个多元混合线性回归对病人进行随机影响,调整年龄、性别、CPAP sVE之间的坚持和持续时间的测量。gydF4y2Ba
在细胞培养实验中,考虑到这样的事实:一些井通常被使用在一个独立的实验中,实验包括线性混合模型中的随机因素,在固定效应是间歇性缺氧和药理治疗风险。我们确认,没有显著的影响实验效果的间歇性缺氧或药物治疗。gydF4y2Ba
结果被认为是重要的,当p < 0.05。值报告为±gydF4y2Ba扫描电镜gydF4y2Ba或中位数(四分位范围(差),根据正态分布。gydF4y2Ba
结果gydF4y2Ba
间歇性低氧刺激VE-cadherin健康个体的乳沟gydF4y2Ba
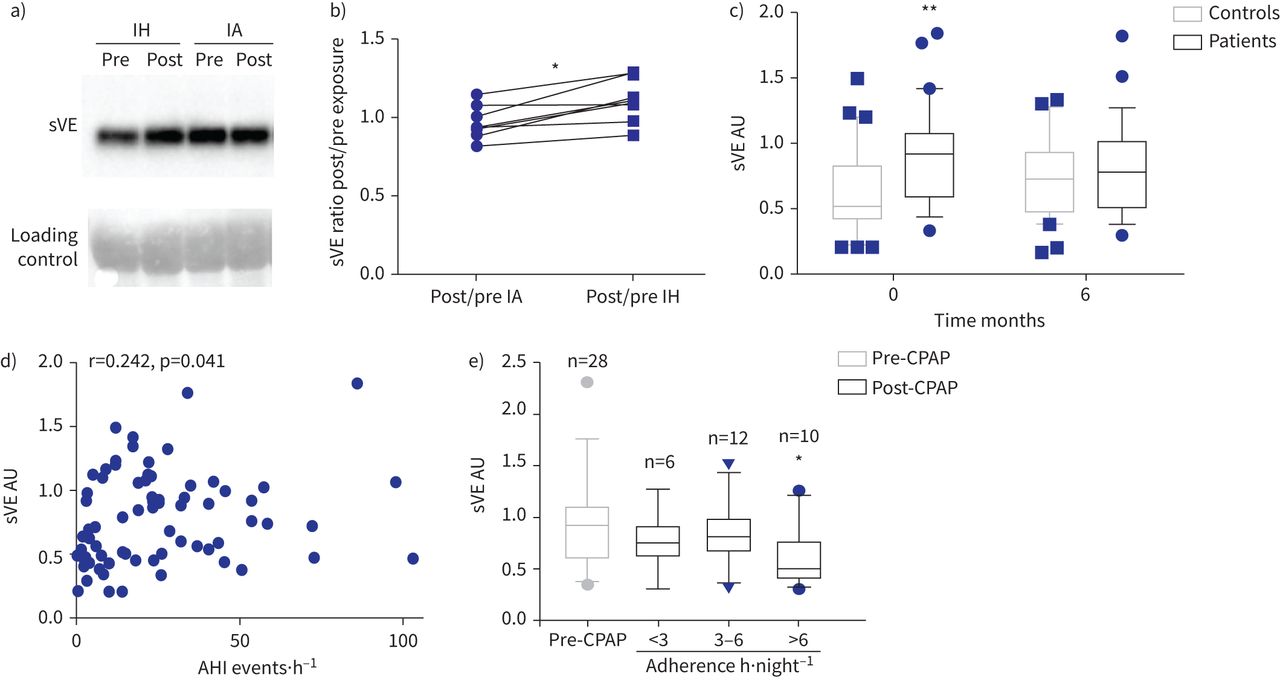
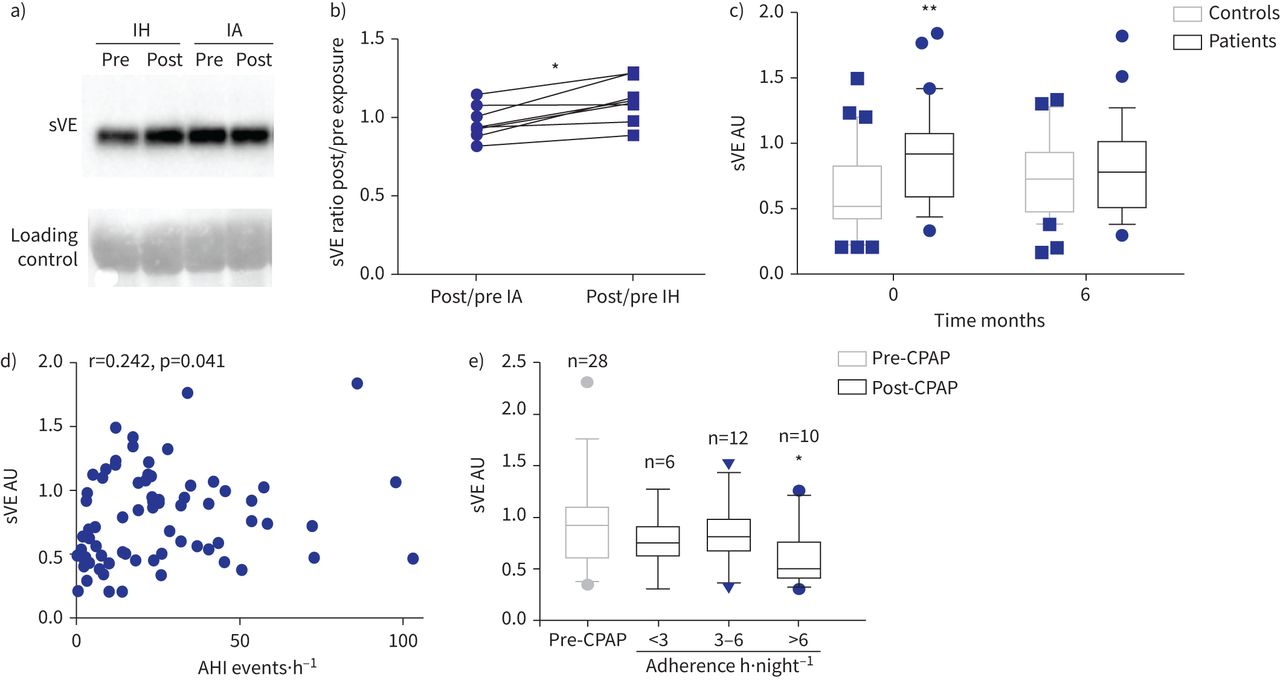
我们评估血清sVE水平的七个健康志愿者随机分配到14晚上间歇性缺氧或间歇空气。受试者的特征是:平均年龄为23.8±3.5年,身体质量指数(BMI) 21.3±2.4公斤·mgydF4y2Ba−2gydF4y2Ba,和五个7∼70%是男性。sVE水平保持稳定接触后14天的间歇空气(中位数post / pre间歇空气比0.94)时增加了19%后14天的间歇性低氧暴露(post / pre间歇性缺氧比中值1.12;p = 0.015gydF4y2Ba与gydF4y2Ba间歇空气)(gydF4y2Ba图1一个gydF4y2Ba和b)。gydF4y2Ba
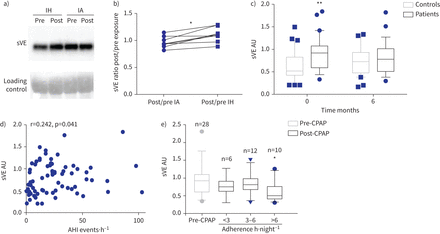
循环溶在血清VE (sVE)增加健康志愿者(控制)受到间歇性缺氧(IH)和阻塞性睡眠呼吸暂停(OSA)患者。IA:间歇空气;你好:apnoea-hypopnoea指数;CPAP:持续气道正压。)免疫印迹显示代表的例子sVE (90gydF4y2BakDa)血清健康志愿者之前和之后的14天的接触或IA。b)比sVE post / IH之前或IA接触计算量化后的西方墨迹了七个人。*:p = 0.015gydF4y2Ba与gydF4y2BaIA;Wilcoxon测试。c) sVE在对照组的病人明显高于基线(月0)(* *:p = 0.0058;重复测量双向方差分析)但不是6gydF4y2Ba个月的CPAP治疗(p = 0.78)。30名学生对于每一列,因为只有科目月0和月6样本可以被包含在这个配对分析。d)枪兵AHI之间的相关性和sVE基线。72年的主题。e)患者高坚持CPAP疗法(> 6gydF4y2Bah·夜gydF4y2Ba−1gydF4y2Ba)后显著降低血清水平sVE的6gydF4y2Ba个月(*:p = 0.034gydF4y2Ba与gydF4y2BasVE级别CPAP治疗前)与附着少患者(< 6gydF4y2Bah·夜gydF4y2Ba−1gydF4y2Ba;p > 0.99gydF4y2Ba与gydF4y2BaCPAP治疗前sVE水平);克鲁斯卡尔-沃利斯测试。图基块表示中位数和四分位范围。gydF4y2Ba
阻塞性睡眠呼吸暂停患者的sVE水平高于健康对照组gydF4y2Ba
研究人口的基线特征所示gydF4y2Ba补充表S1gydF4y2Ba。体重指数、甘油三酯和舒张压在阻塞性睡眠呼吸暂停综合症的人略高,无临床意义。在基线,sVE中值(+ 70%)的患者明显高于对照组(0.90 (0.57 - -1.06)gydF4y2Ba与gydF4y2Ba0.52(0.43 - -0.92)盟;p = 0.0058) (gydF4y2Ba图1 cgydF4y2Ba和gydF4y2Ba补充表S1gydF4y2Ba)。单变量分析表明,在参数与sVE显著相关值是阻塞性睡眠呼吸暂停综合症的严重程度指数apnoea-hypopnoea指数(AHI)和最低血氧饱和度、性别、收缩压和舒张压,血清VEGF浓度(gydF4y2Ba表1gydF4y2Ba)。AHI与sVE呈正相关(r = 0.242, p = 0.041) (gydF4y2Ba图1 dgydF4y2Ba)。在多变量分析中,阻塞性睡眠呼吸暂停综合症相关的独立地位是sVE增加34.6% (p = 0.005)。雄性与sVE增加38.7% (p = 0.01) (gydF4y2Ba表2gydF4y2Ba)。gydF4y2Ba
单变量分析的因素在基线可溶性已经变化gydF4y2Ba
独立基线参数与可溶的多变量分析gydF4y2Ba
sVE CPAP治疗后下降gydF4y2Ba
CPAP治疗6个月后,没有差别sVE水平对照组和患者之间(0.77(0.51 - -0.92)非盟对病人gydF4y2Ba与gydF4y2Ba0.72(0.47 - -0.93)盟控制;p = 0.78) (gydF4y2Ba图1 cgydF4y2Ba)。sVE水平对照组之间倾向于增加月0和6月,但这同样没有显著增加(p = 0.59)。在一个gydF4y2Ba事后gydF4y2Ba分析,减少sVE水平最高的只有发生在病人夜间CPAP用途(p = 0.034 CPAP时用于> 6 h·夜gydF4y2Ba−1gydF4y2Ba与gydF4y2Ba无意义的CPAP使用< 3或3 - 6 h·夜gydF4y2Ba−1gydF4y2Ba)(gydF4y2Ba图1 egydF4y2Ba)。gydF4y2Ba
间歇性低氧增加VE-cadherin表达式和乳沟gydF4y2Ba在体外gydF4y2Ba导致内皮通透性gydF4y2Ba
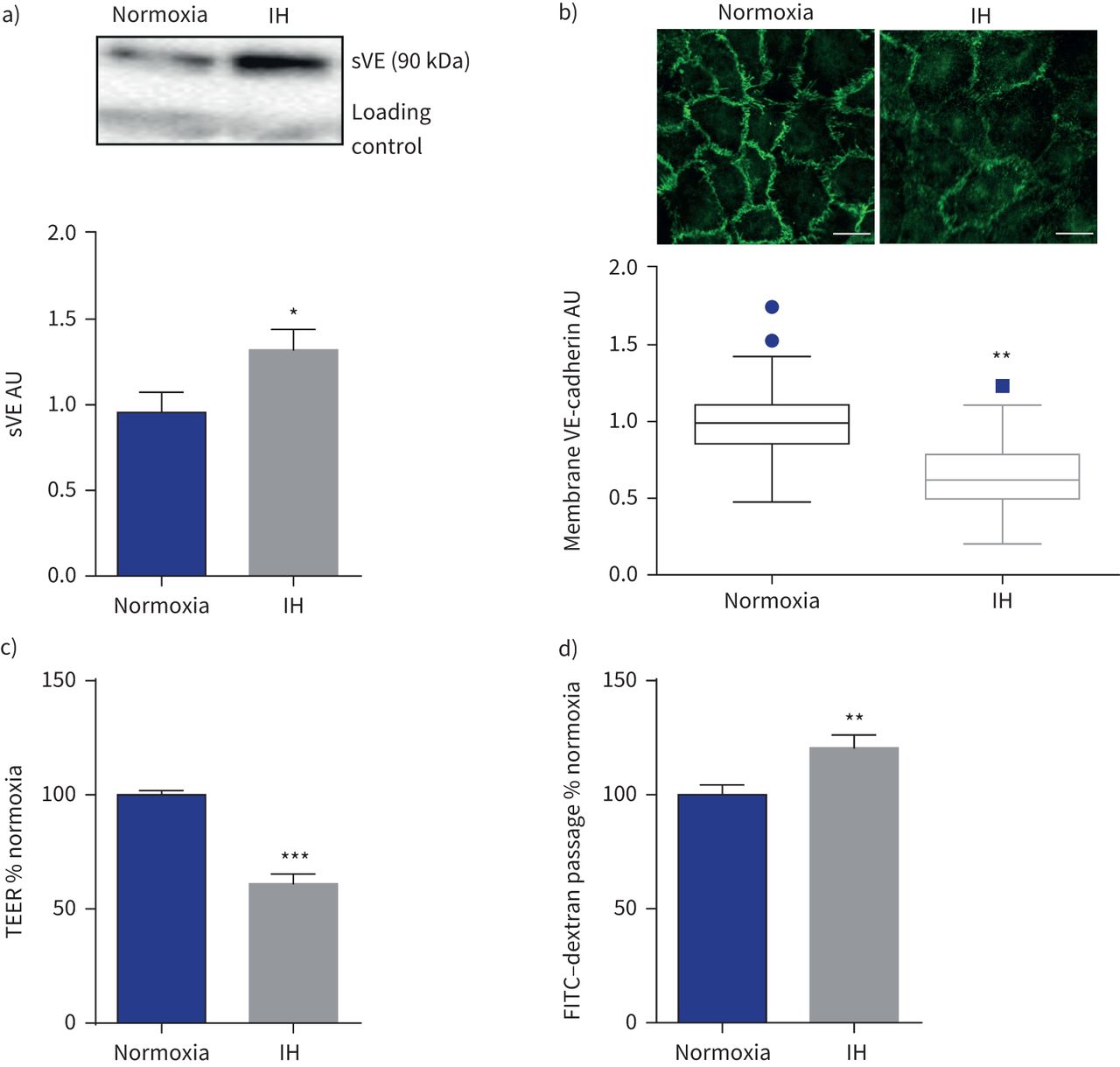
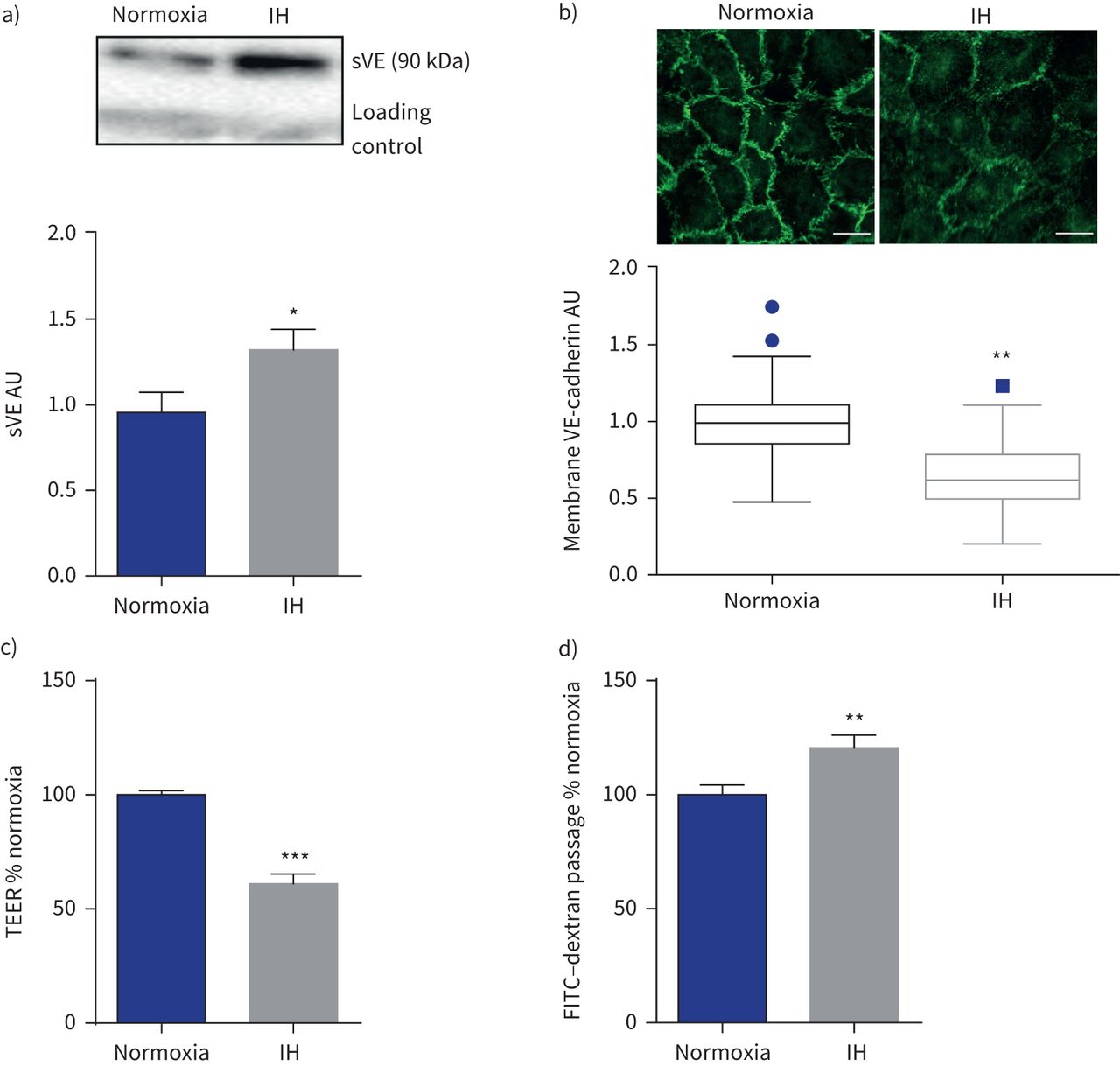
免疫印迹分析显示显著增加sVE HAECs 6 h后上层清液的接触间断缺氧而normoxia (+ 39%;p < 0.05) (gydF4y2Ba图2一个gydF4y2Ba)。这结果是伴随着一个明显减少细胞外的存在域的VE-cadherin细胞膜免疫荧光(如图所示gydF4y2Ba图2 bgydF4y2Ba)。有显著减少37.5%的te HAEC层暴露6 h(间歇性缺氧而normoxia (gydF4y2Ba图2 cgydF4y2Ba),反映内皮通透性的增加。这证实了增加transendothelial FITC-dextran间歇性缺氧后通量(+ 19.5%)(gydF4y2Ba图2 dgydF4y2Ba)。gydF4y2Ba
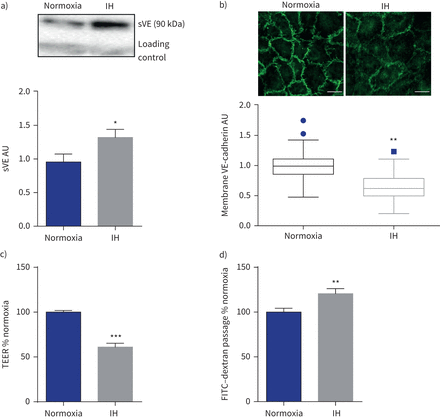
间歇性缺氧(IH)改变了内皮屏障。sVE:可溶;te: transendothelial电阻;FITC:异硫氰酸荧光素。)代表的例子免疫印迹显示更强烈sVE乐队在人类主动脉内皮细胞暴露于IH的上层清液比normoxia(一种免疫球蛋白乐队是用作正常化加载控制)和量化sVE测量细胞上清液后6 h (IH)。*:p < 0.05gydF4y2Ba与gydF4y2Banormoxia;界,n = 15。b)免疫荧光和量化anti-VE-cadherin染色与抗体针对VE-cadherin域显示细胞外膜IH后表达降低。* *:p = 0.01gydF4y2Ba与gydF4y2Banormoxia;Mann-Whitney等级和测试,四个独立的实验,每10 - 14细胞实验。酒吧规模:20µm。c)增加内皮渗透性衡量三通后6 h (IH)。* * *:p < 0.001gydF4y2Ba与gydF4y2Banormoxia;韦尔奇测试,16 transwells从四个独立的实验。d)增加内皮渗透性衡量FITC-dextran transendothelial通量(通道)。* *:p < 0.01gydF4y2Ba与gydF4y2Banormoxia;韦尔奇测试,33 transwells来自12个独立的实验。数据意味着±gydF4y2Ba扫描电镜gydF4y2Ba除了图基图b)中提出了数据的中位数和四分位范围由于nonnormality数据分布。gydF4y2Ba
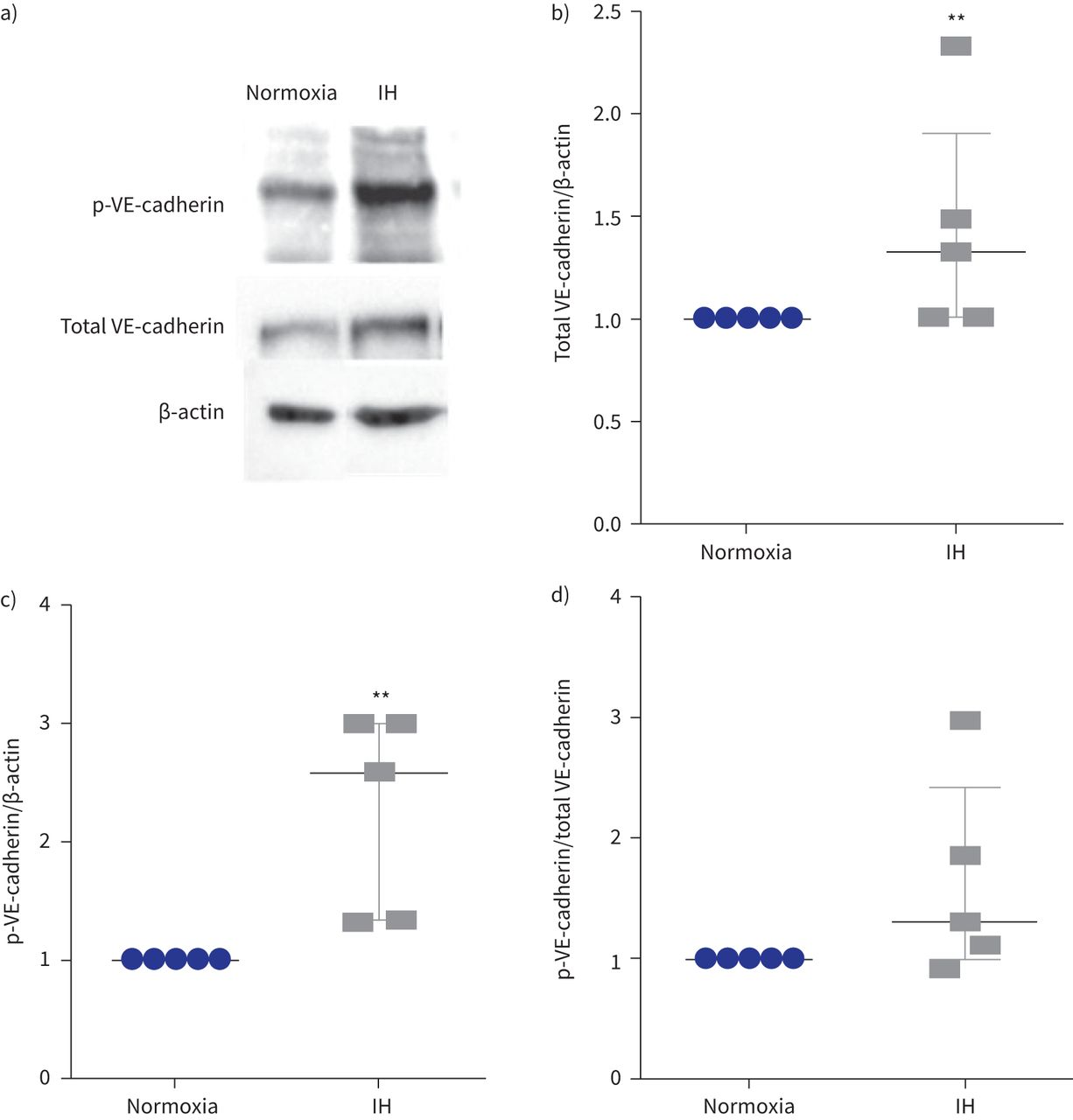
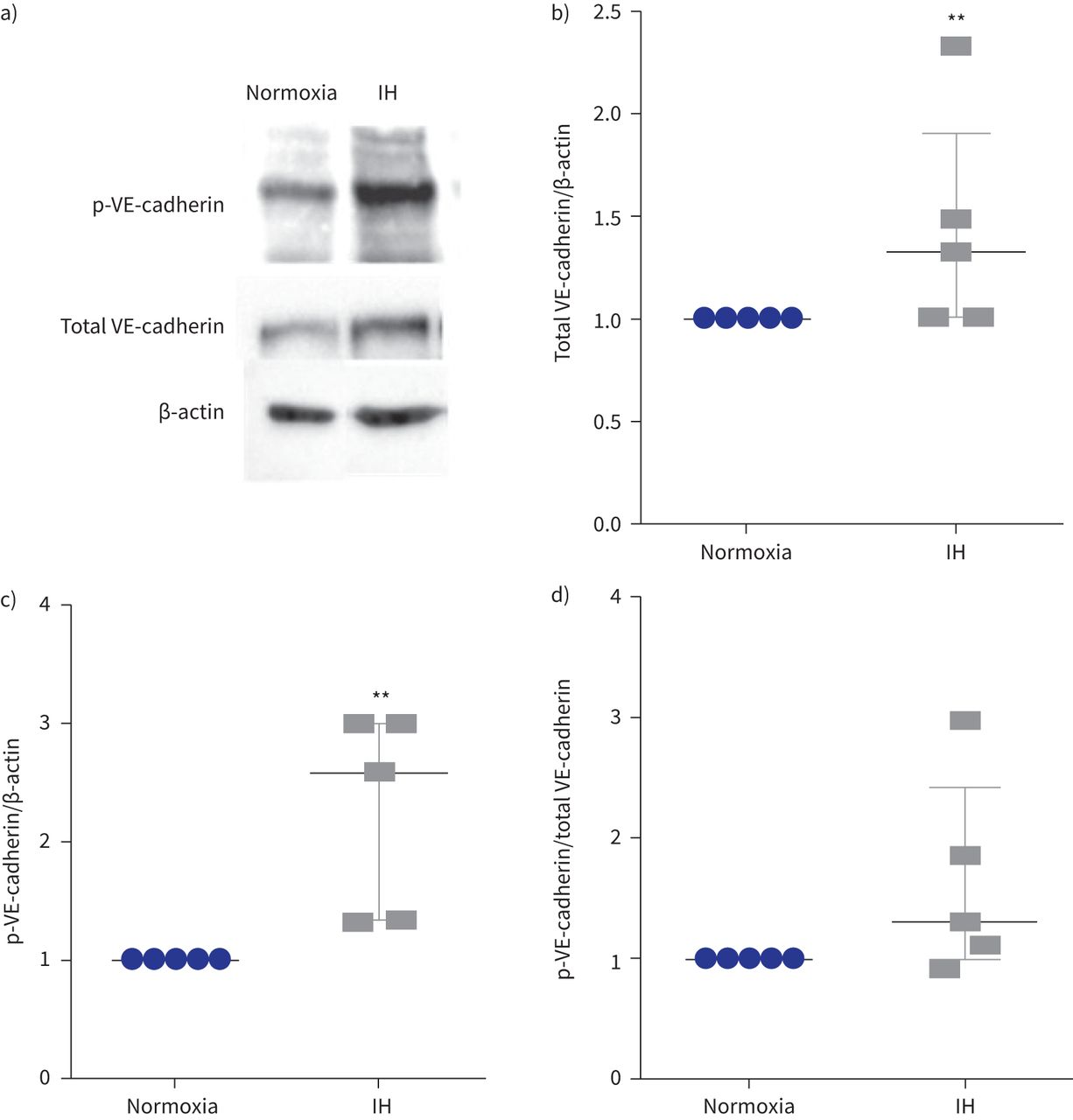
我们还表明,间歇性缺氧诱导的总量增加1.3倍VE-cadherin (gydF4y2Ba图3一gydF4y2Ba和b)伴随着增加2.5倍的磷酸化VE-cadherin酪氨酸(Y685)与β-actin (gydF4y2Ba图3 cgydF4y2Ba)。磷酸化VE-cadherin /总VE-cadherin比率往往升高(中位数(差)比1.3(1.0 - -2.4))虽然没有达到显著性(p = 0.12) (gydF4y2Ba图3 dgydF4y2Ba)。gydF4y2Ba
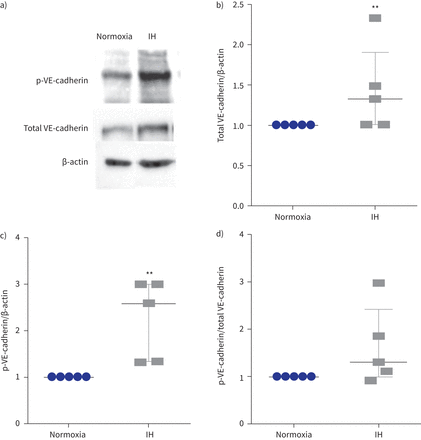
表达和磷酸化的酪氨酸Y685 VE-cadherin间歇性缺氧后(IH)。p:磷酸化。)代表的例子免疫印迹显示更强烈p-VE-cadherin乐队在细胞提取物的人体主动脉内皮细胞暴露于IH normoxia相比。β-actin作为正常化的加载控制。b)量化级别的总VE-cadherin正常化β-actin测量细胞溶解产物。* *:p < 0.01gydF4y2Ba与gydF4y2Banormoxia;Mann-Whitney测试,n = 5。c)水平的量化p-VE-cadherin正常化β-actin以细胞溶解产物。* *:p < 0.01gydF4y2Ba与gydF4y2Banormoxia;Mann-Whitney测试,n = 5。d) p-VE-cadherin /总VE-cadherin比率。p = 0.12;Mann-Whitney测试,n = 5。数据中位数和四分位范围。gydF4y2Ba
信号通路参与间歇性低氧诱导VE-cadherin乳沟gydF4y2Ba
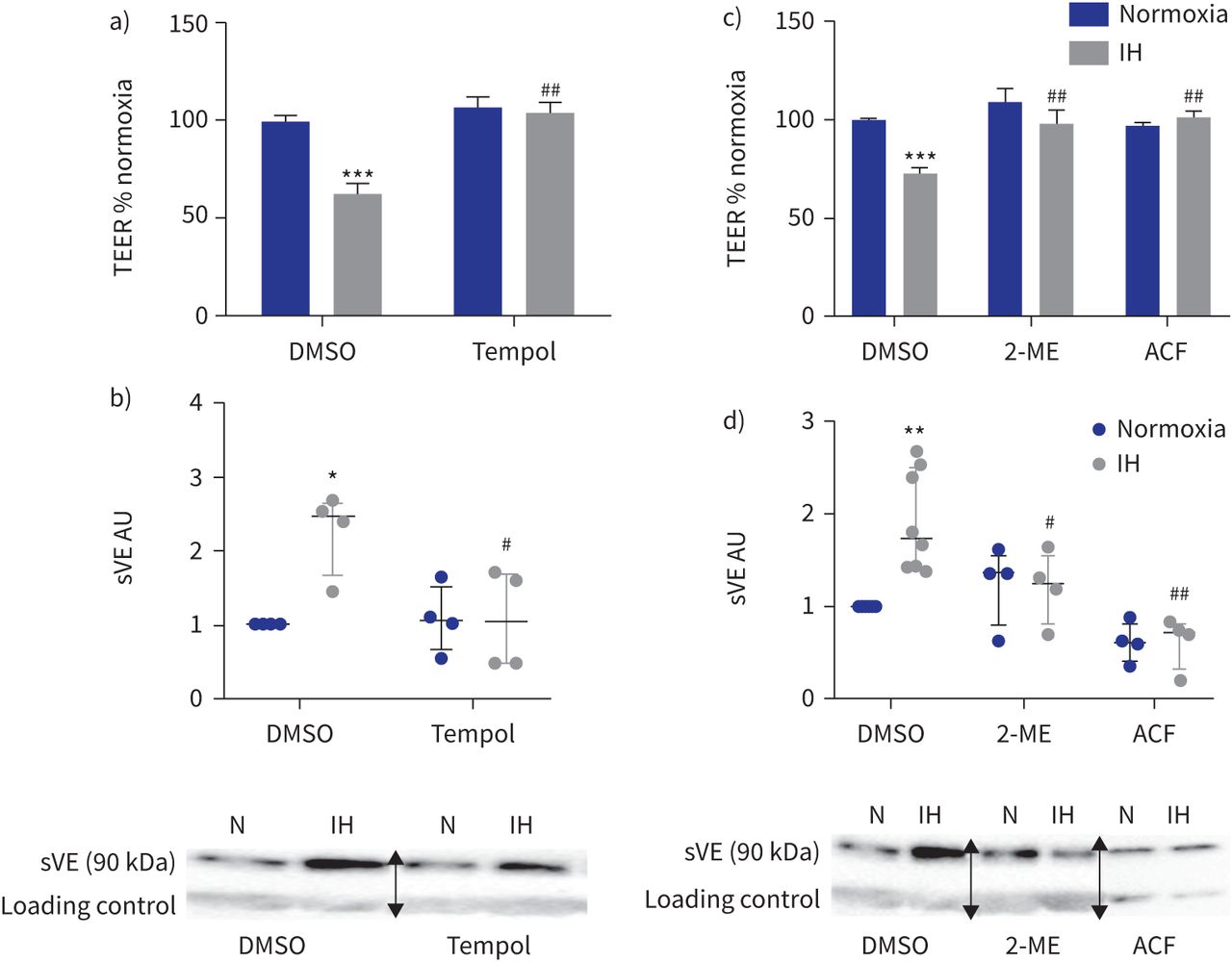
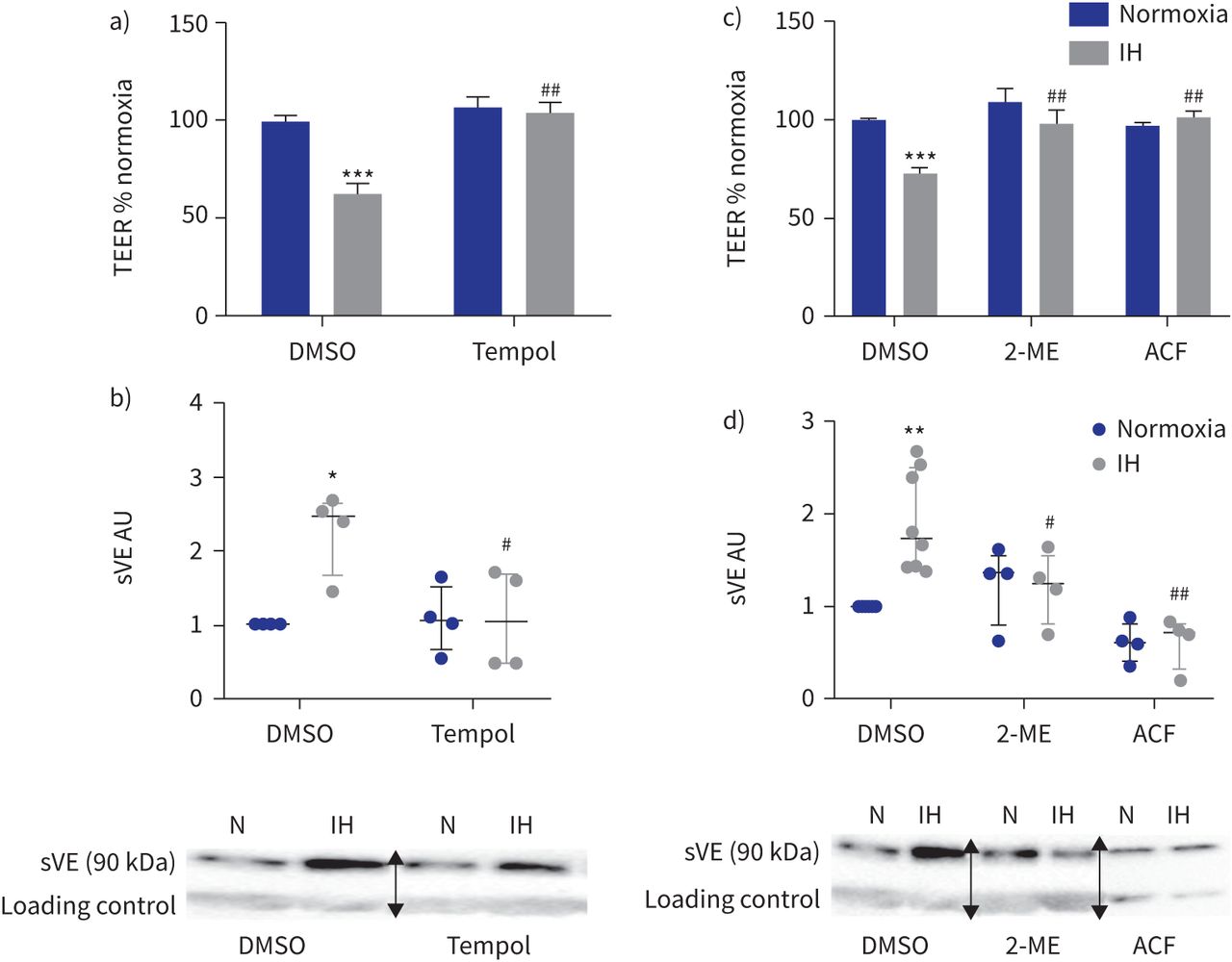
我们研究了氧化应激的作用,HIF-1激活,VEGF信号和酪氨酸激酶活动。首先,氧化应激的作用是通过使用Tempol加以解决,一个强有力的活性氧清除剂作为一种超氧化物歧化酶模拟物。Tempol显著增加t值和减少sVE水平HAEC上层清液下间歇缺氧和DMSO溶液相比,抑制间歇性缺氧影响并返回三通和sVE水平基线(gydF4y2Ba图4一gydF4y2Ba和b)。对te Tempol没有影响,normoxia-exposed sVE水平细胞(gydF4y2Ba图4一gydF4y2Ba和b)。gydF4y2Ba
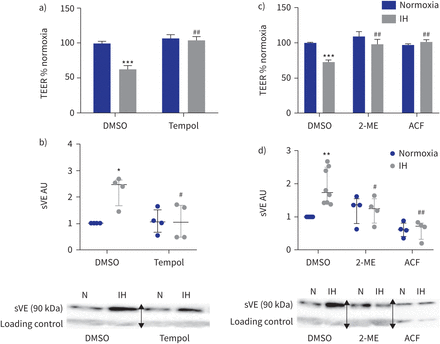
活性氧清除剂的影响(Tempol)和低氧诱导因子(HIF) 1抑制内皮屏障间歇性缺氧(IH)条件下。te: transendothelial电阻;DMSO:二甲亚砜;sVE:可溶;我:2-methoxyestradiol;ACF:吖啶黄;HAEC:人类主动脉内皮细胞。一)Tempol (100gydF4y2BaµM)废除了te IH对内皮渗透性的影响评估。* * *:p < 0.001gydF4y2Ba与gydF4y2Banormoxia + DMSO溶液;gydF4y2Ba# #gydF4y2Ba:p < 0.01gydF4y2Ba与gydF4y2BaIH + DMSO溶液;16 transwells双向方差分析,从四个独立的实验。数据意味着±gydF4y2Ba扫描电镜gydF4y2Ba。b) sVE测量水平的上层清液HAECs Tempol对待。*:p < 0.05gydF4y2Ba与gydF4y2Banormoxia + DMSO溶液;gydF4y2Ba#gydF4y2Ba:p < 0.05gydF4y2Ba与gydF4y2BaIH + DMSO溶液;双向方差分析,n = 4。数据中位数和四分位范围。下面是代表西方墨点法的例子显示那么激烈sVE乐队HAECs的上层清液治疗婴幼儿血管瘤+ Tempol相比之下,IH + DMSO(一种免疫球蛋白乐队作为加载控制正常化)。N: normoxia。c)抑制HIF-1我(1gydF4y2BaµM)或ACF (1gydF4y2BaµM)防止由三通IH对内皮渗透性的影响评估。* * *:p < 0.001gydF4y2Ba与gydF4y2Banormoxia + DMSO溶液;gydF4y2Ba# #gydF4y2Ba:p < 0.01gydF4y2Ba与gydF4y2BaIH + DMSO溶液;双向方差分析,16个独立transwells从四个独立的实验。数据意味着±gydF4y2Ba扫描电镜gydF4y2Ba。d) sVE水平的量化测量的上层清液HAECs对待HIF-1抑制剂。* *:p < 0.01gydF4y2Ba与gydF4y2Banormoxia + DMSO溶液;gydF4y2Ba# #gydF4y2Ba:p < 0.01gydF4y2Ba#gydF4y2Ba:p < 0.05gydF4y2Ba与gydF4y2BaIH + DMSO溶液;双向方差分析,n = 4。数据中位数和四分位范围。下面是代表西方墨点法的例子显示那么激烈sVE乐队HAECs的上层清液治疗婴幼儿血管瘤+ HIF-1抑制剂相比,IH + DMSO(一种免疫球蛋白乐队作为加载控制正常化)。N: normoxia。在西方的屁股,箭头表示图像不同部分的分组相同的凝胶或从不同的凝胶。gydF4y2Ba
同样,抑制HIF-1活动2-methoxyestradiol或吖啶黄完全阻止任何间歇低氧对内皮渗透性的影响和VE-cadherin劈理(gydF4y2Ba图4 cgydF4y2Ba和d)。gydF4y2Ba
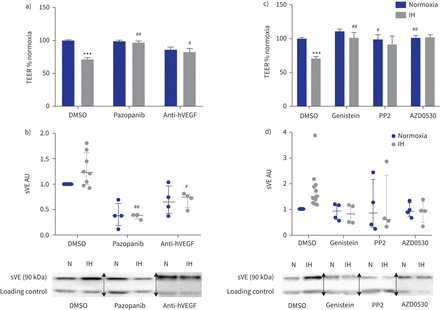
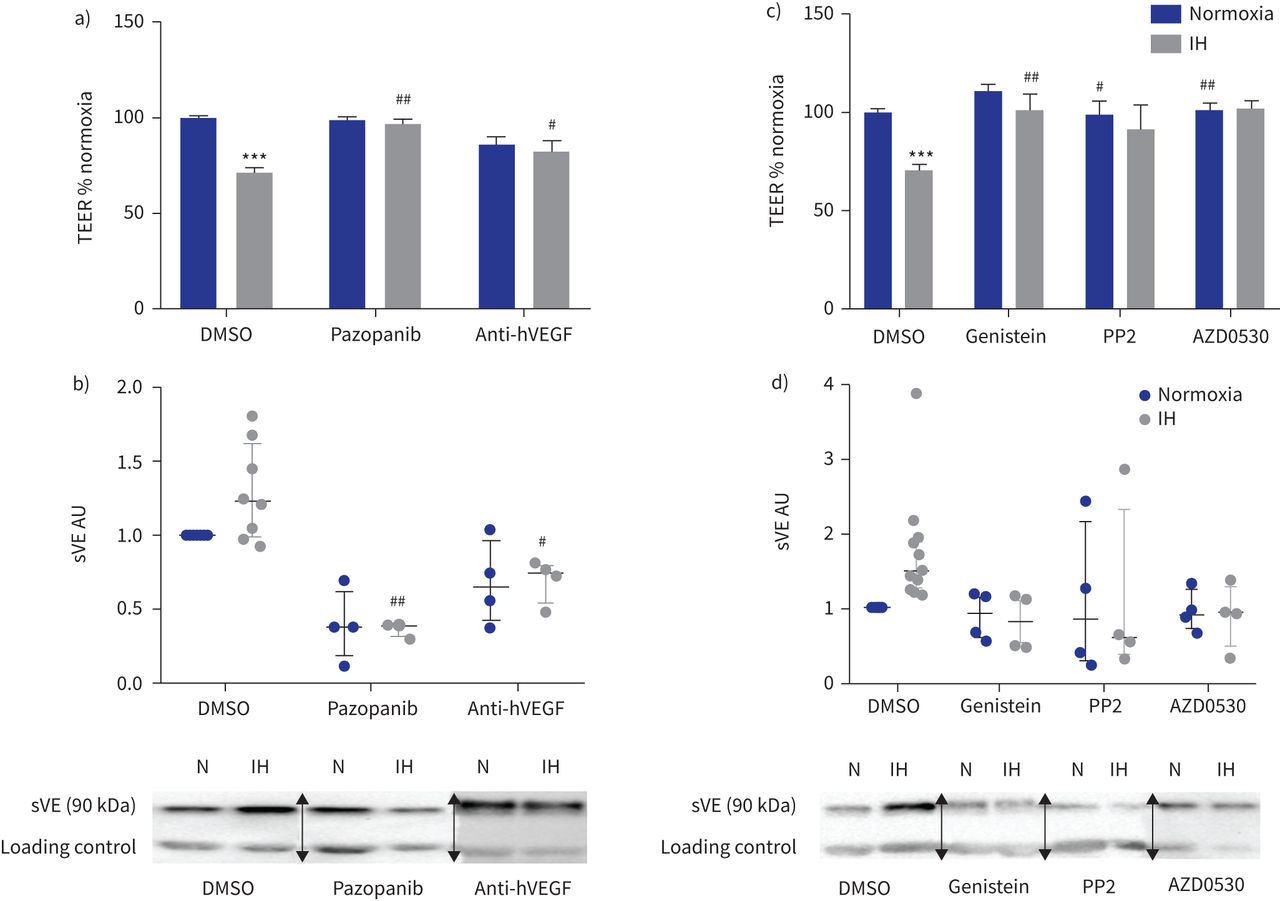
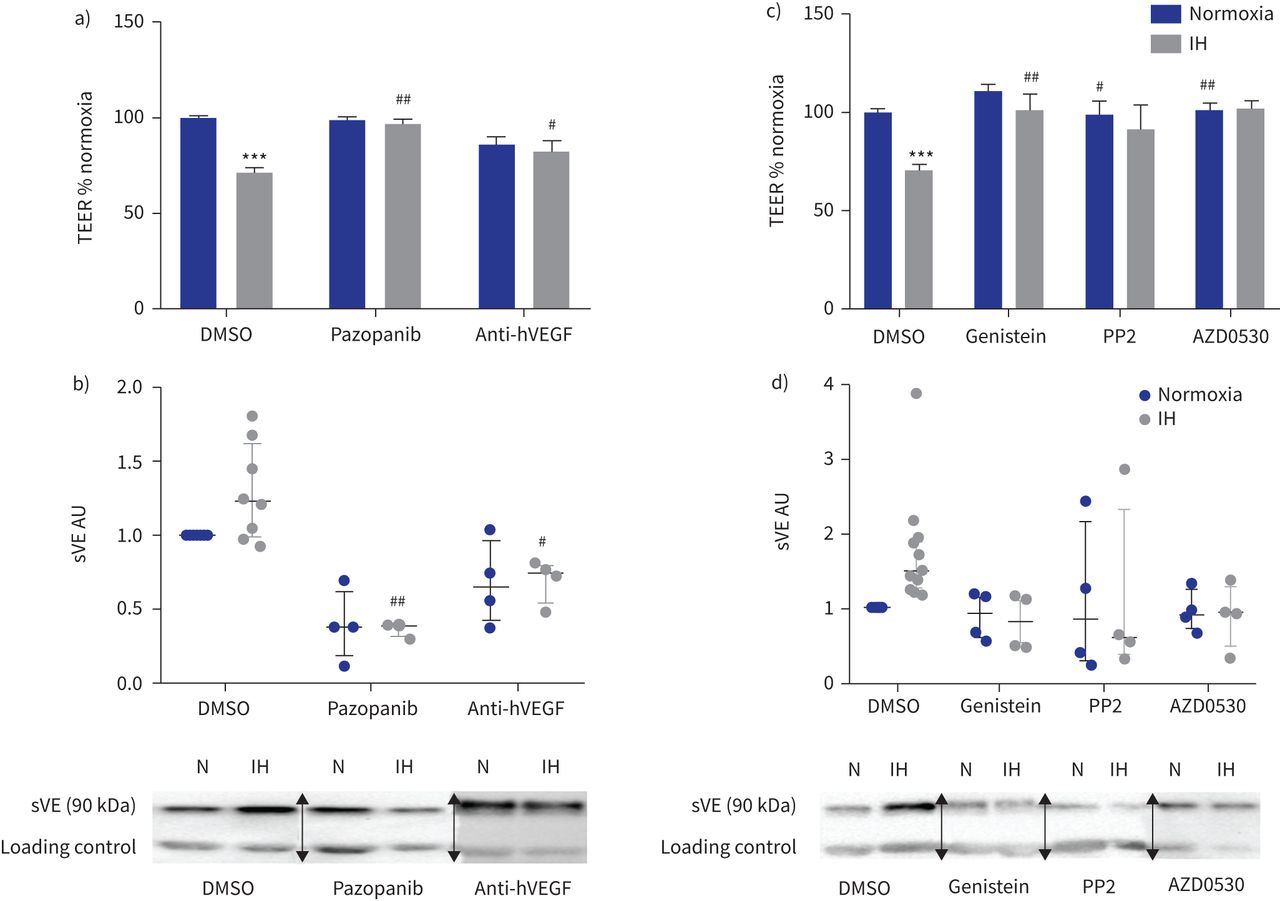
此外,抑制VEGF受体酪氨酸激酶活性的pazopanib或使用了反VEGF抑制性抗体都废除了间歇性低氧对内皮渗透性的影响和VE-cadherin劈理(gydF4y2Ba图5一个gydF4y2Ba和b)。gydF4y2Ba
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
的影响抑制血管内皮生长因子(VEGF)信号和酪氨酸激酶抑制内皮屏障间歇性缺氧(IH)条件下。te: transendothelial电阻;DMSO:二甲亚砜;h:人类;sVE:可溶;HAEC:人类主动脉内皮细胞。由pazopanib)抑制VEGF受体酪氨酸激酶(µg·5毫升gydF4y2Ba−1gydF4y2Ba)和阻塞hVEGF anti-hVEGF抑制性抗体(0.5µg·毫升gydF4y2Ba−1gydF4y2Ba)防止IH对内皮渗透性的影响评估三通。* * *:p < 0.001gydF4y2Ba与gydF4y2Banormoxia + DMSO溶液;gydF4y2Ba# #gydF4y2Ba:p < 0.01gydF4y2Ba#gydF4y2Ba:p < 0.05gydF4y2Ba与gydF4y2BaIH + DMSO溶液;19 transwells双向方差分析,从四个独立的实验。数据意味着±gydF4y2Ba扫描电镜gydF4y2Ba。b)上层清液的sVE测量水平的量化HAECs VEGF信号处理抑制剂。gydF4y2Ba# #gydF4y2Ba:p < 0.01gydF4y2Ba#gydF4y2Ba:p < 0.05gydF4y2Ba与gydF4y2BaIH + DMSO溶液;双向方差分析,n = 4。数据中位数和四分位范围。下面是代表西方墨点法的例子显示那么激烈sVE乐队HAECs的上层清液治疗婴幼儿血管瘤+ VEGF信号抑制剂相比,IH + DMSO(一种免疫球蛋白乐队作为加载控制正常化)。N: normoxia。c)抑制酪氨酸激酶和高金雀花碱(30µM)的Src激酶PP2(10µM)和AZD0530(µM saracatinib, 1)防止由三通IH对内皮渗透性的影响评估。* * *:p < 0.001gydF4y2Ba与gydF4y2Banormoxia + DMSO溶液;gydF4y2Ba# #gydF4y2Ba:p < 0.01gydF4y2Ba#gydF4y2Ba:p < 0.05gydF4y2Ba与gydF4y2BaIH + DMSO溶液;双向方差分析,16 transwells从至少四个独立的实验。数据意味着±gydF4y2Ba扫描电镜gydF4y2Ba。d) sVE水平的量化测量HAEC浮层显示由酪氨酸激酶抑制剂治疗废除IH VE-cadherin乳沟的效果。n = 4。数据中位数和四分位范围。下面是代表西方墨点法的例子显示那么激烈sVE乐队的上层清液HAECs酪氨酸激酶抑制剂治疗+ IH而DMSO + IH(一种免疫球蛋白乐队作为加载控制正常化)。N: normoxia。在西方的屁股,箭头表示图像不同部分的分组相同的凝胶或从不同的凝胶。gydF4y2Ba
最后,我们研究了Src家庭酪氨酸激酶的作用效果间歇性缺氧引起的。染料木黄酮、广谱的酪氨酸激酶抑制剂,以及PP2 AZD0530 (saracatinib),两个特定的Src激酶抑制剂,封锁了间歇性低氧诱导减少三通和VE-cadherin劈理(gydF4y2Ba图5 cgydF4y2Ba和d)。gydF4y2Ba
讨论gydF4y2Ba
本研究首次调查的乳沟VE-cadherin睡眠呼吸暂停患者的具体参与不同间歇hypoxia-activated通路底层内皮通透性的增加。我们演示了sVE的高水平的血清健康志愿者没有任何混杂因素在接触间歇性缺氧和阻塞性睡眠呼吸暂停综合症的病人没有任何已知的心血管疾病。这个海拔血清sVE与AHI,逆转CPAP治疗只在阻塞性睡眠呼吸暂停综合症组依从性高。在培养细胞暴露于间歇性缺氧,我们证明了经典信号通路参与VE-cadherin劈理(VEGF和Src家庭酪氨酸激酶激活)以及已知通路被激活的间歇性缺氧(ROS和HIF-1信号)与间歇性低氧诱导内皮屏障功能障碍。gydF4y2Ba
VE-cadherin乳沟的描述在阻塞性睡眠呼吸暂停综合症患者由于间歇性缺氧是原始的和新的。在多变量分析中,增加的范围sVE在阻塞性睡眠呼吸暂停综合症是近40%的水平,这是肯定的临床相关性。存在sVE血清中是公认的一个标志在几个心血管内皮功能障碍和炎症性疾病(审查,请参阅[gydF4y2Ba16gydF4y2Ba),包括动脉粥样硬化(gydF4y2Ba30.gydF4y2Ba]。VE-cadherin分裂导致的动荡和粘合连接处并且增加内皮渗透性。这内皮杂乱无章,渗透率增加是一个早期的硬化过程中的步骤。在临床研究中,患者常常没有已知心血管并发症的发生与睡眠呼吸暂停和我们的研究结果表明,在这个人口sVE可能代表一个内皮功能障碍的早期生物标志物暴露的起始过程。CPAP治疗对减少事件的影响晚期心血管事件仍争论不休,大多数负设计良好的随机对照试验(审查,请参阅[gydF4y2Ba31日gydF4y2Ba])。包含在这些试验的患者表现出普遍的心血管疾病和任何利益的CPAP干预限制已经现有的不可逆损伤。我们的结果表明,CPAP治疗6个月显著降低VE-cadherin乳沟在阻塞性睡眠呼吸暂停综合症患者中,施加在患者内皮功能保护作用在心血管功能障碍的早期发展阶段。这是重大的领域正在拼命寻找生物标记物预测CPAP心血管反应,以个性化处方(gydF4y2Ba32gydF4y2Ba]。gydF4y2Ba
阻塞性睡眠呼吸暂停综合症的特点是间歇性缺氧与睡眠相关碎片和增加睡眠时呼吸努力。在这些组件中,我们的数据表明,间歇性缺氧可能是主要的因素参与VE-cadherin乳沟。在健康志愿者接触间歇性缺氧模型没有任何混杂因素有力证明血清sVE水平的增加。尽管少量的参与者,我们能够证明显著增加sVE的配对分析,表明这种影响是均匀的患者中,符合我们的假设和另一个gydF4y2Ba在体外gydF4y2Ba和gydF4y2Ba在活的有机体内gydF4y2Ba结果。因此,间歇性缺氧应用于内皮细胞在文化也导致VE-cadherin乳沟。虽然以前的出版物表明膜的表达VE-cadherin减少间歇性缺氧后人工培养的内皮细胞(gydF4y2Ba9gydF4y2Ba),所涉及的机制(gydF4y2Ba即。gydF4y2Ba掩饰或劈理)从来没有调查。我们首次展示,VE-cadherin裂解gydF4y2Ba在活的有机体内gydF4y2Ba在阻塞性睡眠呼吸暂停综合症的病人,以及在人工培养的内皮细胞间歇缺氧后,释放可溶性片段在细胞上清液以及病人的血液中。关于分子机制的级联,阻塞性睡眠呼吸暂停患者的多变量分析发现,VEGF往往是独立与sVE水平相关(p = 0.06)。这些发现符合已知的机制VE-cadherin卵裂后VEGF结合受体(gydF4y2Ba18gydF4y2BaVEGF和支持这个假设,是阻塞性睡眠呼吸暂停患者的升高(gydF4y2Ba6gydF4y2Ba,gydF4y2Ba21gydF4y2Ba,gydF4y2Ba22gydF4y2Ba),可能是一个关键细胞因子信号VE-cadherin乳沟。这证实了证明抗VEGF抗体阻断和抑制剂VEGF受体酪氨酸激酶活性的(pazopanib)阻止了间歇性低氧诱导sVE释放和内皮渗透性gydF4y2Ba在体外gydF4y2Ba。同样,酪氨酸激酶抑制剂,非特异性(染料木素)或针对Src家族酪氨酸激酶(PP2和saracatinib),能够防止间歇hypoxia-initiated后果。最后,我们表明,间歇性缺氧诱导增加VE-cadherin Y685酪氨酸磷酸化,确认间歇性缺氧的主要信号通路激活导致sVE释放(gydF4y2Ba15gydF4y2Ba,gydF4y2Ba19gydF4y2Ba]。gydF4y2Ba
由于VEGF HIF-1转录因子的一个目标,这是众所周知的,被激活在阻塞性睡眠呼吸暂停综合症的人在小鼠模型的间歇性缺氧(审查,请参阅[gydF4y2Ba33gydF4y2Ba]),我们HIF-1参与我们的调查gydF4y2Ba在体外gydF4y2Ba模型。目前没有一个HIF-1α抑制剂是真正具体。我们因此使用两个不同的抑制剂,2-methoxyestradiol (HIF-1抑制剂没有任何动情的活动)(gydF4y2Ba34gydF4y2Ba,gydF4y2Ba35gydF4y2Ba)和吖啶黄(gydF4y2Ba36gydF4y2Ba),加强我们的方法,证明了HIF-1间歇性低氧诱导VE-cadherin乳沟。gydF4y2Ba
最后,在阻塞性睡眠呼吸暂停综合症和间歇性缺氧,HIF-1是稳定和激活hypoxia-reoxygenation重复周期和相关ROS生产(gydF4y2Ba33gydF4y2Ba,gydF4y2Ba37gydF4y2Ba]。我们因此使用一种超氧化物歧化酶模拟物,Tempol,证明ROS确实是间歇性低氧诱导sVE释放和内皮通透性的必要条件。我们的研究结果是一致的结果gydF4y2BaakarenkogydF4y2Baet al。gydF4y2Ba(gydF4y2Ba9gydF4y2Ba)表明,间歇性低氧诱导的渗透率,降低VE-cadherin膜表达都依赖ROS。有趣的是,除了已知的激活HIF-1活性氧,活性氧也被认为是直接激活酪氨酸激酶(gydF4y2Ba28gydF4y2Ba]。HIF-1-independent途径可能参与了间歇性低氧对内皮细胞通透性的影响。同样,我们不能排除其他细胞因子和路径可能导致VE-cadherin乳沟在我们的模型中,比如TNF-α[gydF4y2Ba15gydF4y2Ba]。这需要在将来的研究中探索。gydF4y2Ba
最后,我们表明,间歇性缺氧诱发VE-cadherin乳沟至少部分由活性氧,HIF-1、VEGF和酪氨酸激酶通路。sVE-cadherin碎片被发现在健康志愿者暴露于高浓度间歇性缺氧以及阻塞性睡眠呼吸暂停综合症的病人,并且可能提供了一个创新的生物标志物内皮通透性增加和相关疾病的早期阶段。gydF4y2Ba
补充材料gydF4y2Ba
补充材料gydF4y2Ba
请注意:gydF4y2Ba补充材料并不是由编辑部,编辑和上传已由作者提供。gydF4y2Ba
视觉抽象。gydF4y2Baerj - 04518 - 2020. -抽象gydF4y2Ba
补充材料。gydF4y2Baerj - 04518 - 2020. -补充gydF4y2Ba
补充图S1。间歇性缺氧改变了内皮屏障。这是一样的gydF4y2Ba图2gydF4y2Ba的类型,除了图:手段和SEM取而代之的是散点图中值。)代表的例子免疫印迹显示更强烈的上层清液sVE-band HAECs暴露IH比N(免疫球蛋白乐队被用作加载控制正常化)和量化sVE以HAEC上层清液后6 h (IH) (*: p < 0.05 vs . N,学生学习任务,N = 15)。b)免疫荧光和anti-VE-cadherin染色的量化,与抗体针对VE-cadherin细胞外域,显示减少表达的膜后IH (* *: p = 0.01 IH vs . N, Mann-Whitney等级和测试中,N = 4独立实验和N = 10 - 14每实验细胞)。酒吧规模:20μm。c)增加内皮渗透性衡量三通(* * *:p < 0.001 vs . N, Welch-test, N = 16 transwells从N = 8独立实验)后6 h (IH)。d)增加内皮渗透性衡量FITC-Dextran transendothelial通道(* *:p < 0.01 vs . N, Welch-test, N = 33 transwells从N = 12个独立实验)。值与中值报告为散点图。gydF4y2Baerj - 04518 - 2020. - figure_s1gydF4y2Ba
补充图S2。量化方法HAEC VE-cadherin荧光的细胞。整个内皮细胞被手动定义使用图像J的多边形工具软件(白线)和总荧光(T)测量。然后,荧光的细胞溶质(C)是由相同的方式测量(黄线)。背景是减去从措施和细胞膜荧光(M)计算(M = T - C)。意思是荧光得到总荧光除以面积。对于每一个实验,意味着在IH M细胞中表达意味着在常氧M细胞的百分比。gydF4y2Baerj - 04518 - 2020. - figure_s2gydF4y2Ba
可共享的PDFgydF4y2Ba
确认gydF4y2Ba
我们感谢艾莉森·富特(格勒诺布尔阿尔卑斯大学医院,法国格勒诺布尔)对批判性阅读和英语编辑,和Meriem Benmerad(格勒诺布尔阿尔卑斯大学医院)统计分析支持。gydF4y2Ba
脚注gydF4y2Ba
可以从本文的补充材料gydF4y2Bawww.qdcxjkg.comgydF4y2Ba
这篇文章一篇社论评论:gydF4y2Bahttps://doi.org/10.1183/13993003.01169 - 2021gydF4y2Ba
作者的贡献:a . Briancon-Marjollet o . Harki J-L。Pepin和i Vilgrain构思。r . Tamisier J-L。Pepin招募病人。o . Harki b . Gonthier Mahmani和a . Briancon-Marjollet进行了实验。o . Harki Gonthier, a . Mahmani Bailly和a . Briancon-Marjollet分析数据。o . Harki a . Briancon-Marjollet g . Faury J-L。Pepin起草了手稿。所有作者修订和批准了手稿。gydF4y2Ba
利益冲突:o . Harki没有披露。gydF4y2Ba
利益冲突:r . Tamisier报告奖助金、讲座和其他个人费用(差旅补助)Agiradom(医疗服务提供者),从ResMed赠款和个人费用,从激发赠款,指南针和爵士乐医药、个人费用从Elivie和飞利浦讲座,在提交工作。gydF4y2Ba
利益冲突:J-L。Pepin报告从Agiradom赠款和个人费用,阿斯利康,飞利浦和ResMed,从液化空气集团基金会赠款,费舍尔和Paykel Mutualia, Vitalaire,勃林格殷格翰的发言,爵士乐制药、平衡和Sefam,所有外提交的工作。gydF4y2Ba
贝利的利益冲突:美国没有披露。gydF4y2Ba
利益冲突:a . Mahmani没有披露。gydF4y2Ba
利益冲突:b Gonthier没有披露。gydF4y2Ba
利益冲突:a .所罗门没有披露。gydF4y2Ba
利益冲突:即Vilgrain没有披露。gydF4y2Ba
利益冲突:g . Faury没有披露。gydF4y2Ba
利益冲突:a . Briancon-Marjollet没有披露。gydF4y2Ba
支持声明:这项工作是由大学格勒诺布尔阿尔卑斯,由基金会Agir倒les疾病Chroniques (APMC), du杂音基金会,基金会倒说是在桑特Respiratoire (o . Harki格兰特博士),和德未来基金会。J-L。贝利Pepin, s, r . Tamisier由法国国家研究机构支持的框架“Investissements d未来”计划(ANR-15-IDEX-02)和“e-health和综合护理和轨迹医学和相亲人工智能“椅子的卓越格勒诺布尔大学基金会阿尔卑斯。这部分工作是由相亲@大学格勒诺布尔阿尔卑斯(anr - 19 - p3ia - 0003)。资金信息,本文已沉积的gydF4y2BaCrossref资助者注册表gydF4y2Ba。gydF4y2Ba
- 收到了gydF4y2Ba2020年12月14日。gydF4y2Ba
- 接受gydF4y2Ba2021年2月24日。gydF4y2Ba
- 版权©2021年作者。生殖权利和权限接触gydF4y2Ba权限在}{ersnet.orggydF4y2Ba
引用gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba






