





文摘gydF4y2Ba
上皮细胞管,由两极分化的上皮细胞腔,对器官功能是至关重要的。然而,管形成的分子机制在很大程度上仍未知。在这里,我们报告的功能fibrillin (FBN) 2、细胞外基质(ECM)糖蛋白,气管导管的关键调节器的形成。gydF4y2Ba
我们进行了大规模的遗传屏幕鼠标确定呼吸器官发育和疾病的监管机构。我们确定了gydF4y2BaFbn2gydF4y2Ba突变体表现出较短和缩小气管以及缺陷气管平滑肌细胞排列和极性。gydF4y2Ba
我们发现FBN2对弹性纤维的形成至关重要,在气管平滑肌细胞纤连蛋白的积累。这些过程监管似乎至少部分通过抑制p38-mediated upregulation的基质金属蛋白酶(MMPs),药理减少p38磷酸化或MMP的活动部分减毒gydF4y2BaFbn2gydF4y2Ba突变体气管表型。人类气管组织分析表明,减少ECM蛋白质,包括FBN2和纤连蛋白,与气管软化有关。gydF4y2Ba
我们的研究结果提供了新颖的见解ECM内稳态的作用在气管tubulogenesis间充质细胞分化。gydF4y2Ba
文摘gydF4y2Ba
细胞外基质形成缺陷导致气管狭窄的气管平滑肌组织改变,和药理减少p38磷酸化或基质金属蛋白酶活动部分减弱这些缺陷gydF4y2Bahttp://ow.ly/4zku30mWub3gydF4y2Ba
介绍gydF4y2Ba
平滑肌细胞周围上皮和内皮细胞管在许多器官系统,包括消化道、外分泌腺体、肺、肾脏和脉管系统(gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2gydF4y2Ba]。最近的数据表明,平滑肌细胞所需tubulogenesis在肺、肠道和血管gydF4y2Ba3gydF4y2Ba- - - - - -gydF4y2Ba5gydF4y2Ba]。气管是一个典型的上皮细胞管背包围平滑肌收缩功能提供弹性和控制气管(gydF4y2Ba6gydF4y2Ba]。气管上皮细胞来源于腹前肠内胚层和指定胚胎天(E) 8.25和E9.0之间,其次是分离后从背食道E9.5 [gydF4y2Ba7gydF4y2Ba]。气管平滑肌和软骨来源于周围的内脏中胚层。平滑肌细胞分化在E11.5gydF4y2Ba6gydF4y2Ba],而软骨开发启动最早E9 [gydF4y2Ba8gydF4y2Ba由E13.5[],显示冷凝gydF4y2Ba9gydF4y2Ba,gydF4y2Ba10gydF4y2Ba)和分化E13.5 E15.5 (gydF4y2Ba11gydF4y2Ba]。气管上皮细胞启动平滑肌和软骨的分化成立后(gydF4y2Ba12gydF4y2Ba- - - - - -gydF4y2Ba15gydF4y2Ba]。已报告在人类中,气管导管形成缺陷造成气管狭窄或气管软化,它的特点是一个狭窄的气管腔或缺乏支持软骨,分别和可能导致呼吸困难和死亡gydF4y2Ba16gydF4y2Ba,gydF4y2Ba17gydF4y2Ba]。细胞和分子机制研究气管tubulogenesis大多集中在上皮细胞的作用[gydF4y2Ba18gydF4y2Ba,gydF4y2Ba19gydF4y2Ba)以及上皮和间质之间的复杂信号gydF4y2Ba9gydF4y2Ba,gydF4y2Ba20.gydF4y2Ba,gydF4y2Ba21gydF4y2Ba]。然而,在气管平滑肌和软骨管的角色形成仍然知之甚少。gydF4y2Ba
另一个鲜为人知的方面管发展的角色是细胞外基质(ECM)由平滑肌细胞和软骨细胞分泌。最近的数据表明,ECM线索通过调解控制管形成上皮细胞分化[gydF4y2Ba22gydF4y2Ba,gydF4y2Ba23gydF4y2Ba和重排gydF4y2Ba24gydF4y2Ba),或定向细胞间张力gydF4y2Ba25gydF4y2Ba]。值得注意的是,在我们的研究中,ECM的角色在气管平滑肌细胞排列和极性tubulogenesis尚未解决。gydF4y2Ba
材料和方法gydF4y2Ba
实验动物gydF4y2Ba
30 C57BL / 6 j雄性老鼠接受一个3×100毫克公斤gydF4y2Ba−1gydF4y2Ba剂量的ethylnitrosourea (ENU表示)gydF4y2Ba26gydF4y2Ba从莫妮卡正义)得到(美国贝勒医学院,休斯顿,德克萨斯州)。经过一段时间的10周的恢复生育能力,越过了mutagenised G0雄性雌性小鼠C57BL / 6 j。G1男性与C57BL / 6 j outcrossed女性产生G2女性。四个G2雌性backcrossed G1的父亲,以及由此产生的G3产后一天(P) 0小狗受到气管和肺部解剖和分析。繁殖地都维持在12 h光和12 h暗周期。所有小鼠饲养在标准条件下按照制度执行(马克斯·普朗克协会)和当地伦理委员会(达姆施塔特Regierungsprasidium,黑森州,德国)。动物实验都做符合道德准则和批准的协议。gydF4y2Ba
Whole-exome测序分析和基因分型gydF4y2Ba
基因组DNA分离出两个野生型(WT)和两个突变小鼠中,使用标准的协议使用SureSelect鼠标捕获所有外显子装备V1(美国安捷伦,圣克拉拉,CA),并使用HiSeq 2000测序(Illumina公司、圣地亚哥、钙、美国),最低平均50×目标序列覆盖率(BGI-Hong香港)。排列的顺序读取C57BL / 6 j小鼠参考基因组(mm10)和分析使用CLCBio基因组工作台(试剂盒、希尔登,德国),和GATK软件。减少假阴性,变体电话是设定在5×最小覆盖和≥20%替代读取。序列变异被注释的单核苷酸多态性(snp)对dbSNP128 dbSNP(142年版)和过滤。基因分型结果为gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba老鼠,引物Fwd (5′-TCCAGTCATGTCGAGGGCTA-3′)和牧师(5′-CCAGCTTCTGCTCTTCTGCT-3′)被用来生成一个∼420 - bp PCR扩增子。使用引物PCR产品排序Fwd (5′-TCCAGTCATGTCGAGGGCTA-3′)和使用生物系统序列突变分析扫描仪软件(版本2.0;福斯特城、钙、美国)。gydF4y2BaFbn2gydF4y2Ba零老鼠一直在前面描述的(gydF4y2Ba27gydF4y2Ba]。基因分型结果为gydF4y2BaFbn2gydF4y2Ba如前所述(执行零老鼠,PCRgydF4y2Ba28gydF4y2Ba]。gydF4y2Ba
阿尔新蓝染色的软骨gydF4y2Ba
wholemount染色的气管软骨,解剖气管被固定在95%乙醇12 h与0.03%其次是隔夜染色阿尔新蓝溶解在80%乙醇和20%醋酸。样本清理2%氢氧化钾(KOH)。gydF4y2Ba
Wholemount染色的四肢gydF4y2Ba
执行wholemount染色E13.5前肢和后肢骨骼gydF4y2Ba29日gydF4y2Ba),样本孵化在4°C一夜之间70%的乙醇,95%乙醇1 h和丙酮在室温下过夜。样本然后在0.03%阿尔新蓝染色过夜,清除1% KOH 12 h和孵化50%甘油:50% - 1% KOH溶液在室温下48 h,使组织透明。执行wholemount染色P6的前肢和后肢骨骼,样本在热水浸泡30年代在65°C到删除所有皮肤,在一夜之间95%的乙醇和放置在丙酮孵化12 h在室温下。样本在一夜之间在0.03%阿尔新蓝染色,洗两次在一夜之间在70%乙醇和95%乙醇中孵化。样本在1% KOH 1 h和孵化van buren 0.005%茜素红2 h在室温下。样本保存在50%甘油:50% - 1% KOH成像解决方案。gydF4y2Ba
小鼠气管软骨细胞的主要文化gydF4y2Ba
气管被隔绝E14.5胚胎。分离软骨细胞(gydF4y2Ba30.gydF4y2Ba),每个气管孵化在10毫升的胶原酶D为0.5 mg·毫升的解决方案gydF4y2Ba−1gydF4y2Ba12 h在37°C。样本然后通过2毫升巴斯德吸量管驱散任何细胞聚集。细胞悬液的无菌过滤40μm细胞在50毫升管过滤器和离心机在400×10分钟gydF4y2BaggydF4y2Ba在室温下。的颗粒被2毫升PBS。从每个气管软骨细胞被播种在24-well DMEM / F-12培养基培养细胞培养板和含10%胎牛血清(的边后卫)37°C 5%二氧化碳(有限公司gydF4y2Ba2gydF4y2Ba)孵化器为9天。gydF4y2Ba
阿尔新蓝染色气管软骨细胞培养gydF4y2Ba
小鼠气管软骨细胞在初级文化洗两次在PBS和固定在1毫升4%戊二醛在室温下15分钟。样本洗两次2毫升0.1氯化氢(HCl)其次是阿尔新蓝染色与1%盐酸溶解在0.1 30分钟。后三个洗2毫升的0.1 M盐酸,样本干和水化1毫升的0.1 M盐酸。gydF4y2Ba
量化的相对阿尔新蓝区gydF4y2Ba
阿尔新蓝面积测量使用ImageJ (gydF4y2Bahttp://rsbweb.nih.gov/ij/gydF4y2Ba)。相对阿尔新蓝区被阿尔新蓝的比例计算区总细胞区、WT被指定为1。gydF4y2Ba
气管导管长度测量gydF4y2Ba
气管导管长度决定通过测量第一个和最后一个气管软骨环之间的距离。gydF4y2Ba
呼吸速率的测量gydF4y2Ba
呼吸率取决于测量每分钟呼吸的次数。出乎意料,gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba老鼠展览WT-like呼吸率在休息的时候。它是可能的gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba老鼠将显示在高强度锻炼呼吸困难。gydF4y2Ba
Wholemount疣状gydF4y2Ba
气管被隔绝E12.5 E16.5胚胎,P0和P60老鼠。气管被固定在4%多聚甲醛在一夜之间在4°C和水洗三次PBS 1 h。样本孵化在5%的边后卫Triton X-100/3% / PBS / 0.5%牛血清白蛋白12 h,然后一夜之间在初级抗体在4°C。六洗后在0.5% Triton x - 100 / PBS 1 h,样本孵化在二级抗体在一夜之间在4°C。样本洗6次0.5% Triton x - 100 / PBS 1 h和安装成像。想象平滑肌细胞内层,气管被染色α-smooth肌肉肌动蛋白(SMA)和SOX9,分别。高尔基体的可视化,气管是GM130染色。想象弹性纤维,纤连蛋白(FN),气管被染色弹性蛋白原和FN。gydF4y2Ba
量化的弹性纤维gydF4y2Ba
短的弹性纤维的数量清点每使用ImageJ视野。gydF4y2Ba
cryosections组织化学染色gydF4y2Ba
气管和肺解剖在PBS、固定在4%多聚甲醛在一夜之间在4°C,在10%的蔗糖和30%的蔗糖孵化24小时都在4°C,安装在最佳切削温度(10月)嵌入化合物,和分段10µm厚度。疣状进行了使用标准的协议。疣状phospho-p38和执行α-SMA WT和突变气管部分在同一幻灯片。gydF4y2Ba
量化的SFTPCgydF4y2Ba+gydF4y2Ba细胞比例gydF4y2Ba
表面活性剂蛋白C (SFTPC)gydF4y2Ba+gydF4y2Ba总细胞数是计算使用ImageJ每视野。SFTPCgydF4y2Ba+gydF4y2BaSFTPC细胞比率计算gydF4y2Ba+gydF4y2Ba细胞/总细胞。gydF4y2Ba
有丝分裂的量化的角度gydF4y2Ba
上皮平面由白色虚线标记,和有丝分裂的轴绿色虚线。这两条线之间的角度是有丝分裂角度和使用ImageJ测量。gydF4y2Ba
原位gydF4y2Ba杂交的cryosectionsgydF4y2Ba
气管在PBS解剖,一夜之间在4%多聚甲醛固定在4°C,安装在10月嵌入化合物,和分段10µm厚度。执行gydF4y2Ba原位gydF4y2Ba杂交(gydF4y2Ba31日gydF4y2Ba),cryosections permeabilisedμg·5毫升gydF4y2Ba−1gydF4y2Ba蛋白酶K (04693159001;美国罗氏,印第安纳波利斯)在室温下15分钟,其次是乙酰化作用的2分钟和pre-incubation杂交缓冲3 h在70°C,孵化与digoxigenin(挖)标记反义RNA探针在一夜之间在70°C,水洗,孵化与碱性phosphatase-conjugated anti-digoxigenin抗体(罗氏)一夜之间在4°C,洗,信号检测与电视台/ BCIP染色(罗氏)的解决方案。gydF4y2Ba
西方墨点法gydF4y2Ba
孤立的P7气管细胞溶解使用缓冲区里帕(9806;细胞信号技术,丹弗斯,美国马)补充蛋白酶和磷酸酶抑制剂(5872;细胞信号技术)。溶菌产物在000×离心机gydF4y2BaggydF4y2Ba10分钟,sds - page和转移到硝化纤维膜。探讨了膜主要和辣根peroxidase-conjugated二级抗体(细胞信号技术),使用一个增强化学发光检测系统开发(美国皮尔斯,罗克福德,IL)。gydF4y2Ba
明胶zymographygydF4y2Ba
孤立P7气管是裂解缓冲(0.025 Tris-HCl, pH值7.5,0.1 M氯化钠(氯化钠)),和1% v / v诺乃清洁剂p 40补充了蛋白酶抑制剂Cocktail-EDTA免费(04693159001;罗氏公司)。溶菌产物离心机在16 000 g×10分钟在4°C。上层清液收集和蛋白质浓度测量。等量的蛋白质被加载每车道明胶zymography凝胶。复性的凝胶被孵化的解决方案在室温下30分钟,在300毫升蒸馏水洗两次(dHgydF4y2Ba2gydF4y2BaO),发展中缓冲30分钟和孵化的孵化新的发展中缓冲16 h。与Coomassie蓝染色后,这种凝胶是检查gelatinolytic活动使退色。gydF4y2Ba
量化的免疫印迹的信号gydF4y2Ba
纤连蛋白、活性基质金属蛋白酶(MMP) 2、MMP9,总p38、phospho-p38和3 -磷酸甘油醛脱氢酶(GAPDH)使用ImageJ水平量化。纤连蛋白、活性MMP2、MMP9、总p38和phospho-p38水平正常化GAPDH带来的价值。Phospho-p38褶皱变化是计算的比率Phospho-p38 / p38和WT被分配到1。gydF4y2Ba
定量逆转录聚合酶链反应gydF4y2Ba
总RNA提取使用miRNeasy迷你包(试剂盒)。互补脱氧核糖核酸是使用最大值合成第一链cDNA合成装备(美国马ThermoFisher科学、沃尔瑟姆),根据制造商的指示。定量实时PCR进行使用Eco实时PCR系统(Illumina公司)和Maxima SYBR绿色/荧光素qPCR大师混合(ThermoFisher科学)。平均Ct值所示gydF4y2Ba补充表1gydF4y2Ba。gydF4y2BaActbgydF4y2Ba被用作参考基因。以下引物被使用:gydF4y2BaActbgydF4y2Ba提出5′-CGGCCAGGTCATCACTATTGGCAAC-3′gydF4y2BaActbgydF4y2Ba反向5′-GCCACAGGATTCCATACCCAAGAAG-3′;gydF4y2BaAtf2gydF4y2Ba提出5′-ACTCCAACGCCAACAAGATTC-3′gydF4y2BaAtf2gydF4y2Ba反向5′-TCTACAACAGAAGGCTCCTCAAT-3′;gydF4y2Ba”丛书gydF4y2Ba提出5′-GGGGACAGCCTTTCCTA-3′gydF4y2Ba”丛书gydF4y2Ba反向5′-CTGTCACCGTGGGGATAAAG-3′;gydF4y2Ba小君gydF4y2Ba提出5′-ACGACCTTCTACGACGATGC-3′gydF4y2Ba小君gydF4y2Ba反向5′-CCAGGTTCAAGGTCATGCTC-3′;gydF4y2BaFbn1gydF4y2Ba提出5′-ACGGCTTTACTGGACCCCA-3′gydF4y2BaFbn1gydF4y2Ba反向5′-ACATCTGGTTGCTTACCACAG-3′;gydF4y2BaFbn2gydF4y2Ba提出5′-CCACTCCTATTGCTGCCCAG-3′gydF4y2BaFbn2gydF4y2Ba反向5′-TTGGGGCGGGAACAGAATC-3′;gydF4y2Ba纤连蛋白gydF4y2Ba提出5′-GGCATCGGGGAGTGGCACTG-3′gydF4y2Ba纤连蛋白gydF4y2Ba反向5′-ATTGGGCTGGCTGGGGGTCT-3′;gydF4y2BaFgf10gydF4y2Ba提出5′-CGGGACCAAGAATGAAGACT-3′gydF4y2BaFgf10gydF4y2Ba反向5′-AGTTGCTGTTGATGGCTTTG-3′;gydF4y2BaHspb1gydF4y2Ba提出5′-ATCCCCTGAGGGCACACTTA-3′gydF4y2BaHspb1gydF4y2Ba反向5′-GGAATGGTGATCTCCGCTGAC-3′;gydF4y2BaMmp2gydF4y2Ba提出5′-ACCCAGATGTGGCCAACTAC-3′gydF4y2BaMmp2gydF4y2Ba反向5′-TACTTTTAAGGCCCGAGCAA-3′;gydF4y2BaMmp9gydF4y2Ba提出5′-TAAGGACGGCAAATTTGGTT-3′gydF4y2BaMmp9gydF4y2Ba反向5′-CTTTAGTGGTGCAGGCAGAG-3′;gydF4y2BaNfkb1gydF4y2Ba提出5′-CGTAACAGCAGGACCCAAGG-3′gydF4y2BaNfkb1gydF4y2Ba反向5′-CTGTCATCCGTGCTTCCAGT-3′;gydF4y2BaNfkb2gydF4y2Ba提出5′-TTCAGAGTTGGGAGTGTGCG-3′gydF4y2BaNfkb2gydF4y2Ba反向5′-TGGTTACATGCAGGACACCC-3′;gydF4y2BaRelagydF4y2Ba提出5′-CGGATTCCGGGCAGTGAC-3′gydF4y2BaRelagydF4y2Ba反向5′-GAGGGGAAACAGATCGTCCA-3′。gydF4y2Ba
GM6001 SB203580注射;gydF4y2Ba在体外gydF4y2Ba治疗SB203580和甲胺盐酸盐gydF4y2Ba
GM6001和SB203580溶解在100%二甲亚砜(DMSO)(股票),用0.9%的生理盐水稀释。甲胺盐酸盐是重蒸馏的水(ddH溶解gydF4y2Ba2gydF4y2BaO)(股票)。插入C57BL6 / J女性每日与腹腔内注射治疗GM6001 (50 mg·公斤gydF4y2Ba−1gydF4y2Ba)或SB203580(1μM·公斤gydF4y2Ba−1gydF4y2Ba)E12.5和E14.5之间。车辆用于控制注射DMSO溶液在0.9%生理盐水。为气管隔离E15.5胚胎收获。为gydF4y2Ba在体外gydF4y2Ba治疗SB203580,孤立的气管在DMEM / F-12培养基培养含有0.1% DMSO溶液或1μM SB203580 37°C的5%的股份有限公司gydF4y2Ba2gydF4y2Ba孵化器20 h。为gydF4y2Ba在体外gydF4y2Ba治疗甲胺盐酸盐,孤立的气管包含1.5% ddH DMEM / F-12培养基培养gydF4y2Ba2gydF4y2BaO或30μM甲胺盐酸盐在37°C公司5%gydF4y2Ba2gydF4y2Ba孵化器25 h。gydF4y2Ba
治疗后气管表型分析gydF4y2Ba
评价气管导管长度、平滑肌取向、弹性纤维形成和纤连蛋白积累GM6001 SB203580注入或治疗后进行了基因分型。gydF4y2Ba
抗体gydF4y2Ba
以下使用抗体:鼠标anti-α-SMA-Cy3 (1:1000 C6198;Sigma-Aldrich,圣路易斯,密苏里州,美国);老鼠anti-CDH1 (1:50 0, sc - 59778;美国圣克鲁斯生物技术、达拉斯、TX);兔子anti-SOX9 (1:400 AB5535;美国微孔,伯灵顿,马);山羊anti-SOX9 (1∶AF3075;研发系统,明尼阿波利斯,美国);羊anti-GM130 (1:50, AF8199;研发系统); rabbit anti-Ki67 (1:400, #9027; Cell Signaling Technologies); mouse anti-PCNA (1:400, sc-56; Santa Cruz Biotechnology); rabbit anti-Cleaved Caspase-3 (1:600, #9661; Cell Signaling Technologies); rabbit anti-Phospho-p38 (1:100 for immunostaining and 1:1000 for Western blots, #9215; Cell Signaling Technologies); rabbit anti-p38 (1:1000, #9212; Cell Signaling Technologies); rabbit anti-MMP-9 (1:1000, AB19016; Millipore); rabbit-MMP-2 (1:1000, AB19015; Millipore); rabbit anti-FBN2 (1:5000 for immunostaining and 1:2000 for Western blots); guinea-pig anti-FBN2 (1:500); rabbit anti-Tropoelastin (1:1000 for immunostaining and 1:1000 for Western blots, ab21600; Abcam, Cambridge, UK); sheep anti-Fibronectin (1:1000, AF1918; Novus Biologicals, Centennial, CO, USA); rabbit anti-Fibronectin (1:500 for immunostaining and 1:1000 for Western blots, F3648; Sigma); goat anti-Aggrecan (1:200, AF1220; R&D Systems); rabbit anti-COL2A1 (1:50, AB761; Millipore); rabbit anti-PH3 (1:400, 06-570; Millipore); rat anti-RAGE (1:400, MAB1179; R&D Systems); rat anti-PECAM (1:200, 553370; BD Biosciences, Franklin Lakes, NJ, USA); goat anti-CC10 (1:200, T-18; Santa Cruz); rabbit anti-SFTPC (1:400, AB3786; Millipore); mouse anti-acetylated α-tubulin (1:2000, T-7451; Sigma); goat anti-Vimentin (1:200, C-20; Santa Cruz); and rabbit anti-GAPDH (1:3000, #2118; Cell Signaling Technologies). Polyclonal rabbit and guinea-pig anti-Fibrillin-2 antisera were raised against the N-terminal recombinant human Fibrillin-2 polypeptide rF86 (Gln29日gydF4y2BaaspgydF4y2Ba535年gydF4y2Ba293年巴尔病毒核抗原(EBNA)细胞(gydF4y2Ba32gydF4y2Ba]。抗血清被亲和色谱法纯化使用前一列与抗原与氰bromide-activated琼脂糖(美国通用电气医疗集团,芝加哥,IL)。gydF4y2Ba
外植体培养小鼠胚胎气管和肺,gydF4y2Ba在体外gydF4y2Ba用蛋白酶治疗gydF4y2Ba
气管和肺E13.5胚胎中分离培养使用一个既定协议(gydF4y2Ba33gydF4y2Ba]。为gydF4y2Ba在体外gydF4y2Ba弹性蛋白酶和胶原酶治疗II型、孤立的气管和肺在DMEM培养/ F-12介质或DMEM / F-12介质包含2μ·毫升gydF4y2Ba−1gydF4y2Ba弹性蛋白酶(LS002290;沃辛顿生化公司,莱克伍德,美国新泽西)或DMEM / F-12中包含2μ·毫升gydF4y2Ba−1gydF4y2Ba胶原酶II型(9001-12-1;Gibco,沃尔瑟姆,美国马)37°C公司5%gydF4y2Ba2gydF4y2Ba孵化器28 h。gydF4y2Ba
体外gydF4y2Ba气管生理学gydF4y2Ba
2毫米的气管被隔绝P1幼崽,在柠檬酸溶液(4.7毫米119毫米氯化钠,氯化钾,2.5毫米CaClgydF4y2Ba2gydF4y2Ba,1.17毫米MgSOgydF4y2Ba4gydF4y2Ba,20毫米NaHCOgydF4y2Ba3gydF4y2Ba,1.18毫米KHgydF4y2Ba2gydF4y2Ba阿宝gydF4y2Ba4gydF4y2Ba0.027毫米EDTA 11毫米葡萄糖)充气与carbogen 37°C。气管环安装在wire-myograph系统(610米;丹麦Myo技术,丹麦奥尔胡斯)和2 mN的静息张力申请每个戒指作为基线。收缩反应测定累积管理表示乙酰胆碱浓度。胎儿气管的自发收缩分析,E13.5气管被孤立和保存在PBS延时成像10分钟每1秒蔡司LSM 800反向激光扫描共焦显微镜。的振幅自发收缩被定义为“1−气管平滑肌收缩后最小宽度的比值后最大宽度放松”。gydF4y2Ba
人类气管软化样本gydF4y2Ba
石蜡部分鉴定人类气管软化的样品和健康的气管组织(控制)提供的辛辛那提儿童医院医学中心生物核心设施(Cincinatti,哦,美国)。气管软化的临床诊断证实了组织学分析。研究方案和组织使用被批准的机构伦理委员会。书面知情同意了所有的病人家属之前收集的样本biorepository“儿童更好的结果”。gydF4y2Ba
成像gydF4y2Ba
成像的wholemount气管、肺和四肢,气管和肺部分进行使用尼康SMZ25,尼康90我,蔡司Axio成像仪。A2,蔡司LSM 800倒或880垂直激光扫描共焦显微镜。量化的管长度,距离位于环,环宽度、平滑肌,平滑肌细胞取向和核宽高比,高尔基体的位置相对于原子核,弹性纤维取向,短的弹性纤维,免疫荧光强度、细胞数量和使用ImageJ意味着线性拦截了。直和弯曲气管管长度测量通过徒手线沿外缘ImageJ气管的前后方向。gydF4y2Ba
统计分析gydF4y2Ba
统计分析使用GraphPad软件(美国圣地亚哥,CA)。假定值计算使用t (*: p < 0.05;* *:p < 0.01;* * *:p < 0.001;gydF4y2BansgydF4y2Ba:不重要的)。gydF4y2Ba
结果gydF4y2Ba
Fbn2gydF4y2BaT2547A / T2547AgydF4y2Ba老鼠表现出气管导管形成缺陷gydF4y2Ba
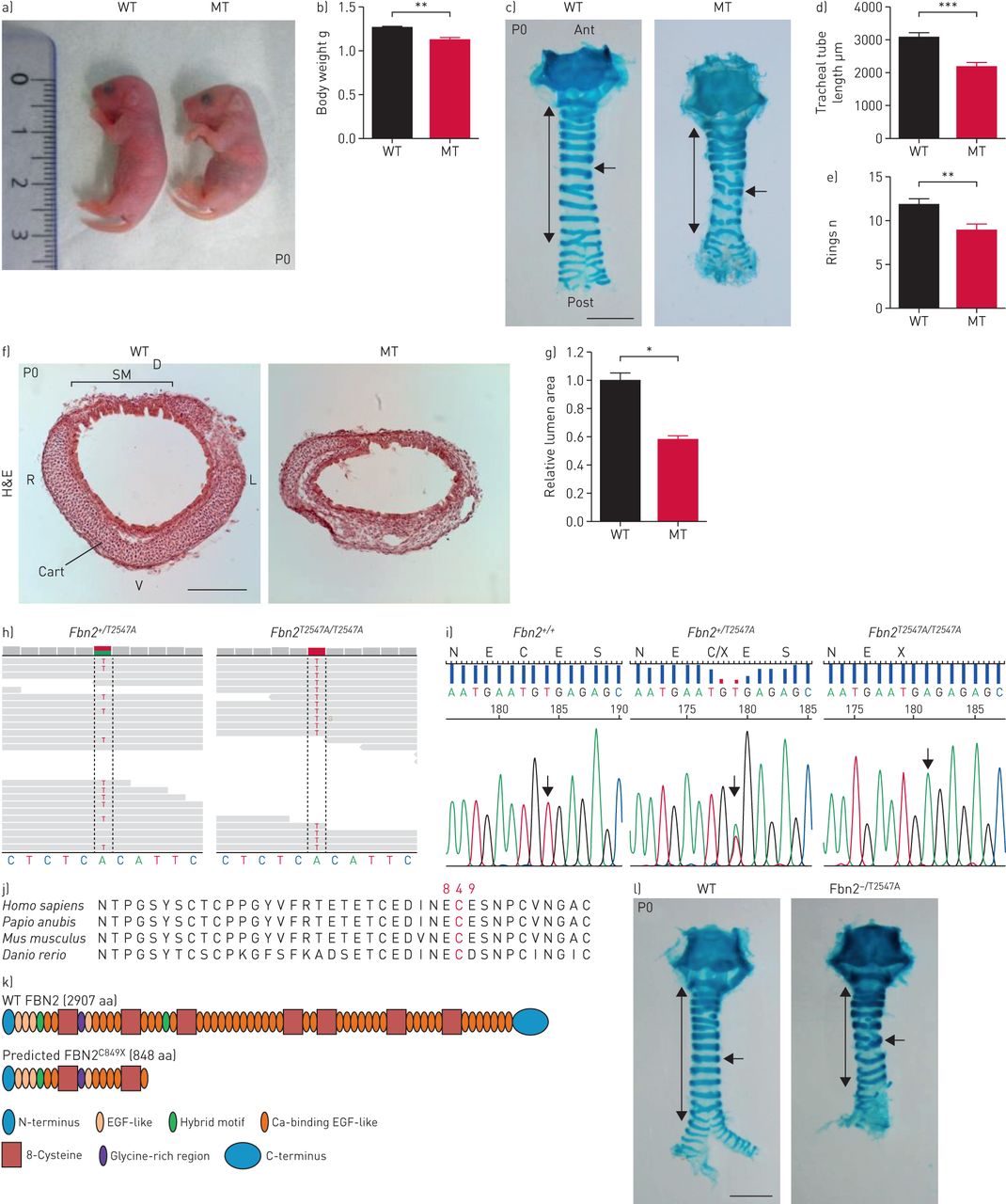
小说来识别位点调节小鼠呼吸器官的形成,我们进行了一次大规模的遗传屏幕使用ENU表示诱变。隐性突变体的发现在这个屏幕展品低体重(gydF4y2Ba图1一个gydF4y2Ba和b和gydF4y2Ba补充图S1a和bgydF4y2Ba),降低鼻肛门长度(gydF4y2Ba补充图S1a和cgydF4y2Ba),缩短气管(gydF4y2Ba图1 c和dgydF4y2Ba和gydF4y2Ba补充图S1dgydF4y2Ba),减少软骨环(gydF4y2Ba图1 cgydF4y2Ba和气管腔缩小(e)gydF4y2Ba图1 fgydF4y2Ba和g),但WT-like呼吸速率(gydF4y2Ba补充图S1egydF4y2Ba)。这些变异动物出生在预期的孟德尔式比例,表明这种突变不会引起胚胎杀伤力。识别phenotype-causing突变,我们执行whole-exome G4基因组DNA的测序样品和确认gydF4y2BaFbn2gydF4y2Ba编码ECM糖蛋白,作为一个候选基因(gydF4y2Ba图1 hgydF4y2Ba)。接下来,我们进行了基因分型的遗传连锁分析189 G4, G5和G6变异动物,发现完整的气管表型和之间的联系gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba等位基因(gydF4y2Ba图1我gydF4y2Ba)。确定等位基因的无义突变(c.T2547A)在一个高度保守的氨基酸(gydF4y2Ba图1 jgydF4y2Ba),这将导致截断和848个氨基酸蛋白质(p.C849X) (gydF4y2Ba图1 kgydF4y2Ba)。然后我们进行了补充试验跨越老鼠携带ENU-inducedgydF4y2BaFbn2gydF4y2Ba等位基因(gydF4y2BaFbn2gydF4y2BaT2547A / +gydF4y2Ba)与老鼠携带gydF4y2BaFbn2gydF4y2Ba删除等位基因(gydF4y2Ba27gydF4y2Ba),发现互补并没有发生gydF4y2BaFbn2gydF4y2Ba- - - - - -gydF4y2Ba/gydF4y2BaT2547AgydF4y2Badouble-heterozygous动物(gydF4y2Ba图1 lgydF4y2Ba),表明损失gydF4y2BaFbn2gydF4y2Ba函数是可能负责观察气管的表型gydF4y2BaFbn2gydF4y2BaT2547AgydF4y2Ba/gydF4y2BaT2547AgydF4y2Ba老鼠。为了进一步测试的作用gydF4y2BaFbn2gydF4y2Ba在气管开发中,我们分析了气管的形成gydF4y2BaFbn2gydF4y2Ba- - - - - -gydF4y2Ba/gydF4y2Ba- - - - - -gydF4y2Ba老鼠。gydF4y2BaFbn2gydF4y2Ba- - - - - -gydF4y2Ba/gydF4y2Ba- - - - - -gydF4y2Ba老鼠表现出缩短气管和软骨环骨折(较少gydF4y2Ba补充图S2a-cgydF4y2Ba)和组织平滑肌(gydF4y2Ba补充图S2c和dgydF4y2Ba)。gydF4y2BaFbn2gydF4y2Ba- - - - - -gydF4y2Ba/gydF4y2Ba- - - - - -gydF4y2Ba老鼠已报告展示骨骼肢体异常(gydF4y2Ba27gydF4y2Ba]。我们因此检查肢体形成gydF4y2BaFbn2gydF4y2BaT2547AgydF4y2Ba/gydF4y2BaT2547AgydF4y2Ba老鼠。有趣的是,gydF4y2BaFbn2gydF4y2BaT2547AgydF4y2Ba/gydF4y2BaT2547AgydF4y2Ba老鼠表现出两国中央数字并趾突起物最早E13.5 (gydF4y2Ba补充图S3agydF4y2Ba),直到成年期(gydF4y2Ba补充图S3b和cgydF4y2Ba)。然而,我们观察到没有明显的表型gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba前肢(gydF4y2Ba补充图S3d-fgydF4y2Ba)。测试可能的作用gydF4y2BaFbn2gydF4y2Ba在肺开发中,我们分析了肺内的气管和肺部。gydF4y2BaFbn2gydF4y2BaT2547AgydF4y2Ba/gydF4y2BaT2547AgydF4y2Ba老鼠表现出软骨形成缺陷在肺内的航空公司(gydF4y2Ba补充图S4a-dgydF4y2Ba)。然而,我们观察到在肺分支形态发生没有明显的表型(gydF4y2Ba补充图S5a和bgydF4y2Ba),alveologenesis (gydF4y2Ba补充图S5c和dgydF4y2Ba),或肺细胞分化gydF4y2Ba补充图S6a-igydF4y2Ba)gydF4y2BaFbn2gydF4y2BaT2547AgydF4y2Ba/gydF4y2BaT2547AgydF4y2Ba动物。此外,我们检查气管导管形成和肢体模式gydF4y2BaFbn2gydF4y2BaT2547A / +gydF4y2Ba动物。gydF4y2BaFbn2gydF4y2BaT2547A / +gydF4y2Ba老鼠展览WT-like气管长度(gydF4y2Ba补充图S7a和bgydF4y2Ba)、软骨的形成(gydF4y2Ba补充图S7a和cgydF4y2Ba)、平滑肌细胞取向(gydF4y2Ba补充图S7d和egydF4y2Ba)和肢体模式(gydF4y2Ba补充图S7fgydF4y2Ba),表明haplosufficiencygydF4y2BaFbn2gydF4y2Ba。总之,这些发现强烈建议gydF4y2BaFbn2gydF4y2Ba调节气管导管的形成。gydF4y2Ba
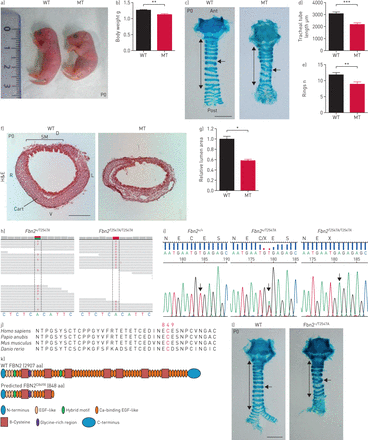
Fbn2gydF4y2BaT2547A / T2547AgydF4y2Ba老鼠表现出气管导管形成缺陷。总值)代表形态的产后的一天(P) 0野生型(WT) (n = 16)和突变体(MT) (n = 16)。b)量化的体重P0 WT (n = 15)和突变体(n = 15)。c)代表图像的腹侧的观点wholemount气管与阿尔新蓝染色P0 WT (n = 24)和突变体(n = 24)。双头箭头表示气管导管的长度。箭头指向气管软骨环。比例尺= 1毫米。d)量化P0 WT (n = 15)和突变(n = 15)气管导管的长度。e)量化气管软骨环的数量从P0 WT (n = 15)和突变体(n = 15)。f)代表图像的横向部分气管与苏木精和伊红染色(圆))从P0 WT (n = 5)和突变体(n = 5)。 Scale bar=200 μm. g) Quantification of tracheal lumen area from P0 WT (n=5) and mutants (n=5). h) Whole-exome sequencing of WT control (Fbn2gydF4y2Ba杂合的,n = 2)和突变(gydF4y2BaFbn2gydF4y2Ba纯合子,n = 2)样本;绿色表示WT核苷酸;红色代表WT的突变体核苷酸t i)序列和突变体基因组DNA损伤。DNA序列色谱显示TGT WT (n = 112)的半胱氨酸(左),TGT和TGA杂合的突变体,和TGA终止密码子的纯合突变体(n = 124)(右)。箭头指向突变。进化守恒p j)。C849残留在脊椎动物。k)域结构WT Fibrillin (FBN) 2和FBN2gydF4y2BaC849XgydF4y2Ba。l)代表图像的腹侧的观点wholemount气管与阿尔新蓝染色从P0 WT (n = 15)gydF4y2BaFbn2gydF4y2Ba——/ T2547AgydF4y2Ba双杂合的动物(n = 15)。双头箭头表示气管导管的长度。箭头指向气管软骨环。比例尺= 1毫米。蚂蚁:前;职位:后;D:背;V:腹侧;R:正确;L:左; Cart: cartilage; SM: smooth muscle; EGF: epidermal growth factor. Data are presented as mean±sdgydF4y2Ba。*:p < 0.05;* *:p < 0.01;* * *:p < 0.001未配对t检验。gydF4y2Ba
FBN2气管平滑肌细胞周围和内层gydF4y2Ba
检查FBN2的时空表达模式在发展中小鼠气管,我们进行免疫染色在胚胎期间和产后阶段使用氨基酸29 - 535引起的多克隆抗体。FBN2似乎本土化气管周围间质包括平滑肌细胞和内层最早E12.5 (gydF4y2Ba图2一个gydF4y2Ba)。有趣的是,FBN2表达水平逐渐减少内层在软骨细胞分化出现E14.5 E18.5 (gydF4y2Ba图2 bgydF4y2Ba),这样FBN2疣状几乎没有可检测气管软骨细胞在产后阶段(gydF4y2Ba图2 bgydF4y2Ba)。然后我们检查gydF4y2BaFbn2gydF4y2Ba在发展中气管mRNA的表达。在E14.5气管,gydF4y2BaFbn2gydF4y2Ba似乎是专门表达间充质细胞,包括内层(gydF4y2Ba补充图S8agydF4y2Ba)。然而,在E16.5,gydF4y2BaFbn2gydF4y2Ba信使rna水平似乎减少气管软骨细胞(gydF4y2Ba补充图S8bgydF4y2Ba),主要是发现在其他间充质细胞,包括气管平滑肌细胞(gydF4y2Ba补充图S8cgydF4y2Ba)。这些数据表明FBN2需要气管平滑肌细胞和软骨细胞在早期胚胎阶段发展。接下来,我们研究了gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba信使rna和蛋白质:突变mRNA水平相比显著降低WT (gydF4y2Ba图2 cgydF4y2Ba和gydF4y2Ba补充表1gydF4y2Ba),这表明mRNA衰变。没有FBN2免疫印迹(gydF4y2Ba图2 dgydF4y2Ba)或疣状(gydF4y2Ba图2 egydF4y2Ba)观察信号变异动物,表明截断gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba蛋白质合成或高度不稳定。这些结果表明,gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba突变会导致减少FBN2 mRNA和蛋白水平。gydF4y2Ba
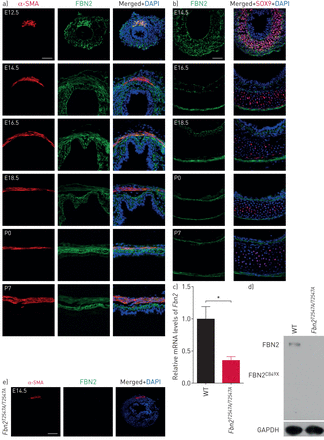
Fibrillin (FBN) 2气管平滑肌细胞周围的本土化和内层。)疣状FBN2(绿色)和α-smooth肌肉肌动蛋白(SMA)(红色),和4′,6-diamidino-2-phenylindole DAPI染色(蓝色)的横向部分野生型(WT)气管(n = 6)在整个气管区域(胚胎(E) 12.5)或平滑肌地区主要在后期。酒吧= 50μm规模。b)疣状FBN2(绿色)和SOX9(红色),和DAPI染色(蓝色)的横向部分WT气管软骨地区(n = 6)主要在几个胚胎和产后阶段。酒吧= 50μm规模。c)定量逆转录PCR分析gydF4y2BaFbn2gydF4y2Ba表达E12.5 WT (n = 4)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 4)气管。d)西方墨点法FBN2和3 -磷酸甘油醛脱氢酶(GAPDH)在产后一天(P) 7 WT (n = 4)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 4)气管。e)疣状FBN2(绿色)和α-smooth肌肉肌动蛋白(SMA)(红色),和DAPI染色(蓝色)E14.5的横截面gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管(n = 6)。酒吧= 100μm规模。数据意味着±gydF4y2BasdgydF4y2Ba。*:p < 0.05;未配对t检验。gydF4y2Ba
Fbn2gydF4y2BaT2547A / T2547AgydF4y2Ba显示缺陷小鼠气管伸长、软骨环形成和平滑肌组织gydF4y2Ba
详细检查气管的形成,我们气管导管发展进行了系统分析。gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba老鼠显示气管导管长度相比没有明显差异在E12.5 WT兄弟姐妹(gydF4y2Ba图3一gydF4y2Ba和c)。然而,从E13.5开始,我们观察到gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管比WT同行(短gydF4y2Ba图3一gydF4y2Ba和c),这种表型变得更加明显在产后阶段(gydF4y2Ba图3 bgydF4y2Ba和c)。这些结果表明,受损平滑肌分化后发生气管导管伸长,这始于E11.5 [gydF4y2Ba6gydF4y2Ba]。确定改变在气管体系结构中,我们进行了结构分析的软骨和平滑肌。软骨的形成发生由于间充质细胞的凝结chondrogenic分化[紧随其后gydF4y2Ba34gydF4y2Ba]。从E13.5开始,chondrogenic结节造成SOX9gydF4y2Ba+gydF4y2Ba间充质细胞凝结观察(gydF4y2Ba图3 dgydF4y2Ba)。然而,gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管表现出数量减少的c形环从E13.5直到成年期(gydF4y2Ba图3 dgydF4y2Ba- f)。此外,从E14.5开始,c形环之间的距离(增加gydF4y2Ba图3 dgydF4y2Ba和g)和减少环宽度(gydF4y2Ba图3 dgydF4y2Ba和h)被观察到gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管。FBN2水平下降期间逐步chondrogenic老鼠ATDC5细胞的分化gydF4y2Ba35gydF4y2Ba),类似于在气管内层(gydF4y2Ba图2 bgydF4y2Ba)。因此,我们假设,FBN2不足可能会导致损害气管软骨细胞分化。为了验证这个假设,我们检查了表达水平aggrecan (AGC1)和II型胶原蛋白(COL2A1) chondrocyte-specific标记(gydF4y2Ba34gydF4y2Ba),观察到AGC1 (gydF4y2Ba补充图S9a和bgydF4y2Ba)和COL2A1 (gydF4y2Ba补充图S9c和dgydF4y2Ba)水平显著降低gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管。相比之下,我们并没有发现明显的变化在细胞凋亡(gydF4y2Ba补充图S10a-cgydF4y2Ba)或SOX9gydF4y2Ba+gydF4y2Ba成软骨细胞增殖(gydF4y2Ba补充图S10d和egydF4y2BaWT之间),gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管,表明FBN2不足导致异常气管环通过抑制软骨细胞分化。然后我们孤立的不成熟的软骨细胞从E14.5气管和培养9天。有趣的是,gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba软骨细胞表现出降低阿尔新蓝染色在初级文化(gydF4y2Ba补充图S11a和bgydF4y2Ba),这表明gydF4y2BaFbn2gydF4y2Ba调节软骨细胞自动的方式形成。因为平滑肌形态发生改变会影响管伸长(gydF4y2Ba4gydF4y2Ba,gydF4y2Ba10gydF4y2Ba,gydF4y2Ba36gydF4y2Ba),我们分析了气管平滑肌的发展。平滑肌细胞,定位在气管背,显示WT,之间没有明显的差异gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba动物在E12.5 (gydF4y2Ba图3我gydF4y2Ba和j),这表明FBN2是可有可无的平滑肌分化和平滑肌形态发生的早期阶段。最早观察到组织平滑肌减少区域的条纹gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管E13.5 (gydF4y2Ba图3我gydF4y2Ba和j)和变得更明显的从E14.5 (gydF4y2Ba图3我gydF4y2Ba相比之下,和j)。gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管平滑肌细胞没有表现出明显的细胞凋亡的变化(gydF4y2Ba补充图S10a-cgydF4y2Ba)或扩散(gydF4y2Ba补充图S10f和ggydF4y2Ba比WT)。同样,有丝分裂纺锤体取向(gydF4y2Ba补充图S12a和bgydF4y2Ba)和细胞增殖(gydF4y2Ba补充图S12c和dgydF4y2Ba)的气管上皮细胞gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba老鼠WT。因为相比似乎不受影响gydF4y2BaFgf10gydF4y2Ba在气管平滑肌细胞表达,调节气道模式(gydF4y2Ba20.gydF4y2Ba),我们还测试了gydF4y2BaFgf10gydF4y2Ba信使rna水平。gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管表现出无显著差异gydF4y2BaFgf10gydF4y2Ba表达水平(gydF4y2Ba补充表1图S12e和补充gydF4y2Ba),这表明gydF4y2BaFbn2gydF4y2Ba信号gydF4y2Ba通过gydF4y2Ba一个gydF4y2BaFgf10gydF4y2Ba独立通路调节气管模式。总的来说,这些结果表明,软骨细胞分化和平滑肌组织对于气管导管的形成至关重要。gydF4y2Ba
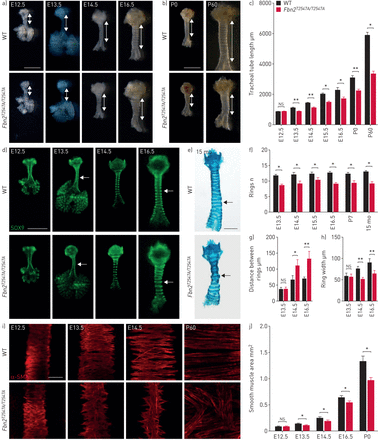
Fbn2gydF4y2BaT2547A / T2547AgydF4y2Ba显示缺陷小鼠气管伸长、软骨环形成和平滑肌组织。野生型的)代表图像腹侧的观点(WT) (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管在几个胚胎阶段。双头箭头表示气管导管的长度。比例尺= 1毫米。b)代表图像的腹侧WT (n = 6)和观点gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba在产后阶段(n = 6)气管。双头箭头表示气管导管的长度。比例尺= 2毫米。c) WT (n = 6)和量化gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管导管的长度。d)腹侧的SOX9疣状(绿色)的WT (n = 7)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 7)气管在几个胚胎阶段。箭头指向气管环。比例尺= 1毫米。e)代表图像的腹侧的观点wholemount气管与阿尔新蓝染色从15个月大(15 mo) WT (n = 7)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba老鼠(n = 7)。箭头指向气管软骨环。比例尺= 2毫米。f)量化的气管软骨环在WT (n = 7)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 7)动物。g)量化气管环之间的距离在WT (n = 7)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 7)动物。h)量化环的宽度在WT (n = 7)和gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 7)动物。我)背视图α-smooth肌肉肌动蛋白(SMA)疣状(红色)的WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管在几个胚胎和产后阶段。酒吧= 100μm规模。j)量化平滑肌在WT (n = 6)和面积gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)动物。艾凡:胚胎;P:产后一天。数据意味着±gydF4y2BasdgydF4y2Ba。*:p < 0.05;* *:p < 0.01;gydF4y2BansgydF4y2Ba:不重要的;未配对t检验。gydF4y2Ba
Fbn2gydF4y2Ba介导平滑肌细胞取向和两极分化gydF4y2Ba
纵向限制肠道平滑肌报道管伸长通过压缩间质和附加上皮(gydF4y2Ba4gydF4y2Ba]。我们需要发现FBN2平滑肌组织让我们假设,气管伸长中观察到的缺陷gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba老鼠可能是由于改变了平滑肌细胞排列。因此,我们进行了系统分析的平滑肌细胞取向。在WT动物分化气管平滑肌细胞表现出圆的形状和逐步发展成纺锤状细胞压痕使管E13.5 (gydF4y2Ba图4一gydF4y2Ba和b),我们观察到在平滑肌对齐之间没有显著差异gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba和WT气管E12.5 (gydF4y2Ba图4 cgydF4y2Ba和d)。然而,在E13.5,gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba平滑肌细胞显示随机排列(gydF4y2Ba图4 cgydF4y2Ba和d),这在成年动物观察表型(gydF4y2Ba图4 cgydF4y2Ba和d)。这种平滑肌表型暂时与气管伸长的缺陷gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba老鼠(gydF4y2Ba图3 cgydF4y2Ba和d)。细胞已被证明的正确取向取决于细胞极性的建立(gydF4y2Ba37gydF4y2Ba]。为了更好地理解平滑肌细胞的两极分化,我们检查了本地化的高尔基体相对于细胞核,使用cis-Golgi矩阵GM130标志,一种广泛使用的方法来确定细胞极性不同细胞类型(gydF4y2Ba38gydF4y2Ba,gydF4y2Ba39gydF4y2Ba]。在WT平滑肌细胞,GM130-labeled高尔基本土化优先由原子核的长边(gydF4y2Ba图4 egydF4y2Ba和f)。相比之下,gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba平滑肌细胞的高尔基体表现出随机排列(gydF4y2Ba图4 egydF4y2Ba和f)。气管平滑肌细胞建立极性E12.5之前,及其破坏原因改变平滑肌由E12.5架构(gydF4y2Ba36gydF4y2Ba]。gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba平滑肌展出WT E12.5(相比无显著差异gydF4y2Ba图4 cgydF4y2Ba这些结果表明,d)。gydF4y2BaFbn2gydF4y2Ba是可有可无的起始的平滑肌细胞极性,但成为必要的维护。组织收缩性报道开车tubulogenesis [gydF4y2Ba3gydF4y2Ba,gydF4y2Ba40gydF4y2Ba]。基于发现的gydF4y2BaFbn2gydF4y2Ba平滑肌细胞需要校准和极性,我们提出,平滑肌细胞取向的中断可能会导致损害圆周气管收缩导致管伸长缺陷gydF4y2BaFbn2gydF4y2Ba突变的老鼠。有趣的是,gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管表现出持续时间增加(gydF4y2Ba图4 ggydF4y2Ba和h和gydF4y2Ba补充电影1gydF4y2Ba)和减少振幅(gydF4y2Ba图4 ggydF4y2Ba和我和gydF4y2Ba补充电影1gydF4y2Ba在E13.5 WT相比)的自发收缩。此外,gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管表现出显著降低收缩力量相比WT (gydF4y2Ba图4 jgydF4y2Ba)。这些发现表明,FBN2是至关重要的维持平滑肌细胞极性直接组织架构和收缩性,最终推动气管导管伸长。gydF4y2Ba
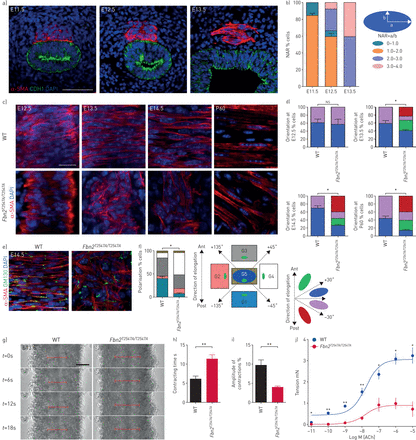
Fbn2gydF4y2Ba调节平滑肌细胞定向分化。)疣状α-smooth肌肉肌动蛋白(SMA)(红色)和背景(绿色),和4′,6-diamidino-2-phenylindole DAPI染色(蓝色)的横向部分野生型(WT)气管(n = 6)在几个胚胎阶段。酒吧= 50μm规模。b)量化气管平滑肌细胞的核宽高比(NAR)。c)的背视图α-SMA疣状(红色)和DAPI染色(蓝色)的WT (n = 7)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 7)气管在几个胚胎(E)和产后(P)阶段。酒吧= 20μm规模。d) WT (n = 7)和量化gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 7)气管平滑肌细胞取向。e)的背视图α-SMA(红色)和GM130(绿色)免疫染色和DAPI染色(蓝色)E14.5 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管。酒吧= 20μm规模。f)的量化E14.5 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)高尔基体位置相对于原子核(蓝色)。g)背视图的延时图像自发收缩E13.5 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管。线表示气管平滑肌宽度。酒吧= 50μm规模。h)收缩时间的量化E13.5 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管。我)的收缩幅度量化E13.5 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管。j)气管压力P1 WT (n = 5)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 5)气管。蚂蚁:前;职位:后。数据意味着±gydF4y2BasdgydF4y2Ba。*:p < 0.05;* *:p < 0.01;gydF4y2BansgydF4y2Ba:不重要的;未配对t检验。gydF4y2Ba
食道和气管分开一个共同的前肠管在E9.5 [gydF4y2Ba41gydF4y2Ba]。因此,我们试图确定gydF4y2BaFbn2gydF4y2Ba也需要食管管形成。在食管平滑肌FBN2高度表达(gydF4y2Ba补充图S13agydF4y2Ba),但我们发现管长度之间的差异不显著gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba和WT老鼠(gydF4y2Ba补充图S13b和cgydF4y2Ba)。接下来,我们检查食管平滑肌形态。平滑肌细胞局部在食管的外层墙gydF4y2Ba42gydF4y2Ba)(gydF4y2Ba补充图S13a和dgydF4y2Ba),主轴gydF4y2Ba- - - - - -gydF4y2Ba的平滑肌细胞被E14.5压痕对齐(gydF4y2Ba补充图S13e和fgydF4y2Ba)。与气管,我们观察到食管平滑肌组织(没有明显的差异gydF4y2Ba补充图S13dgydF4y2Ba)或平滑肌细胞取向(gydF4y2Ba补充图S13e和fgydF4y2Ba)之间的gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba和WT老鼠。这些数据表明,gydF4y2BaFbn2gydF4y2Ba是可有可无的平滑肌发展在食管管形成。gydF4y2Ba
Fbn2gydF4y2Ba调节弹性纤维形成和纤连蛋白积累gydF4y2Ba
探讨分子机制gydF4y2BaFbn2gydF4y2Ba介导的气管导管的形成,我们检查了ECM蛋白质形成和水平。Fibrillin微纤维被提供一个支架的弹性蛋白原沉积,最终成熟弹性蛋白和弹性纤维(gydF4y2Ba43gydF4y2Ba- - - - - -gydF4y2Ba47gydF4y2Ba]。我们提出,gydF4y2BaFbn2gydF4y2Ba缺陷可能导致缺陷的弹性蛋白结构。在E14.5,弹性纤维在WT平滑肌细胞与核的长边平行和近似垂直于管伸长的方向(gydF4y2Ba图5一个gydF4y2Ba和b)。相比之下,弹性纤维取向gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管出现随机的,面向大多数人在一个角度斜管的方向伸长(gydF4y2Ba图5一个gydF4y2Ba和b)。有趣的是,短期弹性纤维也被观察到gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管与WT (gydF4y2Ba图5一个gydF4y2Ba然而,和c)。gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管表现出弹性蛋白的蛋白质含量没有明显变化(gydF4y2Ba图5 dgydF4y2Ba),这表明FBN2弹性纤维的形成需要但不是弹力素蛋白表达。接下来,我们研究了弹性纤维食管平滑肌细胞。之间没有显著差异在弹性纤维的形成gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba和WT oesophagi (gydF4y2Ba补充图S13g-igydF4y2Ba)。gydF4y2BaFbn1gydF4y2Ba据报道调节弹性纤维的形成(gydF4y2Ba48gydF4y2Ba- - - - - -gydF4y2Ba50gydF4y2Ba]。然而,gydF4y2BaFbn2gydF4y2Ba零突变体展览WT-like血管平滑肌细胞和弹性纤维的形成gydF4y2Ba44gydF4y2Ba]。有趣的是,gydF4y2BaFbn1gydF4y2Ba高度表达oesophagi (gydF4y2Ba补充图S13jgydF4y2Ba)和FBN1本土化血管壁(gydF4y2Ba51gydF4y2Ba]。这些结果表明,gydF4y2BaFbn1gydF4y2Ba和gydF4y2BaFbn2gydF4y2Ba可能在食管和血管平滑肌形态发生冗余功能。gydF4y2Ba
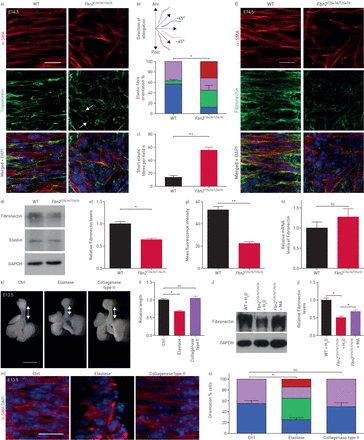
Fbn2gydF4y2Ba调节弹性纤维形成和纤连蛋白积累。)背视图α-smooth肌肉肌动蛋白(SMA)(红色)和弹性蛋白原(绿色)免疫染色和4′,6-diamidino-2-phenylindole DAPI染色(蓝色)的胚胎天14.5 (E)野生型(WT) (n = 8)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 8)气管。箭头指向短弹性纤维。酒吧= 20μm规模。量化的b)弹性纤维取向和c)短弹性纤维在气管平滑肌细胞WT (n = 8)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 8)气管。d)西方墨点法纤连蛋白、弹性蛋白和3 -磷酸甘油醛脱氢酶(GAPDH)在产后一天(P) 7 WT (n = 7)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 7)气管。e)量化纤连蛋白的相对水平P7 WT (n = 7)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 7)气管。f)的背视图α-SMA(红色)和纤连蛋白(绿色)免疫染色和DAPI染色(蓝色)E14.5 WT (n = 8)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 8)气管。酒吧= 20μm规模。g)量化的平均荧光强度纤连蛋白免疫染色在E14.5 WT (n = 8)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 8)气管。h)定量逆转录PCR分析gydF4y2Ba纤连蛋白gydF4y2Ba表达E14.5 WT (n = 5)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(gydF4y2BangydF4y2Ba= 5)气管。我在P7)为纤连蛋白免疫印迹和GAPDH WT (n = 5)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 5) ddHgydF4y2Ba2gydF4y2BaO-treated气管,在gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba甲胺hydrochloride-treated气管(n = 5)。j)量化纤连蛋白的相对水平P7 WT (n = 5)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 5) ddHgydF4y2Ba2gydF4y2BaO-treated气管,在gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba甲胺盐酸盐(MA)治疗气管(n = 5)。腹侧的k)代表图像的观点E13.5气管治疗后28 h DMEM / F-12介质(n = 6), 2μ·毫升gydF4y2Ba−1gydF4y2Ba弹性蛋白酶(n = 6)或2μ·毫升gydF4y2Ba−1gydF4y2Ba胶原酶II型(n = 6)。比例尺= 1毫米。l)量化相对气管导管长度治疗后28 h DMEM / F-12介质(n = 6), 2μ·毫升gydF4y2Ba−1gydF4y2Ba弹性蛋白酶(n = 6)或2μ·毫升gydF4y2Ba−1gydF4y2Ba胶原酶II型(n = 6)。米)背的观点α-SMA疣状(红色)和DAPI染色(蓝色)的E13.5气管治疗后28 h DMEM / F-12介质(n = 6), 2μ·毫升gydF4y2Ba−1gydF4y2Ba弹性蛋白酶(n = 6)或2μ·毫升gydF4y2Ba−1gydF4y2Ba胶原酶II型(n = 6)。酒吧= 20μm规模。n)量化后的气管平滑肌细胞取向治疗28 h DMEM / F-12介质(n = 6)或2μ·毫升gydF4y2Ba−1gydF4y2Ba弹性蛋白酶(n = 6)或2μ·毫升gydF4y2Ba−1gydF4y2Ba胶原酶II型(n = 6)。蚂蚁:前;职位:后。数据意味着±gydF4y2BasdgydF4y2Ba。*:p < 0.05;* *:p < 0.01;gydF4y2BansgydF4y2Ba:不重要的;未配对t检验。gydF4y2Ba
另一个关键ECM蛋白、纤连蛋白(FN),已经被证明可以绑定FBN2 [gydF4y2Ba52gydF4y2Ba,gydF4y2Ba53gydF4y2Ba和促进细胞伸长和对齐gydF4y2Ba54gydF4y2Ba,gydF4y2Ba55gydF4y2Ba]。有趣的是,gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管表现出显著降低FN蛋白质含量与WT (gydF4y2Ba图5 dgydF4y2Ba- g),这表明FN取决于FBN2积累。然而,我们观察到在Fn mRNA水平没有显著差异gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管WT(相比gydF4y2Ba图5 hgydF4y2Ba)。甲胺盐酸盐,溶酶体抑制剂,抑制FN退化(gydF4y2Ba56gydF4y2Ba]。治疗后与甲胺盐酸盐gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管展出部分恢复FN蛋白质水平(gydF4y2Ba图5我gydF4y2Ba和j),这表明FBN2保护FN在溶酶体降解气管导管的形成。测试是否FBN2在弹性纤维的形成过程中发挥作用和FN积累在成年老鼠,我们检查了弹性纤维和FN在成人气管。我们发现gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2BaP70气管表现出随机弹性纤维取向(gydF4y2Ba补充图S14a和bgydF4y2Ba)、短弹性纤维(gydF4y2Ba补充图S14a和cgydF4y2Ba)、FN蛋白质含量降低(gydF4y2Ba补充图S14d和egydF4y2Ba),但弹性蛋白的蛋白质含量没有明显差异(gydF4y2Ba补充图S14f和ggydF4y2Ba)与WT相比,可能表明需要FBN2 ECM内稳态在成熟的老鼠。然而,检查成人的函数gydF4y2BaFbn2gydF4y2Ba,这将是必要的生成和使用液氧gydF4y2BaFbn2gydF4y2Ba等位基因。弹性蛋白缺乏报道改变方向的血管壁平滑肌细胞(gydF4y2Ba57gydF4y2Ba,gydF4y2Ba58gydF4y2Ba]。我们因此提出弹性纤维的破坏可能拟表型gydF4y2BaFbn2gydF4y2Badeficiency-induced气管伸长和平滑肌细胞取向缺陷。有趣的是,在治疗后28 h 2μ·毫升gydF4y2Ba−1gydF4y2Ba弹性蛋白酶,E13.5气管表现出显著减少管长度(gydF4y2Ba图5 kgydF4y2Ba和l)和平滑肌细胞改变方向(gydF4y2Ba图5米gydF4y2Ba和n)与控制相比,表型没有观察到治疗后与胶原酶II型(gydF4y2Ba图5 kgydF4y2Ba- n)。总之,这些结果表明,FBN2-mediated ECM形成和积累是必不可少的气管平滑肌细胞取向和器官伸长。gydF4y2Ba
的差别p38-dependent对这些gydF4y2BaMmp的gydF4y2Ba表达式的中介gydF4y2BaFbn2gydF4y2Ba函数在气管开发gydF4y2Ba
我们试图进一步了解FBN2不足导致受损ECM的形成和积累。观察短期弹性纤维和FN退化促使我们分析的水平MMP2和MMP9,弹性蛋白、纤粘连蛋白的关键gydF4y2Ba- - - - - -gydF4y2Ba裂开的酶(gydF4y2Ba59gydF4y2Ba- - - - - -gydF4y2Ba62年gydF4y2Ba]。gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管表现出显著增加gydF4y2BaMmp2gydF4y2Ba和gydF4y2BaMmp9gydF4y2Ba信使rna水平(gydF4y2Ba图6gydF4y2Ba和gydF4y2Ba补充表S1gydF4y2Ba)以及蛋白质含量增加(gydF4y2Ba图6 bgydF4y2Ba和c)和活动(gydF4y2Ba图6 dgydF4y2Ba相比之下,和e)。gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Baoesophagi表现没有明显的差异gydF4y2BaMmp2gydF4y2Ba信使rna水平(gydF4y2Ba补充图S15a S1和补充表gydF4y2Ba),而gydF4y2BaMmp9gydF4y2Ba信使rna在E14.5察觉WT oesophagi (gydF4y2Ba补充表S1gydF4y2Ba)。由GM6001 (MMP的抑制gydF4y2Ba63年gydF4y2Ba)已经被证明可以减少弹性蛋白劈理(gydF4y2Ba64年gydF4y2Ba)和FN退化(gydF4y2Ba65年gydF4y2Ba]。我们因此检测MMP抑制能否拯救gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管表型。我们管理GM6001(50毫克公斤gydF4y2Ba−1gydF4y2Ba连续三天从E12.5 E14.5)腹腔内。有趣的是,GM6001注射后,gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管展出部分获救管伸长(gydF4y2Ba图6 fgydF4y2Ba和g)和平滑肌取向(gydF4y2Ba图6 hgydF4y2Ba和我)表型,以及部分恢复弹性纤维的形成(gydF4y2Ba图6 hgydF4y2Ba,j和k)和FN积累(gydF4y2Ba图6 hgydF4y2Ba和l)作为盲评估分析。我们旨在发现机制gydF4y2BaFbn2gydF4y2Ba调节gydF4y2BaMmp2gydF4y2Ba和gydF4y2BaMmp9gydF4y2Ba表达式,从而检查的表达gydF4y2BaAtf2gydF4y2Ba和gydF4y2BaHspb1gydF4y2Ba的,潜在的监管机构gydF4y2BaMmp2gydF4y2Ba表达式[gydF4y2Ba66年gydF4y2Ba),以及gydF4y2BaNfkb1gydF4y2Ba,gydF4y2BaNfkb2gydF4y2Ba,gydF4y2BaRelagydF4y2Ba,gydF4y2Ba小君gydF4y2Ba和gydF4y2Ba”丛书gydF4y2Ba的,潜在的监管机构gydF4y2BaMmp9gydF4y2Ba表达式[gydF4y2Ba66年gydF4y2Ba]。gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管WT-like展出的这些基因表达水平(gydF4y2Ba补充图S15b c表S1和补充gydF4y2Ba),这表明gydF4y2BaFbn2gydF4y2Ba调节gydF4y2BaMmp2和Mmp9gydF4y2Ba信使rna水平通过其他途径。gydF4y2Ba
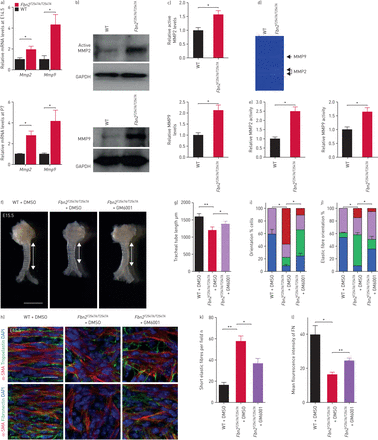
基质金属蛋白酶(MMPs)函数的介质Fibrillin (FBN) 2在气管发展功能。定量逆转录PCR分析gydF4y2BaMmp2gydF4y2Ba和gydF4y2BaMmp9gydF4y2Ba表达在胚胎(E) 14.5和产后一天(P) 7野生型(WT) (n = 5和5)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 5和5)气管。b)西方墨点法MMP2, MMP9和3 -磷酸甘油醛脱氢酶(GAPDH) P7 WT (n = 7)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 7)气管。c)的相对水平量化MMP2和MMP9 P7 WT (n = 7)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 7)气管。d) Zymography分析MMP2和MMP9活动P7 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管提取物。e)量化MMP2和MMP9活动P7 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管提取物。f)代表图像的腹侧的观点E15.5 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管二甲基亚砜(DMSO)注射后,和gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管(n = 6)后GM6001注入。双头箭头表示气管导管的长度。比例尺= 1毫米。g)的量化E15.5 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管导管长度DMSO溶液注射后,和gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管(n = 6)后GM6001注入。h)背视图α-smooth肌肉肌动蛋白(SMA)(红色)和弹性蛋白原(绿色),或纤连蛋白(绿色)免疫染色和4′,6-diamidino-2-phenylindole DAPI染色(蓝色)的E15.5 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管DMSO溶液注射后,和gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管(n = 6)后GM6001注入。酒吧= 20μm规模。量化的我)气管平滑肌细胞取向,j)弹性纤维取向、k)短期弹性纤维和l)平均荧光强度纤连蛋白免疫染色在E15.5 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管DMSO溶液注射后,在gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管(n = 6)后GM6001注入。数据意味着±gydF4y2BasdgydF4y2Ba。*:p < 0.05;* *:p < 0.01;未配对t检验。gydF4y2Ba
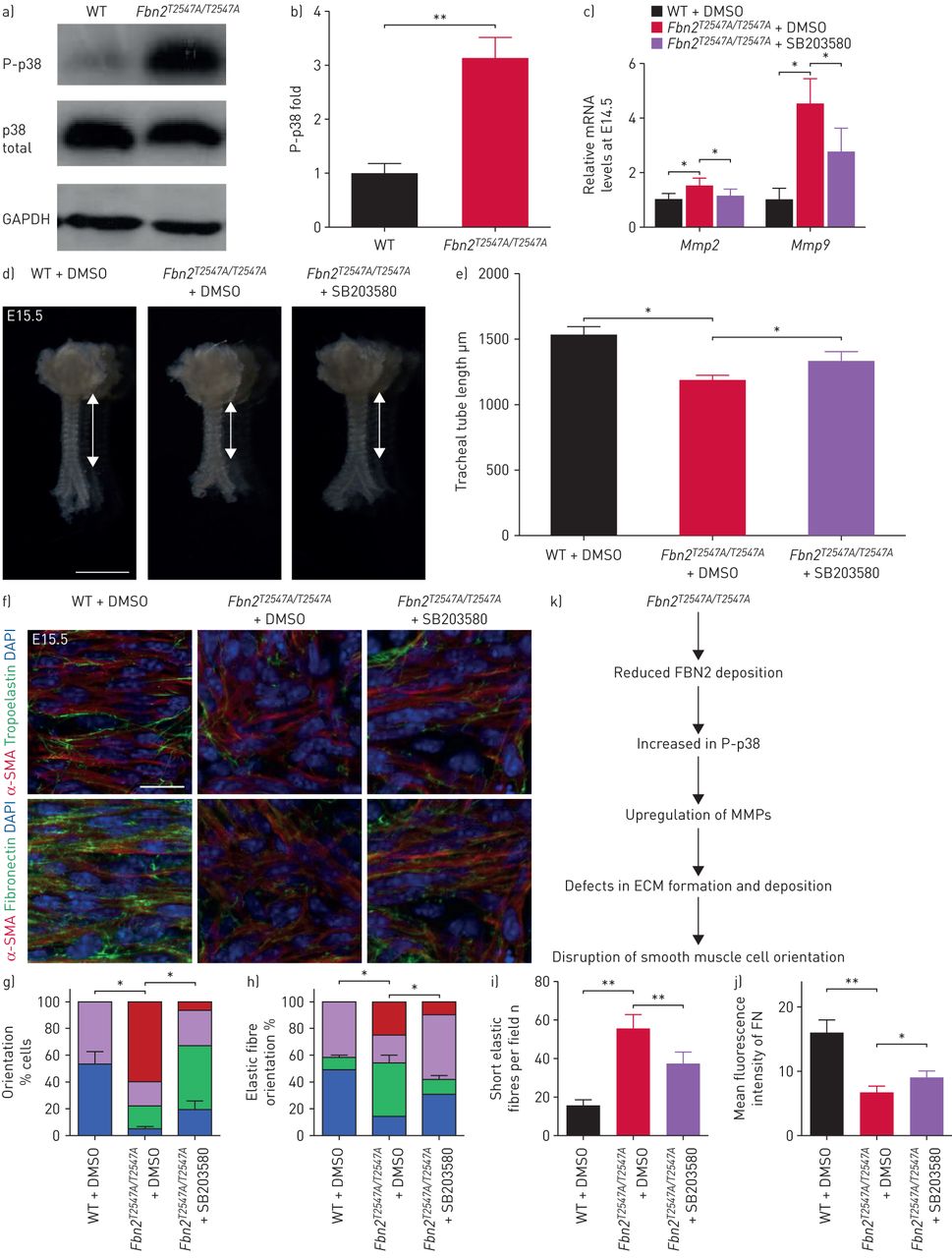
Mmp的gydF4y2Ba年代已确定的下游目标p38 MAPK信号(gydF4y2Ba67年gydF4y2Ba),会使有效抑制p38 MAPK信号通路gydF4y2BaMmp2gydF4y2Ba和gydF4y2BaMmp9gydF4y2Ba表达式[gydF4y2Ba67年gydF4y2Ba,gydF4y2Ba68年gydF4y2Ba]。因此,我们假设,Fbn2不足可能会导致增加p38 MAPK活性。我们观察到的磷酸化水平p38大大升高gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管WT(相比gydF4y2Ba图7gydF4y2Ba和b和gydF4y2Ba补充图S16a和bgydF4y2Ba),没有观察到的表型gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Baoesophagi (gydF4y2Ba补充图S16c和dgydF4y2Ba)。此外,与SB203580治疗后,小分子,减少p38磷酸化(gydF4y2Ba69年gydF4y2Ba),gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管展出部分恢复gydF4y2BaMmp2gydF4y2Ba和gydF4y2BaMmp9gydF4y2Ba信使rna水平(gydF4y2Ba图7 cgydF4y2Ba和gydF4y2Ba补充表S1gydF4y2Ba)。然后我们管理SB203580(1μM·公斤gydF4y2Ba−1gydF4y2Ba连续三天从E12.5 E14.5)腹腔内。值得注意的是,SB203580注射后,gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管展出部分获救管伸长(gydF4y2Ba图7 dgydF4y2Ba和e)和平滑肌取向(gydF4y2Ba图7 fgydF4y2Ba和g)表型,恢复弹性纤维的形成(gydF4y2Ba图7 fgydF4y2Ba,我和h)以及FN积累(gydF4y2Ba图7 fgydF4y2Ba和j)作为盲评估分析。这些结果表明,p38 phosphorylation-mediated的增加gydF4y2BaMmp的gydF4y2Ba水平gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管部分占面向平滑肌细胞表型gydF4y2Ba通过gydF4y2Ba其行动ECM内稳态(gydF4y2Ba图7 kgydF4y2Ba)。gydF4y2Ba
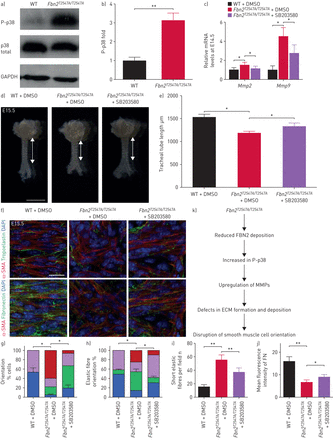
增加p38信号与增加基质金属蛋白酶(gydF4y2BaMmp的gydF4y2Ba)表达式,弹性纤维的形成和纤连蛋白降解受损。phospho-p38)免疫印迹,总p38和3 -磷酸甘油醛脱氢酶(GAPDH)在产后一天(P) 7野生型(WT) (n = 7)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 7)气管。b)量化phospho-p38相对水平的P7 WT (n = 7)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 7)气管。c)定量逆转录PCR分析gydF4y2BaMmp2gydF4y2Ba和gydF4y2BaMmp9gydF4y2Ba表达在胚胎天(E) 14.5 WT (n = 5)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 5) DMSO-treated气管,在gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2BaSB203580-treated气管(gydF4y2BangydF4y2Ba= 5)。d)代表图像的腹侧的观点E15.5 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管二甲基亚砜(DMSO)注射后,和gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管(n = 6)后SB203580注入。双头箭头表示气管导管的长度。比例尺= 1毫米。e)量化的气管导管长度E15.5 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6) DMSO溶液注射后,在gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)后SB203580注入。f)背视图α-smooth肌肉肌动蛋白(SMA)(红色)和弹性蛋白原(绿色),或纤连蛋白(绿色)免疫染色和4′,6-diamidino-2-phenylindole DAPI染色(蓝色)的E15.5 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管DMSO溶液注射后,和gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管(n = 6)后SB203580注入。酒吧= 20μm规模。量化的g)气管平滑肌细胞定位、h)弹性纤维取向,我短的弹性纤维和j)平均荧光强度纤连蛋白免疫染色在E15.5 WT (n = 6)gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba(n = 6)气管DMSO溶液注射后,在gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管(n = 6)后SB203580注入。k)模型的影响gydF4y2BaFbn2gydF4y2BaT2547AgydF4y2Ba突变的细胞外基质(ECM)组织和稳定,和气管平滑肌细胞取向。数据意味着±gydF4y2BasdgydF4y2Ba。*:p < 0.05;* *:p < 0.01;未配对t检验。gydF4y2Ba
FBN2和纤连蛋白减少人类气管软化患者的气管gydF4y2Ba
自gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba老鼠表现出气管软骨缺陷,表型类似观察气管软化患者(gydF4y2Ba图8gydF4y2Ba),我们检查FBN2表达健康对照组的气管和气管软化患者(gydF4y2Ba补充表2gydF4y2Ba)。FBN2局部气管软骨的外围区域(gydF4y2Ba图8 bgydF4y2Ba),没有围绕SOX9gydF4y2Ba+gydF4y2Ba间充质细胞(gydF4y2Ba图8 bgydF4y2Ba人类气管)在产后,表达模式在老鼠身上所观察到的类似产后阶段(gydF4y2Ba图2 bgydF4y2Ba)。我们发现FBN2水平显著降低气管软骨地区从气管软化患者与健康对照组(gydF4y2Ba图8 bgydF4y2Ba和c)。我们还研究了纤连蛋白表达。纤连蛋白局部气管软骨的外围区域(gydF4y2Ba图8 dgydF4y2Ba),没有围绕SOX9gydF4y2Ba+gydF4y2Ba间充质细胞(gydF4y2Ba图8 dgydF4y2Ba人类气管)在产后,表达模式在老鼠身上所观察到的类似产后阶段(gydF4y2Ba补充图肌力gydF4y2Ba)。有趣的是,纤连蛋白水平也似乎减少气管软骨地区从气管软化患者与健康对照组(gydF4y2Ba图8 dgydF4y2Ba和e)。这些结果在小鼠人类的结果是一致的,这表明减少ECM蛋白质,包括FBN2和纤连蛋白,是相关的,和可能导致气管软化的症状。gydF4y2Ba

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
FBN2和纤连蛋白水平降低人类气管软化患者的气管。)代表图像的横向部分气管与苏木精和伊红染色从健康对照组(n = 5)和气管软化患者(n = 7)。箭头指向气管软骨。比例尺= 1毫米。b)疣状SOX9(红色)和FBN2(绿色),和4′,6-diamidino-2-phenylindole DAPI染色(蓝色)的横向部分腹部分气管从健康对照组(n = 4)和气管软化患者(n = 6)。酒吧= 100μm规模。c)相对平均荧光强度的量化FBN2疣状腹部分气管从健康对照组(n = 4)和气管软化患者(n = 6)。d)疣状SOX9(红色)和纤连蛋白(绿色),和DAPI染色(蓝色)的横向部分腹部分气管从健康对照组(n = 5)和气管软化患者(n = 5)。酒吧= 100μm规模。e)相对平均荧光强度的量化纤连蛋白免疫染色腹部分气管从健康对照组(n = 5)和气管软化患者(n = 5)。 Data are presented as mean±sdgydF4y2Ba。*:p < 0.05;未配对t检验。gydF4y2Ba
讨论gydF4y2Ba
理解气管tubulogenesis肺发展领域的基本目标和疾病,主气管软化和先天性气管狭窄等情况出现由于气管导管形成缺陷。我们的工作表明关键角色在气管导管平滑肌细胞和软骨细胞的形成。分化的平滑肌细胞伸长方向垂直于轴的气管伸长和产生周向收缩的力量促进管伸长。在gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管,面向平滑肌细胞异常的限制管伸长通过压缩间质和附加的上皮细胞,如肠道平滑肌限制的伸长(gydF4y2Ba4gydF4y2Ba]。软骨细胞也被报道对气管导管扩张是必要的(gydF4y2Ba36gydF4y2Ba,gydF4y2Ba70年gydF4y2Ba]。值得注意的是,gydF4y2BaSox9gydF4y2Ba基因敲除小鼠表现出缺陷软骨细胞分化,气管软骨形成和气管导管扩张(gydF4y2Ba36gydF4y2Ba,gydF4y2Ba70年gydF4y2Ba]。因此可能gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管软骨细胞分化的缺陷导致受损的软骨形成,最终导致缩小气管导管。有趣的是,动物模型的气管狭窄等gydF4y2Ba雌性生殖道gydF4y2Ba- / -gydF4y2Ba和gydF4y2BaCav3.2gydF4y2Ba- / -gydF4y2Ba老鼠都显示软骨环和管腔形成缺陷(gydF4y2Ba71年gydF4y2Ba- - - - - -gydF4y2Ba73年gydF4y2Ba],表型所观察到的类似gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba老鼠。gydF4y2BaCav3.2gydF4y2Ba调节气管软骨形成,促进软骨细胞分化[gydF4y2Ba73年gydF4y2Ba]。因此,缺陷在软骨细胞分化是一种常见的先天性气管狭窄的原因。gydF4y2Ba
这里我们揭示小说功能的FBN2弹性纤维形成和纤连蛋白积累。gydF4y2BaFbn2gydF4y2Ba弹性纤维形成的突变体表现没有显著差异在食管或血管平滑肌细胞(gydF4y2Ba44gydF4y2Ba]。然而,gydF4y2BaFbn1gydF4y2Ba- / -gydF4y2Ba老鼠表现出弹性纤维组织缺陷在主动脉组织中(gydF4y2Ba44gydF4y2Ba]。gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Baoesophagi展览WT-like p38磷酸化gydF4y2BaMmp2gydF4y2Ba信使rna表达水平。gydF4y2BaFbn1gydF4y2Ba突变体表现出增加p38磷酸化(gydF4y2Ba74年gydF4y2Ba)以及增加MMP2和MMP9在胸主动脉(gydF4y2Ba75年gydF4y2Ba]。有趣的是,这两个gydF4y2BaFbn1gydF4y2Ba- /gydF4y2Ba+gydF4y2Ba;gydF4y2BaFbn2gydF4y2Ba- / -gydF4y2Ba和gydF4y2BaFbn1gydF4y2Ba- / -gydF4y2Ba;和gydF4y2BaFbn2gydF4y2Ba- / -gydF4y2Ba动物显示一个更严重的血管表型gydF4y2BaFbn1gydF4y2Ba- / -gydF4y2Ba老鼠(gydF4y2Ba44gydF4y2Ba),这表明gydF4y2BaFbn1gydF4y2Ba和gydF4y2BaFbn2gydF4y2Ba执行部分重叠的功能在主动脉的发展。因此可能缺乏ECM的缺陷gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Baoesophagi是由于功能冗余gydF4y2BaFbn1gydF4y2Ba。FBN1促进弹性蛋白原凝聚(gydF4y2Ba48gydF4y2Ba,防止弹性蛋白破坏和退化可能通过减少MMP2和MMP9活动(gydF4y2Ba49gydF4y2Ba,gydF4y2Ba50gydF4y2Ba]。然而,gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管似乎表现出弹性蛋白的蛋白质水平没有明显减少,表明FBN2弹性纤维的形成需要但不是蛋白表达在气管导管的形成。colocalise Fibrillin微纤维已报告,与纤连蛋白(gydF4y2Ba52gydF4y2Ba,gydF4y2Ba53gydF4y2Ba),他们的沉积取决于纤连蛋白组装(gydF4y2Ba76年gydF4y2Ba]。有趣的是,我们的研究表明,Fibrillin微纤维纤连蛋白也需要积累。gydF4y2Ba
我们的研究提出了一种新颖的信号轴管理ECM内稳态,通过Fibrillin微纤维,p38增殖活化蛋白激酶(MAPK)和基质金属蛋白酶。gydF4y2BaFbn2gydF4y2Ba不足导致异常激活p38 MAPK信号,进而引发一个upregulation MMP活动扰乱了弹性纤维的形成,降低纤连蛋白。然而,确切的机制FBN2调节p38活动和p38调节gydF4y2BaMmp的gydF4y2Ba表达还有待确定。转化生长因子(TGF) -β异常激活gydF4y2BaFbn2gydF4y2Ba零成骨细胞(gydF4y2Ba77年gydF4y2Ba]。有趣的是,TGF-β抑制报道减少p38磷酸化在平滑肌细胞(gydF4y2Ba78年gydF4y2Ba]。可能没有激活TGF-βFBN2函数信号,从而增加p38磷酸化。已报告的几个目标p38 MAPK调节gydF4y2BaMmp2gydF4y2Ba和gydF4y2BaMmp9gydF4y2Ba表达在转录水平(gydF4y2Ba66年gydF4y2Ba),从而增加p38磷酸化可能导致特定转录因子的激活(s)的目标基因包括gydF4y2BaMmp2gydF4y2Ba和gydF4y2BaMmp9gydF4y2Ba。gydF4y2Ba
Fibrillin微纤维被发现在人类软骨的矩阵(gydF4y2Ba79年gydF4y2Ba),而gydF4y2BaFBN2gydF4y2Ba突变导致病人[皱巴巴的耳朵gydF4y2Ba80年gydF4y2Ba,gydF4y2Ba81年gydF4y2Ba),这可能是由于耳软骨缺陷。需要我们的工作提供了证据表明Fibrillin微纤维软骨的形成可能通过促进软骨细胞分化。值得注意的是,FBN2水平似乎明显减少气管软化患者的气管软骨,这表明Fibrillin微纤维软骨的形成也在人类至关重要。这将是有趣的调查潜在的分子机制。gydF4y2Ba
突变gydF4y2BaFBN2gydF4y2Ba已经与先天性国民间细长指(gydF4y2Ba80年gydF4y2Ba- - - - - -gydF4y2Ba83年gydF4y2Ba和血管疾病在人类gydF4y2Ba84年gydF4y2Ba在斑马鱼)以及心内膜形态发生缺陷gydF4y2Ba85年gydF4y2Ba]。我们的gydF4y2BaFbn2gydF4y2Ba点突变老鼠展品气管狭窄可能是软骨分化造成的缺陷,提供一种新的模式来研究这种疾病的病因学,并制定治疗方法。gydF4y2Ba
补充材料gydF4y2Ba
补充材料gydF4y2Ba
请注意:gydF4y2Ba补充材料并不是由编辑部,编辑和上传已由作者提供。gydF4y2Ba
补充数据和表gydF4y2Baerj - 00840 - 2018 - _supplementgydF4y2Ba
补充的电影。相比WT气管(n = 6),gydF4y2BaFbn2gydF4y2BaT2547A / T2547AgydF4y2Ba气管(n = 6)展览时间收缩时间和减少在E13.5自发收缩幅度。gydF4y2Baerj - 00840 - 2018 - _moviegydF4y2Ba
确认gydF4y2Ba
我们感谢弗朗西斯科·拉米雷斯(伊坎在西奈山医学院,纽约,纽约,美国)提供gydF4y2BaFbn2gydF4y2Ba零老鼠,Radhan阿杨和Yu宣卡罗尔成像援助,鲁本Marin-Juez,保罗·潘,Konstantinos Gkatzis, Srinath拉姆库玛儿,Khrievono Kikhi,杰森Kuan汉Lai沙龙Meaney-Gardian和娜娜福田(马克斯·普朗克心肺研究所坏Nauheim,德国)讨论或援助。gydF4y2Ba
脚注gydF4y2Ba
网上发表07年3月,2019;2019年3月15日,转载来纠正一个错误在生殖免疫印迹图像在图6 b。gydF4y2Ba
可以从本文的补充材料gydF4y2Bawww.qdcxjkg.comgydF4y2Ba
作者的贡献:w .阴和D.Y.R. Stainier构思的项目,设计实验和分析数据;w .阴,谷岩。金,f .谷纳温,r . Li b . Grohmann c·布特尼d .罪人和美国Offermanns导致实验和数据分析;s .小王进行免疫印迹和气管生理学实验和数据分析;g . Sengle导致FBN2抗体生成;w .阴和D.Y.R. Stainier写的手稿。所有作者评论的手稿。gydF4y2Ba
利益冲突:w .阴没有披露。gydF4y2Ba
利益冲突:谷岩。金没有披露。gydF4y2Ba
利益冲突:美国王没有披露。gydF4y2Ba
利益冲突:f .谷纳温没有披露。gydF4y2Ba
利益冲突:r·李没有披露。gydF4y2Ba
利益冲突:c·布特尼没有披露。gydF4y2Ba
利益冲突:b Grohmann没有披露。gydF4y2Ba
利益冲突:g . Sengle没有披露。gydF4y2Ba
利益冲突:d .罪人没有披露。gydF4y2Ba
利益冲突:美国Offermanns没有披露。gydF4y2Ba
利益冲突:D.Y.R. Stainier没有披露。gydF4y2Ba
支持声明:本研究经费是由德意志Forschungsgemeinschaft (DFG) g . Sengle (SFB829 / B11, B12)和马克斯·普朗克社会Offermanns和D.Y.R. Stainier。gydF4y2Ba
- 收到了gydF4y2Ba2018年5月8日。gydF4y2Ba
- 接受gydF4y2Ba2018年11月28日。gydF4y2Ba
- 版权©2019人队gydF4y2Ba
引用gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba










