





文摘
我们调查的角色集落刺激因子(gm - csf)在subchronic香烟烟雾暴露模型(CS)全身炎症使用抗体针对gm - csf或gm - csf受体(GM-CSFR)α-chain。
CS-induced单核细胞和中性粒细胞炎症后4天的CS暴露于BALB / c小鼠评估在支气管肺泡灌洗(BAL)流体。成熟的树突状细胞(dc)的增加(CD11c +和主要组织相容性复合体II +)和Gr-1-high中性粒细胞也观察到流仪分析全部肺组织。
每天i.p。注入400μg gm - csf或GM-CSFR抗体之前每天吸烟暴露减毒球内中性粒细胞60%的累积。成熟的DCs的减少也被观察到。Anti-GM-CSFR抗体政府没有影响肺t细胞的比例;然而,显著减少观察CD69 + CD8 + t细胞激活。Anti-GM-CSFR抗体政府的mRNA和蛋白表达降低interleukin-12 p40和基质金属蛋白酶12。
综上所述,与这种受体抗体结合gm - csf通路干预的一个重要中介烟雾诱发炎症。
同时肺暴露于香烟烟雾中(CS)被确认为是导致慢性阻塞性肺疾病(COPD)、疾病的具体致病机制不理解1]。慢性阻塞性肺病的特点是不可逆气流阻塞和进步的肺部炎症,与疾病严重程度2]。虽然哮喘更与嗜酸性粒细胞的炎症反应有关,在慢性阻塞性肺病,中性粒细胞和巨噬细胞为主3]。
作为一个关键调节器的骨髓细胞生存和激活(4),细胞因子集落刺激因子(gm - csf)已被证明有一个核心作用在维护健康的肺的先天免疫反应(5,6]。它是发布的一系列结构和炎症细胞,如气道上皮细胞、平滑肌细胞、t细胞和巨噬细胞。它也表明,gm - csf高架在COPD患者的肺7- - - - - -9),这表明这种细胞因子可能也扮演了一定的角色夸张的炎症反应的疾病。gm - csf的贡献还不清楚;然而,它已经表明,gm - csf不仅激活巨噬细胞和中性粒细胞与刺激,如脂多糖,可能出现在COPD肺,也促进了细胞因子释放和细胞存活率(2]。最近,它还表明,鼻内交付anti-GM-CSF抗体的小鼠模型CS-induced炎症肺嗜中性变弱的10]。
gm - csf调和它的影响通过gm - csf受体(GM-CSFR)由两个亚基组成,一个α-subunit,结合亲和力较低的细胞因子,和一个更大的β-subunit(β普遍;βc),负责信号,形成三元复杂的受体(11]。信号转导的细胞因子白介素(IL) 3和IL-5也是由βc;因此,受体特异性是由于GM-CSFRα[12]。因为CS是慢性阻塞性肺病的主要病原体之一,许多CS-exposure模型已经开发出来,表现为表型类似于慢性阻塞性肺病,如肺气肿和肺发炎(13- - - - - -15),这些模型提供平台评估新疗法的疗效和机制。生物制剂越来越被开发为慢性疾病,如肿瘤坏死因子(TNF) -α抗体和受体拮抗剂在关节炎(16- - - - - -18],anti-immunoglobulin (Ig) E抗体在哮喘19]。此外,抗体试验在慢性阻塞性肺病也正在这一目标IL-1β(canakinumab) [20.]。目前许多anti-GM-CSF和anti-GM-CSFRα[4)方法在早期类风湿性关节炎的临床开发。迄今为止,只有一项研究进行了调查gm - csf CS-exposure模型的作用通过鼻内交货(10]。在目前的研究中,我们继续这项工作通过评估中和抗体gm - csf (22 e9)和GM-CSFRα(凸轮- 3003)抗体前subchronic CS模型通过系统性舱确定抑制这个途径的影响,通过临床相关的抗体管理路线。此外,我们还探讨了gm - csf对下游通路封锁细胞因子的作用,及其对树突状细胞(DC)的子集的影响在肺癌和CD8 + t细胞激活。
材料和方法
动物
雌性BALB / c小鼠(6 - 8周的年龄)从查尔斯河实验室购买(加拿大蒙特利尔,QC)。il - 1受体敲除和野生型(WT)控制C57BL / 6小鼠获得杰克逊实验室(美国我巴尔港)。老鼠保持在特定的无菌条件下,在12 h明暗周期,提供食物和水随意。麦克马斯特大学的动物研究伦理委员会(汉密尔顿,加拿大)批准所有的实验。
FDCP-1细胞增殖试验
FDCP-1细胞被维护在杜尔贝科修改鹰的介质(表达载体,伯灵顿,加拿大)含10%胎牛血清(的边后卫;美国KS JRH生物科学,Lenexa ng)和5毫升−1小鼠gm - csf(研发系统,明尼阿波利斯,美国)。试验前,细胞培养在没有37°C的gm - csf 30 h 5%的二氧化碳。细胞被resuspended 1.5×105细胞·毫升−1100年中期和μL这个暂停添加到每个平底的好,96孔板。细胞孵化37°C 5%二氧化碳与抗体的小鼠gm - csf 16 h。1 - 2点20μL氚化胸苷(5.0 mCi·毫升−1)被添加到每个收获前和孵化4 h对GF / C Unifilter™板块(珀金埃尔默,弗吉尼亚州伍德布里奇,加拿大)。胸腺嘧啶核苷掺入决心使用闪烁计数器(Packard Topcount;美国帕卡德仪器公司,梅里登,CT)。数据分析使用棱镜软件(GraphPad、圣地亚哥、钙、美国)。
CS-exposure协议
老鼠暴露在CS从12 2 r4f参考香烟过滤器移除(烟草与健康研究所,列克星敦,肯塔基州,美国)∼50分钟,每日两次,4天使用全身烟雾暴露系统(SIU-48;Promech实验室AB, Vintrie,瑞典)如前所述21]。CS曝光后最初的适应阶段,1天,老鼠放入20分钟的曝光盒,2天,30分钟和3天,50分钟。对照组(假的),被暴露于室内空气。
i.p。政府的抗体
BALB / c小鼠注射i.p。与400年μg anti-GM-CSFR(凸轮- 3003;鼠标IgG1)或anti-GM-CSF配体(MP122E9克隆;老鼠IgG2a;研发系统)抗体前12 h每天的第一个CS曝光。老鼠牺牲12 - 18 h后CS曝光。
支气管肺泡灌洗和微分细胞计数
支气管肺泡灌洗(BAL)流体收集如下。简而言之,肺解剖,插管和0.25毫升冰冷的1×PBS灌输,其次是0.2毫升1×PBS。总细胞使用血球计人数统计。Cytospins准备微分细胞计数和彩色丙烯酸- 3(生化科学Inc . Swedesboro,新泽西,美国)。500细胞数/ cytospin识别单核细胞、中性粒细胞和嗜酸性粒细胞。
ELISA和中尺度发现分析
gm - csf的ELISA试剂盒购自研发系统和使用根据制造商的协议。细胞因子检测干扰素(IFN) -γIL-1β,il - 4, keratinocyte-derived细胞因子(KC)和il - 12进行使用multi-array小鼠炎性和辅助细胞(Th) 1 / Th2型细胞因子面板检测系统(MesoScaleDevices,盖瑟斯堡,医学博士,美国)。
Fluidigm mRNA表达分析
从单个叶使用RNA提取试剂盒RNeasy纤维组织包根据制造商的协议(试剂盒、希尔登,德国)。RNA是量化和正常化,和完整性评估使用安捷伦RNA 6000纳米工具包(美国安捷伦,圣克拉拉,CA)。互补脱氧核糖核酸的一代是用超级脚本三世工具包(美国生活技术,Carlsbard, CA)。相对转录表达评估使用Fluidigm Biomark动态数组(Fluidigm,阿姆斯特丹,荷兰)装载探针对感兴趣的记录。
隔离肺单核细胞
肺单核细胞分离(如前所述21]。简而言之,肺灌注了1×PBS和细胞悬浮液由解集和孵化1 h在150年37°C U mL−1胶原酶III在羟乙基哌嗪乙磺酸acid-buffered生理盐水(表达载体)。碎片被通过尼龙网。细胞被resuspended 1×PBS含有0.3%牛血清白蛋白(BSA)(英杰公司)或RPMI补充10%的边后卫(Sigma-Aldrich,奥克维尔,加拿大),1%l谷氨酰胺和1%青霉素和链霉素(表达载体)。
流仪结果
1×106肺单核细胞被洗一次1×PBS / 0.3% BSA和沾直接共轭主要抗体30分钟在4°C。105生活事件是获得使用LSR II (BD生物科学,米西索加、加拿大)流式细胞分析仪和FlowJo分析软件(TreeStar Inc .)、亚什兰,美国)。以下使用抗体:荧光素isothiocyanate-conjugated anti-CD11c,藻红蛋白(PE)共轭anti-CD11b, PE-Alexa萤石610 -共轭CD8 PE-cy5.5-conjugated anti-CD19, PE-cy7-conjugated anti-CD69, allophycocyanin (APC)共轭anti-major组织相容性复合体(MHC)二类,Alexa萤石700 -共轭anti-CD86 APC-cy7-conjugated anti-CD45,太平洋Blue-conjugated anti-CD3和太平洋Orange-conjugated anti-Gr-1。这些抗体购自BD生物科学(美国加利福尼亚州圣何塞)或eBioscience(美国圣地亚哥,CA)除了CD86 (BioLegend、圣地亚哥、钙、美国)。量子点(Qdot) 605 -共轭anti-CD4和Qdot655-conjugated anti-B220买来英杰公司(美国加利福尼亚州卡尔斯巴德)。
统计分析
数据分析使用棱镜软件版本5 (Graphpad)和表达为±扫描电镜。正态分布统计分析是由测试使用Kolmogorov-Smirnov测试紧随其后的是一个未配对t检验,单向方差分析或双向方差分析Bonferroni检测后,在数据如上所述。非参数测试进行使用Kruskall-Wallis Mann-Whitney测试后测试。被认为具有统计显著性差异p < 0.05时。
结果
CS-induced炎症与gm - csf mRNA和蛋白表达增加有关
为了研究CS暴露的影响在肺部,gm - csf表达我们暴露BALB / c小鼠为4天CS。控制动物暴露于室内空气(假的)。CS暴露导致显著增加细胞总数落下帷幕,在这主要反映出中性粒细胞(图1一个)。CS显著增加肺的gm - csf水平(图1 b)。
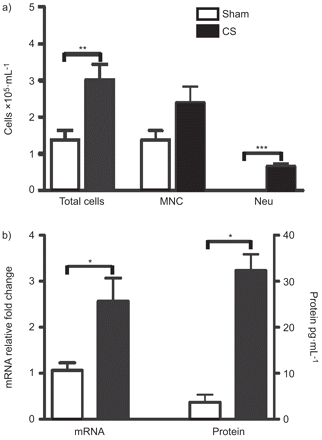
支气管肺泡灌洗(BAL)细胞轮廓和集落刺激因子(gm - csf)表达在香烟烟雾(CS)暴露的老鼠。BALB / c小鼠骗局——或者CS-exposed 4天。BAL总细胞数,单核细胞(跨国公司)和中性粒细胞(Neu)。n = 5 /组。b) gm - csf mRNA(骗局:n = 3;c: n = 8)和蛋白质表达每组(n = 5)在肺部的骗局,CS-exposed老鼠。使用非参数统计分析进行Kruskall-Wallis Mann-Whitney测试后测试。酒吧代表手段和胡须代表的标准错误的意思。*:p < 0.05;* *:p < 0.01; ***: p<0.001.
gm - csf表达机制
尽管多个通路可能导致CS-induced炎症,接下来我们试图调查是否表达gm - csf烟雾暴露后il - 1受体(IL-1R) 1-dependent。这个实验是基于最近的一份报告打et al。(22)展示在CS-induced IL-1R1炎症的一个重要的角色。与打的结果一致et al。(22),CS-induced嗜中性IL-1R-deficient小鼠明显减弱(图2一个)。失去IL-1R表达式并没有显着影响的CS曝光后单核细胞的增加。我们进一步分析了gm - csf mRNA表达小鼠C57BL / 6 (WT)和IL-1R-deficient虚假或CS暴露。gm - csf mRNA表达废除IL-1R-deficient老鼠(图2 b)。

支气管肺泡灌洗(BAL)细胞轮廓和集落刺激因子(gm - csf)表达在香烟烟雾(CS)暴露interleukin-1受体基因敲除小鼠(IL-1R)。一)野生型和IL-1R- / -老鼠被暴露于室内空气(假的)或CS 4天。矿山总细胞数,单核细胞(跨国公司)和中性粒细胞(Neu)所示。b) gm - csf mRNA表达虚假的,CS-exposed野生型IL-1R- / -老鼠。酒吧代表手段和胡须代表的标准错误的意思。n = 5 /组。执行统计分析使用双向方差分析与野生型的Bonferroni测试后与淘汰赛。*:p < 0.05;* *:p < 0.01。
Anti-GM-CSF配体和受体抗体减弱CS-induced肺部炎症
为了测试的重要性,gm - csf CS-induced气道炎症,我们管理anti-ligand (22 e9)或receptor-blocking抗体(凸轮- 3003)骗局——或者CS-exposed老鼠。的活动中的每个抗体确认FDCP-1扩散试验(图3)。22 e9的抑制浓度中位数和凸轮- 3003是919点(95%可信区间694 - 1218点)和130点(95%可信区间99 - 171点),分别。由于不同的效能和同形像这些抗体,完全饱和剂量的抗体被用于这些研究。药代动力学分析凸轮- 3003进行确认抗体之间的接触水平骗局,CS-treated动物肺和血液隔间。之间没有区别观察抗体接触骗局,CS-treated动物(图3 b)。在活的有机体内anti-GM-CSF anti-GM-CSFR抗体管理局导致类似,显著减毒CS-induced中性粒细胞炎症。控制老鼠收到同形像控制抗体表明没有attentuation CS-induced炎症(图3 c和d)。为了确定在中性粒细胞数间质改变,流式细胞仪肺单细胞的悬浮液进行。Gr-1-high细胞的百分比显著增加了CS暴露与sham-treated动物相比(p < 0.05)。之间无显著差异观察CS-exposed同形像控制——anti-GM-CSF——或者anti-GM-CSFR-treated老鼠(图4)。
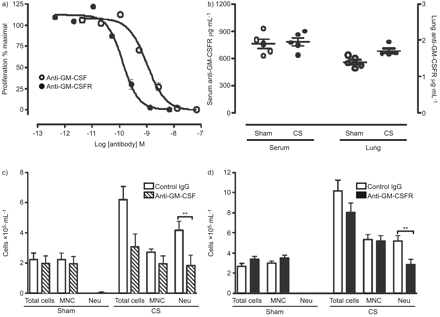
集落刺激因子(gm - csf)中和活动和支气管肺泡灌洗细胞在香烟烟雾(CS)暴露小鼠注射anti-GM-CSF配体或anti-GM-CSF受体(anti-GM-CSFR)抗体。)效力比较anti-GM-CSF和anti-GM-CSFR鼠标gm - csf FDCP-1扩散试验。细胞培养小鼠gm - csf和一系列稀释的抗体16 h。细胞增殖是量化后进一步4 h与氚化胸苷孵化。b)终端接触anti-GM-CSFR水平血清和肺匀浆的骗局,CS-exposed老鼠。BALB / c小鼠暴露于CS 4天。小白鼠注射i.p。与c) anti-GM-CSF配体或d) anti-GM-CSFR,或同形像控制每日CS曝光前18 h抗体。数据显示细胞总数,单核细胞(跨国公司)和中性粒细胞(Neu)。酒吧代表手段和胡须代表的标准错误的意思。n = 5 /组。统计分析了使用双向方差分析Bonferroni测试后的骗局与CS组。* *:p < 0.01。
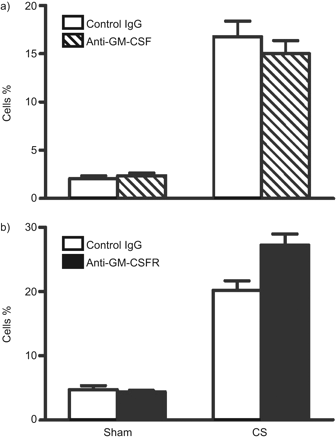
流仪分析中性粒细胞(Gr-1-high)骗局和香烟(CS)暴露小鼠注射anti-granulocyte-macrophage集落刺激因子(gm - csf)配体或anti-GM-CSF受体(GM-CSFR)抗体。BALB / c小鼠暴露于CS 4天。小白鼠注射i.p。)anti-GM-CSF或b) anti-GM-CSFR抗体,或同形像控制抗体。中性粒细胞(Gr-1-high主要组织相容性复合体类II-high)在全部肺检查,通过流式细胞术单细胞悬浮体。统计分析了使用双向方差分析Bonferroni测试后的骗局与烟。
Anti-GM-CSF配体和受体抗体减弱CS-induced直流扩张和t细胞激活
人们普遍认为gm - csf促进DC的成熟,一个关键的步骤,抗原驱动(综述[t细胞反应4])。因此,我们调查是否干预与anti-GM-CSF或受体抗体衰减直流扩张和t细胞活化后CS曝光。Anti-GM-CSF配体抗体干预废除CS-induced增加骨髓DCs (mdc) (CD11c-high MHC II-high)和血浆DCs(髓)(CD11c-high MHC II + B220 +) DCs (图5 a和b)。管理一个anti-GM-CSFR抗体也减小了mDCs频率(图5 c和d),而没有观察pDCs效应。

流仪分析树突细胞(DC)子集并激活t细胞在香烟烟雾(CS)暴露小鼠注射anti-granulocyte-macrophage集落刺激因子(gm - csf)配体或anti-GM-CSF受体(GM-CSFR)抗体。BALB / c小鼠暴露于CS 4天。小白鼠注射i.p。用a、b、e) anti-GM-CSF配体抗体或c, d, f) anti-GM-CSFR抗体,或同形像控制抗体。a, c)骨髓DCs (CD11c-high主要组织相容性复合体(MHC)类II-high B220)和b, d)血浆DCs (CD11c-high MHC II-high B220 +),或e, f)激活CD69 + CD4 +和CD8 + t细胞亚群在全部肺单细胞通过流式细胞仪检测。统计分析了使用双向方差分析Bonferroni测试后的骗局与CS。搞笑:免疫球蛋白。*:p < 0.05;* *:p < 0.01。
Anti-GM-CSF配体抗体干预废除CS-induced增加激活CD69 + CD4 +和CD8 + t细胞与同形像control-treated动物。ligand-blocking抗体相比,减少只是观察CD69 + CD8 + T细胞,但不是CD4 + T细胞(图5 e和f),在小鼠anti-GM-CSFR抗体。
Anti-GM-CSFR抗体治疗可以降低il - 12,基质金属蛋白酶12 mRNA和蛋白表达在CS-exposed BALB / c小鼠
为了研究CS-induced炎症的机制,我们分析了肺癌细胞因子和趋化因子的表达,以及基质金属蛋白酶(MMPs),使用Fluidigm分析。CS暴露导致显著增加mRNA表达:细胞因子IL-12p40, IL-1β,IL-33, il - 6,粘蛋白5 ac, TNF-α;趋化因子KC,单核细胞趋化蛋白1,主题趋化因子配体(CCL) 3,亚兰,CCL9,集落刺激因素3和科学家图案配体(CXCL) 2;和MMP9 MMP12 (表1)。Anti-GM-CSFR抗体明显减毒的mRNA表达IL-12p40 (图6)和MMP-12 (图6 b相对于同形像控制抗体治疗(图6和表1)。没有观察到显著差异与TNF-α或CXCL2相对于同形像控制(图6 c和d)。
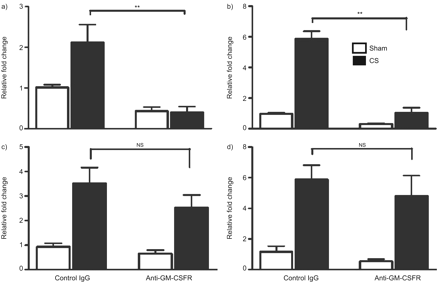
一)Interleukin-12 p40, b)基质金属蛋白酶12 c)肿瘤坏死factor-α和d)科学家配体2 mRNA的表达。BALB / c小鼠暴露于室内空气(假的)或香烟(CS) 4天。小白鼠注射i.p。与anti-granulocyte-macrophage集落刺激因子受体(GM-CSFR)或同形像控制抗体。所有值正常化三管家基因和表达为相对褶皱变化sham-exposed老鼠。酒吧代表手段和胡须代表的标准错误的意思。n = 5 /组。统计分析进行了比较CS-exposed控制免疫球蛋白G (Ig)与anti-GM-CSFR antibody-treated老鼠使用Mann-Whitney紫外线测试。ns:不重要的。* *:p < 0.001。
为了确认mRNA表达数据,我们对细胞因子的表达IL-1βKC, IFN-γ,2、il - 12、il - 4。IL-1β表达的增加,KC,观察il - 12在CS-exposed动物注射同形像抗体控制。anti-GM-CSFR抗体可以显著降低表达il - 12在球和肺匀浆,虽然IL-1β或KC水平保持不变。类似的观察是为动物治疗IL-1β水平anti-ligand抗体(补充表1)。IFN-γ、2、il - 4含量均低于在虚假的检测极限,CS-treated组(表2)。
讨论
中性粒细胞和巨噬细胞经常与COPD的发病机制。以往的经验显示,随着疾病的严重程度的发展,在气道炎症细胞的比例增加(2]。因此,有人提出干扰机制,缓解这些细胞活动的可以提供临床益处。因此,gm - csf在慢性阻塞性肺病被视作潜在的中介。
尽管gm - csf高架在COPD患者的肺或慢性支气管炎(8),针对CS-induced肺部炎症模型这一途径的干预措施并没有被广泛描述(10]。在这里,我们表明,4天的CS接触后,我们观察到一个一致的增加总细胞落下帷幕,绝大多数被中性粒细胞。此外,gm - csf转录和蛋白水平升高在这些动物的肺。有趣的是,虽然多等。(10]证明了衰减与抗体gm - csf CS-induced肺部炎症,他们无法测量球的gm - csf蛋白质。同样,他们也展示了衰减的中性粒细胞和巨噬细胞;CS-induced炎症的然而,在我们的模型中,我们观察到中性粒细胞增加和随后的衰减,使用抗体。的注意,CS-exposure的类型系统和抗体交付的路线是不同的,因此,可能占本研究观察之间的细微差别和先前描述10]。
为了确定gm - csf的招聘途径中扮演了重要角色炎症细胞进入落下帷幕,我们评价一个anti-GM-CSF中和抗体(22 e9)和一个anti-GM-CSFRα-chain抗体(CAM3003)在这个模型。,排除潜在的依赖抗体的细胞毒性anti-GM-CSFR抗体是专门表示为一个鼠标IgG1抗体。因此,没有证据表明在观察外周血中性粒细胞减少这种抗体(数据没有显示)。使用方法,我们抑制中性粒细胞的招聘剂量的落下帷幕通过系统性舱;然而,我们并没有减少中性粒细胞数量的组织,所定义的流式细胞术。这个观察球与先前的观察,鼻内交付一致相同的gm - csf抗体(e9 22日)据报道,钝CS-induced嗜中性粒细胞和巨噬细胞招聘在球10,23]。虽然我们没有观察到显著增加巨噬细胞和淋巴细胞在CS曝光,我们观察一个温和但显著改变的百分比成熟肺DCs减毒与抗体。在体外,它已经表明,柴油废气particle-conditioned媒介从上皮细胞可以诱导GM-CSF-dependent DC成熟(24),这表明CS暴露也可能对DC成熟以类似的方式作出贡献。这是支持的观察,反过来,CD4 +和CD8 + t细胞CD69的表达增加CS曝光后如前所述[21),这部分被封锁的gm - csf途径。有趣的是只有CD8 + t细胞激活抑制的机制,但是,CD4 + CD69 +激活t细胞不受anti-GM-CSFR治疗的影响。这样做的原因是不清楚和进一步研究。然而,t细胞不表达GM-CSFRα-chain;因此,这些数据表明,减少CS-induced t细胞激活可能间接影响gm - csf刺激骨髓细胞,而不是直接gm - csf对t细胞的影响。
最近的研究(22]表明MyD88, toll样受体4 - IL-1R-deficient老鼠都有肺嗜中性和细胞因子水平降低后的肺CS曝光。因此,我们研究了gm - csf表达CS-exposed IL-1R-deficient老鼠。在这个系统中,我们证实CS-induced嗜中性镇压,KC mRNA水平显著地抑制(数据没有显示)。根据我们的观察,得出的结论是,gm - csf在CS中扮演着中心角色肺部炎症,可能IL-1R通路的下游。类似的观察也被观察到在一个小鼠模型的IL-1β-driven monoarthitis [25]。
为了确定gm - csf封锁的机制,我们研究了各种蛋白质和mrna的表达水平的变化在肺。当CS曝光增强一系列细胞因子的表达,挡住了gm - csf途径并没有抑制细胞因子通常与中性粒细胞炎症有关,如IL-1β和KC。gm - csf激活中性粒细胞,促进肺血管内皮的粘附(26,招聘27和敏化作用,趋化因子28]。缺乏直接影响KC, IL-1β下游和TNF表明gm - csf可能运作这些细胞因子,可能呈现中性粒细胞减少响应这些分子的直接影响,也可以占的比例没有明显区别从CS-exposed小鼠中性粒细胞在肺匀浆处理一个anti-ligand抗体或anti-receptor抗体。它也表明,gm - csf促进中性粒细胞生存通过阻止细胞凋亡(29日),因此,gm - csf封锁也可能增强中性粒细胞凋亡和巨噬细胞间隙。进一步的研究是必要的,以便更好地了解矿山的中性粒细胞数量减毒和肺内的居民中性粒细胞的激活状态。
然而,gm - csf的抑制途径在CS接触抑制IL-12p40和白介素转录和蛋白质的生产水平,分别。在体外,gm - csf此前从DCs和巨噬细胞产生il - 1230.]。此外,在gm - csf暴露小鼠缺乏病毒(31日)或细菌(32)病原体证明il - 12的减少生产。此外,过度的gm - csf在正常小鼠的肺刺激增加il - 12的生产(33]。同样,我们表明,IL-12p40调节肺的CS-exposed老鼠和这些水平减毒与抗体,表明IL-12p40表达式CS-induced gm - csf的下游生产。慢性暴露于CS老鼠+添加病毒或病毒模仿(polyI: C)已经被证明可以促进增加IL-12p40生产(34];但是,这是第一次这海拔已被证明在这种subchronic模型。然而,相对很少有人知道这个分子的表达及其在慢性阻塞性肺病(作用24),因此,进一步的研究是必要的。
除了upregulation IL-12p40, subchronic烟雾暴露也刺激增加MMP-12信使rna,与先前的报告相一致(35]。组织改造和蛋白水解损伤是慢性阻塞性肺病的特征,和蛋白酶,如MMP9 MMP12,已经被证明是提高COPD肺内(36,37]。此外,老鼠缺乏MMP-12不存在与一个气肿慢性吸烟暴露后表型(38]。中性粒细胞和巨噬细胞都是已知的丰富来源MMP-12 [39),他们的数量与疾病严重程度在慢性阻塞性肺病40]。在另一个CS模型引起的肺部炎症,巨噬细胞被称为蛋白酶的主要来源(35]。在目前的研究中,我们已经证明,阻断gm - csf信号衰减MMP-12表达的肺部组织。这些数据符合观察gm - csf能诱导MMP-12生产从人类外周血monocyte-derived巨噬细胞(41,42]。有趣的是,当我们观察肺MMP-12 mRNA水平的变化,最近描述(10),我们没有观察到显著改变TNF-α信使rna或巨噬细胞炎性蛋白2α(图6 c和d)在此系统中,这表明交货路线可能会略有不同影响mRNA肺内的资料。这种差异并不是由于抗体接触,药代动力学分析凸轮- 3003确认显著水平的抗体血清和肺匀浆图3 b)。
总之,目前的研究报告,系统交付anti-GM-CSF或anti-GM-CSFR抗体减弱CS-induced嗜中性。此外,gm - csf似乎在激活居民肺癌DCs和淋巴细胞中发挥作用,提供一个初始先天反应之间的联系subchronic CS曝光和支持适应性反应。描述这一关系,进一步的研究是必要的阐明潜在作用的细胞因子在慢性阻塞性肺病。
确认
作者欣然承认j . Kasinska和美国的专家技术支持Kianpour(汉密尔顿,麦克马斯特大学,加拿大),j·艾文(MedImmune公司,剑桥,英国),p . Brohawn和a·凯勒(美国马里兰州盖瑟斯堡两MedImmune LLC,),和杂种细胞小组落实(剑桥)。
脚注
可以从本文的补充材料www.www.qdcxjkg.com
支持声明
J.K. Nikota是由安大略省的早期研究奖计划支持(时代)。核磁共振Stampfli持有加拿大卫生研究院为研究新研究员奖。
感兴趣的语句
感兴趣的语句N.H.E.戴维斯,静电的科恩I.K.安德森,A.J. Coyle, r . Kolbeck教训了,核磁共振Stampfli和硕士Sleeman可以找到www.www.qdcxjkg.com/site/misc/statements.xhtml
- 收到了2010年5月14日。
- 接受2011年2月2日。
- ©2011人队











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}