





摘要
ω-3脂肪酸如二十二碳六烯酸(DHA)对心脏的保护作用已得到很好的证实,但DHA对肺动脉高压患者血管张力和压力的调节作用尚不清楚。
DHA是一种有效的钾调节剂+通道,我们假设DHA通过K+从而对肺血管张力和压力产生影响。
我们发现DHA引起钙活化K的剂量依赖激活+(KCa)电流在原发性人PASMCs和内皮依赖性肺动脉舒张。KCa的血管舒张明显减弱- / -(Kcnma1- / -老鼠)。在活的有机体内,急性DHA使慢性缺氧肺动脉高压动物模型右心室收缩压恢复到常氧动物水平。有趣的是,在特发性肺动脉高压中,KCa通道及其亚单位上调。DHA激活了这些人PASMCs中的KCa通道,并使特发性肺动脉高压PASMCs的膜电位与健康供体PASMCs的膜电位超极化。
我们的研究结果表明,DHA激活肺动脉高压的PASMC KCa通道,导致血管舒张。这种效应可能为先前未描述的DHA在肺动脉高压中作为急性血管扩张剂的作用提供了分子解释。
摘要
IPAH重塑肺动脉中KCa通道的增加可能是DHA治疗的契机http://ow.ly/rTUl300pn7R
简介
肺动脉高压(PAH)是一种罕见的危及生命的肺血管疾病,其特征是肺血管阻力的进行性增加,最终导致右心室衰竭。内源性血管扩张性因子的丧失和肺血管扩张能力是该疾病病理特征的主要因素,血管扩张剂治疗多环芳烃的疗效已得到证实[1,2].在正常生理条件下,肺循环是一个低压、低阻力的系统。肺脉管系统的低血管张力主要由肺动脉平滑肌细胞(PASMCs)的静息膜电位维持,主要但不完全由K引起+电导(3.- - - - - -5].因此,K+通道的活性和表达直接影响PASMCs静息膜电位,从而影响肺血管张力。文献提示,特发性多环芳烃(IPAH;原原发性肺动脉高压患者可能有更多的去极化膜电位(他们是慢性去极化),有助于血管收缩和重塑[6],尽管支持这一假设的数据非常稀少。以往研究显示,内源性血管活性物质(如。内皮素-1 (ET-1)、血管紧张素II及血清素)[7,8]、缺氧或药物(达沙替尼及培高利特)[9- - - - - -11]参与肺动脉高压的病理生理,通过抑制K+并导致血管收缩。因此,激活K+通道电流(一氧化氮、伊洛前列素或曲前列素)引起PASMCs膜超极化,导致血管舒张[12].
大规模流行病学研究和随机对照试验证明ω-3脂肪酸(二十碳五烯酸、二十二碳六烯酸(DHA)和α-亚麻酸)可显著降低冠状动脉疾病患者心血管疾病事件的发生[13].因此,美国心脏协会(AHA)的指南建议经常摄入ω-3脂肪酸[14].此外,膳食鱼油可降低慢性缺氧诱导肺动脉高压期间的肺动脉高压和血管重塑[15].然而,为了解释脂肪酸对肺循环的有益作用,还需要继续进行分子机制的研究。DHA可防止ET-1诱导的细胞内游离钙增加,并抑制PASMC增殖[16].此外,我们之前的研究对这些慢性抑制作用背后的分子事件进行了全面分析,并表明DHA促进钙依赖性氧化应激,导致人类原发性pasmc中的未折叠蛋白反应,从而抑制增殖[17,18].DHA是钙依赖性钾的有效激活剂+(KCa)在大鼠冠状动脉平滑肌细胞中的通道电流,促进孤立的小冠状动脉扩张[19,20.].已知DHA可可逆激活由成孔Slo1亚基(Kcnma1)和辅助亚基β1 (Kcnmb1)组成的大电导KCa (BKCa)通道[21].然而,这种重要脂肪酸在肺循环和肺动脉高压中的急性作用的详细机制尚不清楚。
在这项研究中,我们假设DHA激活K+PASMCs中的通道导致静息膜电位的超极化和肺动脉高压的逆转。本研究考察了DHA在肺循环中的急性作用,特别是对K+原代人pasmc的通道功能和膜电位。我们还研究了DHA对肺动脉高压小鼠模型中肺动脉压(PAP)的影响。使用BKCa (Kcnma1- / -)敲除小鼠后,我们研究了KCa通道对DHA在肺循环中的影响的具体贡献。此外,我们还利用了特定区域的分析通过激光捕获显微解剖,并研究了IPAH患者和健康捐赠者肺动脉中BKCa通道和亚单位的表达。最后,我们验证了DHA的效果,观察到在活的有机体内而且在体外该研究主要针对从IPAH患者和健康捐赠者中分离出的原发性人类pasmc。我们发现来自IPAH的pasmc明显去极化,而DHA使这种去极化正常化通过激活KCa。
材料与方法
详细的资料和方法可以在网上的补充资料中找到。
从IPAH和供体组织中分离人pasmc
本研究涉及的所有IPAH患者和供体组织样本的研究均得到奥地利维也纳医科大学伦理委员会的批准,并获得所有研究参与者的书面知情同意。肺组织取自维也纳医科大学胸外科接受肺移植的IPAH患者。未移植的供体肺,为移植而采集,但未被使用(如。因为缩小了肺移植),作为对照。部分样品在液氮中快速冷冻或在4% (w/v)甲醛中固定,分别进行激光显微解剖或组织组织学检查。原发性PASMCs是从患者的抵抗性肺动脉中分离出来的,PASMCs的特征如前所述[10].在研究中给出了供体和患者的临床特征表1.
动物研究
所有实验都是由地方当局根据国家动物实验条例批准的(奥地利教育、科学和文化部;bmwf - 66.010/0076 - 2 / 3 b / 20116)。
统计分析
数值以均数±表示扫描电镜.使用Prism 5 (GraphPad, San Diego, CA, USA)进行统计分析。非参数资料采用Mann-Whitney u检验。采用配对t检验评估dha输注前后右心室收缩压(RVSP)的变化。在组间差异的情况下,评估单向方差分析,然后Tukey's事后测试。p值<0.05为有统计学意义。
结果
DHA激活人原代pasmc中的KCa通道
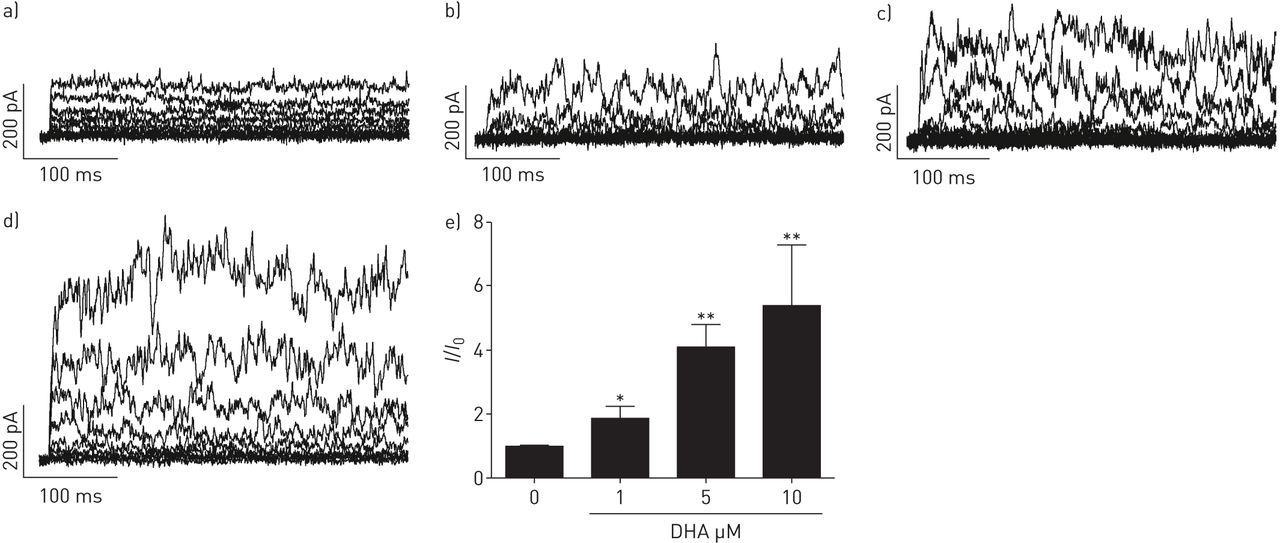
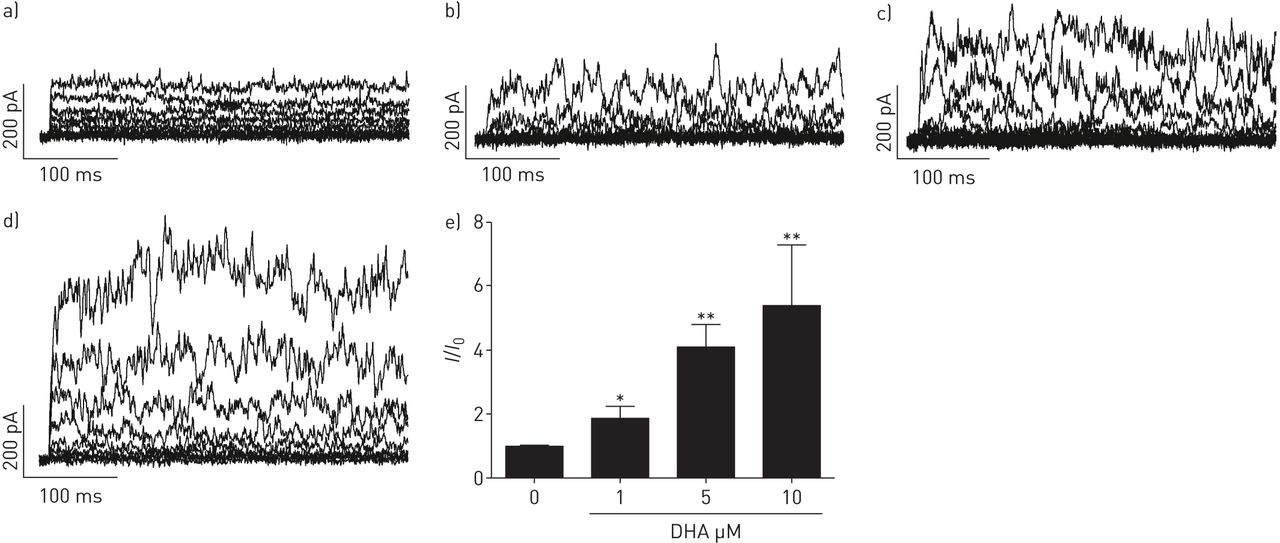
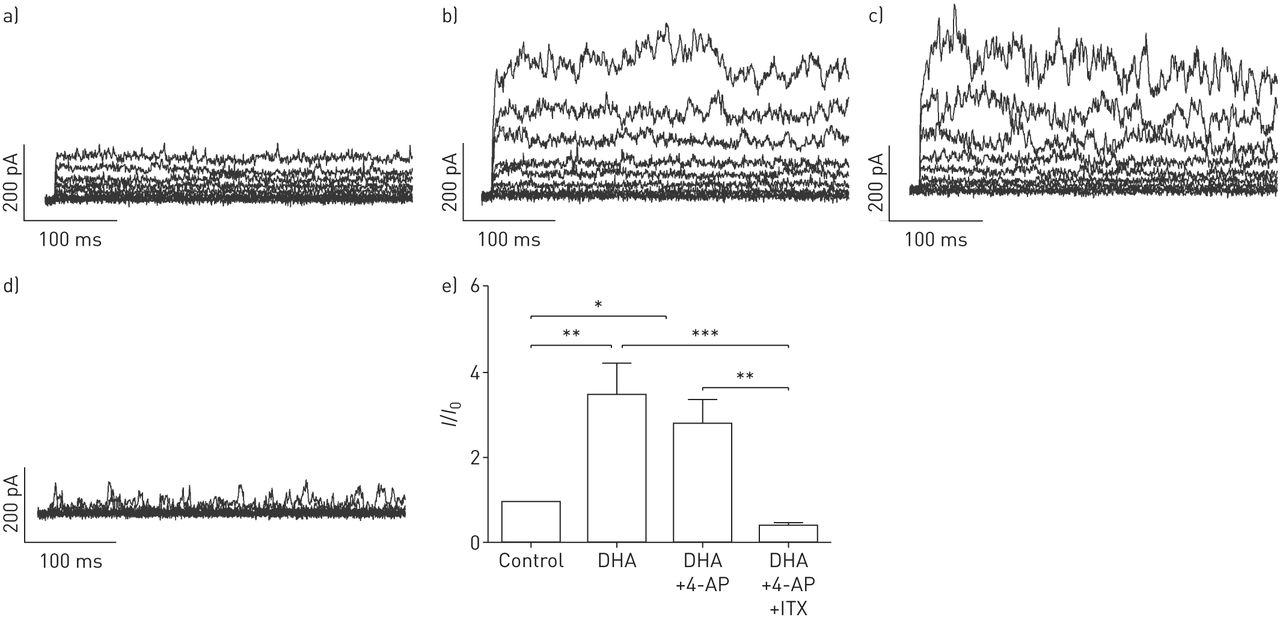
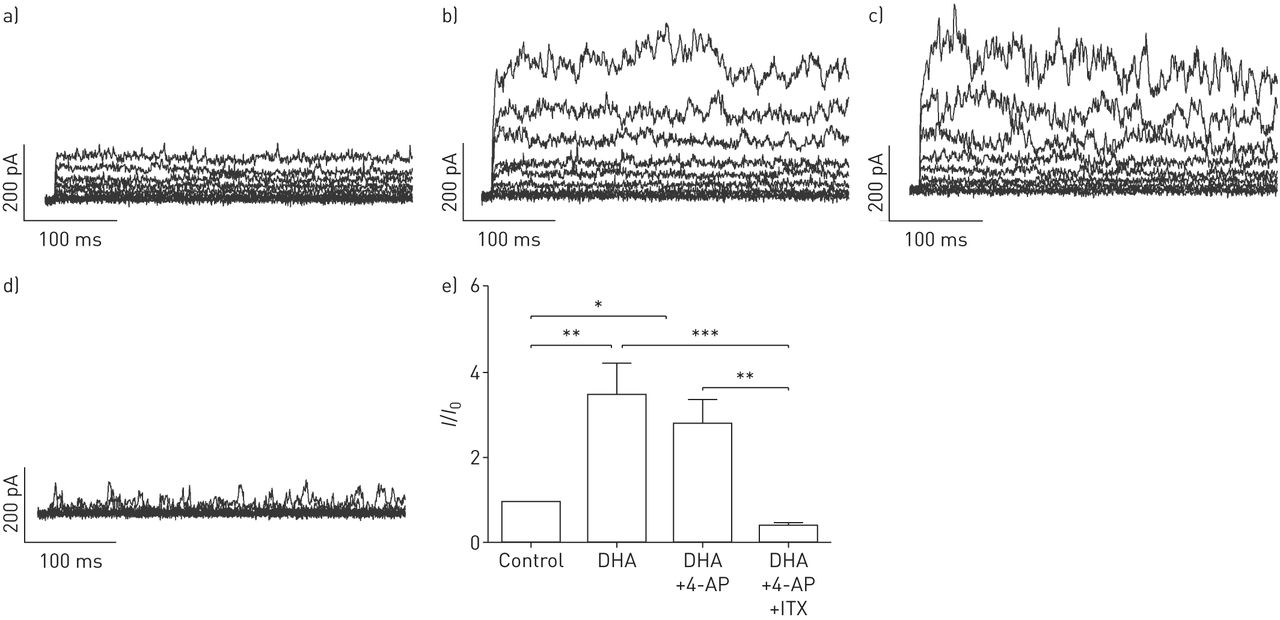
DHA显著且可逆地提高了全细胞钾含量+人PASMCs中保持电位为- 20mv的电流以剂量依赖的方式记录(图1).接下来,要解决激活电流的来源,不同的K+使用通道抑制剂。电压激活K对DHA的增强作用没有影响+通道抑制剂4-氨基吡啶(4-AP, 5µM;图2 d),但由于KCa通道抑制剂伊比利亚毒素(ITX, 100 nM;图2 d).相对K的变化+电流振幅(我/我0)所引起的疾病,总结于图2 e.显然,ITX的降低明显大于4-AP,分数变化在有4-AP和没有4-AP的记录之间没有显著变化。我们的结果表明dha激活电流主要来源于人pasmc中的KCa通道。
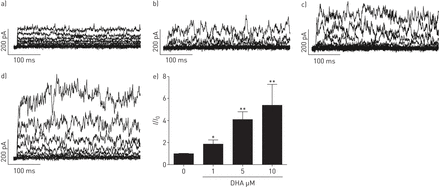
二十二碳六烯酸(DHA)剂量依赖性地激活全细胞钾+电流在人肺动脉平滑肌细胞(PASMCs)。原全细胞K+电流示踪在人pasmc中表现为a)无浓度和随浓度增加b) 1、c) 5和d) 10 μM DHA。e)总结相对大电导KCa (BKCa)通道电流(我/我0)而增加人类主要pasmc中DHA的浓度。每个设置N = 4-6。*: p < 0.05;* *: p < 0.01。
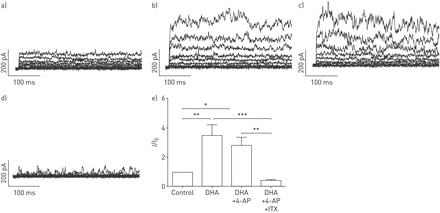
二十二碳六烯酸(DHA)激活人原代肺动脉平滑肌细胞(PASMCs)大导KCa通道(BKCa)。原全细胞K+电流示踪呈现a)不带和带b) DHA, c)带电压门控K的DHA+通道阻滞剂4-氨基吡啶(4-AP), d) DHA以及电压门控K+通路阻滞剂4-AP和BKCa通路阻滞剂伊比利亚毒素(ITX)在原发性人pasmc。e) KCa电流的相对变化总结(我/我0)在人原代pasmc中被DHA诱导。每组N = 5-6。*: p < 0.05;* *: p < 0.01;* * *: p < 0.001。
DHA诱导快速内皮依赖性肺舒张
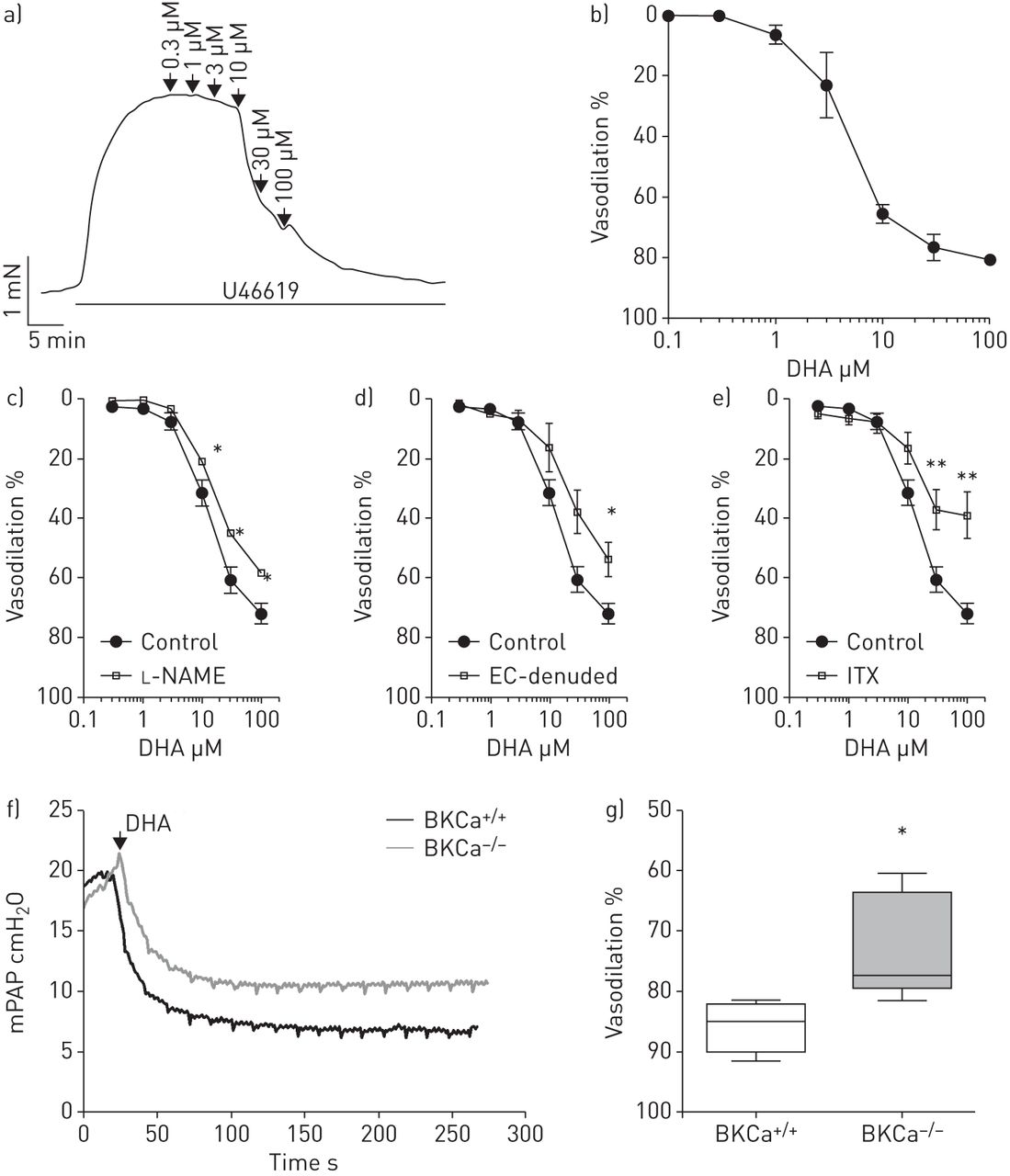
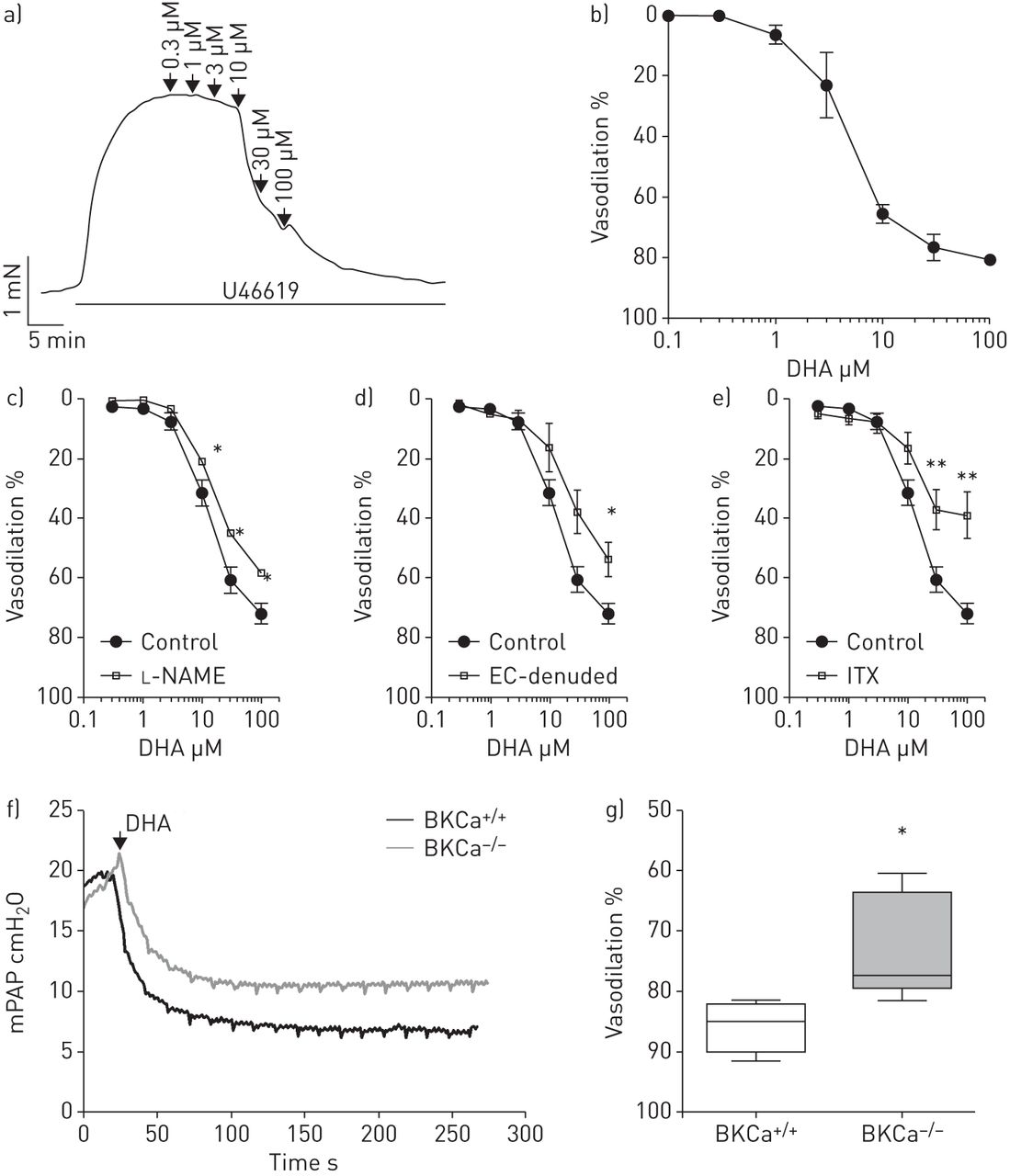
研究我们在人类pasmc中观察到的相关性在体外对肺动脉张力的影响,我们测定了DHA对孤立的预狭窄肺动脉张力的影响。用钢丝肌描记术进行的等长张力测量显示,DHA可诱导U46619 (图3一).DHA的放松作用与浓度有关,最大有效浓度的一半(EC50) 3.5µM (n=6;图3 b).接下来,我们还研究了内皮细胞在dha介导的肺血管舒张中的作用。一氧化氮合酶抑制剂的应用l- name (l-NG-硝基精氨酸甲酯,n=3;图3 c)或内皮剥蚀(n=4;图3 d在等距张力测量中,与对照组(n=6)相比,在大鼠肺动脉中显著降低了对DHA浓度增加的血管舒张反应。接下来,我们研究了BKCa通道在dha诱导弛豫中的作用。存在BKCa通道阻滞剂ITX (n=4;图3 e),与对照组(n=6)相比,大鼠肺内动脉对增加DHA浓度的血管舒张反应显著降低。进一步,我们研究了DHA在完整肺中对PAP的调节。在体外在小鼠离体灌注肺模型中,单独服用DHA没有改变基础平均PAP (mPAP)(在线补充图S1A)。当在DHA应用之前对肺进行U46619处理时,DHA(10µM)导致mPAP大幅下降(11.6±0.9,n=6)(在线补充图S1B)。为了阐明KCa通道在DHA对肺血管舒张功能反应中的作用,小鼠缺乏大导KCa (BKCa) α-亚基- / -)和它们的野生型对应物(BKCa+/+)用于小鼠离体灌注肺模型(图3 f).u46619诱导的前音不受BKCa通道缺失的影响。然而,dha诱导的肺血管舒张在BKCa中被显著抑制- / -小鼠与BKCa比较+/+老鼠(73±4%与86±2%,每组n=5;图3 g).
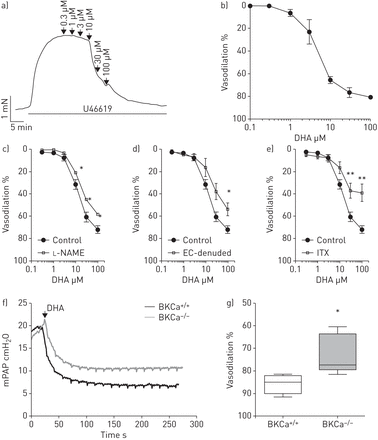
二十二碳六烯酸(DHA)引起肺内动脉内皮依赖性血管舒张。a) U46619缩窄前,在离体小鼠肺内动脉中记录DHA添加浓度的代表性张力。b) DHA对肺内动脉的累积浓度依赖性血管舒张作用。在存在/不存在c时,对DHA浓度增加的反应中,预收缩的离体大鼠肺内动脉的放松l-NAME (n=3), d)内皮细胞(EC) (n=4)或e)伊比利亚毒素(ITX) (n=4)。血管舒张的百分比表示相对于U46619的最大反应。dha诱导的血管舒张作用在BKCa中受损- / -老鼠。f) BKCa的代表性平均肺动脉压(mPAP)示踪+/+和BKCa- / -小鼠离体灌注通气肺模型。g) DHA对BKCa中U46619预张力的急性血管扩张作用+/+和BKCa- / -老鼠。数据以中值、四分位范围、最小值和最大值表示。每组N =5人。*: p < 0.05;* *: p < 0.01。
DHA导致低氧诱导的肺动脉高压的快速肺血管舒张
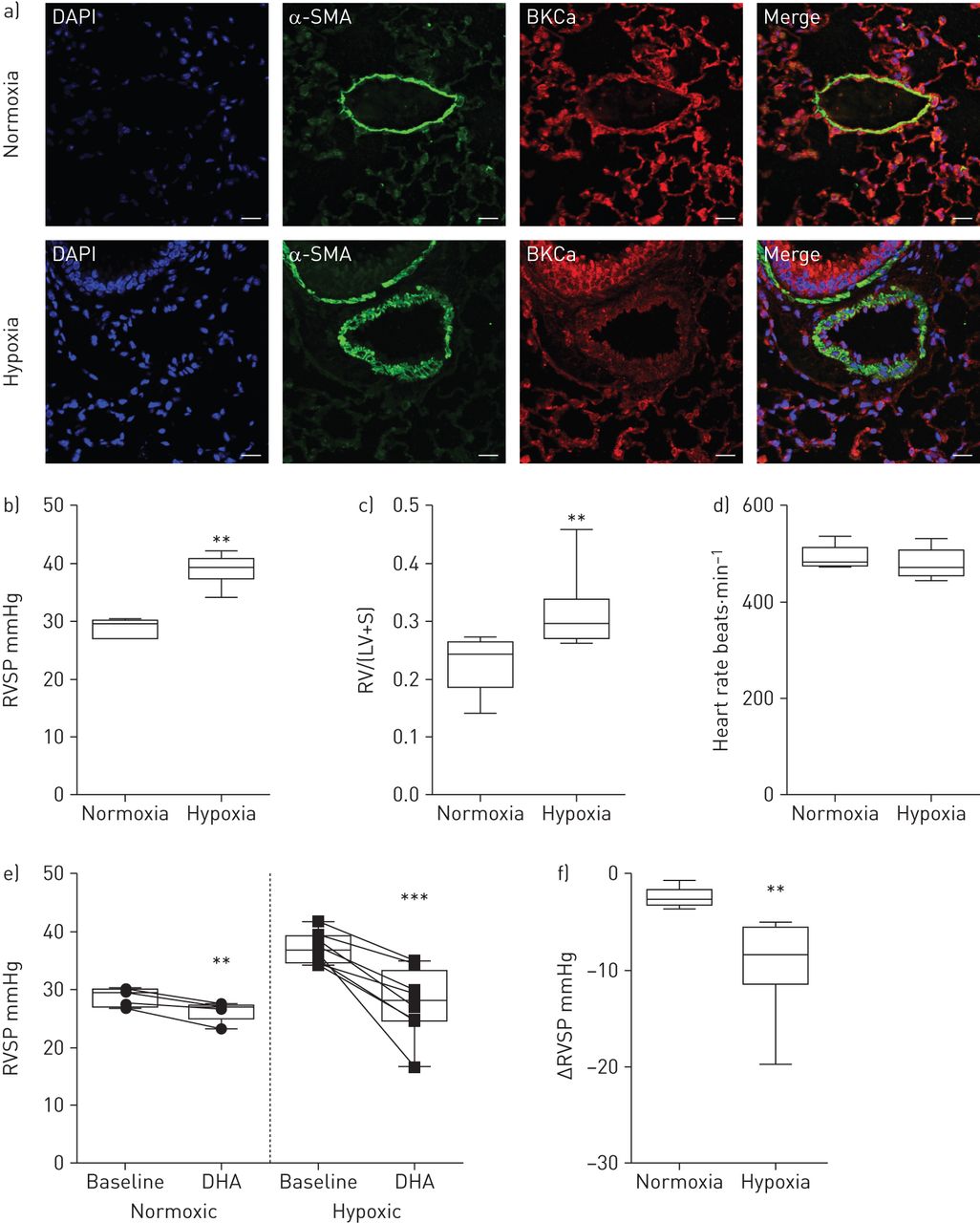
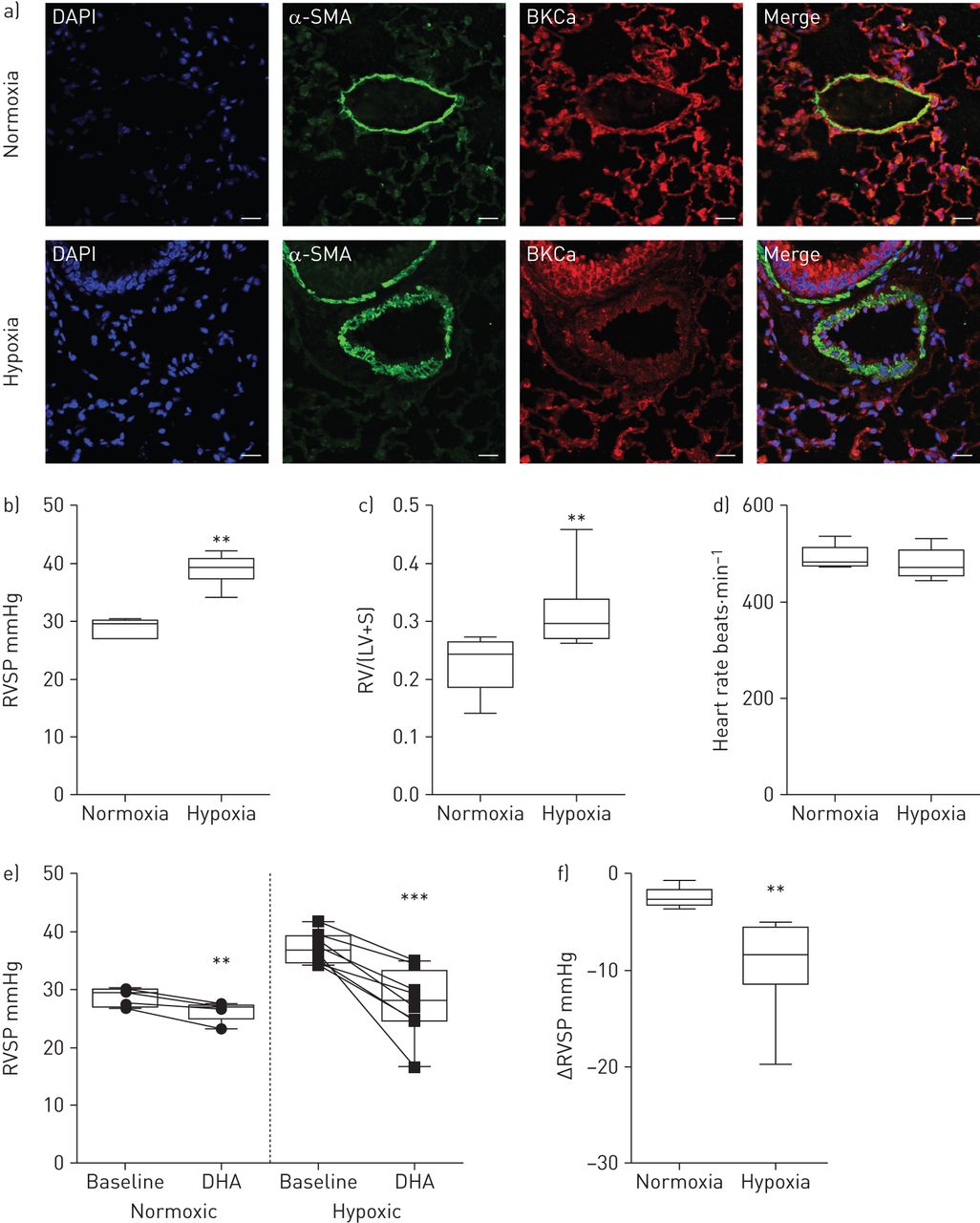
为了深入了解急性DHA应用是否在模拟人类肺血管重塑疾病的动物模型中表现出血管舒张作用,我们使用了慢性缺氧的肺动脉高压小鼠模型。通过对α-平滑肌肌动蛋白(α-SMA)阳性细胞和对照(常氧)动物和缺氧处理小鼠重构肺动脉中的BKCa通道的双重免疫标记,证实了肺动脉中存在BKCa通道(图4一).用特异性阻断肽和肺组织染色检测BKCa的抗体特异性+/+或BKCa- / -小鼠(在线补充图S3A和B)。为确认动物出现肺动脉高压,通过右心导管测量RVSP (图4 b),而两组之间的心率并无差异(图4 d).随后测量右心室肥厚,将右心室与左心室和室间隔完全分离。将分离的心室称量,以获得右心室与左心室加室间隔的比值(图4 c).在麻醉小鼠的中央静脉注射一剂DHA的效果是通过右心导管检测的。图4 e显示在常氧组和低氧组中,DHA应用后RVSP显著降低。然而,DHA在低氧组中比在常氧组中有更明显的作用(−9.4±1.7)与−2.5±0.4 mmHg, n=8与5;图4 f).在DHA应用于缺氧处理的小鼠中,RVSP从37±0.9下降到27.2±2 mmHg (n=8),使慢性疾病模型中的RVSP恢复到常氧小鼠的水平。因此,我们的结果表明dha诱导的血管舒张是通过激活肺动脉高压模型中的KCa通道发生的。
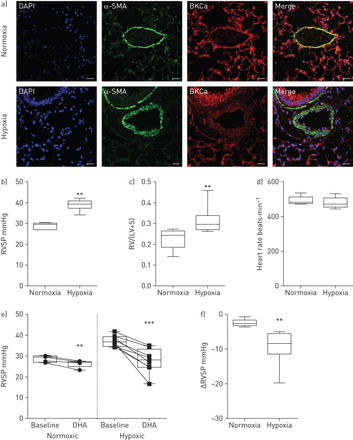
二十二碳六烯酸(DHA)在小鼠肺动脉高压模型中诱导增强的肺血管扩张。a)常氧和慢性缺氧小鼠肺双免疫荧光染色显示存在大导KCa (BKCa)通道(红色)和α-平滑肌肌动蛋白(α-SMA;绿色)在肺动脉。DAPI: 4 ', 6-diamidino-2-phenylindole。比例尺:20µM。b)右心室收缩压(RVSP), c)右心室肥厚(右心室(RV)从左心室(LV)和室间隔(S)完全剥离)和d)常氧和低氧小鼠的心率。e)在常氧和慢性缺氧治疗小鼠单丸DHA给药前(基线)和后的RVSP。f)总结常氧和缺氧小鼠DHA作用下RVSP的变化。数据以中值、四分位范围、最小值和最大值表示。常氧和缺氧时N =5和8。 **: p<0.01; ***: p<0.001.
DHA导致IPAH患者的pasmc超极化
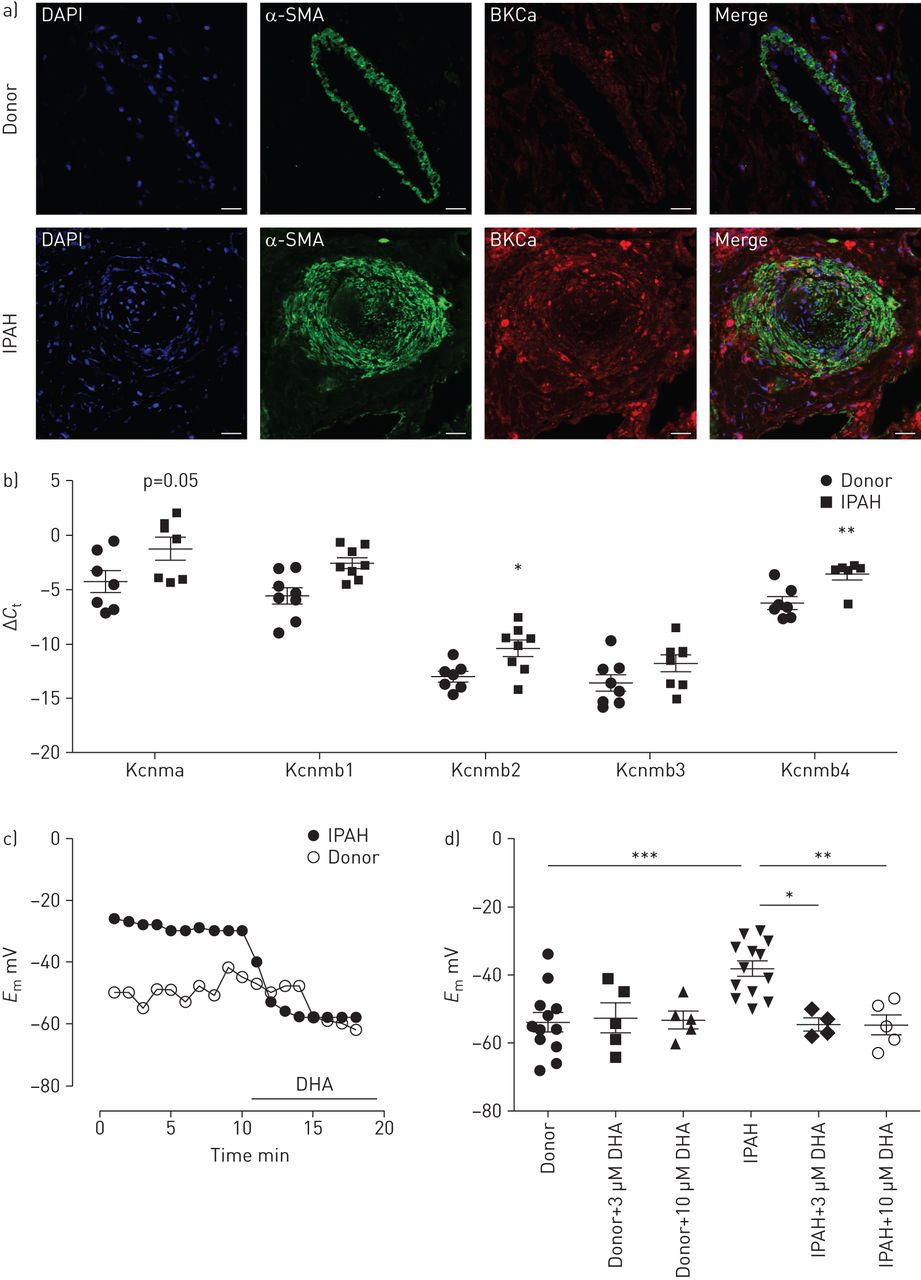
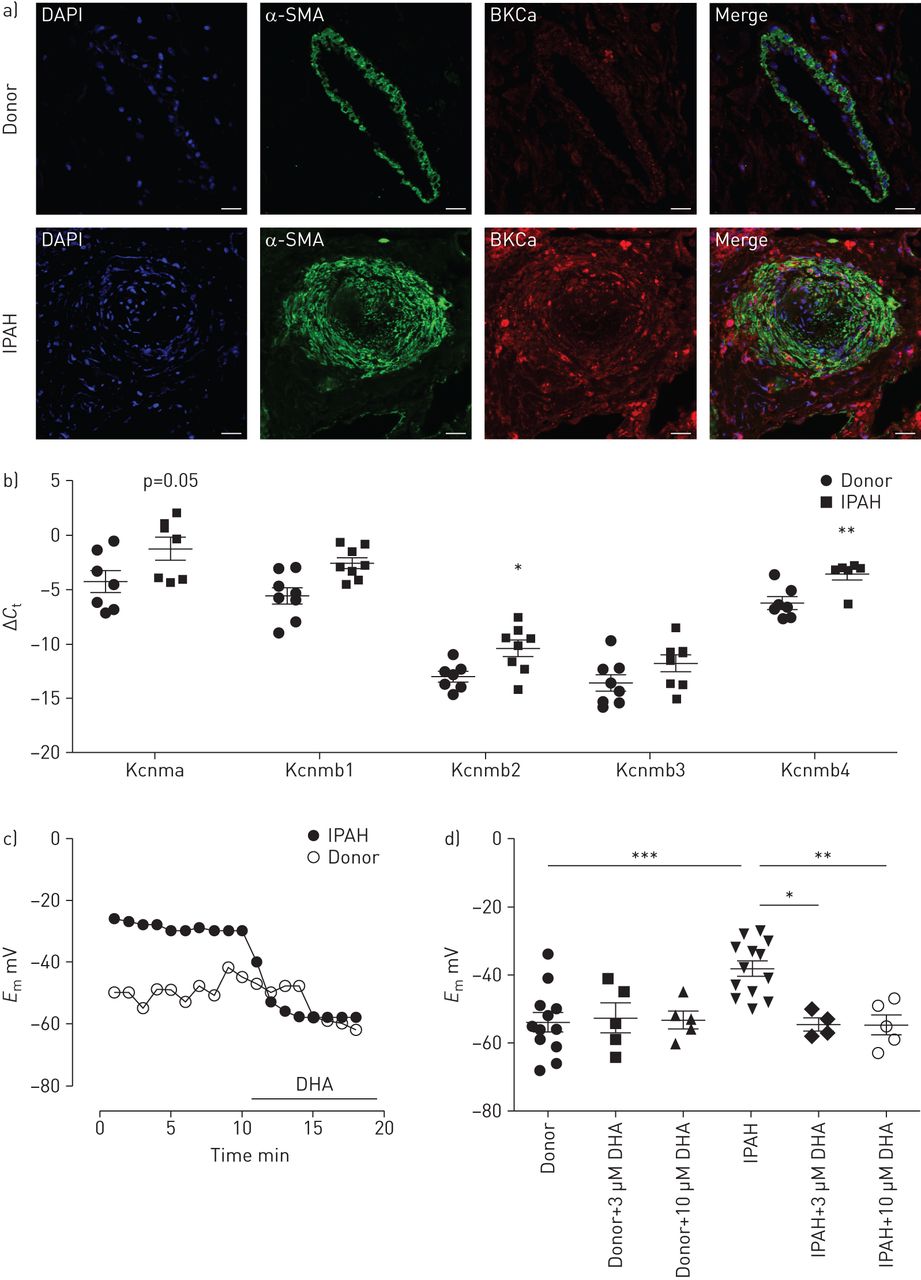
从DHA在动物模型中的有益作用出发,接下来我们研究了DHA对从IPAH患者和健康捐赠者获得的原发性人类pasmc的影响。双免疫标记显示来自健康供体肺的肺动脉平滑肌层和IPAH患者重塑肺动脉中的BKCa通道(图5一个).
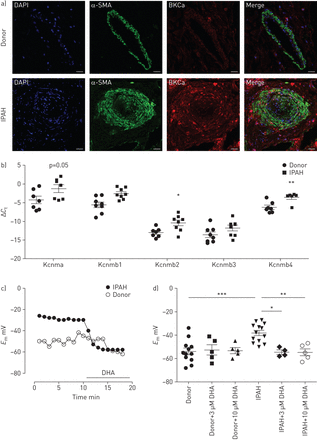
二十二碳六烯酸(DHA)可引起特发性肺动脉高压(IPAH)患者肺动脉平滑肌细胞(PASMCs)的过度极化。a)双免疫荧光染色显示大电导KCa (BKCa)通道(红色)和α-平滑肌肌动蛋白(α-SMA;绿色)在供体和IPAH人类肺的肺动脉中。DAPI: 4 ', 6-diamidino-2-phenylindole。比例尺:50µM。b)供体(n=7或8例)和IPAH (n=6或8例)患者激光显微解剖肺动脉中含α-亚基(Kcnma)及其β-亚基(Kcnmb1-4)的BKCa通道表达。c)代表性膜电位(E米)在急性DHA暴露时从供者和IPAH pasmc进行追踪。d)总结了供体和IPAH pasmc中DHA浓度增加和不含DHA时膜电位的变化。b、d)数据以均数±的单个点表示扫描电镜.*: p < 0.05;* *: p < 0.01;* * *: p < 0.001。
在健康供体和IPAH患者的激光微解剖肺动脉中研究BKCa通道的表达。IPAH患者的肺动脉中BKCa通道α-亚基(Kcnma1)以及β-亚基(Kcnmb1, Kcnmb2和Kcnmb4)明显上调(图5 b),而Kcnmb3则保持不变。然后,我们探索了DHA对从两组获得的原代人pasmc膜电位的影响。代表录音(图5 c)表明,急性灌注DHA使IPAH PASMCs的静息膜电位超极化,但不影响健康供体的PASMCs。值得注意的是,与健康供体的pasmc相比,IPAH患者的pasmc的膜电位明显去极化(−38.1±2.2)与−54±2.8 mV, n=12和13;图5 d).我们进一步的分析显示,急性DHA给药(3和10µM)没有改变对照供体pasmc的膜电位。相反,暴露于DHA可逆转IPAH患者的PASMCs去极化(3µM DHA后−54.5±1.8 mV)与−38.1±2.2 mV基线,n=4和−54.6±2.9 mV后10µM DHA与−38.1±2.2 mV基线,n=5;图5 d),使其恢复到健康捐赠者的水平。这些数据表明DHA对去极化的人pasmc产生选择性和直接的影响。
讨论
我们报道了DHA作为KCa通道激活剂在肺循环中的作用及其在肺动脉高压模型和IPAH样本中的急性作用。我们的研究表明,急性DHA管理迅速激活全细胞K+电流,表明这种激活电流主要来自KCa通道,而不是来自电压门控K+频道。KCa通道的激活是剂量依赖性的,降低了肺血管张力,并通过孤立的预狭窄的肺内动脉中的BKCa通道引起内皮依赖性和非依赖性肺血管舒张。内皮细胞的相对贡献与PASMC BKCa通道对DHA的血管舒张反应仍有待确定。此外,DHA显著降低U46619在离体灌注肺中升高的PAP;然而,它没有改变基线PAP。dha诱导的血管舒张对BKCa的影响显著降低- / -这表明DHA的急性肺血管舒张作用部分是通过激活BKCa通道介导的。重要的是,在小鼠慢性缺氧肺动脉高压疾病模型中在活的有机体内,急性DHA应用使右心室收缩压恢复到对照动物的水平,这表明DHA是肺动脉高压的有效血管扩张剂。此外,我们观察到IPAH患者重构肺动脉中BKCa通道的上调。与这些发现一致,我们还证明DHA使IPAH患者的原发性人类pasmc的膜电位超极化,在基线时被发现是去极化的,但健康捐赠者的情况则不是这样。
肺动脉的血管张力由pasmc控制;在生理(放松)条件下,它们具有相对稳定的膜电位,主要由K驱动+渗透性,尽管氯化物电导不应被忽视[5,22].因此,膜电位是血管张力的重要控制者:膜电位/血管张力斜率非常陡。因此,即使是几毫伏的膜电位变化也会引起血管直径的显著变化。任何改变膜电位的生理或药理因素都会引起压力的显著变化。因此,很明显,膜超极化,通过激活K+通道,是一个强大的机制,以降低压力通过血管扩张。KCa通道是由形成孔隙的α-亚基(Kcnmb1)和辅助β-亚基(Kcnmb1 - 4)构建的,在几乎所有类型的平滑肌细胞中都有发现,当激活时,会引起令人印象深刻的超极化和血管扩张[23,24].除了细胞内钙增加2 +而KCa激活的最有力的机制是由cAMP-或cgmp依赖的蛋白激酶(蛋白激酶a或G)介导的通道磷酸化[12,25,26].这对于KCa的生理功能非常重要,并为对抗肺动脉高压提供了强大的药理学工具,因为前列腺素、磷酸二酯酶-5抑制剂或可溶性胍酰环化酶激活剂在一定程度上发挥了其血管扩张作用通过激活KCa。
BKCa通道表达和功能在慢性缺氧小鼠和pasmc中升高[27,28].与肺动脉高压模型一致的是,我们首次证明了通过激光捕获IPAH患者的显微解剖获得的重塑肺动脉中BKCa通道和β亚基上调。因此,人类PASMCs中的BKCa通道可能在慢性血管收缩的背景下为DHA提供一个靶点,并可能作为一个反馈途径来控制膜去极化和血管收缩的程度,如模型所示(在线补充图S4)。
DHA是ω-3脂肪酸的主要成分之一,主要通过食用海洋食品获得。流行病学研究和临床试验表明,DHA对人类健康有益,因此美国心脏协会建议人们定期从鱼类中摄入DHA,或将其作为预防心血管疾病的膳食补充剂[14,29].DHA及其代谢衍生物对肺循环的慢性影响已在PASMCs和增殖表型上进行了研究在体外研究证明DHA能够抵消增强的人类PASMC增殖,这是肺动脉重塑的标志[30.].长期DHA治疗抑制细胞外信号调控的激酶激活,并引发未折叠蛋白反应,引起周期阻滞和细胞凋亡诱导[17].此外,一项早期研究表明,DHA结合并刺激g蛋白偶联受体GPR120与EC50的~ 10 μM [31],表明DHA的免疫调节作用可能存在新机制。此外,一项专注于DHA诱导平滑肌变化的研究提出,DHA通过分解成其代谢产物,可能抑制Rho激酶途径,降低钙敏感性[32].
相比之下,DHA的急性作用最近才被研究出来。在肺循环中,研究了DHA在急性缺氧环境下对BKCa通道的缺氧抑制,其中DHA的使用逆转了BKCa通道的缺氧抑制[33].此外,DHA暴露引起基础张力迅速下降,从而导致体循环血管舒张,这表明DHA对K有直接影响+导致平滑肌细胞膜超极化,从而导致血管舒张的通道[21].事实上,Hoshi等.[34]为DHA直接结合到人胚胎肾细胞中表达的大电导KCa的成孔Slo1和辅助β1亚基提供了证据。我们的观察结果与该研究一致,表明DHA直接作用于膜片钳测量的KCa通道活性,因为当KCa通道调节剂被抑制时,人类pasmc中KCa通道的激活保持不变(在线补充图S2A)。与之前在体循环中的发现相反,在我们的研究中,DHA不影响孤立灌注小鼠肺的基础肺动脉张力,在常氧状态下的小鼠中,DHA引起的急性RVSP降低很小。但是,在肺动脉预收缩后体外模型以及在在活的有机体内结果表明,DHA可显著降低肺压,提示DHA对收缩的肺血管有选择性作用。在BKCa敲除小鼠预收缩的孤立灌注肺中,DHA继续引起一些血管舒张,这表明它具有额外的血管舒张特性。因此,DHA之前被报道为一种强大的体循环血管扩张剂,但未能改变BKCa敲除小鼠的全身血压,这一事实表明,DHA的作用强烈依赖于两种血管的特定细胞环境和基础张力水平[21].最后,最近有报道称,含ω-3多不饱和脂肪酸的脂质输注可诱导慢性肺高压胎羊模型的有效和持续的血管扩张[35].目前的研究与此一致,因此能够进一步证明当PAP增加时,DHA作为急性血管扩张剂的相关性。
使用体外而且在活的有机体内肺动脉高压动物模型,结合激光显微解剖重塑肺动脉的研究,以及对IPAH患者分离的pasmc的膜片钳研究,我们进行了详细的实验表征,并对dha诱导的肺动脉高压血管舒张提供了分子上的见解。据我们所知,这是第一个表明DHA能迅速降低肺血管张力和压力的研究。这种减少是通过激活PASMCs中的KCa通道来实现的通过直接调节膜电位导致血管松弛。我们还证明DHA激活KCa通道对已建立的肺动脉高压模型是有益的,更重要的是,DHA只使IPAH患者获得的pasmc高极化,而不是对照的pasmc。此外,我们在人IPAH中检测到BKCa通道的增加,这表明KCa开启器在PAH的药理干预中具有实用性。因此,我们提出DHA是一种有效的血管扩张剂,通过激活KCa通道治疗肺动脉高压。IPAH患者中KCa通道表达的增加可能进一步引起人们对DHA作为肺动脉高压潜在治疗选择的兴趣。
确认
我们非常感谢Maria Helene Schloffer, Simone Tischler和Elisabeth Blanz(奥地利格拉茨医科大学麻醉和重症监护医学系实验麻醉学)出色的技术支持。
脚注
这篇文章有补充资料可从www.qdcxjkg.com
支持声明:A.L. Meredith由国家心肺血液研究所(资助号:HL102758)支持。本文的资助信息已存入FundRef.
利益冲突:可以在本文旁边的网站上找到信息披露www.qdcxjkg.com
- 收到了2015年11月3日。
- 接受2016年5月14日。
- 版权所有©ERS 2016











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}