





数据
-
图1
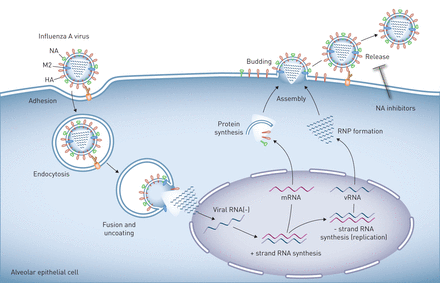
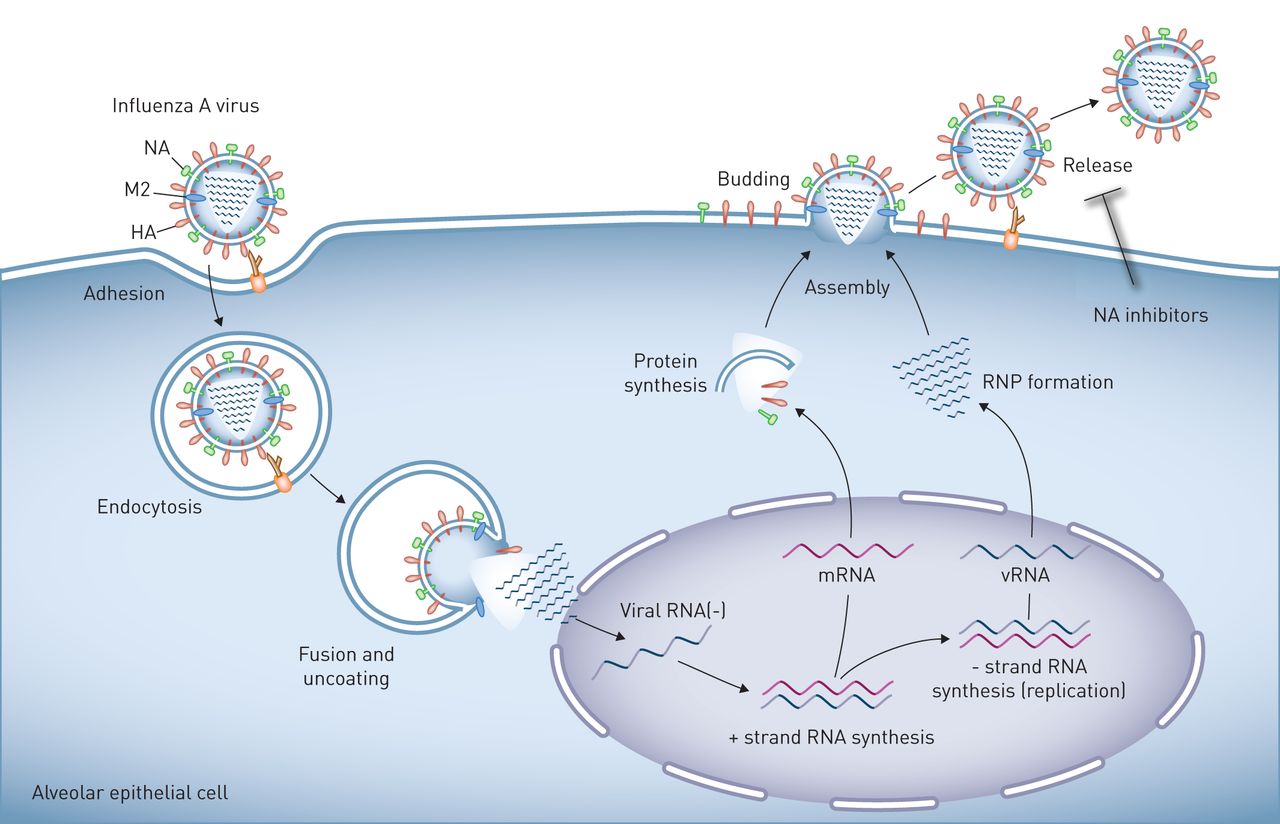
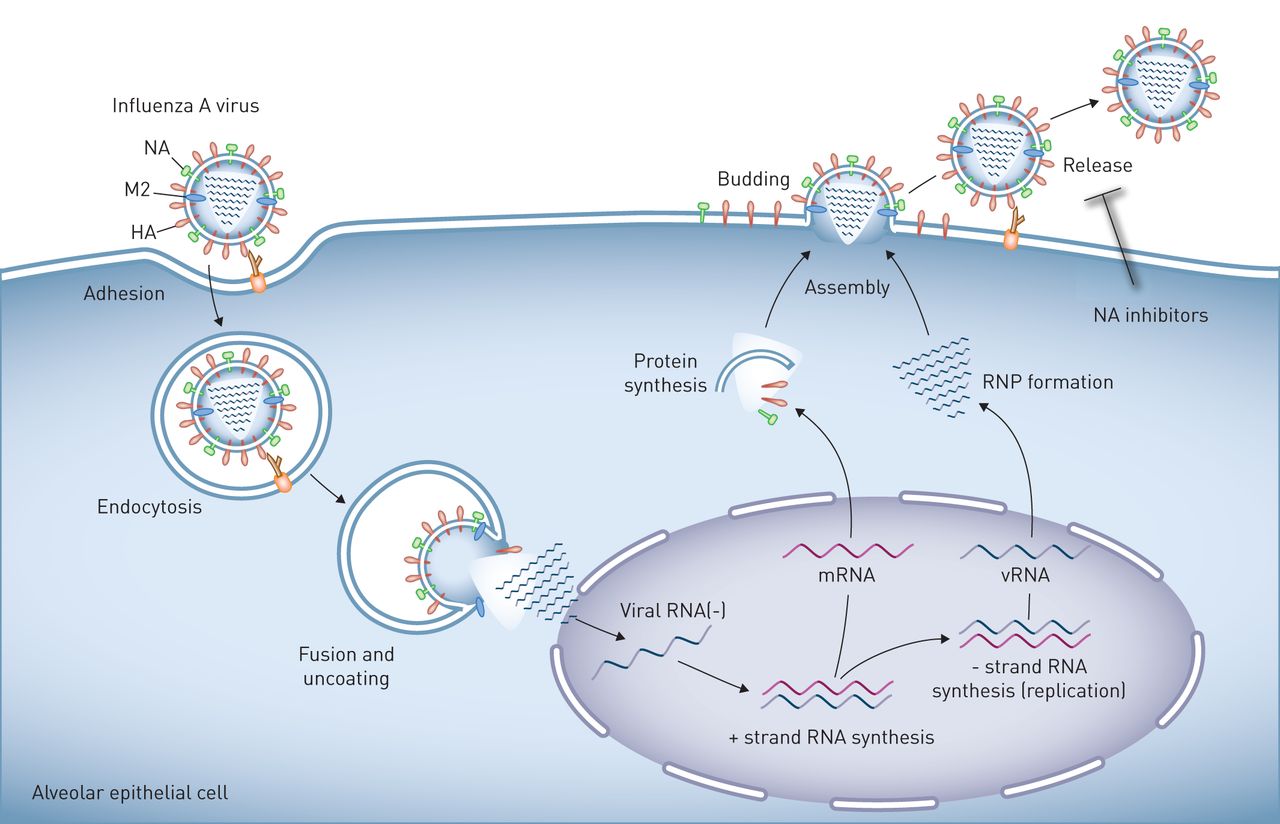
A型流感病毒在肺上皮细胞的复制在流感病毒粒子表面表达的血凝素(HA)与连接到细胞表面聚糖的唾液酸残基结合,可诱导病毒粒子与靶细胞的质膜结合和融合。人病毒中的HA与与表面聚糖相连的唾液酸残基相互作用通过α-2,6连锁,在人气道上、下上皮细胞和肺泡II型细胞中发现。相反,禽病毒中的HA与唾液酸残基以α-2,3连接相互作用。然后病毒进入细胞通过内吞作用或小胞饮作用,并被转运到溶酶体,在那里酸化激活质子选择性基质蛋白-2病毒通道(M2),诱导膜融合和病毒核糖核蛋白(RNP)核心的解离,然后转运到发生病毒RNA复制的细胞核。子代病毒RNP核心在细胞质中产生,与病毒表面蛋白、HA和神经氨酸酶(NA)以及其他病毒蛋白一起集中在质膜的脂筏中或附近。这些质膜区域的出芽形成完整的病毒子代,通过HA/唾液酸相互作用与质膜相连。神经氨酸酶对唾液酸残基的裂解会释放病毒子代,这样它们就可以自由地感染其他细胞,这可以通过NA抑制剂来阻止。以肺泡上皮细胞为例,但其生命周期在气道上皮细胞中相似。
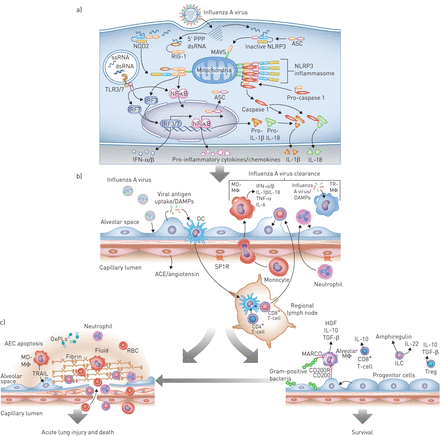
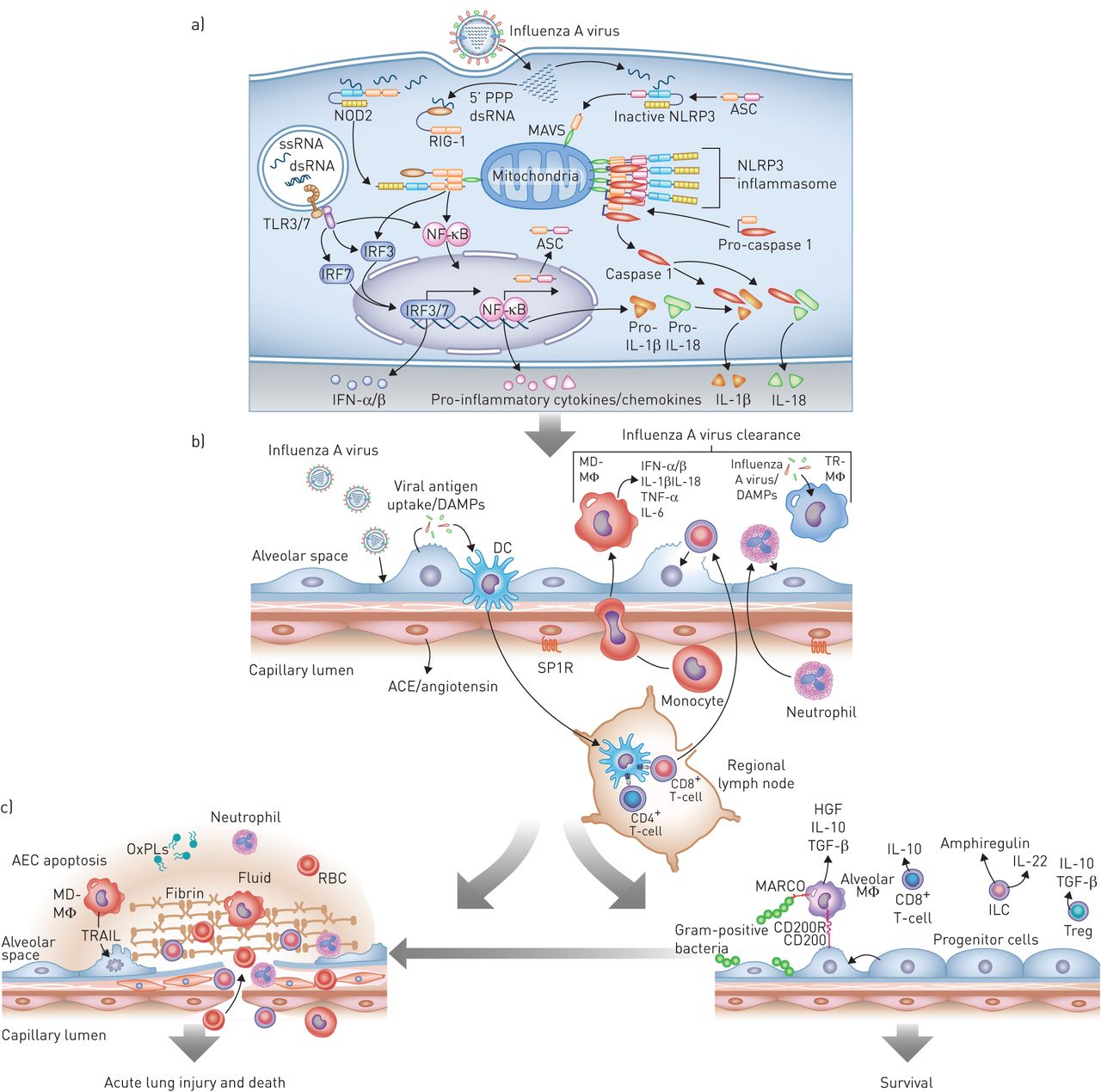
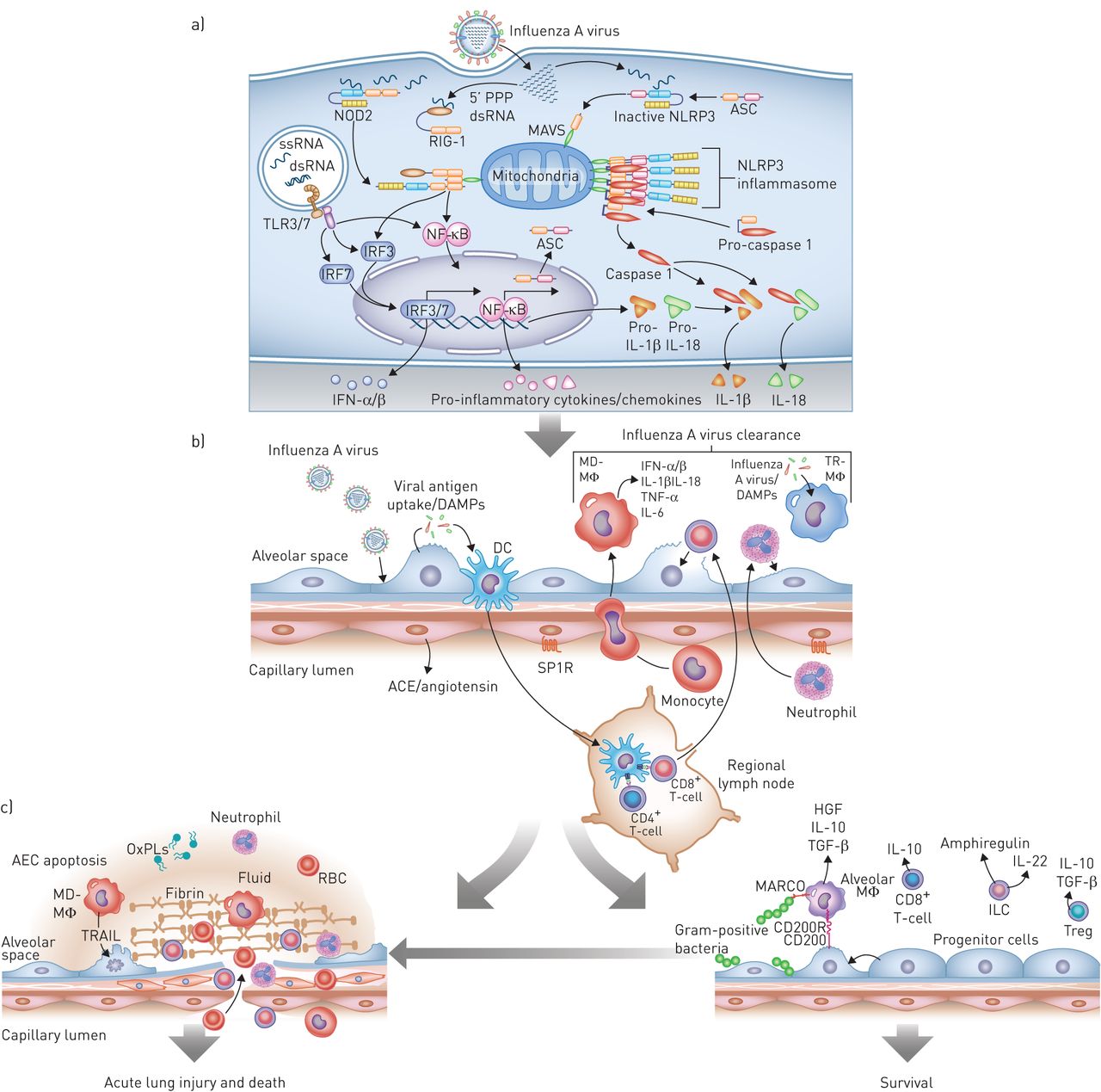
流感感染导致肺中有益和有害的宿主免疫通路的连续激活。a)最早的反应见于感染的气道或肺泡上皮细胞(AEC)。从左到右,细胞内病毒RNA的存在激活toll样受体(TLRs),主要是TLR7和TLR3,以诱导干扰素调节因子(IRF)3或IRF7的激活途径,从而增加I型干扰素(IFN)-α/β的转录。该通路的激活还可以通过激活核因子(NF)-κB诱导促炎细胞因子和趋化因子的转录。流感感染期间释放到细胞质中的5 '三磷酸化双链RNA (5 ' -PPP dsRNA)诱导维甲酸诱导基因i (RIG-I)的构象变化,该基因i与线粒体抗病毒信号蛋白(MAVS)相互作用,使其激活核苷酸结合寡聚结构域含蛋白(NOD)2。这也诱导I型ifn的转录通过IRF3和促炎细胞因子通过NF -κB。NF-κB诱导炎性小体蛋白ASC(包含CARD的适配蛋白凋亡相关斑点样蛋白)、IL -1β和IL-18的表达。在病毒RNA存在的情况下,ASC与NLR家族、含pyrin结构域(NLRP)3和MAVS相互作用,诱导NLRP3炎性小体的激活,NLRP3炎性小体裂解并激活caspase-1生成IL-1β和IL-18,从而放大炎症级联反应。b)气道或肺泡II型细胞感染导致损伤相关分子模式分子(DAMPs)和病原体相关分子模式分子(PAMPs)的释放,常驻树突状细胞(DCs)可以感知这些分子模式分子。DCs迁移到区域淋巴结激活细胞毒性(CD8+)和辅助细胞(CD4+) t细胞,以及罕见的记忆t细胞能够诱导特定的抗病毒反应(未显示)。与此同时,DAMPs, PAMPs,内吞病毒,可能还有流感感染本身会诱导组织内的肺泡巨噬细胞(TR-MΦ)和dc(未显示)释放I型ifn和炎症细胞因子。这些细胞因子/趋化因子诱导中性粒细胞的募集以及外周血单核细胞募集和分化为单核细胞来源的肺泡巨噬细胞(MD-MΦ)。中性粒细胞和MD-MΦ都能放大炎症反应。底层内皮的损伤导致炎症细胞募集和炎症的负调节因子的丢失,包括通过SP1R的信号传递和血管紧张素的释放通过血管紧张素转换酶(ACE),从而放大炎症反应。c) I型干扰素和炎症细胞因子的释放以及细胞毒性t细胞的作用对病毒至关重要
表
-
表1
A型流感病毒(IAV):一种节段反义RNA病毒,编码11种蛋白质
病毒蛋白 位置 函数 评论 新的抗病毒疗法在临床研究中得到验证 血凝素 病毒囊膜 附着在呼吸道和肺泡上皮上的唾液酸残基,膜融合和病毒进入 在人类气道中,蛋白水解裂解和疾病严重程度与宿主蛋白酶TMPRSS2和HAT有关[25-27]。n链糖基化的加入掩盖了血凝素作为抗原表位[28-31],但触发c型凝集素的识别[32-34]。 DAS181(唾液酸裂解),flufirvitide(与血凝素结合的肽抑制剂) Neuraminadase 病毒囊膜 新形成的病毒粒子末端唾液酸残基的酶解和子代的释放 目前批准的IAV感染治疗(osteltamivir, zanamivir, laninamivir和peramivir)靶向神经氨酸酶 矩阵protein-2 病毒囊膜 病毒进入、组装和出芽所需的质子选择性离子通道 基质蛋白-2被认为通过抑制自噬[35]促进细胞凋亡,并诱导ENaC和CFTR的降解[36,37],从而影响肺泡液的清除 AVI-7100(干扰M段基因表达) 矩阵蛋白1 病毒矩阵 膜和核糖核蛋白核心之间的结构联系 聚合酶蛋白:PB1、PB2、PA 核糖核蛋白核心 RNA聚合酶复合蛋白 聚合酶亚基与病毒致病性有关,并与>300细胞蛋白相互作用[38]突变可以改变复制效率;如。单一氨基酸替代(E627K)在1918年、1957年和1968年流行的IAV毒株中诱导了更高的致病性[39,40],并在流行的H5N1和H7N9变种中发现 Favipiravir(靶向PB1的核苷抑制剂) 核衣壳蛋白 核糖核蛋白核心 病毒基因组的包装 非结构蛋白1 在复制过程中表达,不是成熟病毒粒子的一部分 免疫逃避 它与病毒RNA、RIG-1和TRIM25复合物结合,抑制下游抗病毒信号通路[41,42]。它还诱导IFN-α受体[43]的下调和JAK/STAT信号通路抑制剂(SOCS1和SOCS3)的上调[43,44]。在感染期间,非结构蛋白-1也通过蛋白激酶R阻断促凋亡信号通路[45,46],并阻止NF-κB[47]的激活 非结构化protein-2 在复制过程中表达,不是成熟病毒粒子的一部分 病毒复制过程中从细胞核输出的病毒核糖核蛋白 PB1-F2# 在复制期间表示 诱导宿主细胞凋亡 PB1-F2通过与线粒体膜蛋白ANT3和VDAC相互作用提高线粒体膜通透性,增强BAK/ bax介导的细胞色素C释放[48-50] 成熟的病毒粒子包括8种结构蛋白;其他三种在病毒复制过程中表达。血凝素和神经氨酸酶分别是病毒结合和释放所必需的,存在于病毒包膜中。血凝素和神经氨酸酶共同决定了病毒的抗原特性,并被用来定义不同的病毒株,如。导致2009年流感大流行的H1N1毒株,以及导致最近亚洲爆发禽流感的H7N9毒株。一些病毒蛋白可能是抗病毒治疗的新靶点。TMPRSS2:跨膜蛋白酶丝氨酸2;人气道胰样蛋白酶;ENaC:上皮钠通道;CFTR:囊性纤维化跨膜传导调节剂;RIG-1:视黄酸诱导基因-1;干扰素:干扰素;JAK: janus激酶;STAT:信号转换器和转录激活器; NF-κB: nuclear factor-κB; VDAC: voltage-dependent anion channels.#:所有病毒不表达。











{kind=link}
{kind=link}
{kind=link}
{kind=link}