





文摘gydF4y2Ba
肺动脉高压(PAH)的特点是过度肺血管重塑涉及管制在内膜增殖的细胞,媒体以及动脉外膜。肺动脉内皮细胞(PAEC)和生存基础内皮增生疾病的病理学。gydF4y2Ba
不可或缺的Notch1参与动脉内皮细胞表型和血管生成提供了有趣的前景为其参与PAH的发病机理。gydF4y2Ba
我们观察到的表达增加Notch1特发性肺的PAH (IPAH)患者和缺氧/ SU5416 (SUHx)老鼠与健康受试者相比。gydF4y2Ba在体外gydF4y2Ba损失和功能研究表明Notch1增加人类PAECs扩散(hPAECs)gydF4y2Ba通过gydF4y2Badownregulation p21和抑制细胞凋亡gydF4y2Ba通过gydF4y2Babcl - 2和存活素。使用γ-secretase抑制剂抑制Notch信号dibenzazepine hPAECs剂量依赖性降低扩散和迁移。值得注意的是,Notch1表达和转录活动增加缺氧下hPAECs和击倒Notch1抑制低氧诱导的细胞增殖。此外,gydF4y2Ba在活的有机体内gydF4y2Ba治疗γ-secretase抑制剂(AMG2008827)显著降低右心室收缩压和右心肥大SUHx老鼠。gydF4y2Ba
这里,我们得出这样的结论:Notch1起着关键作用的多环芳烃和切口抑制剂可能是一个有前途的多环芳烃的治疗选择。gydF4y2Ba
文摘gydF4y2Ba
增加Notch1调节内皮细胞增殖、迁移和生存在PAH,使其小说的目标gydF4y2Bahttp://ow.ly/Z7iD3008DxngydF4y2Ba
介绍gydF4y2Ba
肺动脉高压(PAH)是一种进行性疾病的特点是增加了肺血管阻力(PVR)导致右心肥大并最终死亡的病人由于右心衰。PAH的发病机制涉及到一个复杂和多因子的过程中,内皮细胞功能障碍似乎扮演不可或缺的角色在介导肺血管的结构变化(gydF4y2Ba1gydF4y2Ba]。直接证据支持人口增殖内皮细胞抗凋亡gydF4y2Ba在活的有机体内gydF4y2Ba已经被增加的检测提供了表达各种增殖标记(ki - 67 / MIB-1) (gydF4y2Ba2gydF4y2Ba和血管生成和survival-related分子gydF4y2Ba3gydF4y2Ba,gydF4y2Ba4gydF4y2Ba),如血管内皮生长因子(VEGF)和VEGF受体(VEGFR) 2以及低氧诱导因子(HIF) 1α1β(gydF4y2Ba4gydF4y2Ba]。内皮细胞进一步港口伯灵顿基因的突变,以及减少伯灵顿的表情和caspase-3 [gydF4y2Ba5gydF4y2Ba,gydF4y2Ba6gydF4y2Ba]。此外,内皮细胞与特发性肺动脉高压(IPAH)患者显示增殖增加以应对各种生长因子gydF4y2Ba在体外gydF4y2Ba(gydF4y2Ba5gydF4y2Ba]。因此,针对高度增殖和apoptosis-resistant内皮细胞可以提供多环芳烃的治疗干预。gydF4y2Ba
Notch信号与血管发育,体内平衡和损伤反应(gydF4y2Ba7gydF4y2Ba- - - - - -gydF4y2Ba9gydF4y2Ba]。切口受体(Notch1-4)是跨膜蛋白,显示在细胞表面形成。受体和配体之间的相互作用(锯齿状或Delta-like配体(DLL))相邻细胞膜上表达导致激活信号。结合配体触发两个切口受体蛋白水解分裂。第二个乳沟由γ-secretase释放胞内域研究所,然后把原子核。在细胞核中,镍镉与dna结合蛋白质中超,也称为复合信号序列结合蛋白Jκ(RBP-Jκ),导致切口目标基因的转录,进而控制大量的细胞过程,如干细胞维护,细胞命运规范、分化、增殖和凋亡[gydF4y2Ba10gydF4y2Ba]。gydF4y2Ba
以前,我gydF4y2Ba我gydF4y2Baet al。gydF4y2Ba(gydF4y2Ba11gydF4y2Ba]报道的激活增加Notch3信号在肺动脉平滑肌细胞(PASMCs) PAH患者和突变Notch3已确定患者的多环芳烃和促进细胞增殖和生存能力gydF4y2Ba12gydF4y2Ba]。然而,一些研究也证实了有关Notch1信号血管发展和伤害。Notch1基因敲除小鼠由于缺陷导致胚胎杀伤力somitogenesis和心血管异常(gydF4y2Ba13gydF4y2Ba]。Cre-mediated删除内皮Notch1胚胎发育期间也是致命的,证明逮捕增长与损失有关的基因在船舶/心内膜。减少neointima形成杂合的Notch1gydF4y2Ba+ /−gydF4y2Ba老鼠(gydF4y2Ba14gydF4y2Ba)和Hey2gydF4y2Ba−−/gydF4y2Ba(gydF4y2Ba15gydF4y2Ba)后小鼠颈动脉结扎提供强有力的证据在血管损伤反应的作用。一些报告进一步联系Notch1内皮增生(gydF4y2Ba16gydF4y2Ba- - - - - -gydF4y2Ba19gydF4y2Ba]。虽然在系统性血管特征(gydF4y2Ba7gydF4y2Ba和血管损伤gydF4y2Ba8gydF4y2Ba,gydF4y2Ba9gydF4y2Ba),在肺血管Notch1知之甚少。这使我们调查的表达式和潜在贡献Notch1信号在多环芳烃的发病机制。具体来说,我们旨在:1)概要Notch1表达式从IPAH或从患者肺部缺氧/ SU5416 (SUHx)老鼠,2)确定Notch1表达和功能在人类PAECs缺氧(hPAECs), 3)调查Notch1增益的影响,丧失hPAECs增殖和凋亡,和4)调查的影响锅切口hPAECs抑制剂(γ-secretase抑制剂)gydF4y2Ba在体外gydF4y2Ba和SUHx老鼠gydF4y2Ba在活的有机体内gydF4y2Ba。gydF4y2Ba
材料和方法gydF4y2Ba
主细胞和细胞培养gydF4y2Ba
hPAECs买来Lonza(瑞士巴塞尔)。gydF4y2Ba
质粒和小干扰RNAgydF4y2Ba
pFLAG-CMV2-Notch1胞内域(Notch1ICD)质粒是一种礼物Cheol o·乔教授(韩国先进科学技术研究所、大田、韩国)。Notch1子质粒是一种礼物从教授g .保罗Dotto(瑞士洛桑大学、洛桑)。小核RNA)针对Notch1 mRNA (si-Notch1)和小干扰RNA (si-Scr)匆忙从SABiosciences购买(美国马里兰州弗雷德里克)。gydF4y2Ba
RNA隔离和实时逆转录酶聚合酶链反应gydF4y2Ba
从组织和细胞总RNA转录使用ImProm-II逆转录系统(WI Promega,麦迪逊,美国),其次是各种基因的实时PCR分析使用在线补充表中描述的引物S2。gydF4y2Ba
免疫印迹分析gydF4y2Ba
组织和hPAECs细胞溶解在radioimmunoprecipitation分析裂解缓冲(美国马热科学、沃尔瑟姆)和溶解产物受到西方印迹使用各种抗体(在线补充表S3)。gydF4y2Ba
评估hPAECs的增殖和细胞凋亡gydF4y2Ba
增殖和凋亡的hPAECs评估使用溴脱氧尿苷掺入试验(瑞士巴塞尔罗氏公司)和gydF4y2Ba原位gydF4y2Ba细胞死亡检测设备(罗氏)根据制造商的指示。伯灵顿/ bcl - 2比例是用来评估细胞凋亡的程度。gydF4y2Ba
评估迁移和管hPAECs的形成gydF4y2Ba
迁移hPAECs评估使用修改后的transwell Boyden室,而使用人工基底膜管的形成进行了分析™试验。gydF4y2Ba
荧光素酶检测gydF4y2Ba
人类胚胎肾(HEK)细胞转染Notch1启动子/报告基因构建缺氧暴露(1% O紧随其后gydF4y2Ba2gydF4y2Ba),使用双荧光素酶报告实验测量的荧光素酶活性(WI Promega,麦迪逊,美国)。gydF4y2Ba
免疫组织化学gydF4y2Ba
石蜡包埋组织切片染色了Notch1使用束射功率管™辣根过氧化物酶底物工具包(美国向量,伯林盖姆,CA)根据制造商的指示。连续切片染色与血管性血友病因子(一种内皮标记)和α-smooth肌肉肌动蛋白(α-actin;另外平滑肌标记)。gydF4y2Ba
超声心动图、血流动力学和右心室肥大测量gydF4y2Ba
经胸廓的超声心动图,血液动力学和组织准备进行如前所述[gydF4y2Ba20.gydF4y2Ba),详细描述在线补充材料。gydF4y2Ba
统计分析gydF4y2Ba
数据意味着±gydF4y2Ba扫描电镜gydF4y2Ba。除非另有说明,统计比较样本由单向方差分析Dunnett的紧随其后gydF4y2Ba事后gydF4y2Ba测试。两个样品比较采用t检验。gydF4y2Ba
结果gydF4y2Ba
Notch1表达和本地化IPAH和供体肺gydF4y2Ba
mRNA的表达Notch1和蛋白质的表达Notch1ICD强烈调节患者的肺匀浆IPAH与捐助者(gydF4y2Ba图1一个gydF4y2Ba- - - - - -gydF4y2BacgydF4y2Ba)。进一步Notch1表达式在microdissected肺血管评估和丛状的病变(gydF4y2Ba图1 dgydF4y2Ba和在线补充图S2)。mRNA的表达有显著增加Notch1 microdissected船只从IPAH患者的观察与供体组织相比,但不是在丛状的病变。同样,在蛋白质水平,我们观察到大幅提高Notch1ICD表达IPAH肺动脉(gydF4y2Ba图1 egydF4y2Ba和gydF4y2BafgydF4y2Ba)。强烈的免疫反应性Notch1观察肺动脉内膜的层的IPAH肺(gydF4y2Ba图1 ggydF4y2Ba),这显然colocalised与增殖细胞核抗原(PCNA)。重大变化,信使rna和蛋白质水平,没有观察到其他受体和配体在IPAH肺匀浆除了DLL1配体(在线补充图S1)。gydF4y2Ba
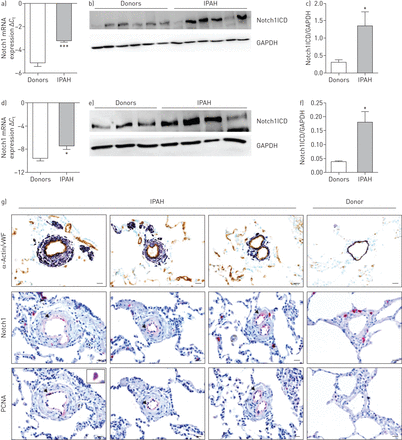
Notch1表达和本地化人力捐赠者和特发性肺动脉高血压(IPAH)肺。信使rna表达的Notch1)肺匀浆和d)肺动脉的捐赠者和IPAH病人。监管在mRNA水平被实时PCR分析使用周期阈值(gydF4y2BaCgydF4y2BatgydF4y2Ba)方法(n = 7)。免疫印迹分析Notch1胞内域(Notch1ICD) b)肺匀浆和e)肺动脉的捐赠者和IPAH病人,其次是c、f)光密度分析。3 -磷酸甘油醛脱氢酶(GAPDH)作为加载控制。*:p < 0.05;* * *:p < 0.001gydF4y2Ba与gydF4y2Ba捐助者。疣状显微图g)代表的人类肺部分从捐赠者和IPAH病人。染色进行Notch1,增殖细胞核抗原(PCNA)、血管性血友病因子(vWF);棕色的;内皮细胞)和α-actin(紫色;平滑肌细胞)。酒吧规模:20μm。gydF4y2Ba
的表达在hPAECs Notch1缺氧对低氧诱导扩散及其影响gydF4y2Ba
暴露hPAECs缺氧(1% OgydF4y2Ba2gydF4y2Ba)导致upregulation Notch1 mRNA表达(gydF4y2Ba图2一个gydF4y2Ba)和Notch1ICD蛋白表达(gydF4y2Ba图2 bgydF4y2Ba和c)。基于存在缺氧反应元素网站Notch1启动子(在线补充图S3),进行双荧光素酶测定与pGL4-Notch1 HEK细胞启动子构建。荧光素酶活性显著增加后缺氧暴露与normoxia (gydF4y2Ba图2 dgydF4y2Ba)。更有趣的是,低氧诱导hPAECs扩散是减少Notch1击倒而si-Scr-transfected细胞(gydF4y2Ba图2 egydF4y2Ba),表明中央的角色Notch1 hPAECs增殖表型的缺氧。特定的击倒Notch1ICD击倒效率(85%)以及减少切口Hes1目标基因的mRNA表达是通过Notch1 Notch2ICD和Notch4ICD (siRNA没有影响gydF4y2Ba图3 bgydF4y2Ba和gydF4y2BacgydF4y2Ba和在线补充图S4A-C)。gydF4y2Ba
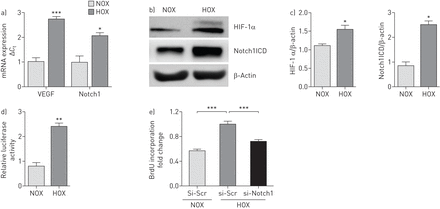
Notch1的表达在人类肺动脉内皮细胞(hPAECs)缺氧对低氧诱导扩散及其影响。hPAECs被暴露于低氧(1%啊gydF4y2Ba2gydF4y2Ba;HOX)和normoxia (NOX) 24 h。一)mRNA的表达血管内皮生长因子(VEGF)和Notch1 HOX hPAECs暴露。监管在mRNA水平被实时PCR分析使用周期阈值(gydF4y2BaCgydF4y2BatgydF4y2Ba)方法。*:p < 0.05;* * *:p < 0.001gydF4y2Ba与gydF4y2Ba氮氧化物的控制。b)免疫印迹分析Notch1胞内域(Notch1ICD)和低氧诱导因子(HIF) 1后面αc)光密度分析。β-Actin被加载控制。*:p < 0.05gydF4y2Ba与gydF4y2Ba氮氧化物。d)人类胚胎肾细胞转染人类Notch1启动子/记者构造和暴露于低氧24 h,和相对荧光素酶活性在细胞提取物用光度计测量,表示为相对荧光素酶活性。所有的值代表的意思是±gydF4y2Ba扫描电镜gydF4y2Ba(n = 9)。* *:p < 0.01gydF4y2Ba与gydF4y2Ba氮氧化物。e) hPAEC扩散以溴脱氧尿苷(BrdU)合并后48 h小干扰RNA (siRNA)针对Notch1 mRNA (si-Notch1)缺氧暴露紧随其后。* * *:p < 0.001gydF4y2Ba与gydF4y2Ba小干扰rna (si-Scr)炒(HOX)。gydF4y2Ba
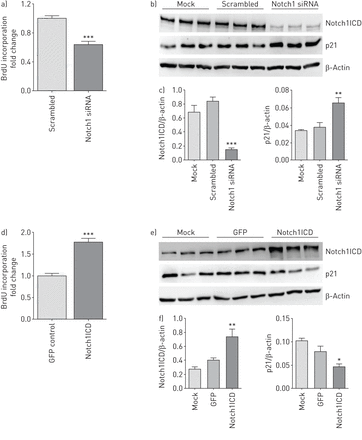
Notch1击倒效果和过度增长medium-induced增殖和细胞周期调控人类肺动脉内皮细胞(hPAECs)。hPAECs是暂时性的转染小核RNA)针对Notch1 mRNA (si-Notch1)或Notch1胞内域(Notch1ICD)质粒48 h。hPAECs对待转染试剂单独是模拟的。hPAEC扩散引起的生长培养基以溴脱氧尿苷(BrdU)合并后48 h) si-Notch1和d) Notch1ICD质粒转染。* * *:p < 0.001gydF4y2Ba与gydF4y2Ba炒核(si-Scr)或绿色荧光蛋白(GFP)控制(n = 5)。b) si-Notch1和e) Notch1ICD超表达在细胞周期调控p21和抗体免疫印迹分析,其次是c、f)的光密度分析的屁股。β-Actin被加载控制。*:p < 0.05;* *:p < 0.01;* * *:p < 0.001gydF4y2Ba与gydF4y2Ba模拟。每组(n = 3。gydF4y2Ba
Notch1击倒效果和超表达hPAECs扩散gydF4y2Ba
在体外gydF4y2Ba研究进行了使用小干扰rna (si-Notch1) Notch1和Notch1ICD质粒在hPAECs Notch1的功能作用进行调查。重要的是,Notch1击倒降低增长medium-induced扩散(∼40%)相比,si-Scr-transfected hPAECs (gydF4y2Ba图3一gydF4y2Ba)。此外Notch1击倒p21的表达增加,内源性细胞周期抑制剂(gydF4y2Ba图3 bgydF4y2Ba和gydF4y2BacgydF4y2Ba),这表明Notch1参与细胞周期调控。gydF4y2Ba
功能性Notch1ICD的过度表达,证实了显著增加mRNA的表达Hes1(在线补充图S5B),导致经济增长大幅增加medium-stimulated hPAECs扩散(77%)与空vector-transfected细胞(gydF4y2Ba图3 dgydF4y2Ba)和相应的减少p21的表达(gydF4y2Ba图3 egydF4y2Ba和gydF4y2BafgydF4y2Ba)。据报道,除了内皮细胞,Notch1在血管平滑肌细胞中表达gydF4y2Ba14gydF4y2Ba,gydF4y2Ba21gydF4y2Ba];然而,并没有改变人类PASMCs观察生长情况的medium-stimulated扩散(hPASMCs)击倒或组成型表达Notch1(在线补充图S6A和F)。gydF4y2Ba
hPAECs的Notch1击倒和超表达对细胞凋亡的影响gydF4y2Ba
击倒Notch1导致∼20%凋亡细胞的数量增加而si-Scr-transfected细胞TUNEL衡量(末端转移酶的dUTP尼克end-labelling)试验(gydF4y2Ba图4一gydF4y2Ba和gydF4y2BabgydF4y2Ba)。此外,免疫印迹分析表明显著增加si-Notch1-transfected伯灵顿/ bcl - 2比例的细胞(gydF4y2Ba图4 cgydF4y2Ba和gydF4y2BadgydF4y2Ba),表明细胞凋亡增加。转染的hPAECs Notch1ICD表达质粒导致显著降低伯灵顿/ bcl - 2比例(gydF4y2Ba图4 egydF4y2Ba和gydF4y2BafgydF4y2Ba)与空向量。生存素的mRNA表达被Notch1击倒而减少gydF4y2Ba反之亦然gydF4y2Ba这是观察与Notch1ICD超表达(gydF4y2Ba图4 ggydF4y2Ba和gydF4y2BahgydF4y2Ba)。gydF4y2Ba
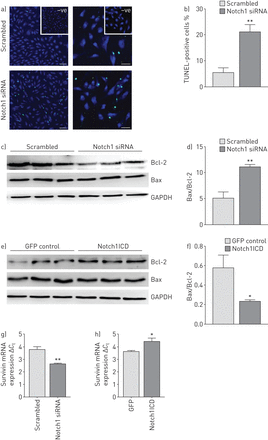
Notch1击倒和超表达对细胞凋亡的影响人类肺动脉内皮细胞(hPAECs)gydF4y2Ba。gydF4y2Ba)代表的图像炒小干扰RNA (siRNA) (si-Scr)和核针对Notch1 mRNA (si-Notch1)转染细胞凋亡使用进行评估gydF4y2Ba原位gydF4y2Ba细胞死亡检测方法(TUNEL末端转移酶的dUTP尼克end-labelling)测定)转染后48 h。酒吧规模:50μm。b)凋亡的比例(TUNEL-positive)细胞使用STEPanizer软件计算。c) Notch1击倒效果和e) Notch1胞内域(Notch1ICD)分析了过度凋亡蛋白通过与抗体蛋白免疫印迹bcl - 2、Bax紧随其后的是d, f)的光密度分析的屁股。3 -磷酸甘油醛脱氢酶(GAPDH)作为加载控制。*:p < 0.05;* *:p < 0.01gydF4y2Ba与gydF4y2Bab, c) si-Scr或d)绿色荧光蛋白(GFP)控制。每组(n = 3。mRNA的表达凋亡蛋白生存素的实时PCR在g) si-Notch1 - h) Notch1ICD-transfected细胞。*:p < 0.05;* *:p < 0.01gydF4y2Ba与gydF4y2Basi-Scr或GFP控制。gydF4y2Ba
药理效应抑制Notch信号扩散,迁移和血管生成反应hPAECsgydF4y2Ba
治疗dibenzazepine (DBZ;γ-secretase抑制剂)γ-secretase活动降低了50%在hPAECs 8 h后所观察到的Notch1ICD(蛋白质含量降低gydF4y2Ba图5一个gydF4y2Ba和gydF4y2BabgydF4y2Ba)和降低Hes1 mRNA表达(gydF4y2Ba图5 cgydF4y2Ba)。5%胎牛血清(FCS) hPAECs扩散导致显著增加(88%),这是减少到只有不到一半(66%)20μM DBZ (gydF4y2Ba图5 dgydF4y2Ba)。增殖诱导VEGF,有趣的是,hPAECs强有力的有丝分裂原,也有效地减少了20μM DBZ 56% (gydF4y2Ba图5 egydF4y2Ba)。此外,FCS-induced hPAECs迁移明显减少DBZ剂量依赖性的方式与20μM DBZ显示减少50% (gydF4y2Ba图5 fgydF4y2Ba和gydF4y2BaggydF4y2Ba),尽管没有影响的观察DBZ管形成响应的hPAECs(在线补充图S7)。数据从工作解剖Notch3 PAH透露,榫眼信号(gydF4y2BaNgydF4y2Ba- - - - - - (gydF4y2BaNgydF4y2Ba(3,5-difluorophenacetyl)gydF4y2BalgydF4y2Ba丙氨酰)-gydF4y2Ba年代gydF4y2Ba苯基甘氨酸gydF4y2BatgydF4y2Ba丁酯),另一个γ-secretase抑制剂,能够减少扩散hPASMCs [gydF4y2Ba11gydF4y2Ba]。同样,DBZ显著降低DNA合成5% FCS -和血小板源生长因子(PDGF) -BB-stimulated hPASMCs剂量依赖性的方式(在线补充图S8a和b)。gydF4y2Ba
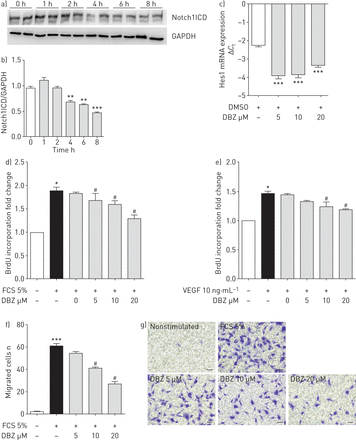
药理效应抑制Notch信号在血清或全身的血管内皮生长因子(VEGF)的肺动脉内皮细胞增殖和迁移(hPAECs)。)免疫印迹分析Notch1胞内域(Notch1ICD)治疗后的hPAECs dibenzazepine (DBZ)和3 -磷酸甘油醛脱氢酶(GAPDH)作为加载控制。b)免疫印迹的量化。GAPDH是作为加载控制。* *:p < 0.01;* * *:p < 0.001gydF4y2Ba与gydF4y2Ba0 h。c) mRNA表达的Hes1 hPAECs 24 h后治疗与DBZ实时PCR分析。所有的值表示为ΔgydF4y2BaCgydF4y2BatgydF4y2Ba。* * *:p < 0.001gydF4y2Ba与gydF4y2Badimethylsuphoxide (DMSO)控制。hPAECs血清饥饿和刺激d) 5%胎牛血清(FCS)或e) 10 ng·毫升gydF4y2Ba−1gydF4y2BaVEGF在各种浓度的DBZ或DMSO(-)和测量扩散的变化由溴脱氧尿苷(BrdU)合并。所有数据都表示为褶皱增加nonstimulated集团,代表的意思是±gydF4y2Ba扫描电镜gydF4y2Ba(n = 5)。f) hPAECs血清饥饿12 h和10米丝裂霉素C处理2 h。细胞被播种transwell插入和降低5%的房间充满了FCS在各种浓度的DBZ或DMSO (-)。16小时后,细胞迁移是通过计算测量晶体violet-positive细胞在显微镜下。*:p < 0.05;* * *:p < 0.001gydF4y2Ba与gydF4y2Banonstimulated;gydF4y2Ba#gydF4y2Ba:p < 0.05gydF4y2Ba与gydF4y2Bad, f) FCS -或e) VEGF-treated组。g)代表transwell迁移试验的照片。酒吧规模:100μm。gydF4y2Ba
Notch1表达SUHx老鼠和治疗评估SUHxγ-secretase抑制剂(AMG2008827)的老鼠gydF4y2Ba
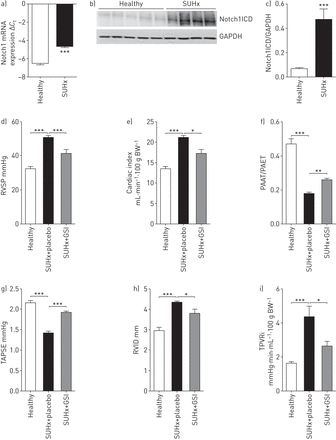
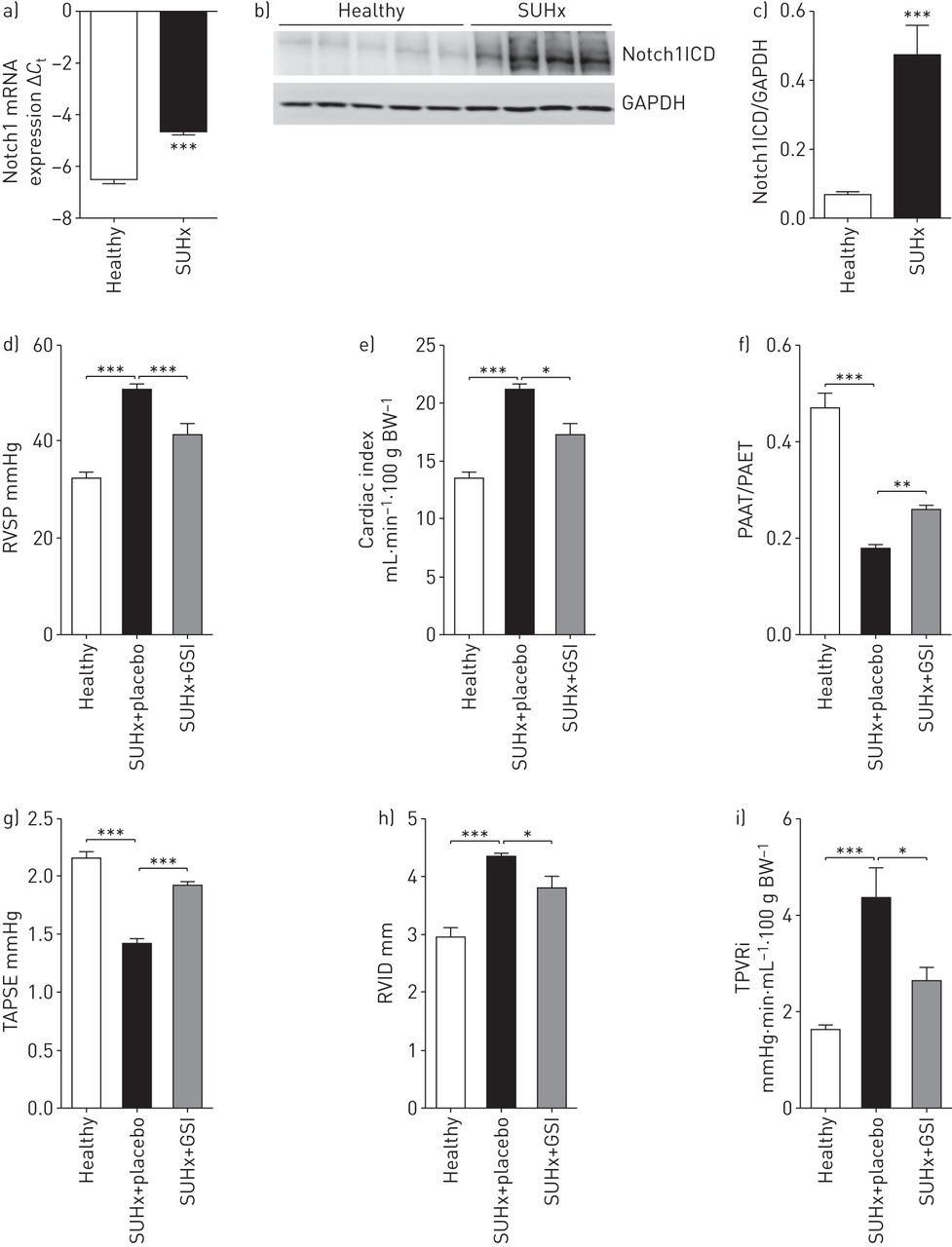
先前的报道表明,注射后大鼠暴露于低氧VEGFR-2抑制剂SU5416展览血管内膜病变,特点是hyperproliferative和apoptosis-resistant内皮细胞(gydF4y2Ba22gydF4y2Ba]。我们观察到一个强大的upregulation Notch1 SUHx信使rna和蛋白质水平的肺的老鼠相比,健康大鼠(gydF4y2Ba图6gydF4y2Ba- - - - - -gydF4y2BacgydF4y2Ba),同时与我们的发现在IPAH肺。同时,Notch3表达式被发现强烈调节(在线补充图S9)。评估在SUHx陷波抑制肺动脉高压的治疗潜力,老鼠Sugen治疗3周后注入和缺氧暴露γ-secretase抑制剂(助教;AMG2008827;安进,千橡市、钙、美国),抑制Notch1信号在细胞记者分析集成电路gydF4y2Ba50gydF4y2Ba(1.28 nM的一半最大抑制浓度)(在线补充图S10),或安慰剂为14天。助教治疗SUHx老鼠导致显著降低右心室收缩压(gydF4y2Ba图6 dgydF4y2Ba)和增加心脏指数(gydF4y2Ba图6 egydF4y2Ba)与安慰剂组相比SUHx老鼠。此外,GSI治疗显著增加肺动脉加速度时间/肺动脉射血时间比和三尖瓣环平面收缩偏差(gydF4y2Ba图6 fgydF4y2Ba和gydF4y2BaggydF4y2Ba),降低肺血管阻力指数和右心室内部直径(gydF4y2Ba图6 hgydF4y2Ba和gydF4y2Ba我gydF4y2Ba以超声心动图。对血管重塑,GSI治疗导致减少neointima /媒体比和内侧壁厚(在线补充图S11)。gydF4y2Ba
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Notch1表达和本地化的肺部缺氧/ SU5416 (SUHx)治疗大鼠和γ-secretase抑制剂的效果(助教;AMG2008827)在大鼠SUHx-induced肺动脉高压。一)mRNA的表达Notch1健康SUHx老鼠的肺匀浆。监管在mRNA水平被实时PCR分析使用周期阈值(gydF4y2BaCgydF4y2BatgydF4y2Ba)方法(n = 4)。b)免疫印迹分析Notch1胞内域(Notch1ICD)在健康SUHx鼠肺匀浆,紧随其后的是c)光密度分析。3 -磷酸甘油醛脱氢酶(GAPDH)作为加载控制。* * *:p < 0.001gydF4y2Ba与gydF4y2Ba健康。d - i)助教对血液动力学的影响和对心脏功能SUHx老鼠。BW:体重。进行超声心动图其次是生理测量GSI-treated BW (3 mg·公斤gydF4y2Ba−1gydF4y2Ba)/安慰剂组SUHx老鼠和健康老鼠SUHx治疗开始后的35天。d)右心室收缩压(RVSP)以右心catheterisation,心脏指数和e), f)肺动脉加速时间(非洲锥虫病)/肺动脉射血时间)(尔马斯·帕依特一起,g)三尖瓣环平面收缩偏差(TAPSE), h)右心室内部直径(RVID)和我)总肺血管阻力指数(TPVRi)老鼠以超声心动图。n = 5健康,n = 7 SUHx安慰剂,n = 5 GSI-treated SUHx。*:p < 0.05;* *:p < 0.01;* * *:p < 0.001gydF4y2Ba与gydF4y2BaSUHx +安慰剂。gydF4y2Ba
讨论gydF4y2Ba
我们的调查Notch信号在多环芳烃的作用让我们以下关键的观察。1)Notch1表达和乳沟增加肺和肺动脉IPAH病人和SUHx老鼠的肺。2)Notch1积极调节hPAECs扩散gydF4y2Ba通过gydF4y2Bap21,消极的调节细胞凋亡gydF4y2Ba通过gydF4y2Babcl - 2和存活素,没有影响hPASMCs扩散,支持endothelial-specific Notch1的影响。3)缺氧是一种刺激导致增加hPAECs Notch1信号。4)抑制信号gydF4y2Ba在体外gydF4y2Ba由DBZ有效减毒扩散hPAECs和hPASMCs,和gydF4y2Ba在活的有机体内gydF4y2Ba管理的GSI (AMG2008827)有说服力地改善右心室收缩压和SUHx老鼠的心脏功能。gydF4y2Ba
肺动脉内皮细胞的PAH患者表现出hyperproliferative和抗凋亡表型(gydF4y2Ba5gydF4y2Ba,gydF4y2Ba23gydF4y2Ba]。体细胞突变骨形态形成蛋白受体II型(BMPRII)和伯灵顿基因已经被鉴定IPAH患者(gydF4y2Ba6gydF4y2Ba,gydF4y2Ba24gydF4y2Ba),这可能会改变内皮细胞的生长优势。然而,BMPRII突变只观察到70%的家族多环芳烃和不是所有内膜的损伤港口体细胞突变。因此,其他分子玩家需要调查。基于我们目前的研究,它是令人信服的猜测Notch1能够成为这样的一个分子中介和Notch1信号调制的PAH患者的内皮细胞可能是有益的。gydF4y2Ba
一个初始的相关性提示Notch1信号在多环芳烃是由我们提供筛选研究显示增加Notch1信号的表达和激活IPAH肺血管的病人。另外,我们观察到增加Notch1表达式在IPAH血管内皮细胞与供体血管。这种变化是与PCNA染色增加,表明Notch1可能参与内皮细胞增殖和neointima形成。我们所知,这是第一次研究证明增加Notch1激活IPAH肺血管的病人。SUHx老鼠,据报道,反映了hyperproliferative内皮细胞严重的人类多环芳烃(病因学gydF4y2Ba22gydF4y2Ba,gydF4y2Ba25gydF4y2Ba),表现出增加Notch1表达和乳沟在肺部与健康的老鼠相比,进一步表明Notch1在内皮细胞增殖和生存gydF4y2Ba体内。gydF4y2Ba切口的激活受体与配体依赖于交互显示在相邻细胞(gydF4y2Ba10gydF4y2Ba]。DLL1配体的表达增加IPAH病人可以代表一个可能的方式诱导Notch1激活。然而,某些报告建议ligand-independent Notch信号激活。一个gydF4y2Ba在体外gydF4y2Ba研究表明增加Notch1的乳沟和激活胚胎干细胞VEGFR-2gydF4y2Ba+gydF4y2Ba剪切应力下细胞(gydF4y2Ba26gydF4y2Ba]。Hyperproliferative改变内皮细胞通常观察IPAH患者的肺动脉分支点被认为是剪切应力增加的结果(gydF4y2Ba23gydF4y2Ba)和SUHx治疗大鼠也已被证明能够反映这些剪切应力条件(gydF4y2Ba22gydF4y2Ba),暗示剪切应力直接Notch1活化剂。此外,额外的多环芳烃因素如活性氧或HIF-1α激活也与Notch1激活。为了定义Notch1激活的机制,我们研究了缺氧作为最初的刺激。缺氧被描述为一个重要的触发导致肺动脉高压(船舶改造gydF4y2Ba27gydF4y2Ba]。在我们的研究中,Notch1表达式,乳沟和启动子活动强烈调节hypoxia-exposed hPAECs HEK细胞(双荧光素酶测定)。这增加可能归因于HIF-1αHIF-1α-binding网站存在于Notch1启动子序列。有趣的是,发现高浓度的HIF-1α动脉病变在肺IPAH和IPAH PAECs [gydF4y2Ba28gydF4y2Ba]。最近的一项研究进一步揭示了激活Notch1癌症细胞系在缺氧gydF4y2Ba通过gydF4y2BaHIF-1α,导致增加增殖、迁移和生存的细胞(gydF4y2Ba29日gydF4y2Ba]。在hPAECs Notch1抑制了低氧诱导扩散。因此,我们得出结论,缺氧会使Notch1 hPAECs信号,进而导致增加扩散。gydF4y2Ba
相似性PAH患者的癌细胞和改变内皮细胞(gydF4y2Ba3gydF4y2Ba,gydF4y2Ba30.gydF4y2Ba),建立Notch1作为癌基因的作用[gydF4y2Ba31日gydF4y2Ba,gydF4y2Ba32gydF4y2Ba),让我们在内皮细胞增殖和细胞凋亡研究的贡献。在我们的研究中,导致差别Notch1对这些∼hPAECs扩散减少40%,而Notch1ICD过度导致显著增加∼75%增殖。先前的报告涉及Notch1在内皮细胞增殖的规定,虽然相互矛盾的结论。在人类主动脉内皮细胞,Notch1ICD过度抑制增殖的细胞gydF4y2Ba通过gydF4y2Ba抑制增殖作用的蛋白激酶和磷酸肌醇3-kinase / Akt通路(gydF4y2Ba16gydF4y2Ba]和p21 [gydF4y2Ba17gydF4y2Ba]。反驳这些报告,TgydF4y2BaakeshitagydF4y2Baet al。gydF4y2Ba(gydF4y2Ba18gydF4y2Ba证明Notch1对VEGF-induced扩散至关重要,迁移和内皮细胞的生存。另一项研究有关减少Notch1乳沟在内皮细胞PCNA水平下降,当cocultured Jagged1-deficient血管平滑肌细胞(gydF4y2Ba19gydF4y2Ba]。探索所涉及的机制,细胞周期调控蛋白的表达。我们观察到的蛋白质含量明显增加,细胞周期抑制剂p21 Notch1击倒,而Notch1ICD超表达与p21的表达下降有关。p21,著名的细胞周期蛋白依赖性激酶抑制剂,被描述为直接目标RBP-Jκ-dependent Notch1信号(gydF4y2Ba17gydF4y2Ba,gydF4y2Ba33gydF4y2Ba],作者证明p21子活动的镇压下Notch1人类脐静脉内皮细胞激活。我们的研究结果支持了积极作用的Notch1 hPAECs扩散gydF4y2Ba通过gydF4y2Ba影响p21的表达。有趣的是,siRNA-mediated Notch1击倒或Notch1ICD超表达对hPASMCs扩散没有任何影响,表明其他切口同系物在调节PASMCs的增殖潜力更重要。事实上,Notch3被描述为异常的基本扩散PASMCs IPAH患者(gydF4y2Ba11gydF4y2Ba]。这一发现进一步支持我们的假设endothelial-specific Notch1的影响。gydF4y2Ba
抗细胞凋亡改变的另一个重要特点是患者内皮细胞中发现多环芳烃(gydF4y2Ba5gydF4y2Ba,gydF4y2Ba6gydF4y2Ba]。Notch1击倒了一个小但重要(20%)TUNEL-positive凋亡内皮细胞的数量增加。许多报道提出,进一步证明了bcl - 2 /伯灵顿比率可能控制细胞凋亡刺激的敏感性(gydF4y2Ba34gydF4y2Ba]。有趣的是,我们能够观察到明显降低的伯灵顿/ bcl - 2比,表明增加细胞凋亡与si-Notch1 hPAECs治疗。在协议,组成型表达Notch1 ICD导致伯灵顿/ bcl - 2比例的增加。生存素,哺乳动物的一员“细胞凋亡抑制剂”家庭,显示被表达在癌症和改建PAH患者的肺动脉(gydF4y2Ba35gydF4y2Ba,gydF4y2Ba36gydF4y2Ba]。我们观察到的积极影响Notch1存活素mRNA表达,表明生存素是下游Notch1信号的目标,符合各种研究表明Notch1上游是一个监管机构的生存素在癌症gydF4y2Ba37gydF4y2Ba]。先前的研究已经定义了一个Notch1信号的动物和抗凋亡的作用gydF4y2Ba在体外gydF4y2Ba细胞模型(gydF4y2Ba38gydF4y2Ba),在这个领域的发展和癌症。然而,没有多少人知道Notch1参与内皮细胞凋亡。我们的研究结果表明Notch1的角色在促进生存hPAECs通过调节bcl - 2的表达和存活素。gydF4y2Ba
药物抑制Notch信号通过γ-secretase抑制剂已广泛应用于各种研究利用不同癌症细胞株和动物模型(gydF4y2Ba39gydF4y2Ba,gydF4y2Ba40gydF4y2Ba)建立的潜在效用γ-secretase inhibitor-based治疗。为了检查的相关性γ-secretase抑制剂治疗实验性肺动脉高压,我们DBZ用于gydF4y2Ba在体外gydF4y2Ba研究。DBZ能够显著降低血清和VEGF-induced内皮细胞扩散。事实上,相声VEGF和等级之间的信号已经被报道。VEGF是证明增加内皮细胞受体和配体的表达(gydF4y2Ba41gydF4y2Ba]。此外,VEGF-induced产后介导的血管生成包括内皮细胞增殖和迁移gydF4y2Ba通过gydF4y2Baγ-secretase Notch1激活(gydF4y2Ba18gydF4y2Ba]。血浆VEGF水平升高的患者严重的多环芳烃,VEGF和VEGFR-2强烈表达丛状的病变(gydF4y2Ba4gydF4y2Ba]。DBZ进一步表现出强烈抑制作用迁移FCS hPAECs刺激的,虽然没有显著影响血管生成反应。据报道,IPAH PAECs显示增加的迁徙可能与控制细胞。但是,没有更大的血管生成的潜力IPAH PAECs观察,而是少有序与薄管形成分支。肺血管生成在开发过程中多环芳烃可能充当保护机制对肺微脉管系统的损失由于内皮损伤和功能障碍,尽管严重的多环芳烃的特点是解除血管生成在内皮细胞增生性病变包括一塌糊涂,交替区域与坚实的细胞和其他领域的核心变量阶段的毛细管/血管形成[gydF4y2Ba42gydF4y2Ba]。切口的实质性影响调制观察增殖,迁移和细胞凋亡hPAECs,但不是血管生成,强烈暗示对切口的有益作用抑制治疗对肺血管生成多环芳烃没有任何负面影响。符合之前的结果(gydF4y2Ba11gydF4y2Ba),血清和PDGF-induced hPASMCs扩散被DBZ有效减少。在PDGF-BB DBZ诱导增殖的影响可以是由于直接影响细胞增殖的切口标记或由于PDGF相声信号。事实上,Notch3显示调节PDGF受体β表达在血管平滑肌细胞(gydF4y2Ba43gydF4y2Ba]。gydF4y2Ba
重要的是,治疗的潜力GSI (AMG2008827;3 mg·公斤gydF4y2Ba−1gydF4y2Ba)gydF4y2Ba在活的有机体内gydF4y2Ba在几个疾病参数建立SUHx-induced肺动脉高压大鼠的评估。抑制剂显著改善血液动力学的参数(右心室收缩压和心脏指数)和右SUHx老鼠相比安慰剂组大鼠心脏功能。此外,减少neointima形成和内侧壁厚与GSI治疗也被观察到。然而,高剂量的抑制剂或长期管理可能需要进一步显示持续的影响。综上所述,这些数据显示实验PAH陷波抑制的保护作用。gydF4y2Ba
研究的局限性gydF4y2Ba
本研究阐明了一个明确的机制调节内皮细胞增殖和生存的Notch1信号gydF4y2Ba在体外gydF4y2Ba和gydF4y2Ba在活的有机体内gydF4y2Ba。还需要进一步的研究来剖析的分子机制(配体依赖和独立)导致Notch1激活处于疾病状态和评估的有效性GSI治疗与其他化合物结合之前报道在SUHx降低肺动脉高压模型。gydF4y2Ba
结论gydF4y2Ba
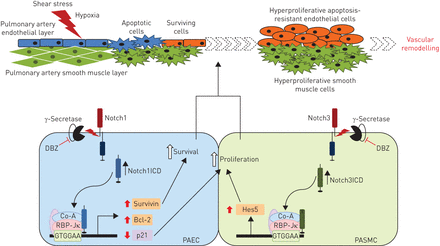
这些数据描述小说涉及Notch1信号通路,调节的前导和pro-survival PAECs表型(gydF4y2Ba图7gydF4y2Ba)。这个途径似乎进一步激活缺氧刺激。这两个gydF4y2Ba在体外gydF4y2Ba和gydF4y2Ba在活的有机体内gydF4y2Ba数据使用药理表明Notch1γ-secretase抑制剂是一个潜在的治疗目标在多环芳烃。gydF4y2Ba
{kind=link}
{kind=link}
Notch信号在肺动脉高血压(PAH)。提出角色等级的示意图表示信号在多环芳烃的肺血管重塑。Notch1ICD: Notch1胞内域;DBZ: dibenzazepine;Co-A:共激活剂;RBP-Jκ:复合信号序列结合蛋白Jκ;PAEC:肺动脉内皮细胞;PASMC:肺动脉平滑肌细胞;Hes5:转录因子Hes5。gydF4y2Ba
确认gydF4y2Ba
我们承认Ewa Bieniek和克里斯蒂娜发呜呜声(吉森和马尔堡大学肺中心,吉森,德国),和Yanina Knepper(马克斯·普朗克心肺研究所坏Nauheim,德国)为他们的有价值的技术援助。gydF4y2Ba
脚注gydF4y2Ba
可以从本文的补充材料gydF4y2Bawww.qdcxjkg.comgydF4y2Ba
支持声明:这项工作得到了资助吉森和马尔堡大学的肺中心(UGMLC)和马克斯·普朗克(MPI)的社会。为这篇文章一直存放在资助信息gydF4y2BaFundRefgydF4y2Ba。gydF4y2Ba
利益冲突:没有宣布。gydF4y2Ba
- 收到了gydF4y2Ba2015年5月18日。gydF4y2Ba
- 接受gydF4y2Ba2016年5月6日。gydF4y2Ba
- 版权©2016人队gydF4y2Ba
引用gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba










