





摘要gydF4y2Ba
本研究首次分析了暴露于异弗尔酮二异氰酸酯(IPDI)后支气管上皮分泌的可溶性因子,这些因子可增加原代正常人支气管平滑肌细胞(BSMCs)的迁移和增殖。gydF4y2Ba
我们用IPDI处理不朽的、非致瘤的人支气管上皮细胞(BEAS-2B细胞系)和原代正常人支气管上皮细胞(HBEC),然后收集条件培养基(IPDI-BEAS-2B- cm和IPDI-HBEC- cm),并将其添加到BSMCs中。gydF4y2Ba
暴露于IPDI的BEAS-2B细胞和HBECs增加了白细胞介素(IL)-8的产生。采用IPDI-BEAS-2B-CM和ipdi - hbecc - cm培养BSMC可增加BSMC的增殖和迁移,这是哮喘相关气道重塑的主要特征。IPDI-BEAS-2B-CM和ipdi - hbeck - cm诱导BSMC增殖和迁移与局灶黏附激酶(FAK)、Src、细胞外信号调节激酶(ERK)1/2和AKT活化有关。通过抑制ERK1/2的激活,用特异性抑制剂阻断FAK可显著降低BSMC的迁移和增殖。FAK和ERK1/2抑制剂也能降低IPDI-BEAS-2B-CM-、ipdi - hbecc - cm -和重组人il -8介导的BSMC增殖和迁移,而使用小干扰RNA阻断Rnd3并不能影响BSMC增殖,提示Rnd3仅参与BSMC迁移的调控。gydF4y2Ba
我们的研究表明,抑制IL-8或IL-8介导的FAK/ERK/Rnd3信号是ipdi介导的哮喘有吸引力的治疗靶点。gydF4y2Ba
暴露于异氰酸酯和其他低分子量化学品导致职业性哮喘的5-15%,这是一个主要的健康问题gydF4y2Ba1gydF4y2Ba.异福尔酮二异氰酸酯(IPDI),一种用于制造聚氨酯塑料的脂肪族二异氰酸酯,据报道引起职业性成人哮喘gydF4y2Ba2gydF4y2Ba,gydF4y2Ba3.gydF4y2Ba.二异氰酸酯哮喘患者和实验动物的支气管和淋巴结活检特征显示出与特应性哮喘相似的病理特征,包括炎症和t辅助细胞2型反应增加,气道重塑增强gydF4y2Ba1gydF4y2Ba,gydF4y2Ba4gydF4y2Ba.气道重塑的主要特征包括:上皮完整性丧失、上皮下纤维化、杯状细胞和粘膜下腺增大、支气管平滑肌增重和血管生成增加gydF4y2Ba5gydF4y2Ba.人支气管平滑肌细胞(BSMCs)在气道张力调节中起关键作用。在哮喘气道中,BSMCs不仅增加分泌和增殖能力,而且还迁移到上皮下区域gydF4y2Ba5gydF4y2Ba,gydF4y2Ba6gydF4y2Ba.此外,BSMCs还释放促炎细胞因子,这是导致哮喘发病进程的原因。支气管平滑肌质量的变化程度与哮喘的严重程度有关gydF4y2Ba6gydF4y2Ba.gydF4y2Ba
黏附灶激酶(Focal adhesion kinase, FAK)是一种非受体蛋白酪氨酸激酶,参与信号级联的介导,在细胞增殖、存活、黏附和运动中发挥重要作用gydF4y2Ba7gydF4y2Ba,gydF4y2Ba8gydF4y2Ba.FAK的活化受第6个酪氨酸残基的磷酸化调控。FAK在397酪氨酸处的自磷酸化形成Src的结合位点,而Src又在Tyr 576/577处磷酸化FAK,这对FAK激酶的最佳活性至关重要。FAK - src复合物还在Tyr 925处磷酸化FAK,这为adaptor生长因子受体结合蛋白-2形成一个对接位点,并激活有丝分裂原激活蛋白激酶(MAPK)途径gydF4y2Ba9gydF4y2Ba.Rho鸟苷三磷酸酶Rnd3/RhoE可被Raf-MEK-ERK激活gydF4y2Ba10gydF4y2Ba,已被报道参与许多细胞功能,包括细胞迁移、增殖、细胞周期进展、凋亡和分化gydF4y2Ba11gydF4y2Ba- - - - - -gydF4y2Ba13gydF4y2Ba.在成纤维细胞和上皮细胞中,Rnd3/RhoE过表达破坏肌动蛋白细胞骨架组织和局灶性粘连gydF4y2Ba14gydF4y2Ba.例如,Rnd3的高表达已经被发现可以增加黑素瘤的侵袭能力gydF4y2Ba11gydF4y2Ba,但Rnd3在BSMCs中的功能研究较少。gydF4y2Ba
我们假设IPDI可能导致上皮细胞产生可溶性因子,进而增加BSMCs的增殖和迁移。因此,我们用ipdi处理的支气管上皮细胞条件培养基处理BSMC,并评估BSMC增殖和迁移的机制。该模型提供了支气管上皮细胞与BSMC相互作用的证据,以及IPDI暴露后BSMC迁移的机制。gydF4y2Ba
方法gydF4y2Ba
细胞培养和条件培养基gydF4y2Ba
不朽的,非致瘤的人支气管上皮细胞(细胞系BEAS-2B;美国型培养收集号为CRL-9609),培养于添加10%热灭活胎牛血清的Dulbecco改良Eagle’s培养基/F-12gydF4y2BalgydF4y2Ba谷氨酰胺,100 U·毫升gydF4y2Ba−1gydF4y2Ba青霉素100 μg·mLgydF4y2Ba−1gydF4y2Ba在含5% CO的湿化空气中37°C下的链霉素gydF4y2Ba2gydF4y2Ba.原发正常人BSMCs和原发正常人支气管上皮细胞(HBECs)取自Lonza (Walkersville, MD, USA)。BSMCs在平滑肌生长培养基2 (Lonza)中培养,HBECs在支气管上皮细胞生长培养基(Lonza)中培养。gydF4y2Ba
获得ipdi处理的BEAS-2B和hbech条件培养基(分别为IPDI-BEAS-2B-CM和ipdi - hbech - cm)、BEAS-2B细胞和HBECs (2×10gydF4y2Ba6gydF4y2Ba细胞培养皿),用载体对照或不同浓度的IPID处理6 h。处理后,更换培养液,培养24 h后收集上清液。使用抗IL-8抗体(2 μg·mL)从IPDI-BEAS-2B-CM和IPDI-HBEC-CM中去除白细胞介素(IL)-8gydF4y2Ba−1gydF4y2Ba)和Sepharose A/G珠按标准免疫沉淀协议。使用IL-8 ELISA试剂盒(R&D Systems Europe, Abingdon, UK)证实细胞因子耗竭。gydF4y2Ba
酶联免疫吸附试验和细胞因子阵列gydF4y2Ba
采用elisa试剂盒(R&D Systems Europe)检测IL-8、CXC趋化因子配体(CXCL)5和IL-1β水平。根据生产说明书进行酶联免疫吸附试验。RayBio®Human Inflammation Antibody Array (RayBiotech, Inc., Norcross, GA, USA)还根据制造商说明书评估了ipdi处理的BEAS-2B细胞表达的细胞因子。gydF4y2Ba
细胞增殖gydF4y2Ba
细胞(4×10gydF4y2Ba3.gydF4y2Ba每孔),置于96孔培养板中。孵育24 h后,用载体控制条件培养基IPDI-BEAS-2B-CM或ipdi - hbecc - cm处理细胞72 h。根据制造商的说明书,使用预混WST-1细胞增殖试剂(Clontech Laboratories Inc., Mountain View, CA, USA)测定BSMCs的增殖情况。gydF4y2Ba
细胞迁移试验gydF4y2Ba
根据制造商的说明,使用QCM Chemotaxis 8-μm细胞迁移检测系统(Millipore Corp., Bedford, MA, USA)进行细胞迁移。细胞被播种到迁移室,IPDI-BEAS-2B-CM, ipdi - hbecc - cm,载体控制条件培养基,il -8耗尽的IPDI-BEAS-2B-CM, il -8耗尽的ipdi - hbecc - cm或含有20ng·mL的培养基gydF4y2Ba−1gydF4y2Ba重组人(rh)IL-8置于下腔。细胞迁移24小时后,通过细胞膜迁移的细胞在520 nm的微孔板上进行染色、裂解和定量。gydF4y2Ba
免疫印迹和免疫沉淀反应gydF4y2Ba
使用M-PER裂解试剂(Pierce, Rockford, IL, USA)将细胞冰裂解15分钟。细胞裂解液14000 ×离心gydF4y2BaggydF4y2Ba静置15分钟,收集上清液进行免疫印迹。等量(20 μg·mLgydF4y2Ba−1gydF4y2Ba),用十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(8-12%)分离,转移到聚偏氟乙烯膜上。在5%脱脂奶粉和tris缓冲盐水中封闭1小时后,用一抗孵育膜1-16小时。然后用适当的过氧化物酶结合的二抗处理膜,根据制造商的说明书使用增强化学发光试剂盒(Millipore)检测免疫反应蛋白。gydF4y2Ba
实时RT-PCR和微阵列gydF4y2Ba
使用TRIzol试剂(Invitrogen, Carlsbad, CA, USA)进行RNA分离。采用低聚脱氧胸腺嘧啶引物和逆转录酶(Takara,志贺,日本)按照标准协议制备cDNA。采用SYBR Green在ABI 7500 Real-time PCR系统(Applied Biosystems, Foster City, CA, USA)上进行实时PCR。每个PCR反应混合物中含有200 nM的引物、10 μL的2× SYBR Green PCR Master Mix (Applied Biosystems)、5 μL的cDNA和无核糖核酸水,总体积为20 μL。PCR反应在95°C变性10 min,然后在95°C变性15 s, 60°C变性1 min, 40个循环。所有PCR重复三次,并归一化为内控甘油醛-3-磷酸脱氢酶(GAPDH) mRNA。相对表达式为2gydF4y2Ba- - - - - -δδCTgydF4y2Ba,式中,δ代表“变化”,CT代表浓度。gydF4y2Ba
微阵列实验程序是按照制造商的协议进行的。用Agilent Quick Amp Labeling Kit (Agilent Technologies, Santa Clara, CA, USA)扩增总RNA (1 μg)。IPDI-BEAS-2B-CM-和ipdi - hbeccm处理的BSMC RNA用Cy5标记,对照培养液处理的BSMC RNA用Cy3标记gydF4y2Ba在体外gydF4y2Ba转录过程。0.825μg Cy-labelled cRNA裂解的平均大小大约50 - 100核苷酸通过孵化碎片缓冲区(安捷伦科技)60°C 30分钟。等量的Cy-labelled cRNA汇集和杂交是安捷伦整个人类基因组4×44 k低聚糖微阵列(安捷伦科技)为17 65°C h。洗后用氮枪吹干后,用安捷伦微阵列扫描仪(Agilent Technologies)扫描微阵列,Cy3和Cy5分别在535 nm和625 nm处扫描。扫描图像使用特征提取软件10.5版本(安捷伦技术)进行分析,这是一个用于量化每个特征的信号和背景强度的软件包,它使用秩一致性滤波局部加权散点图平滑方法对数据进行了基本的归一化。gydF4y2Ba
小干扰RNA敲除gydF4y2Ba
用1 μM非靶向或Rnd3小干扰(si)RNA库(Accell;darmacon, Lafayette, CO, USA)在Accell交付介质(Dharmacon),根据制造商的说明。实验采用阳性对照、Accell GAPDH siRNA和非靶向Accell siRNA库。转染72 h后,将培养基交换为完全培养基,细胞因子和紫菜芪的混合物再处理15 h。如前所述,通过实时荧光定量PCR检测Rnd3表达的变化。gydF4y2Ba
统计分析gydF4y2Ba
数据以平均值±表示gydF4y2BasdgydF4y2Ba.采用方差分析对结果进行统计比较。采用Dunnett’s检验分析两组均值之间的显著差异(p<0.05)。gydF4y2Ba
结果gydF4y2Ba
IPDI-BEAS-2B-CM和ipdi - hbecc - cm可促进BSMCs的增殖和迁移gydF4y2Ba
BSMCs体积增大,与支气管上皮细胞距离减小是哮喘气道壁重塑的重要特征gydF4y2Ba5gydF4y2Ba.BSMCs的这些变化可由上皮细胞产生的多种因子诱导gydF4y2Ba5gydF4y2Ba.我们收集IPDI-BEAS-2B-CM和ipdi - hbecc - cm,评估这两种条件培养基对BSMCs增殖和迁移的影响。所示gydF4y2Ba图1一个gydF4y2Ba, IPDI-BEAS-2B-CM和ipdi - hbecc - cm在72 h后以剂量依赖的方式增加BSMC的增殖。此外,IPDI-BEAS-2B-CM和ipdi - hbecc - cm以浓度依赖的方式增加BSMC的迁移(gydF4y2Ba图1 bgydF4y2Ba).gydF4y2Ba
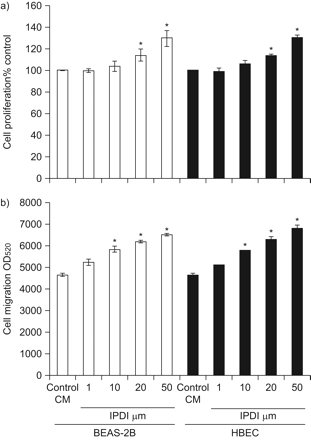
IPDI处理的人支气管上皮细胞(HBEC)-CM (IPDI-HBEC-CM)和IPDI处理的人支气管上皮细胞(HBEC)-CM (IPDI-HBEC-CM)对支气管平滑肌细胞(BSMCs)增殖和迁移的影响。a) IPDI-BEAS-2B-CM和ipdi - hbecc - cm促进BSMC增殖。b) IPDI-BEAS-2B-CM和ipdi - hbecc - cm可增强BSMCs的迁移能力。BEAS-2B细胞和HBECs (2×10gydF4y2Ba6gydF4y2Ba用载体控制或不同浓度IPDI处理6 h。用新鲜培养基替换培养基,孵育24 h后收获细胞。收获培养基定义为IPDI-BEAS-2B-CM或IPDI-HBEC-CM。用Wst-1细胞增殖试剂检测IPDI-BEAS-2B-CM和ipdi - hbech - cm对BSMC增殖的影响。使用QCM趋化细胞迁移检测系统评估BSMC的迁移。ODgydF4y2Ba530gydF4y2Ba:在波长为520 nm处测量的光密度。*: p < 0.05。gydF4y2Ba
IPDI-BEAS-2B-CM和ipdi - hbecc - cm增加炎症、粘附和趋化因子的表达gydF4y2Ba
BSMCs炎症反应的增加是气道重塑发展的主要特征gydF4y2Ba15gydF4y2Ba.我们评估IPDI是否通过上皮细胞和支气管平滑肌的交互作用增加炎症反应。所示gydF4y2Ba图2一个gydF4y2BaIPDI-BEAS-2B-CM可增加炎症因子的转录,包括IL-1β、IL-6、IL-8、CXCL2、CXCL3、CXCL5、C-C motif ligant (CCL)5和细胞间粘附分子。这些数据通过IPDI-BEAS-2B-CM和ipdi - hbecc - cm处理的BSMCs (gydF4y2Ba图2 bgydF4y2Ba).同样,IPDI-BEAS-2B-CM和ipdi - hbecc - cm也能在蛋白水平上增加IL-8、IL-1β和CXCL5 (gydF4y2Ba图2汉英gydF4y2Ba).gydF4y2Ba
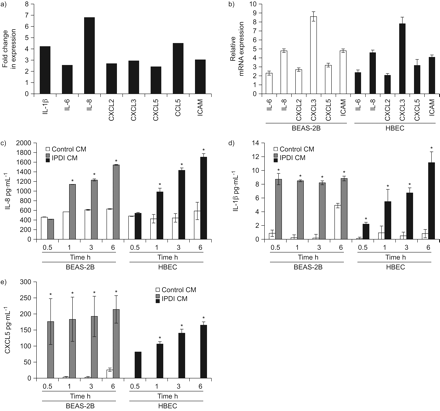
50 μM异氟酮二异氰酸酯(IPDI)处理的BEAS-2B细胞条件培养基(CM) (IPDI-BEAS-2B-CM)和IPDI处理的人支气管上皮细胞(HBEC)-CM (IPDI-HBEC-CM)可增加支气管平滑肌细胞(BSMCs)的炎症和趋化反应。IPDI-BEAS-2B-CM和ipdi - hbecc - cm在转录水平上增加了炎症因子和趋化性的表达,这通过a)微阵列和b)实时PCR进行了评估。IPDI-BEAS-2B-CM和ipdi - hbecc - cm在蛋白水平上增加了c)白细胞介素(IL)-8, d) IL-1β和e) CXC趋化因子配体(CXCL)5的数量。用IPDI-BEAS-2B-CM和ipdi - hbecc - cm处理BSMCs 6 h,通过a)芯片和b)实时PCR检测mRNA表达。c-e)用IPDI-BEAS-2B-CM和ipdi - hbecc - cm处理BSMCs,在规定的时间内,用ELISA检测不同蛋白的含量。ICAM:细胞间粘附分子。*: p < 0.05。gydF4y2Ba
IL-8参与IPDI-BEAS-2B-CM-和ipdi - hbecc - cm介导的增殖和迁移gydF4y2Ba
上皮炎症反应的增加在哮喘的发展中起重要作用gydF4y2Ba16gydF4y2Ba,gydF4y2Ba17gydF4y2Ba.因此,我们评估了IPDI是否增加了上皮细胞中炎症因子的产生,从而增强了BSMCs的增殖和迁移。我们首先通过细胞因子阵列分析确定了IPDI治疗后BEAS-2B细胞的细胞因子谱。数据显示IPDI仅增加IL-8 (gydF4y2Ba图3 bgydF4y2Ba),经酶联免疫吸附试验证实。所示gydF4y2Ba图3 bgydF4y2Ba, IPDI以剂量依赖的方式增加BEAS-2B细胞和HBECs中IL-8的含量。相比之下,IPDI未能影响BEAS-2B和HBECs中的IL-6、CCL5/RANTES(受激活、正常t细胞表达和分泌调节)和CXCL5水平(数据未显示)。我们还评估了rhIL-8对BSMCs增殖和迁移的影响。结果表明,rhIL-8不仅增加了BSMC的增殖(gydF4y2Ba图3 cgydF4y2Ba),但也增强了BSMC迁移(gydF4y2Ba图3 dgydF4y2Ba).gydF4y2Ba
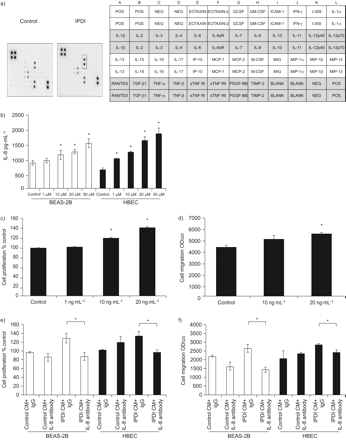
白介素(IL)-8参与了IPDI处理的BEAS-2B细胞条件培养基(CM) (IPDI-BEAS-2B-CM)和IPDI处理的人支气管上皮细胞(HBEC)-CM (IPDI-HBEC-CM)诱导的支气管平滑肌细胞(BSMC)增殖和迁移。通过a)细胞因子阵列分析和b) ELISA检测,IPDI增加了BEAS-2B细胞中IL-8的含量。重组人IL-8的加入增加了BSMC c)增殖和d)迁移。从IPDI-BEAS-2B-CM和ipdi - hbecc - cm中去除IL-8可降低BSMC e)增殖和f) IPDI-BEAS-2B-CM和ipdi - hbecc - cm诱导的迁移。用IPDI (50 μM)处理BEAS-2B细胞和HBECs细胞6 h,收集上清,通过细胞因子阵列分析和ELISA检测IPDI-BEAS-2B- cm和IPDI- HBECs - cm的IL-8水平。POS:积极控制;否定:负控制;GCSF:粒细胞集落刺激因子;GM-CSF:粒细胞-巨噬细胞集落刺激因子;ICAM:细胞间粘附分子; IFN: interferon; sR: soluble receptor; IP: IFN-γ-induced protein; MCP: monocyte chemotactic protein; M-CSF: macrophage colony-stimulating factor; MIG: monokine induced by IFN-γ; MIP: macrophage inflammatory protein; TGF: transforming growth factor; TNF: tumour necrosis factor; sTNFR: soluble TNF receptor; PDGF: platelet-derived growth factor; TIMP: tissue inhibitor of metalloproteases; OD520gydF4y2Ba:在波长为520 nm处测量的光密度。*: p < 0.05。gydF4y2Ba
接下来,我们清空IL-8的IPDI-BEAS-2B-CM和ipdi - hbecc - cm,以确定IL-8在BSMC增殖和迁移中的作用。IL-8 ELISA证实IPDI-BEAS-2B-CM和ipdi - hbecc - cm中IL-8的成功消耗(数据未显示)。所示gydF4y2Ba图3 egydF4y2Ba将il -8耗尽的IPDI-BEAS-2B-CM和ipdi - hbecc - cm加入BSMCs中,有效逆转IPDI-BEAS-2B-CM和ipdi - hbecc - cm引起的BSMCs增殖。同样,IPDI-BEAS-2B-CM和ipdi - hbecc - cm诱导的BSMC迁移增加在IL-8耗尽后终止(gydF4y2Ba图3 fgydF4y2Ba).gydF4y2Ba
IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8对FAK/Rnd3信号通路的影响gydF4y2Ba
FAK可能整合生长因子、细胞因子和整合素信号,参与细胞迁移。我们评估了IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8是否诱导BSMC迁移gydF4y2Ba通过gydF4y2BaFAK。所示gydF4y2Ba图4一gydF4y2BaIPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8增加了BSMCs中FAK Tyr397、Tyr576/577和Tyr925位点的磷酸化。然而,IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8治疗并没有引起总FAK蛋白水平的任何变化。BSMCs暴露于IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8可增加Src的活性形式(Tyr416磷酸化),减少Src的非活性形式(Tyr527)。此外,在IPDI-BEAS-2B-CM-、ipdi - hbecc - cm -和rhil -8处理的BSMC中,FAK和Src的相关性以时间依赖性的方式增加,通过免疫沉淀法(gydF4y2Ba图4 bgydF4y2Ba).FAK的另外两个下游靶点,ERK1/2和AKT的磷酸化形式也被观察到类似的反应(gydF4y2Ba图4gydF4y2Ba).gydF4y2Ba
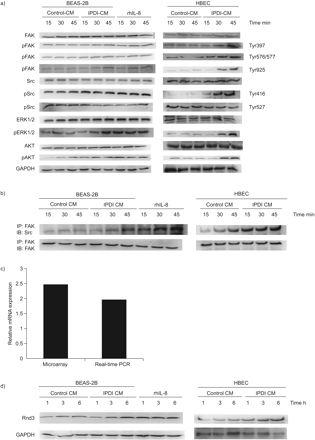
50 μM异氟酮二异氰酸酯(IPDI)处理的BEAS-2B细胞条件培养基(CM) (IPDI-BEAS-2B-CM)、IPDI处理的人支气管上皮细胞(HBEC)-CM (IPDI-HBEC-CM)和重组人白细胞介素(rhIL)-8激活局部粘附激酶(FAK)信号。a) IPDI-BEAS-2B-CM、ipdi - hbeck - cm和rhIL-8可增加支气管平滑肌细胞(BSMCs)中FAK、Src、ERK 1/2和AKT的磷酸化。b) IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8增强了FAK与Src的相互作用。IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8在c) mRNA和d)蛋白水平上都增加了Rnd3的数量。用IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8 (20 ng·mL)处理BSMCsgydF4y2Ba−1gydF4y2Ba)以查询指定的时间。通过免疫印迹(IB)检测磷酸化(p)和未磷酸化(p)蛋白,通过免疫沉淀(IP)检测FAK和Src的相互作用。采用芯片分析和实时荧光定量PCR检测Rnd3 mRNA的表达量。GAPDH:甘油醛磷酸脱氢酶。gydF4y2Ba
因为Rnd3/RhoE是MEK-ERK1/2信号通路的靶点gydF4y2Ba12gydF4y2Ba,我们评估Rnd3在BSMCs中的表达。所示gydF4y2Ba图4 cgydF4y2Bad, IPDI-BEAS-2B-CM, ipdi - hbecc - cm和rhIL-8增加了BSMCs中Rnd3的mRNA和蛋白水平。gydF4y2Ba
FAK和ERK在BSMC增殖和迁移中的作用gydF4y2Ba
为了研究Rnd3上游的因素,我们使用特定的化学抑制剂评估了FAK和ERK1/2在Rnd3表达中的作用。用FAK抑制剂14 (FAK抑制剂)和PD98059 (ERK1/2抑制剂)预处理BSMCs,然后用IPDI-BEAS-2B-CM、ipdi - hbech - cm和rhIL-8处理细胞。然后检测FAK和ERK抑制剂对Rnd3表达、细胞迁移和增殖的影响。用FAK抑制剂预处理BSMCs可减少IPDI-BEAS-2B-CM、ipdi - hbecc - cm或rhIL-8诱导的FAK磷酸化(gydF4y2Ba图5gydF4y2Ba).在IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8处理后,特异性FAK抑制剂停止BSMCs中ERK1/2磷酸化和Rnd3上调(gydF4y2Ba图5gydF4y2Ba).IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8处理后,BSMCs细胞增殖和迁移也停止(gydF4y2Ba图5中gydF4y2Ba).gydF4y2Ba
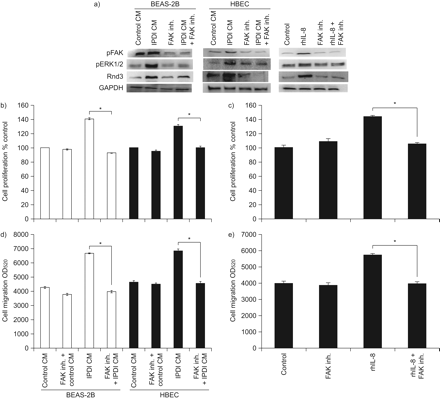
局灶黏附激酶(FAK)在支气管平滑肌细胞(BSMC)增殖和迁移中的作用。a) FAK抑制剂(inh.)对FAK和细胞外信号调节激酶(ERK)1/2磷酸化(p)和Rnd3表达的影响。FAK inh的效果。b)异氟酮二异氰酸酯(IPDI)处理的BEAS-2B细胞条件培养基(CM) (IPDI-BEAS-2B-CM)和IPDI处理的人支气管上皮细胞(HBEC)-CM (IPDI-HBEC-CM)和重组人白细胞介素(rhIL)-8对BSMC增殖的影响。FAK inh的效果。d) IPDI-BEAS-2B-CM和ipdi - hbecc - cm诱导BSMC迁移,e) rhIL-8诱导BSMC迁移。用FAK inh预处理BSMCs。1 h后,再分别暴露于IPDI-BEAS-2B-CM、ipdi - hbecc - cm和hil -8 (ERK磷酸化45 min, Rnd3表达3 h,增殖72 h,迁移24 h)。GAPDH:甘油醛磷酸脱氢酶;ODgydF4y2Ba520gydF4y2Ba:在波长为520 nm处测量的光密度。*: p < 0.05。gydF4y2Ba
PD98059特异性抑制ERK1/2激活可降低BSMCs中IPDI-BEAS-2B-CM-、ipdi - hbecc - cm -和rhil -8诱导的Rnd3上调(gydF4y2Ba图6gydF4y2Ba).与FAK抑制相似,PD98059也降低了IPDI-BEAS-2B-CM、ipdi - hbecs - cm和rhIL-8对BSMC增殖和迁移的影响(gydF4y2Ba图6中gydF4y2Ba).gydF4y2Ba
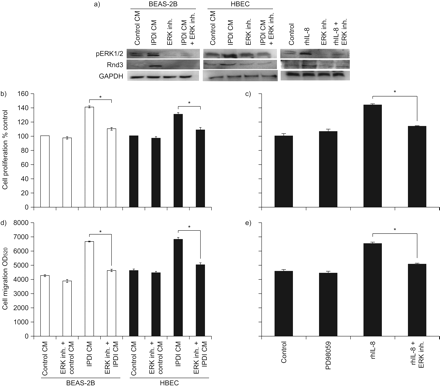
细胞外信号调节激酶(ERK)1/2在支气管平滑肌细胞(BSMC)增殖和迁移中的作用。a) ERK抑制剂(inh.)对ERK1/2磷酸化(p)和Rnd3表达的影响。ERK inh的作用。b)异氟酮二异氰酸酯(IPDI)处理的BEAS-2B细胞条件培养基(CM) (IPDI-BEAS-2B-CM)和IPDI处理的人支气管上皮细胞(HBEC)-CM (IPDI-HBEC-CM)和重组人白细胞介素(rhIL)-8诱导BSMC增殖。ERK inh的作用。d) IPDI-BEAS-2B-CM和ipdi - hbecc - cm, e) rhIL-8引起的BSMC迁移。用PD98059 (20 μM)预处理1 h后,将BSMCs暴露于IPDI-BEAS-2B-CM、ipdi - hbecc - cm和hil -8 (ERK磷酸化45 min, Rnd3表达3 h,增殖72 h,迁移24 h)。GAPDH:甘油醛磷酸脱氢酶;ODgydF4y2Ba520gydF4y2Ba:在波长为520 nm处测量的光密度。*: p < 0.05。gydF4y2Ba
Rnd3在IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8诱导的BSMC迁移中的作用gydF4y2Ba
为了证实Rnd3在IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8诱导的BSMC迁移中的核心作用,我们用Rnd3 siRNA转染BSMC。所示gydF4y2Ba图7gydF4y2Ba与对照siRNA转染相比,Rnd3 siRNA降低了Rnd3表达约90%。特异性敲低Rnd3表达可降低IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8对BSMC迁移的影响,但对BSMC增殖没有影响(gydF4y2Ba图7中gydF4y2Ba).这些结果表明,Rnd3可能在ipdi介导的BSMC迁移中发挥关键作用。gydF4y2Ba
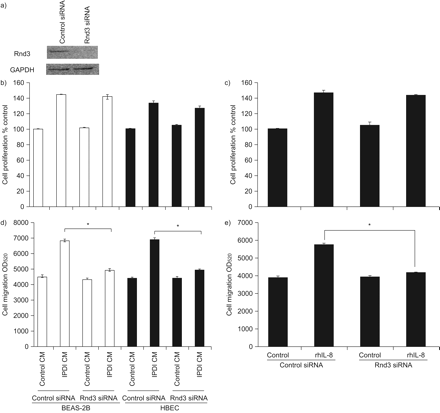
Rnd3在支气管平滑肌细胞(BSMC)增殖和迁移中的作用。a)用小干扰(si)RNA敲除Rnd3。Rnd3抑制不影响50 μM异氟酮二异氰酸酯(IPDI)处理的BEAS-2B细胞条件培养基(CM) (IPDI-BEAS-2B-CM)或IPDI处理的人支气管上皮细胞(HBEC)-CM (IPDI-HBEC-CM)或20 ng·mL的诱导效果gydF4y2Ba−1gydF4y2Ba重组人白细胞介素(rhIL)-8对BSMC增殖的影响。Rnd3抑制减弱d) IPDI-BEAS-2B-CM-和ipdi - hbecc - cm -, e) rhil -8介导的BSMC迁移。GAPDH:甘油醛磷酸脱氢酶。*: p < 0.05。gydF4y2Ba
讨论gydF4y2Ba
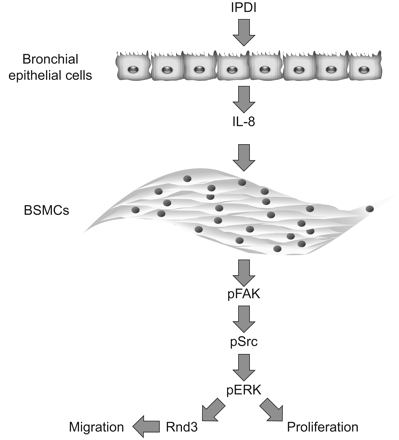
本研究首次探讨暴露于环境化学剂IPDI后气道上皮与平滑肌的相互作用。IPDI可使永生化、非致瘤性人支气管上皮细胞(BEAS-2B细胞系)和原代正常HBECs产生IL-8,从而增加原代正常人BSMCs的增殖和迁移。此外,IPDI-BEAS-2B-CM、ipdi - hbecc - cm和hil -8增加了BSMCs中FAK的激活,随后增加了ERK1/2和AKT的激活。总之,使用特异性抑制剂阻断上游分子FAK或下游因子ERK1/2的激活,可以有效逆转其对BSMC增殖和迁移的诱导作用。此外,siRNA抑制Rnd3也可降低IPDI-BEAS-2B-CM-、ipdi - hbecc - cm -和rhil -8介导的BSMC迁移。这些数据表明,FAK、ERK1/2和Rnd3在ipdi介导的哮喘中发挥关键作用(gydF4y2Ba图8gydF4y2Ba).gydF4y2Ba
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
异佛尔酮二异氰酸酯(IPDI)诱发哮喘的分子机制。IPDI可使支气管上皮细胞产生白介素(IL)-8,增加支气管平滑肌细胞(BSMCs)的增殖和迁移。条件培养基中的IL-8增强了BSMC的生长和运动,而局灶黏附激酶(FAK)、细胞外信号调节激酶(ERK)和Rnd3被发现负责BSMC的迁移。p:磷酸化。gydF4y2Ba
CXCL8/IL-8是一种促炎CXC趋化因子,由多种暴露于促炎细胞因子(如IL-1和肿瘤坏死因子-α)的人支气管上皮细胞或BSMCs分泌,并促进炎症反应gydF4y2Ba17gydF4y2Ba,gydF4y2Ba18gydF4y2Ba.据报道,IL-8通过功能CXC趋化因子受体(CXCR)1和CXCR2触发BSMCs中钙的释放、收缩和迁移gydF4y2Ba18gydF4y2Ba,gydF4y2Ba19gydF4y2Ba.另一种异氰酸酯,甲苯二异氰酸酯,已被发现可增加支气管上皮细胞IL-8的产生,IL-8反过来招募中性粒细胞,导致炎症反应gydF4y2Ba20.gydF4y2Ba,gydF4y2Ba21gydF4y2Ba.然而,IL-8在ipdi相关性哮喘中的作用尚不清楚。我们发现IPDI可以促进BEAS-2B细胞和HBEC分泌IL-8,从而促进BSMCs的增殖和迁移。然而,从IPDI-BEAS-2B-CM和IPDI-HBEC-CM中消耗IL-8逆转了这一趋势。IPDI治疗BSMC也可促进BSMC的增殖和迁移。这与我们发现ipdi介导的哮喘中IL-8水平升高的临床意义具有重要的相关性。gydF4y2Ba
FAK是一种非受体酪氨酸激酶,已被证明是多种细胞类型中细胞增殖、存活和迁移的关键调解者gydF4y2Ba22gydF4y2Ba.许多研究报道,FAK的激活与血管平滑肌细胞的生长和迁移密切相关,而FAK激活的缺失抑制了平滑肌细胞的增殖和迁移gydF4y2Ba22gydF4y2Ba,gydF4y2Ba23gydF4y2Ba.活化的FAK可与Src结合形成复合物,解除Tyr527位点的失活磷酸化,并促进Tyr416位点的活化自磷酸化,导致Src激活。活化Src已被证明可使FAK的多个酪氨酸残基磷酸化,随后激活AKT和ERK1/2,并促进细胞增殖和迁移。虽然林gydF4y2Baet al。gydF4y2Ba22gydF4y2Ba报道反义寡脱氧核苷酸抑制FAK可抑制人肺动脉平滑肌细胞增殖,导致细胞凋亡,但FAK对支气管平滑肌的影响尚不清楚。在我们的研究中,我们发现IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8增加了FAK在三个不同酪氨酸位点的磷酸化。并增强了FAK和Src的相互作用。此外,BSMCs暴露于IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8也可激活AKT和ERK1/2。抑制FAK活化降低了ERK1/2的活化,说明FAK是ERK1/2的上游分子。此外,化学抑制剂对FAK和ERK1/2的选择性抑制也降低了IPDI-BEAS-2B-CM、ipdi - hbecs - cm或rhIL-8对BSMC增殖和迁移的影响,提示FAK/ERK1/2在IPDI-BEAS-2B-CM、ipdi - hbecs - cm和rhIL-8介导的BSMC变化中发挥了关键作用。gydF4y2Ba
Rnd3/RhoE是一种非典型Rho家族蛋白,在细胞骨架形成、细胞存活、细胞凋亡、细胞周期进展和分化等细胞过程中发挥调节作用gydF4y2Ba23gydF4y2Ba,gydF4y2Ba24gydF4y2Ba.与典型的Rho蛋白不同,Rho蛋白的活性依赖于鸟苷三或二磷酸结合状态,RhoE的激活是由转录或转录后修饰控制的gydF4y2Ba25gydF4y2Ba,gydF4y2Ba26gydF4y2Ba.然而,内源性Rnd3在BSMCs中的作用尚不清楚。最近的研究表明,内源性Rnd3与rho相关蛋白激酶介导的细胞凋亡和成肌细胞排列有关gydF4y2Ba14gydF4y2Ba.在这项研究中,我们证明了用IPDI-BEAS-2B-CM、ipdi - hbecc - cm或rhIL-8治疗BSMCs可在转录和翻译水平上增加Rnd3的表达。通过抑制FAK和ERK1/2, IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhIL-8抑制Rnd3的上调,表明FAK/ERK1/2是Rnd3的上游事件。此外,通过siRNA下调Rnd3可以降低IPDI-BEAS-2B-CM、ipdi - hbecc - cm和rhil -8介导的BSMC迁移,表明Rnd3参与了BSMC的迁移。Rnd3在BSMC迁移中的调节作用为Rnd3参与气道重塑提供了重要的新功能。gydF4y2Ba
综上所述,我们的研究结果表明,ipdi处理的上皮细胞条件培养基刺激支气管上皮细胞增殖和迁移。IL-8在条件培养基中增强了BSMC的生长和运动,而FAK, ERK和Rnd3被发现负责BSMC的迁移。鉴于这一发现,抑制IL-8信号通路是ipdi诱发职业性哮喘的一个有吸引力的治疗靶点。gydF4y2Ba
脚注gydF4y2Ba
支持声明gydF4y2Ba
本研究由高雄医科大学环境医学卓越中心(KMU-EM- 98-4)、高雄医科大学研究基金会(KMUQ099002)及高雄医科大学医院(KMUH98-8R21)资助。gydF4y2Ba
感兴趣的语句gydF4y2Ba
没有宣布。gydF4y2Ba
- 收到了gydF4y2Ba2009年12月4日。gydF4y2Ba
- 接受gydF4y2Ba2010年8月16日。gydF4y2Ba
- ©2011人队gydF4y2Ba
参考文献gydF4y2Ba










