





抽象的gydF4y2Ba
凝血调节蛋白(TM)的凝集素样结构域在无菌炎症条件中起重要的调节作用,但其在严重革兰氏阳性感染性疾病中的作用尚不清楚。gydF4y2Ba肺炎链球菌肺炎料gydF4y2Ba是社区获得的肺炎最常见的原因。本研究的目的是确定凝集素状的TM在鼠肺炎骨球菌肺炎的角色的作用。gydF4y2Ba
野生型(WT)小鼠和缺乏TM凝集素样结构域(TMgydF4y2BaLED / LEDgydF4y2Ba用可行的鼻内感染gydF4y2BaS.肺炎gydF4y2Ba并且在感染后在生存研究中观察或安乐死6,24或48小时。gydF4y2Ba
TM值gydF4y2BaLED / LEDgydF4y2Ba与野生型小鼠相比,小鼠在肺炎球菌性肺炎中有明显更好的生存。感染后48小时gydF4y2Ba肺炎链球菌,gydF4y2BaTM值gydF4y2BaLED / LEDgydF4y2Ba小鼠血液和肝脏中的细菌负荷较低,表现出较少的肺部炎症,表现为肺组织病理学较低,中性粒细胞流入较少,细胞因子和趋化因子水平较低。TM患者血浆促炎细胞因子水平也降低gydF4y2BaLED / LEDgydF4y2Ba小鼠暴露于感染后。gydF4y2Ba
缺失TM的凝集素样域改善了肺炎球菌肺炎的宿主防御。TM的凝集素状结构域可以响应于革兰氏阳性或革兰氏阴性细菌而具有差异作用。gydF4y2Ba
社区获得性肺炎的主要原因是gydF4y2Ba肺炎链球菌肺炎料gydF4y2Ba[gydF4y2Ba1gydF4y2Ba].每年,在美国发生〜570 000例肺炎骨膜肺炎病例,占~175 000住院治疗[gydF4y2Ba2gydF4y2Ba].当与败血症相关时,肺炎球菌性肺炎的死亡率可超过20% [gydF4y2Ba3.gydF4y2Ba].在世界范围内,gydF4y2BaS.肺炎gydF4y2Ba负责每年估计的1000万人死亡[gydF4y2Ba4gydF4y2Ba].与对该病原体的抗生素抗性的发病率升高相同[gydF4y2Ba1gydF4y2Ba],现在迫切需要扩大我们对影响肺炎球菌肺炎和败血症结果的宿主防御机制的知识。gydF4y2Ba
血栓调节蛋白(TM) CD141是一种跨膜、多结构域糖蛋白受体,主要表达于血管内皮细胞,但也表达于单核细胞、中性粒细胞、成骨细胞、滑膜细胞和树突状细胞[gydF4y2Ba5gydF4y2Ba- - - - - -gydF4y2Ba7gydF4y2Ba].TM在很大程度上凝结,纤维蛋白溶解,先天免疫和炎症的凝固,纤维蛋白溶解,先天免疫和炎症的几种不同的性质。TM的中心结构域由六个表皮生长因子(EGF) - 样重复组成,其中三种,其中三个是用于凝血酶介导的活化蛋白C(APC)和活性凝血酶激活纤维蛋白解抑制剂(TAFIA)的关键辅因子活性.蛋白质C通过凝血酶-TM激活,进一步通过内皮蛋白C受体增强[gydF4y2Ba5gydF4y2Ba- - - - - -gydF4y2Ba7gydF4y2Ba].APC通过裂解和灭活凝血辅助因子Va和viia来抑制凝血酶的进一步生成。APC还具有深刻的消炎特性[gydF4y2Ba8gydF4y2Ba- - - - - -gydF4y2Ba10gydF4y2Ba].血浆羧肽酶B,Tafia通过改性纤维蛋白原来抑制纤维蛋白溶解,使抗纤溶酶原料的纤维蛋白抗性,并限制纤溶酶产生[gydF4y2Ba11gydF4y2Ba].TAFIa还灭活补体片段和过敏毒素C3a和C5a,从而抑制先天免疫反应[gydF4y2Ba12gydF4y2Ba].除了在激活蛋白C和TAFI方面的关键作用gydF4y2Ba8gydF4y2Ba,gydF4y2Ba11gydF4y2Ba, TM也有直接的抗炎特性[gydF4y2Ba5gydF4y2Ba- - - - - -gydF4y2Ba7gydF4y2Ba至少部分是由其n端c型凝集素样结构域介导的。这种结构干扰中性粒细胞粘附、补体激活和细胞因子的产生[gydF4y2Ba13gydF4y2Ba- - - - - -gydF4y2Ba15gydF4y2Ba[缺少TM的凝集素样域的小鼠(TMgydF4y2BaLED / LEDgydF4y2Ba小鼠)在内毒素血症、肺和心肌缺血-再灌注和炎症性关节炎模型中对组织损伤的敏感性增加[gydF4y2Ba13gydF4y2Ba- - - - - -gydF4y2Ba15gydF4y2Ba].gydF4y2Ba
虽然已经对肺炎和败血症中凝血系统调节的改变进行了广泛的研究[gydF4y2Ba8gydF4y2Ba],目前对TM在宿主对细菌感染反应中的作用了解有限。由于TM在肺中高表达,特别是肺泡毛细血管内皮细胞[gydF4y2Ba16.gydF4y2Ba,gydF4y2Ba17.gydF4y2Ba],我们一直在评估其对呼吸道感染的反应的参与。具有编码TM基因中具有点突变的小鼠,其显着降低了其产生APC的能力,降低了TM抗原水平,令人惊讶地没有表现出对内毒素的肺反应或呼吸道病原体的改变,包括gydF4y2BaS.肺炎gydF4y2Ba[gydF4y2Ba18.gydF4y2Ba].使用TM.gydF4y2BaLED / LEDgydF4y2Ba小鼠及其对手野生型(WT)小鼠,我们寻求确定gydF4y2Ba体内gydF4y2BaTM的凝集素样结构域在应对肺炎球菌肺炎中的作用。gydF4y2Ba
材料和方法gydF4y2Ba
动物gydF4y2Ba
TM值gydF4y2BaLED / LEDgydF4y2Ba如上所述产生小鼠[gydF4y2Ba13gydF4y2Ba],回交8次获得C57BL/6遗传背景。TM值gydF4y2BaLED / LEDgydF4y2Ba小鼠表达正常的Tm抗原水平,并激活蛋白C的完整性,不受粘合剂状结构域的缺失的影响[gydF4y2Ba13gydF4y2Ba].C57BL/6 WT小鼠购自Charles River (Maastricht, Netherlands)。所有的实验都是由阿姆斯特丹大学(Amsterdam, Netherlands)动物保护和使用委员会批准的。gydF4y2Ba
实验感染和取样gydF4y2Ba
肺炎患者肿瘤接种诱导〜5×10gydF4y2Ba4gydF4y2BaCFUgydF4y2BaS.肺炎gydF4y2Ba血清型3(ATCC 6303;美国型文化收藏,Rockville,MD,USA)如上所述[gydF4y2Ba18.gydF4y2Ba,gydF4y2Ba19.gydF4y2Ba].观察1周(每组n=14只),然后在预定的时间点处死或安乐死(每组每个时间点n=8只)。从下腔静脉采血制备柠檬酸血浆。器官匀浆按所述制备[gydF4y2Ba18.gydF4y2Ba,gydF4y2Ba19.gydF4y2Ba].简而言之,收集左肺和大约一半的脾脏和肝脏,使用组织匀浆器(Biospect Products, Bartlesville, UK)在4°C下用四卷无菌盐水匀浆。器官匀浆中的细菌生长如所述[gydF4y2Ba18.gydF4y2Ba,gydF4y2Ba19.gydF4y2Ba].对于肺组织的进一步测量,用裂解缓冲液(300mM NaCl,30mM Tris,2mM MgCl稀释1:2稀释肺匀浆gydF4y2Ba2gydF4y2Ba, 2 mM CaClgydF4y2Ba2gydF4y2Ba, 1%(体积/体积)Triton X-100, pH 7.4),添加蛋白酶抑制剂混合物(4-(2-氨基乙基)苯磺酰氟(AEBSF)), EDTA-NAgydF4y2Ba2gydF4y2Ba,胃蛋白酶和leapeptin,所有购自MP生物医学(荷兰埃因霍温),并在冰上孵育30分钟,然后在680×以680×离心gydF4y2BaggydF4y2Ba10分钟。将上清液储存在-20℃直至分析。gydF4y2Ba
在单独的一系列实验中,暴露气管,用无菌22号abbocat导管(Abott, Sligo, Ireland)插管,然后通过灌注和取出两个0.5 mL等量的生理盐水(每组每个时间点n=8)进行支气管肺泡灌洗(BAL)。用血球计(Beckman Coulter, Fullerton, CA, USA)测定每个BAL液(BALF)样本的细胞计数。Giemsa染色(diffa - quick;Dade Behring AG, Düdingen,瑞士)。gydF4y2Ba
组织学和免疫组织化学gydF4y2Ba
右肺在10%福尔马林/PBS中室温固定24 h,随后石蜡包埋;切片厚度为5 μm。使用异硫氰酸荧光素标记的抗小鼠ly - 6g单克隆抗体(Pharmingen, San Diego, CA, USA)进行粒细胞染色,如所述[gydF4y2Ba20.gydF4y2Ba].Ly-6G染色玻片用配备数码相机(徕卡CTR500;徕卡微系统公司,Wetzlar,德国)。每张幻灯片随机拍摄10张照片。染色区域使用Image Pro Plus (Media Cybernetics, Bethesda, MD, USA)进行分析,并以总表面积的百分比表示。切片也用苏木精和伊红染色,并由不了解不同组身份的病理学家进行分析。为了对肺部炎症和损害进行评分,对整个肺切片进行以下参数分析:支气管炎、间质炎症、水肿、内皮炎、胸膜炎和血栓形成。每个参数按0 - 4分等级(0:缺席;1:温和;2:温和; 3: severe; and 4: very severe). The total histopathological score was expressed as the sum of the scores for the different parameters.
化验gydF4y2Ba
ELISA测量了MyeloceRoxidase(MPO)(Hycort Biotechnology,荷兰,荷兰)。肿瘤坏死因子(TNF)-α,白细胞介素(IL)-6,单核细胞化学侵入剂蛋白(MCP)-1,IL-12P70,细胞率珠阵列多重测定测量干扰素(IFN)-γ和IL-10(BD Biosciences,圣何塞,加州,美国)。通过ELISA(R&D Systems,Minneapolis,Mn,USA)测量角质形成细胞衍生的趋化因子(KC)和巨噬细胞炎症蛋白(MIP)-2。通过ELISA(西门子医疗保健诊断,德国Marburg)测量凝血酶 - 抗凝血酶复合物(TATC)。gydF4y2Ba
统计分析gydF4y2Ba
数据可以用盒状和须状图来描述最小观测值、下四分位数、中位数、上四分位数和最大观测值,也可以用四分位数区间的中位数或Kaplan-Meier图来表示。组间差异采用Mann-Whitney u检验或log rank检验。采用GraphPad Prism 4.0版本(GraphPad Software, San Diego, CA, USA)进行分析。p值<0.05认为有统计学意义。gydF4y2Ba
结果gydF4y2Ba
TM值gydF4y2BaLED / LEDgydF4y2Ba小鼠在肺炎球菌感染后表现出存活的优势gydF4y2Ba
为了响应致命剂量,确定TM的凝集素样域是否改变生存gydF4y2BaS.肺炎gydF4y2Ba, TM值gydF4y2BaLED / LEDgydF4y2Ba和WT小鼠鼻内接种细菌并监测1周(gydF4y2Ba无花果。1gydF4y2Ba).TM值gydF4y2BaLED / LEDgydF4y2Ba与野生型小鼠相比,小鼠对致死感染有明显的保护作用(p<0.05),且开始死亡的时间较野生型小鼠晚。14个TM中的5个gydF4y2BaLED / LEDgydF4y2Ba小鼠在研究结束时活着,而只有14只WT小鼠中只有一个幸存下来。gydF4y2Ba
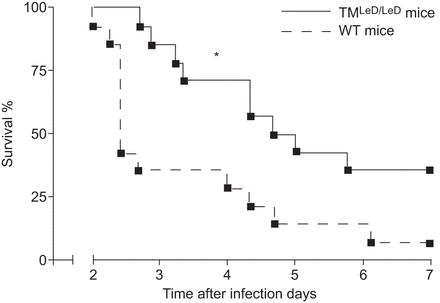
血栓调节蛋白的凝集素样结构域(TMgydF4y2BaLED / LEDgydF4y2Ba)降低了鼠肺炎球菌肺炎的死亡率。野生型(WT)小鼠的存活和缺乏TM的小鼠gydF4y2BaLED / LEDgydF4y2Ba经鼻内感染5×10gydF4y2Ba4gydF4y2Ba肺炎链球菌肺炎料gydF4y2BaCFU(每组14只小鼠)。*:P <0.05,统计显着性与WT使用日志等级测试相比。gydF4y2Ba
TM值gydF4y2BaLED / LEDgydF4y2Ba小鼠显示感染的传播减少gydF4y2Ba
以确定TM之间的生存差异gydF4y2BaLED / LEDgydF4y2Ba和WT小鼠可归因于抗菌防御的差异,我们测得肺部,血液和迟到的器官(脾脏和肝脏)的感染后细菌过度6,24和48小时(gydF4y2Ba无花果。2gydF4y2Ba).6小时,TMgydF4y2BaLED / LEDgydF4y2Ba小鼠肺内细菌载量略低于野生型小鼠(p<0.05) (gydF4y2Ba无花果。2AgydF4y2Ba).然而,在随后的时间点上,在不同基因型小鼠(gydF4y2Ba无花果。2AgydF4y2Ba).而在感染后的最初24小时,血液和远处器官的培养几乎都是阴性的(gydF4y2Ba图2罪犯gydF4y2Ba),在48小时,大多数小鼠显示系统性感染。值得注意的是,在这个时间点,TMgydF4y2BaLED / LEDgydF4y2Ba小鼠血液中的细菌计数显著降低(gydF4y2Ba无花果。2B.gydF4y2Ba)及肝脏(gydF4y2Ba无花果。2DgydF4y2Ba)与野生型小鼠比较(均p<0.05),而脾脏肺炎球菌载量(gydF4y2Ba无花果。2CgydF4y2Ba组之间没有显着差异。gydF4y2Ba
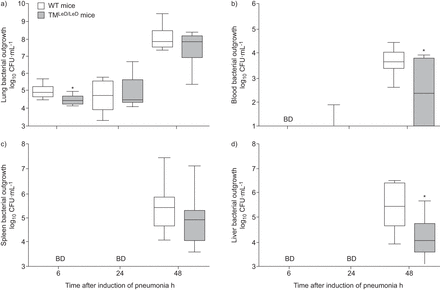
血液和肝脏中的细菌生长缺乏血栓调节蛋白的凝集素样结构蛋白(TMgydF4y2BaLED / LEDgydF4y2Ba与野生型小鼠相比,野生型小鼠感染肺炎球菌后的存活率明显降低。a)肺,b)血液,c)脾脏和d)肝6,24和48h诱导肺炎后血糖接种,5×10gydF4y2Ba4gydF4y2Ba肺炎链球菌肺炎料gydF4y2Bacfu在wt和tmgydF4y2BaLED / LEDgydF4y2Ba老鼠。数据表示为描绘最小观察,低四分位数,中值,上四分位数和最大观察(每组八只小鼠)的数据表示为箱子和晶须图。BD:低于检测。*:P <0.05,统计显着性与WT使用Mann-Whitney U-Test相比。gydF4y2Ba
TM的肺部gydF4y2BaLED / LEDgydF4y2Ba小鼠对促炎作用相对抵抗gydF4y2BaS.肺炎gydF4y2Ba
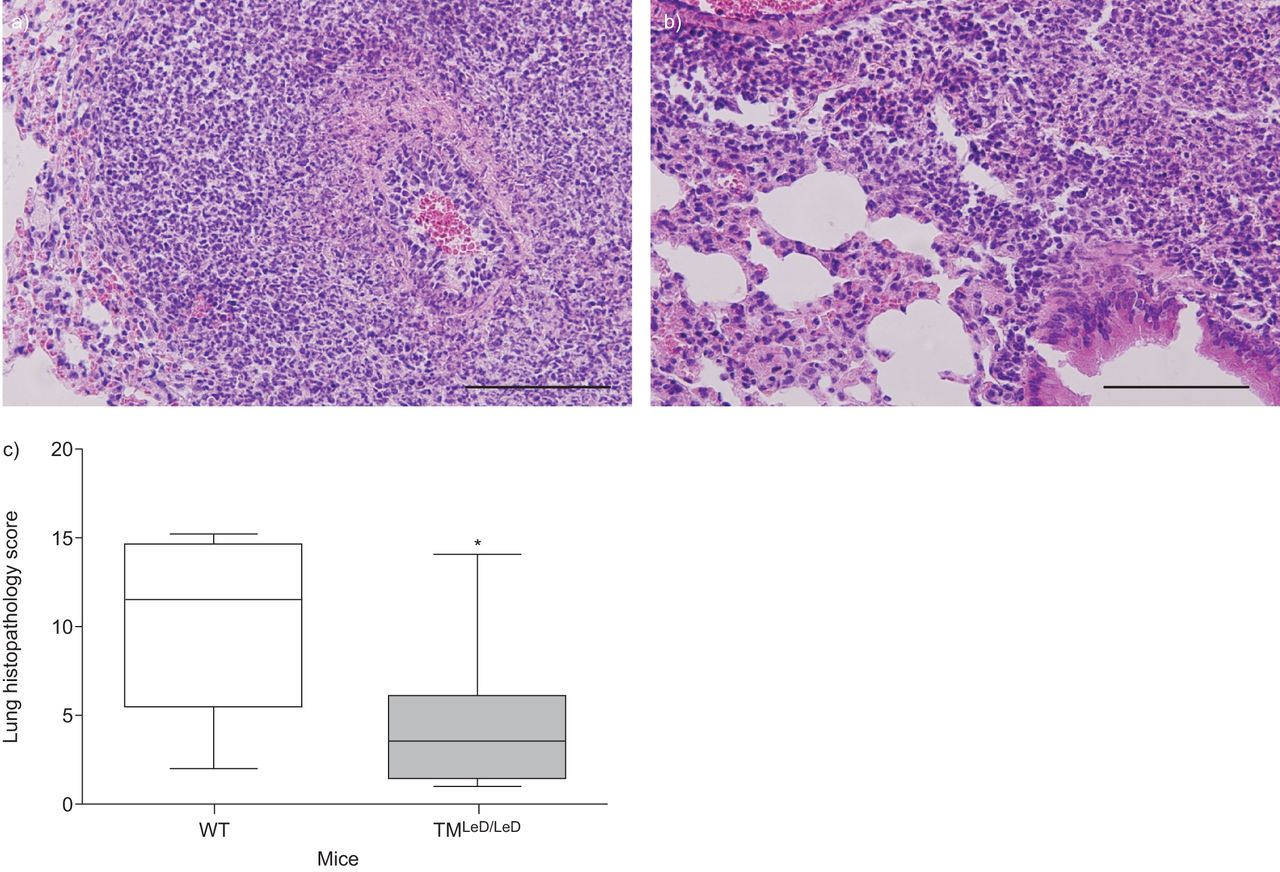
肺炎球菌肺炎与肺炎炎症和损伤有关,嗜中性粒细胞进入肺部的肺炎和支气管炎,间质炎症,水肿和内皮炎的发生。在小鼠中没有凝集素样结构域导致在内毒素吸入后的肺中的增强中性粒细胞积累[gydF4y2Ba13gydF4y2Ba].为了评估肺炎球菌肺炎中TM凝集素样结构域对中性粒细胞流入肺腔的作用,我们在肺部切片上进行了Ly-6G染色,并在感染后24和48小时测量肺匀浆中MPO的水平。虽然在24小时没有差异(数据未显示),但在48小时,TM肺中的Ly-6G阳性显著降低gydF4y2BaLED / LEDgydF4y2Ba小鼠比WT小鼠,表明中性粒细胞累积减少(gydF4y2Ba无花果。3A-C.gydF4y2Ba).在TM的肺匀浆中也显着降低了MPO水平gydF4y2BaLED / LEDgydF4y2Ba小鼠48小时(gydF4y2Ba图3 dgydF4y2Ba).与这些结果一致,同一时间点肺切片的平均组织病理学评分在TM较低gydF4y2BaLED / LEDgydF4y2Ba老鼠(gydF4y2Ba无花果。4gydF4y2Ba).为了探讨肺组织中性粒细胞中源粒细胞中源粒细胞中的差异是否导致支气管肺泡室中的细胞流入的差异,在诱导肺炎球菌肺炎后,在6,24和48小时内测定了总细胞计数,中性粒细胞数和MPO。TM之间的总细胞计数和中性粒细胞数量没有差异gydF4y2BaLED / LEDgydF4y2Ba在研究的任何时间点(gydF4y2Ba表格1gydF4y2Ba).此外,BALF MPO水平并不不同(未显示)。gydF4y2Ba
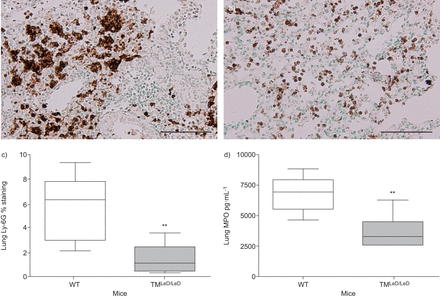
肺中性粒细胞流入和髓过氧化物酶(MPO)水平缺乏血栓调节蛋白的凝集素状结构域的小鼠(TMgydF4y2BaLED / LEDgydF4y2Ba与野生型小鼠相比,野生型小鼠感染肺炎球菌后的存活率明显降低。经鼻内接种5×10诱导肺炎48小时后,肺异thyocyanate荧光素抗小鼠Ly-6G染色(棕色)的代表性玻片gydF4y2Ba4gydF4y2Ba肺炎链球菌肺炎料gydF4y2BaCFU在a)wt小鼠和b)tmgydF4y2BaLED / LEDgydF4y2Ba老鼠。比例尺条= 100μm。c)在WT小鼠和TM中诱导肺炎球菌肺炎后肺炎术后肺炎术18小时的定量gydF4y2BaLED / LEDgydF4y2Ba老鼠。d)MPO 48小时的肺水平在WT小鼠和TM中患有肺炎球菌肺炎的感染后感染gydF4y2BaLED / LEDgydF4y2Ba老鼠。数据表示为描绘最小观察,低四分位数,中值,上四分位数和最大观察(每组八只小鼠)的数据表示为箱子和晶须图。**:P <0.01,统计显着性与WT使用Mann-Whitney U-Test相比。gydF4y2Ba
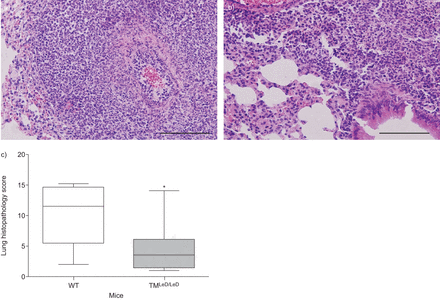
小鼠的总组织病理学评分缺乏血栓调节蛋白的凝集素样结构蛋白(TMgydF4y2BaLED / LEDgydF4y2Ba与野生型小鼠相比,野生型小鼠感染肺炎球菌后的存活率明显降低。肺血红素和eosin染色的代表性载玻片48小时诱导肺炎,鼻内接种5×10gydF4y2Ba4gydF4y2Ba肺炎链球菌肺炎料gydF4y2BaCFU在a)wt小鼠和b)tmgydF4y2BaLED / LEDgydF4y2Ba老鼠。比例尺条= 100μm。c) WT小鼠和TM感染肺炎球菌肺炎48小时后的总病理评分gydF4y2BaLED / LEDgydF4y2Ba老鼠。数据表示为描绘最小观察,低四分位数,中值,上四分位数和最大观察(每组八只小鼠)的数据表示为箱子和晶须图。*:P <0.05,统计显着性与WT使用Mann-Whitney U-Test相比。gydF4y2Ba
接下来,我们量化了感染后6、24和48小时肺部匀浆中几种趋化因子和细胞因子的水平(gydF4y2Ba表2.gydF4y2Ba).在第6和24 h,两组间肺趋化因子和细胞因子检测无差异。然而,在48小时,TM的肺匀浆gydF4y2BaLED / LEDgydF4y2BaTNF-α (p<0.001)、IL-6、IL-10、IFN-γ和MCP-1 (p<0.05)水平显著降低(p<0.05) (gydF4y2Ba表2.gydF4y2Ba).gydF4y2Ba
TM患者血浆细胞因子水平降低gydF4y2BaLED / LEDgydF4y2Ba老鼠后gydF4y2BaS.肺炎gydF4y2Ba感染gydF4y2Ba
为了进一步探讨TM的凝集素样结构域的作用,响应肺炎球菌肺炎,我们测量血浆中的炎症标志物。TM之间的差异炎症反应gydF4y2BaLED / LEDgydF4y2Ba在感染后24小时的血浆中已经在血浆中显而易见的是,TM的TNF-α,IL-6和MCP-1的点水平显着降低gydF4y2BaLED / LEDgydF4y2Ba小鼠比在wt小鼠中(gydF4y2Ba表3gydF4y2Ba),持续到48小时的差异。IL-10,IL-12和IFN-γ的血浆水平低于测定的检测限(未示出)。gydF4y2Ba
TM值gydF4y2BaLED / LEDgydF4y2Ba小鼠对gydF4y2BaS.肺炎gydF4y2Ba
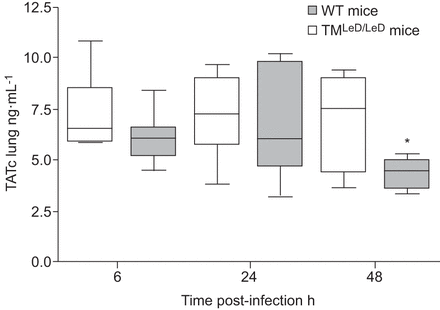
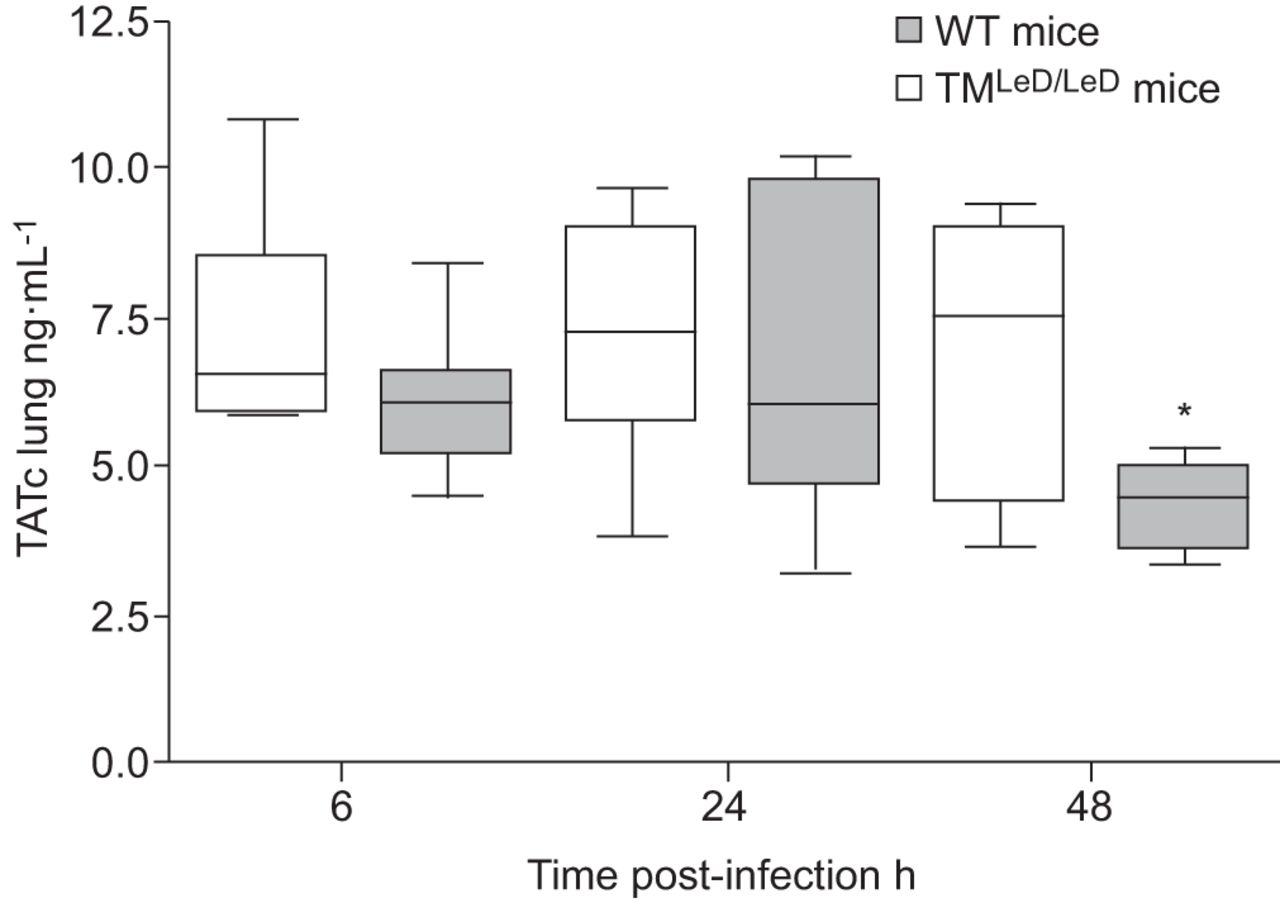
我们还通过测量肺匀浆和血浆中的TATC水平来评估TM改性肺炎球菌肺炎的凝集素状的肺炎肺炎肠道肺炎诱导的变化。在血浆或肺匀浆中感染后6和24小时,注意到TATC水平的差异。然而,在48小时,当两组小鼠(未显示的数据)仍保持相似的血浆TATC水平时,来自TM的肺组织中TATC水平显着降低gydF4y2BaLED / LEDgydF4y2Ba与野生型小鼠比较(p<0.05) (gydF4y2Ba无花果。5gydF4y2Ba).gydF4y2Ba
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
缺乏血栓调节蛋白的凝集素状结构蛋白的小鼠的肺组织中凝血的激活减少了(TMgydF4y2BaLED / LEDgydF4y2Ba)与肺炎球菌肺炎后野生型(WT)小鼠相比。用5×10后感染血浆抗凝血酶复合物(TATC)6,24和48小时的肺匀浆水平gydF4y2Ba4gydF4y2Ba肺炎链球菌肺炎料gydF4y2Bacfu在wt和tmgydF4y2BaLED / LEDgydF4y2Ba老鼠。数据表示为描绘最小观察,低四分位数,中值,上四分位数和最大观察(每个时间点的八只小鼠)的盒子和晶须图。*:P <0.05,统计显着性与WT使用Mann-Whitney U-Test相比。gydF4y2Ba
讨论gydF4y2Ba
凝血和炎症在宿主对感染的即时反应中是高度整合和相互作用的参与者。TM在凝血和炎症的调节中发挥着核心作用,其特定的功能依赖于其独特结构域的完整性。因此,egf样重复为凝血酶介导的蛋白C和TAFI激活提供辅助因子活性,从而抑制凝血、纤溶和补体,而n端凝集素样结构域显示出许多抗炎特性[gydF4y2Ba5gydF4y2Ba- - - - - -gydF4y2Ba7gydF4y2Ba].使用鼠标模型研究对细菌肺炎的宿主反应,我们先前已经研究了eGF样重复TM的作用,这是蛋白C活化的原因。TM值gydF4y2BaPRO / PROgydF4y2Ba具有适度降低的TM抗原水平的小鼠和其支持凝血酶介导的蛋白C的蛋白C活化的容量显着降低,并且与WT小鼠相比,对细菌肺炎的促凝血剂或炎症反应的TM依赖性变化没有表现出TM依赖性变化[gydF4y2Ba18.gydF4y2Ba].在这里,我们类似地研究了凝集素样域的TM在社区获得的肺炎的小鼠模型中的作用gydF4y2BaS.肺炎gydF4y2Ba.TM的此域不会参与APC的生成。与无菌炎症模型相比,在哪个凝集素样结构域的TM渲染小鼠对肺和心肌缺血再灌注损伤,炎症性关节炎和内毒素血症的丧失[gydF4y2Ba13gydF4y2Ba- - - - - -gydF4y2Ba15gydF4y2Ba],在TM的改善的存活,降低的细菌生长和传播中,术后,TM的凝集素样结构域的损失似乎保护宿主响应于肺炎球菌肺炎,并且在TM中减毒和衰减炎症和促凝血剂反应而反映gydF4y2BaLED / LEDgydF4y2Ba与wt小鼠相比的小鼠。gydF4y2Ba
这是第一个报告,其中使用可行的革兰氏阳性细菌来评估gydF4y2Ba体内gydF4y2Ba凝集素状的TM的角色。在我们的研究中,肺炎球菌载荷在TM的肺部瞬时降低gydF4y2BaLED / LEDgydF4y2Ba小鼠6小时后感染后。这种差异不能归因于改变的炎症细胞数,因为WT和TM之间的BALF总细胞计数和中性粒细胞数量不差异gydF4y2BaLED / LEDgydF4y2Ba在感染后的早期时间点。值得注意的是,未感染TMgydF4y2BaLED / LEDgydF4y2Ba小鼠在Puribronchial位置的肺插形中显示出明显增强的中性粒细胞积累[gydF4y2Ba13gydF4y2Ba]预计这预计会促进在较低航空的感染时立即进行抗菌反应[gydF4y2Ba21.gydF4y2Ba].然而,当发现血液和肝脏中的细菌载量时,差异在48小时内变得更加突出,在TM中显着降低gydF4y2BaLED / LEDgydF4y2Ba比野生型小鼠更明显。这些数据表明gydF4y2BaS.肺炎gydF4y2Ba在肺血界面,gydF4y2Ba即。gydF4y2Ba在内皮细胞层或系统间室本身,在缺乏TM的凝集素样结构域时增强。这可能是由于TM凝集素样结构域的缺失导致该位点补体活化过度,否则会下调补体活化[gydF4y2Ba22.gydF4y2Ba].的确,TM.gydF4y2BaLED / LEDgydF4y2Ba小鼠具有较低的基底血浆C3水平和降低的CH 50(导致50%溶解敏感的绵羊红细胞溶解的血清的量),两者都表明过量的补体激活[gydF4y2Ba15gydF4y2Ba].考虑到补体激活在宿主防御肺炎球菌中发挥的重要作用gydF4y2Ba体内gydF4y2Ba[gydF4y2Ba23.gydF4y2Ba],可以想到,TM的保护表型gydF4y2BaLED / LEDgydF4y2Ba小鼠至少可以部分解释为血液中肺炎球菌的清除增加了gydF4y2Ba通过gydF4y2Ba增强了补体系统的激活。gydF4y2Ba
在我们的研究中,TMgydF4y2BaLED / LEDgydF4y2Ba小鼠响应的局部和全身炎症的证据显示gydF4y2BaS.肺炎gydF4y2Ba感染。这反映了较低的TNF-α,IL-6和MCP-1的血浆水平,肺部损伤的较少组织学证据,肺匀浆中的细胞因子和趋化因子的细胞因子和趋化因子的组织学证据较少,感染肺中的嗜中性粒细胞较少,减少了凝血的二次活化通过TATC测量,最后通过相对抗致死剂量的细菌。虽然看起来与以前的报告有关gydF4y2BaLED / LEDgydF4y2Ba小鼠表现出高度的炎症反应,这些研究与内毒素或非分泌胁迫进行,而不是活革兰氏阳性细菌进行[gydF4y2Ba13gydF4y2Ba].实际上,目前的研究结果表明TMgydF4y2Ba通过gydF4y2Ba它的凝集素样结构域可能介导宿主对革兰氏阴性菌的差异反应gydF4y2Ba相对gydF4y2Ba革兰氏阳性细菌。TM的凝集素样结构域与革兰氏阴性菌的碳水化合物Lewis Y抗原结合,诱导革兰氏阴性菌的凝集和调理作用,包括gydF4y2Ba大肠杆菌gydF4y2Ba和gydF4y2BaKlebsiella肺炎gydF4y2Ba,增强巨噬细胞的吞噬作用[gydF4y2Ba24.gydF4y2Ba].通过与Lewis Y抗原相互作用,TM的凝集素状结构曲面可以干扰其与CD14的脂多糖结合,其其同源受体,从而阻尼下游促炎信号途径和促炎细胞因子的释放,以及诱导的一氧化氮合酶表达.实际上,TM的凝集素状结构域的重组可溶性形式的TM降低的克雷布氏菌gydF4y2Ba-gydF4y2Ba诱发的炎症反应和致死率[gydF4y2Ba24.gydF4y2Ba].如果不调用其他机制,这种有益的反应在我们的革兰氏阳性肺炎模型中不一定是预期的,因为内毒素没有参与。认识到这一发现的临床相关性很重要,因为可溶性血栓调节蛋白正被研究用于临床[gydF4y2Ba25.gydF4y2Ba]并识别适当的应用对于确保最佳益处至关重要。而且,虽然在非分泌胁迫模型中,抗炎反应倾向于改善损伤,在活细菌模型中,需要大量的炎症反应来杀死细菌;通过减少炎症反应干扰细菌杀伤可以增强细菌产物,从而导致不良结果。gydF4y2Ba
我们的研究有局限性。如上所述,我们在单个感染模型中研究了单一病原体。目前尚不清楚我们的研究结果是否与其他模型和/或使用其他细菌相当。此外,TM的遗传淘汰可能是可能的gydF4y2BaLED / LEDgydF4y2Ba在小鼠生长发育过程中引起了未知的代偿性变化。在这方面,研究干扰对TM功能的影响是很有意义的gydF4y2BaLED / LEDgydF4y2Ba而不是打倒TMgydF4y2BaLED / LEDgydF4y2Ba例如,通过使用阻断抗体。gydF4y2Ba
总之,我们表明,在小鼠中的凝集素样结构域的缺失导致对肺炎球菌肺炎的改善,是严重的社区获得感染的常见原因。额外的研究是必要的,以进一步了解革兰氏阳性凝集素样TM域的可能差异作用gydF4y2Ba相对gydF4y2Ba革兰氏阴性感染,因为这将确定如何最佳设计靶向疗法,以增强功能或干扰这种有趣的内皮C型凝集素结构的功能,从而在不同的临床情况下获得最大益处。gydF4y2Ba
致谢gydF4y2Ba
作者感谢以下人员,全部在荷兰阿姆斯特丹学术医疗中心,Amsterdam的学术和分子医学中心工作:M. Ten Brink和J. Daalhuisen在动物实验期间为他们的技术援助,R. De Beer用于表演(immuno)histopathological stainings and A. Achouiti and F.E. van den Boogaard for assisting in cytokine measurements.
脚注gydF4y2Ba
支持声明gydF4y2Ba
M. Schouten得到了研究的支持gydF4y2Ba荷兰血栓形成基础gydF4y2Ba(授予号码TSN 2005-1)。J.D. De Boer由授权支持gydF4y2Ba荷兰哮喘基金会gydF4y2Ba(项目编号3.2.08.009)。gydF4y2Ba
兴趣表gydF4y2Ba
没有宣布。gydF4y2Ba
- 收到了gydF4y2Ba2012年1月25日。gydF4y2Ba
- 接受gydF4y2Ba2012年7月10日。gydF4y2Ba
- ©2013人队gydF4y2Ba
参考文献gydF4y2Ba







