





文摘gydF4y2Ba
气道上皮屏障功能受损已成为一个关键因素在过敏性哮喘的发病机制。我们旨在辨别的参与表皮生长因子受体(EGFR)在allergen-induced上皮屏障损伤,正如我们以前观察到屋尘螨通过表皮生长因子受体(HDM)信号。gydF4y2Ba
我们研究了人类支气管上皮细胞的交界完整性使用电动细胞基质阻抗传感和immunofluorescent染色。gydF4y2Ba
HDM诱导迅速,瞬态上皮抵抗力下降,伴随delocalisation钙粘蛋白和带occludens(佐薇)1,和蛋白水解乳沟后者。EGFR抑制由AG1478降低HDM-triggered上皮在减少阻力和提高恢复上皮连接。同样,AG1478增加上皮屏障复苏electroporation-induced受伤后,虽然延迟伤口愈合的迁移阶段的反应。HDM-promoted再分配钙粘蛋白介导的gydF4y2Ba通过gydF4y2BaEGFR-dependent激活protease-activated受体(PAR) 2,而随之而来的是杆/ EGFR-independent ZO-1退化。重要的是,纤维化细胞因子的转化生长因子(TGF) -β长期HDM-induced EGFR磷酸化,抑制ligand-induced EGFR掩饰/退化,导致持续的钙粘蛋白和ZO-1再分配。gydF4y2Ba
因此,allergen-induced杆/ EGFR-mediated信号降低上皮抵抗力,促进结拆卸。TGF-β-enhanced EGFR信号可能是一个重要的贡献者屏障功能障碍,增加上皮漏洞响应HDM。gydF4y2Ba
- 钙粘蛋白gydF4y2Ba
- 上皮细胞连接gydF4y2Ba
- protease-activated受体2gydF4y2Ba
- 改变增长factor-βgydF4y2Ba
- 带occludens-1gydF4y2Ba
过敏性哮喘的特点是allergen-induced气道炎症,高反应性(AHR)和改造。呼吸道上皮细胞形成第一个障碍对沉积高空过敏症和过敏反应的起始中扮演着关键的角色,以及气道重塑。破裂的细胞间连接不仅促进运输过敏原,但也可能促进炎性反应gydF4y2Ba1gydF4y2Ba和生长因子释放gydF4y2Ba2gydF4y2Ba上皮细胞。因此,战略指向维护和/或恢复上皮屏障功能可能是至关重要的。细胞间连接上皮屏障功能的结构基础,包括紧,。粘合连接处并且是由粘合连接处并且粘附分子钙调节上皮结构通过亲同种抗原的交互和被连环蛋白与细胞骨架gydF4y2Ba3gydF4y2Ba。越水、局部紧密连接的主要贡献者是上皮抵抗,制约paracellular渗透率,是由相互作用的跨膜蛋白,gydF4y2Ba如。gydF4y2Baoccludin claudins,由带固定在细胞骨架occludens(佐薇)1和cingulin。gydF4y2Ba
在哮喘气道上皮完整性往往是妥协,超然的纤毛细胞,减少钙粘蛋白表达在这些网站,紧密连接的破坏和增加渗透率过敏原gydF4y2Ba4gydF4y2Ba- - - - - -gydF4y2Ba6gydF4y2Ba。许多过敏原,包括屋尘螨(HDM),含有蛋白水解活性,导致上皮损伤gydF4y2Ba7gydF4y2Ba,gydF4y2Ba8gydF4y2Ba。此外,蛋白酶作用于protease-activated受体(PAR) 2,气道上皮细胞和表达的受体与细胞内炎性信号通路gydF4y2Ba9gydF4y2Ba。最近,杆激活被描述为扰乱E-cadherin-mediated联系人gydF4y2Ba10gydF4y2Ba和影响上皮完整,以及炎性活动gydF4y2Ba1gydF4y2Ba。此外,是一个关键组成部分差别钙粘蛋白对这些epithelial-to-mesenchymal过渡(EMT),一个参与组织修复和改造的过程。重要的是,最近的研究表明,EMT的发展有助于哮喘的表型异常上皮gydF4y2Ba11gydF4y2Ba,gydF4y2Ba12gydF4y2Ba。gydF4y2Ba
然而,潜在的结构性变化机制proteolytically活跃,但通常无害,哮喘的过敏原仍然定义。哮喘上皮可能更加脆弱gydF4y2Ba6gydF4y2Ba和/或效率低下在再生损伤,随着支持增加水平的修复标记转化生长因子(TGF) -β和表皮生长因子受体(EGFR)在哮喘气道gydF4y2Ba2gydF4y2Ba,gydF4y2Ba13gydF4y2Ba,gydF4y2Ba14gydF4y2Ba。尽管有证据表明,表皮生长因子受体是调节在哮喘gydF4y2Ba15gydF4y2Ba、功能意义还不清楚。我们曾观察到表皮生长因子受体在支气管上皮HDM-induced信号是至关重要的gydF4y2Ba16gydF4y2Ba。这里我们试图确定表皮生长因子受体也参与结构变化对HDM曝光。我们发现HDM瞬变干扰上皮路口,表皮生长因子受体的激活显著贡献。此外,存在TGF-β持续EGFR-mediated上皮连接中断,因此,可能会增加HDM脆弱性。gydF4y2Ba
方法gydF4y2Ba
细胞培养和激励gydF4y2Ba
人类支气管上皮细胞系16 hbe14o - (16 hbe)请提供的特区Gruenert(美国加州大学,旧金山,CA)和培养EMEM介质/ 10%胎牛血清(Biowhittaker, Verviers,比利时)与100 U·毫升gydF4y2Ba−1gydF4y2Ba青霉素/ 100μg·毫升gydF4y2Ba−1gydF4y2Ba链霉素对collagen-coated烧瓶如前所述gydF4y2Ba1gydF4y2Ba。主支气管上皮细胞来自Lonza (Walkersville,医学博士,美国),低温贮藏1通过正常的人类支气管上皮细胞(NHBE)。在激素补充支气管上皮细胞培养生长培养基(Lonza),胶原蛋白/ fibronectin-coated烧瓶gydF4y2Ba17gydF4y2Ba通道2,用于实验。细胞被播种在50×10gydF4y2Ba3gydF4y2Ba细胞·毫升gydF4y2Ba−1gydF4y2Ba在盘子或75×10gydF4y2Ba3gydF4y2Ba细胞·数组gydF4y2Ba−1gydF4y2Ba电动细胞基质阻抗传感通性和融合发展。表皮生长因子受体抑制剂AG1478(σ,圣路易斯,密苏里州,美国),亚当抑制剂TAPI-2 (Calbiochem、圣地亚哥、钙、美国),和环己酰亚胺(CHX;σ)添加了60 - 120分钟前刺激与HDM(爱格里尔实验室、数控、美国),TGF-β(σ),杆激活肽或表皮生长因子(EGF);σ)。丝氨酸肽酶抑制剂AEBSF添加到螨虫提取为60分钟37°C。此外,主要来自支气管上皮细胞刷在哮喘患者培养和用于实验如前所述gydF4y2Ba16gydF4y2Ba。gydF4y2Ba
欧洲互通性系统委员会gydF4y2Ba
电特性的融合性的或受伤上皮测量使用电动欧洲互通性系统委员会的如前所述gydF4y2Ba18gydF4y2Ba。细胞粘附的测量是基于电阻/电容电流的变化应用于不同频率(应用生物物理学,特洛伊,纽约,美国)。细胞接种在75×10gydF4y2Ba3gydF4y2Ba细胞·好gydF4y2Ba−1gydF4y2Ba400年μL副本和电阻/电容测量在400和40000赫兹。伤害是由电穿孔用电压脉冲的5 V和40 kHz 30年代。gydF4y2Ba
Immunodetection通过免疫印迹gydF4y2Ba
总细胞溶解产物得到的再悬浮细胞Laemmli缓冲区。Immunodetection如前所述gydF4y2Ba1gydF4y2Ba使用anti-EGFR, anti-phospho-EGFR(圣克鲁斯生物技术、圣克鲁斯,CA), anti-phospho-extracellular-regulated激酶(ERK;细胞信号技术,英国),anti-occludin和anti-ZO-1(酶实验室、旧金山、钙、美国)。gydF4y2Ba
Immunofluorescent染色gydF4y2Ba
细胞生长与PBS / CaCl LabTeks被清洗gydF4y2Ba2gydF4y2Ba,固定在冰冷的丙酮(90%)为30分钟,阻止了PBS / 5%牛血清白蛋白为60分钟,60分钟孵化与初级抗体对表皮生长因子受体(1:200),钙粘蛋白(Santa Cruz)和ZO-1(酶)和随后孵化为60分钟FITC-labelled anti-rabbit (1:200;DAKO诊断,米西索加、加拿大)或Rhodamine-labelled anti-mouse免疫球蛋白G轭合物(1:400;杰克逊Immunoresearch实验室、西树林,PA,美国)和分析如前所述gydF4y2Ba19gydF4y2Ba。gydF4y2Ba
透射电子显微镜法gydF4y2Ba
细胞被固定在室温下1 h和隔夜PFA在4°C 2%, 2.5%戊二醛和0.1摩尔·LgydF4y2Ba−1gydF4y2Ba钠甲次砷酸盐,pH值7.4和准备TEM如前所述。TEM进行西奈山医院核心设施(多伦多,加拿大)。gydF4y2Ba
细胞表面生物素酰化gydF4y2Ba
细胞被洗了四次与包含1毫米MgCl PBSgydF4y2Ba2gydF4y2Ba和0.1毫米CaClgydF4y2Ba2gydF4y2Ba和1.5 mg·毫升孵化gydF4y2Ba−1gydF4y2Basulfosuccinimidyl 2 - (biotinamido) ethyl-dithioproprionate (sulfo-NHS-SS-biotin)两次20分钟,清洗与50 mM NH紧随其后gydF4y2Ba4gydF4y2Ba在PBS / MgCl ClgydF4y2Ba2gydF4y2Ba/ CaClgydF4y2Ba2gydF4y2Ba淬火免费sulfo-NHS-SS-biotin,紧随其后的是四个附加在PBS / MgCl耐洗gydF4y2Ba2gydF4y2Ba/ CaClgydF4y2Ba2gydF4y2Ba。细胞然后用冰冷的PBS包含1毫米原钒酸钠(NagydF4y2Ba3gydF4y2Ba签证官gydF4y2Ba4gydF4y2Ba200年)和1毫米PMSF和随后细胞溶解μL Triton裂解缓冲(30更易·LgydF4y2Ba−1gydF4y2Ba玫瑰(pH值7.4),100年更易·LgydF4y2Ba−1gydF4y2Ba生理盐水1更易·LgydF4y2Ba−1gydF4y2BaEGTA 20更易·LgydF4y2Ba−1gydF4y2Ba氟化钠,1% Triton x - 100 1更易·LgydF4y2Ba−1gydF4y2BaPMSF 20μl·毫升gydF4y2Ba−1gydF4y2Ba蛋白酶抑制鸡尾酒(BD生物科学、圣地亚哥、钙、美国),和1更易·LgydF4y2Ba−1gydF4y2BaNagydF4y2Ba3gydF4y2Ba签证官gydF4y2Ba4gydF4y2Ba在冰上20分钟。细胞溶解产物在10000×离心机gydF4y2BaggydF4y2Ba15分钟和孵化与链霉亲和素小珠收集生物素化的蛋白质。这之后,免疫复合物与裂解缓冲洗了三次。沉淀进行SDS 10%页面,在硝化纤维膜钙粘蛋白和表皮生长因子受体免疫印迹检测使用增强化学发光,这是执行根据制造商的指导方针。在所有情况下,膜沾着朱红色年代(σ),以确保平等的加载和转移的蛋白质。gydF4y2Ba
统计分析gydF4y2Ba
我们学生魏克森讯号等级测试用于成对观测之后,分析或Mann-Whitney紫外线测试分析的意义不同上皮电阻测量的条件。gydF4y2Ba
结果gydF4y2Ba
HDM诱发短期变化的上皮屏障的形成gydF4y2Ba
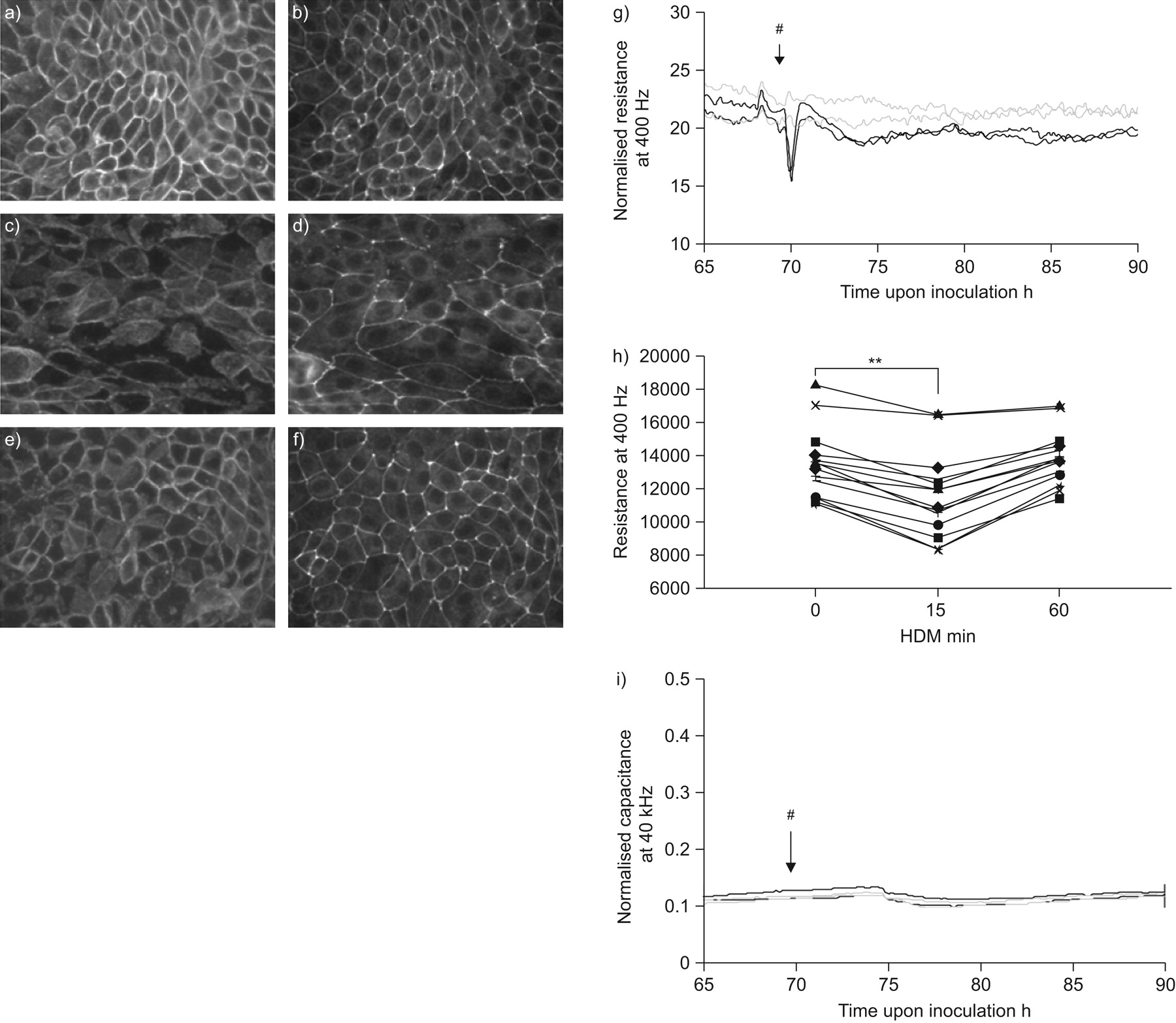
交界的损失与破坏细胞间粘附接触,观察哮喘,可能对气道上皮细胞功能有重要影响。因此,接触EMT的损伤是一个关键组成部分gydF4y2Ba20.gydF4y2Ba,这一过程可能导致过敏性哮喘气道上皮重建gydF4y2Ba11gydF4y2Ba,gydF4y2Ba12gydF4y2Ba。因此,我们旨在调查HDM对细胞间连接的影响。我们观察到ZO-1和钙粘紧的标志,粘合连接处并且分别是局部的细胞间连接在静止条件下,形成一个连续的环。HDM曝光后,钙粘蛋白采用了参差不齐的安排和ZO-1染色变得不连续偶尔完全丧失15分钟内相邻细胞边界(gydF4y2Ba图1 fgydF4y2Ba)。60分钟后,上皮屏障得以恢复,所观察到的re-localisation ZO-1和钙粘蛋白膜(gydF4y2Ba图1 fgydF4y2Ba)。gydF4y2Ba
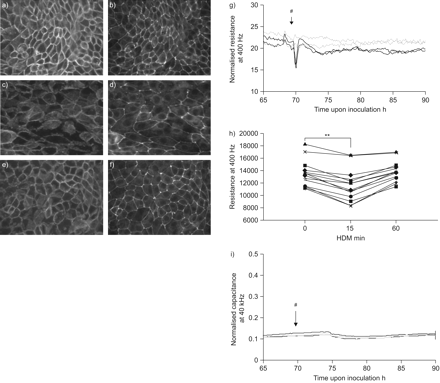
屋尘螨(HDM)诱发短期上皮的变化和信息联系。16 hbe细胞被播种在LabTeks或电动细胞基质阻抗传感通性数组副本,增长3 - 5天,serum-deprived一夜之间,HDM(μg·50毫升gydF4y2Ba−1gydF4y2Ba)或车辆(媒介)。a、b)控制、氟)细胞暴露于HDM c, d) 15分钟和e, f) 60分钟,c, e)钙粘蛋白和b, d, f)带occludens-1被immunofluorescent染色检测。代表三个独立的实验。g)电阻为400赫兹。正确的井间方差,阻力值正常化的起点发现电极。正常化的一个代表性的实验从60 - 85 h。HDM(#)添加在接种后∼70 h。车辆:- - - - -;HDM: - - - - -。h)绝对阻力值(t = 0)和前15和HDM曝光后60分钟。 n = 14. **: p<0.01. i) Capacitance was measured at 40 kHz using ECIS and normalised capacitance of a representative experiment from 60–85 h is shown. HDM (#) was added at ∼70 h upon inoculation. ––––: vehicle; ––––: HDM.
接下来我们测试了这些变化是否在屏障结构平行的屏障功能的变化。我们测量上皮阻力位在400赫兹(通过欧洲互通性系统委员会),因为低频电阻已经证明为屏障的变化最敏感的紧张gydF4y2Ba18gydF4y2Ba。相比之下,电容测量40 kHz是最敏感的参数监控细胞对矩阵的变化,细胞间接触形成相对不敏感gydF4y2Ba18gydF4y2Ba。我们观察到的HDM诱导迅速和大量低频电阻下降(∼20%)(gydF4y2Ba图1 g, hgydF4y2Ba),但不是高频电容(gydF4y2Ba图1我gydF4y2Ba),表明选择性破坏细胞间的粘连gydF4y2Ba18gydF4y2Ba。这种效果是暂时的:阻力回到原来的值在∼1 h,伴随的re-localisation ZO-1和钙粘蛋白膜。值得注意的是在几个小时内,第二个趋势平缓,持久减少阻力可以观察到HDM-treated (p = 0.07gydF4y2Ba与gydF4y2Ba控制值)。gydF4y2Ba
HDM蛋白酶依赖的方式激活表皮生长因子受体导致钙粘蛋白的重新分配和delocalisation ZO-1gydF4y2Ba
解开HDM-induced接触损伤的机制,我们认为表皮生长因子受体信号可能导致这种类型的损伤,基于我们之前的发现表明EGFR活动起着至关重要的作用在支气管哮喘上皮HDM-induced信号gydF4y2Ba16gydF4y2Ba。事实上,我们观察到HDM激活表皮生长因子受体下游信号,当检测到水平的提高phospho-EGFR(酪氨酸1173)和phospho-ERK,最大的影响在HDM曝光(5 - 20分钟gydF4y2Ba图2一个gydF4y2Ba)。这次课程是按照瞬态屏障功能下降。这可能涉及到杆的激活,导致ADAM17-dependent heparin-bound-EGF脱落gydF4y2Ba16gydF4y2Ba,gydF4y2Ba21gydF4y2Ba- - - - - -gydF4y2Ba23gydF4y2Ba或TGF-αgydF4y2Ba24gydF4y2Ba。过敏原Der p 1是已知的作为半胱氨酸肽酶,而Der 3 p, Der p 6和Der 9含有丝氨酸蛋白酶活性。值得注意的是,丝氨酸蛋白酶在HDM提取物已被证明激活杆gydF4y2Ba22gydF4y2Ba。我们使用的提取是proteolytically活跃,可检测的丝氨酸,但不是半胱氨酸肽酶活动(数据未显示)。感兴趣的注意,heat-inactivation,以及治疗与丝氨酸和半胱氨酸蛋白酶抑制剂,我们仍然发现大量的蛋白水解活性水平(数据没有显示)。这表明,另外,heat-insensitive然而定义蛋白酶也可能导致HDM的效果。因此,HDM-induced EGFR磷酸化丝氨酸蛋白酶抑制剂预处理后反应较AEBSF,表明这是至少在一定程度上依赖于丝氨酸蛋白酶活动(gydF4y2Ba图2一个gydF4y2Ba)。评估是否丝氨酸protease-dependent / PAR2-mediated机制可能涉及,我们使用PAR2-activating肽(PAR2-ap)和phospho-EGFR观察到类似的增加。这取决于亚当活动,因为它被亚当抑制剂TAPI-2 (gydF4y2Ba图2 bgydF4y2Ba)。此外,杆下游信号依赖于表皮生长因子受体,PAR2-ap-induced phospho-ERK-1/2反应是抑制表皮生长因子受体抑制剂AG1478 (gydF4y2Ba图2 bgydF4y2Ba)。此外,PAR2-ap和EGF可以模仿HDM-induced钙粘蛋白的再分配。符合这一点,AEBSF和AG1478封锁了HDM-induced钙粘蛋白再分配(gydF4y2Ba图2 cgydF4y2Ba)。TAPI-2产生了类似的效果(数据没有显示)。这些结果说明参与的蛋白酶/杆/亚当/ EGFR-dependent通路HDM-induced动荡的钙粘蛋白。发现进一步支持ADAM-dependent HDM脱落的表皮生长因子受体配体,我们使用中和α-EGFR阻止ligand-dependent受体的激活。我们观察到HDM不再能够增加EGFR磷酸化的α-EGFR (gydF4y2Ba图2 cgydF4y2Ba)。此外,AG1478 ZO-1的膜本地化HDM暴露增加(gydF4y2Ba图2 dgydF4y2Ba)。这是反映上皮屏障功能的变化;AG1478显著降低(虽然不能完全阻止)HDM-induced上皮抵抗力下降(gydF4y2Ba图2 e, fgydF4y2Ba)。gydF4y2Ba
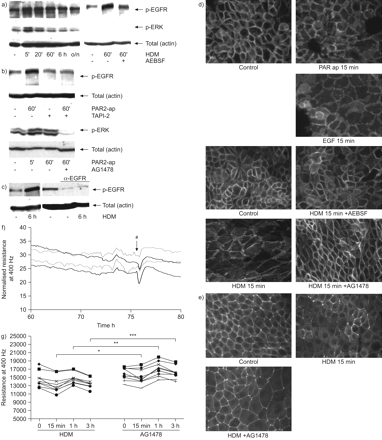
Protease-dependent激活表皮生长因子受体(EGFR)屋尘螨(HDM)是参与细胞间联系的中断。16 hbe细胞生长3 - 5天24-well盘子,LabTeks或电动细胞基质阻抗传感阵列,血清剥夺一夜之间,有或没有孵化表皮生长因子受体抑制剂AG1478(1μM) TAPI-2(22.5μM), AEBSF(1海里)或α-EGFR(1μg·毫升gydF4y2Ba−1gydF4y2Ba60 - 120分钟),暴露于HDM(μg·50毫升gydF4y2Ba−1gydF4y2Ba),protease-activated受体2激活肽(PAR2-ap;50μM)或表皮生长因子(EGF;10 ng·毫升gydF4y2Ba−1gydF4y2Ba)表示。两者总细胞溶解产物是准备和phospho-extracellular-regulated激酶(ERK)或phospho-EGFR被免疫印迹检测(箭头所指)。β-actin被用作控制加载。代表三个独立的实验。d)钙粘蛋白和e)带occludens-1被immunofluorescent染色检测。代表三个独立的实验。f)正常阻力的实验从60 - 70 h代表。HDM(#)添加∼75 h后接种。AG1478: - - - - -;控制:- - - - - -。g)绝对阻力值(t = 0)和前15分钟,1 h和3 h后HDM暴露所示。 n = 9. *: p<0.05; **: p<0.01; ***: p<0.001. Resistance was measured at 400 Hz. AG1478 (1 μM) was added 2 h prior to the addition of HDM/vehicle.
表皮生长因子受体参与上皮重组时通过电穿孔伤gydF4y2Ba
类似于HDM-induced受伤,我们观察到表皮生长因子受体抑制改进上皮重建在电穿孔伤。在这种情况下,一个相对小的区域受伤,上皮完整性预计将在几小时内恢复,gydF4y2Ba即。gydF4y2Ba通过细胞迁移/传播,而不是扩散。单层一旦恢复,电阻可能进一步增加由于恢复细胞间接触。的确,细胞添满2 h内电极,观察到稳定的高频电容(gydF4y2Ba图3 bgydF4y2Ba)。随后,阻力水平慢慢的继续增加,表明细胞间联系的形成(gydF4y2Ba图3gydF4y2Ba)。AG1478明显延迟2 h内电极的重新受伤后,19±7%gydF4y2Ba与gydF4y2Ba增加一个51±10%。这种延迟也观察高频电容(gydF4y2Ba图3 bgydF4y2Ba),可能是由于抑制细胞/迁移扩散。相反,AG1478增加了复苏的上皮屏障2 h后,所反映的一个额外的42.2±9%增加阻力gydF4y2Ba与gydF4y2Ba10±12%的增长在其缺席(n = 4)。添加EGF受伤前有一个相反的效果;它促进了迁移阶段(gydF4y2Ba图3 c, dgydF4y2Ba),但受损上皮屏障的恢复(gydF4y2Ba图3 cgydF4y2Ba),这表明EGFR活动促进迁移阶段和抑制上皮再生期间接触重组。gydF4y2Ba
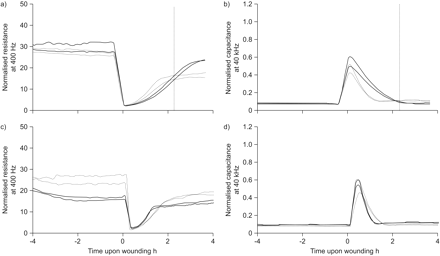
抑制表皮生长因子受体(EGF)活动增加传播/附件,降低细胞间联系的形成在通过电穿孔伤。16 hbe细胞被播种在重复电动细胞基质阻抗传感通性数组,增长了3天,serum-deprived一夜之间,预处理(- - -)或没有(·······),b) AG1478(1μM)或c, d) EGF (10 ng·毫升gydF4y2Ba−1gydF4y2Ba电穿孔)6 - 8 h和受伤。a, c)电阻为400 Hz和b, d)电容测量使用欧洲互通性系统委员会40 kHz。正常化的电阻和电容代表实验显示4 h受伤前4 h后受伤。gydF4y2Ba
HDM诱发EGFR-independent乳沟交界的蛋白质gydF4y2Ba
最初HDM-induced阻力下降,出现部分独立于表皮生长因子受体,可能造成的短期变化紧密连接密封。除了ZO-1再分配,退化的紧密连接蛋白可能参与其中gydF4y2Ba25gydF4y2Ba,gydF4y2Ba26gydF4y2Ba。我们调查是否HDM影响额外的——和/或细胞内紧密连接组件,gydF4y2Ba如。gydF4y2Baoccludin ZO-1,通过这样的机制。的确,HDM诱导部分occludin的乳沟,减少其80 kD的增长表明过度磷酸化形式gydF4y2Ba27gydF4y2Ba较小的乳沟和外观的片段(45∼∼30 kD)。我们还观察到部分乳沟ZO-1 HDM暴露成更小的产品(192∼∼165 kD)。正如所料,AG1478不能防止ZO-1或occludin的存在退化(gydF4y2Ba图4gydF4y2Ba)。此外,无论是蛋白质的乳沟是预防AEBSF(数据未显示),建议其他蛋白水解酶的参与活动或HDM-activated蛋白酶。除了丝氨酸肽酶活动,提取含有几丁质酶活性(∼20 U·μLgydF4y2Ba−1gydF4y2Baβ-N-acetylglucosaminidase),但目前未知occludin能否作为几丁质酶的底物。HDM提取物通常含有内毒素。然而,内毒素水平在我们的提取是较低的(gydF4y2Ba即。gydF4y2Ba0.5欧盟·μLgydF4y2Ba−1gydF4y2Ba,由鲎变形细胞溶解产物内毒素测试,渲染最终欧盟·2.5毫升的浓度gydF4y2Ba−1gydF4y2Ba在我们的细胞)。脂多糖含量高达50000欧盟·毫升gydF4y2Ba−1gydF4y2Ba并不影响上皮完整性(gydF4y2Ba图4 bgydF4y2Ba)。因此,参与类毒素似乎不太可能。gydF4y2Ba
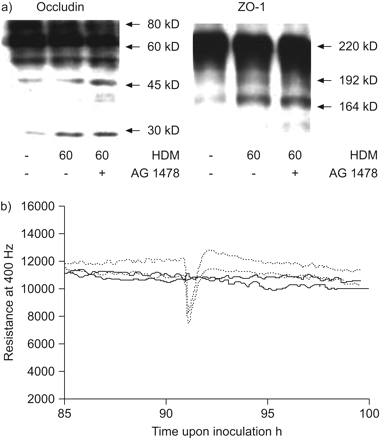
屋尘螨(HDM)诱发短期乳沟的TJ蛋白表皮生长因子receptor-independent方式。16 hbe细胞3 - 5天24-well盘子,serum-deprived一夜之间,HDM(μg·50毫升gydF4y2Ba−1gydF4y2Ba)60分钟。AG1478(1μM)添加2 h与HDM刺激之前。全长occludin (60 - 80 kD),带occludens-1 (ZO-1;220 kD)和乳沟产品被免疫印迹检测,用箭头表示。b)暴露在脂多糖(LPS);500年欧盟·10gydF4y2Ba−1gydF4y2BaμL)不减少上皮屏障功能。16 hbe细胞生长3 - 5天在电动细胞基质阻抗数组,血清剥夺了一夜之间和暴露于HDM(μg·50毫升gydF4y2Ba−1gydF4y2Ba;·····)或有限合伙人(——)。电阻测量在400赫兹。电阻的实验显示了从85 - 100 h代表。HDM和LPS被添加在接种∼90 h。gydF4y2Ba
信号通过表皮生长因子受体和随后的变化和信息接触形成推动了TGF-β-primed上皮gydF4y2Ba
在一起,上面的数据显示,信号抑制表皮生长因子受体信号,gydF4y2Ba如。gydF4y2Baligand-induced EGFR掩饰/溶酶体降解gydF4y2Ba28gydF4y2Ba从受伤,可能会改善上皮恢复。相反,持续的表皮生长因子受体信号可能会减弱恢复上皮屏障。因为我们之前的研究结果表明,TGF-β延长ligand-induced表皮生长因子受体激活gydF4y2Ba11gydF4y2Ba,我们检查TGF-β是否能够推迟终止EGFR信号,从而对上皮屏障功能的影响。我们首先研究了TGF-β是否改变了表皮生长因子受体的细胞命运通过内吞作用和退化的影响。长期治疗与EGF(6小时)的差别导致了对这些表皮生长因子受体(gydF4y2Ba图5gydF4y2Ba)。重要的是,TGF-β预处理大大减轻这个(gydF4y2Ba图5gydF4y2Ba),而TGF-β就没有这样的效果,甚至增加表皮生长因子受体的蛋白表达(gydF4y2Ba图5 bgydF4y2Ba)。当新的蛋白质合成受到抑制CHX,暴露引起的细胞EGF表皮生长因子受体表达的急剧减少,揭露健壮EGF-promoted受体退化。再次,预处理TGF-β几乎完全废除了EGF-induced表皮生长因子受体退化(gydF4y2Ba图5度gydF4y2Ba)。此外,EGF造成完全掩饰的表皮生长因子受体为endosomal水泡隔间。TGF-β-treated细胞,除了水泡积累,不同的细胞质和外围EGFR标签仍然是可见的。这也是证明了更高水平的生物素化的表皮生长因子受体在细胞表面gydF4y2Ba与gydF4y2BaEGF治疗(gydF4y2Ba图5 d, egydF4y2Ba)。在一起,这些发现表明,TGF-β可能有助于维护的表皮生长因子受体移植EGFR表达,减少表皮生长因子受体掩饰和防止退化性表皮生长因子受体。gydF4y2Ba
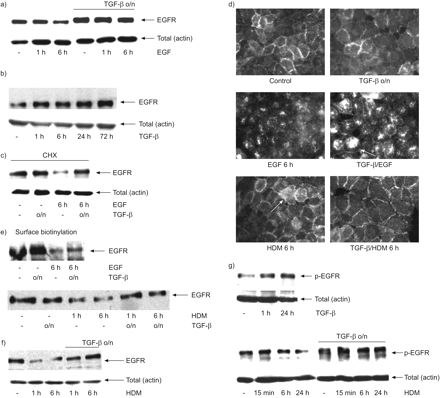
转化生长因子(TGF) -β防止表皮生长因子受体(EGFR)降解,减少表皮生长因子受体的内吞作用,延长屋尘螨(HDM)全身表皮生长因子受体信号。16 hbe细胞生长24-well盘子或LabTeks 3天,血清剥夺4 h,预处理的一夜之间有或没有TGF-βng(2毫升gydF4y2Ba−1gydF4y2Bang)和随后刺激TGF-β(2毫升gydF4y2Ba−1gydF4y2Ba)、表皮生长因子(EGF);10 ng·毫升gydF4y2Ba−1gydF4y2Ba)或HDM(μg·50毫升gydF4y2Ba−1gydF4y2Ba)表示,在和无环己酰亚胺(CHX;μg·10毫升gydF4y2Ba−1gydF4y2Ba)。两者总细胞溶解产物是准备和EGFR检测到免疫印迹(箭头所指)。β-actin被用作控制加载。代表三个独立的实验。d)细胞生长在LabTeks和表皮生长因子受体被immunofluorescent染色检测。表皮生长因子受体在细胞膜局部是箭头所示。e)生物素化的细胞表面和细胞溶解。使用链霉亲和素生物素化的蛋白质被找到珠子和sds - page分析。表皮生长因子受体被免疫印迹检测(箭头所指)。f, g)总细胞溶解产物制备和表皮生长因子受体,phospho-EGFR检测到免疫印迹(箭头所指)。 β-actin was used as a control for equal loading. Representatives of three independent experiments are shown.
符合磷酸化在HDM暴露时间延长,降低HDM EGFR表达的差别也引起对这些(gydF4y2Ba图5 fgydF4y2Ba)和表皮生长因子受体引起掩饰,这是预防TGF-β预处理(gydF4y2Ba图5 d, egydF4y2Ba)。TGF-βEGFR磷酸化增加(gydF4y2Ba图5克gydF4y2Ba),基底和TGF-β-induced EGFR磷酸化被HDM进一步增强,产生影响,持续长时间TGF-β-treated细胞(gydF4y2Ba图5克gydF4y2Ba)。gydF4y2Ba
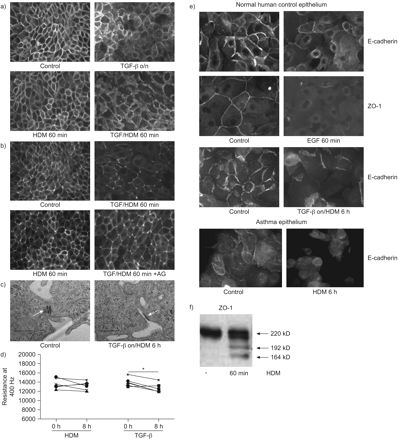
接下来我们测试持续的表皮生长因子受体信号在TGF-β治疗的后果。钙粘蛋白本地化(TGF-β本身没有重大影响gydF4y2Ba图6 a, bgydF4y2Ba),然而,它延长了HDM-induced delocalisation钙粘蛋白以及ZO-1,后仍然可以观察到60分钟(gydF4y2Ba图6 a, bgydF4y2Ba)。AG1478 ZO-1封锁了再分配(gydF4y2Ba图6gydF4y2Ba)。此外,电子显微镜分析发现主要减少细胞间紧密连接在细胞暴露于TGF-β和HDMgydF4y2Ba与gydF4y2Ba控制细胞(gydF4y2Ba图6 cgydF4y2Ba)。因此,虽然独自TGF-β没有明显影响上皮阻力,减少HDM-induced上皮抵抗被TGF-β强,成为重要的经过长时间的接触(如图所示的8小时时间点)(gydF4y2Ba图6 dgydF4y2Ba)。gydF4y2Ba
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
转化生长因子(TGF) -β预处理延长屋尘螨(HDM)全身上皮连接中断。a、b)在LabTeks 16 hbe细胞培养3天,血清剥夺4 h,预处理的一夜之间有或没有TGF-βng(2毫升gydF4y2Ba−1gydF4y2Ba),随后刺激HDM(μg·50毫升gydF4y2Ba−1gydF4y2Ba)表示,在存在和缺乏AG1478(1μM)。a)上皮型和b)带occludens(佐薇)1被immunofluorescent染色检测。代表三个独立的实验。c) 16 hbe细胞种植在封面3天,血清剥夺4 h,预处理的一夜之间有或没有TGF-βng(2毫升gydF4y2Ba−1gydF4y2Ba),随后刺激HDM(μg·50毫升gydF4y2Ba−1gydF4y2Ba)6 h。电镜分析表明执行紧密连接(白色箭头)。代表电子显微镜图像显示。d)细胞生长在电动细胞基质阻抗传感通性数组的副本3天,血清剥夺4 h,预处理的一夜之间有或没有TGF-βng(2毫升gydF4y2Ba−1gydF4y2Ba),随后刺激HDM(μg·50毫升gydF4y2Ba−1gydF4y2Ba)/车辆。电阻测量在0和8 h后使用欧洲互通性系统委员会在400 Hz HDM治疗。n = 6。*:p < 0.05。中间值表示。正常的主支气管上皮细胞获得Lonza (Walkersville,医学博士,美国)e)和上皮细胞来自支气管哮喘刷牙gydF4y2Ba16gydF4y2Ba成长为融合在LabTeks 3 - 5天,生长因子/ hormone-deprived 4 h,预处理的一夜之间有或没有TGF-β(2 ng·毫升gydF4y2Ba−1gydF4y2Ba),随后刺激HDM(μg·50毫升gydF4y2Ba−1gydF4y2Ba)或表皮生长因子表示。钙粘蛋白和ZO-1被immunofluorescent染色检测。代表三个独立的实验。f)总细胞溶解产物从支气管哮喘上皮细胞被准备gydF4y2Ba16gydF4y2Ba和ZO-1检测到蛋白免疫印迹(箭头所指)。代表三个独立的实验。gydF4y2Ba
最后,为了测试我们的研究结果的相关性,我们研究NHBE。EGF诱导钙粘蛋白和ZO-1 delocalisation NHBE细胞和HDM + TGF-β导致长时间的组合联接的钙粘蛋白的损失,而HDM仅在6 h(不产生明显影响gydF4y2Ba图6 egydF4y2Ba)。此外,我们发现单独接触HDM诱导持续delocalisation在支气管哮喘上皮钙粘蛋白(gydF4y2Ba图6 egydF4y2Ba)。符合16 hbe的影响(gydF4y2Ba图4gydF4y2Ba)和NHBE(未显示),HDM导致哮喘上皮(ZO-1乳沟gydF4y2Ba图6 fgydF4y2Ba)。gydF4y2Ba
讨论gydF4y2Ba
上皮连接已成为过敏敏化作用的重要目标。他们的崩溃可能促进过敏原的运输,促进上皮细胞的炎性反应gydF4y2Ba1gydF4y2Ba基底和允许访问的生长因子受体gydF4y2Ba29日gydF4y2Ba。re-epithelisation这可能为正常,但也可能导致异常的维修改造和正在进行的组织。因此,有效恢复紧密连接在损伤可能是患有过敏性哮喘的关键。gydF4y2Ba
此前,HDM被描述上皮通透性增加gydF4y2Ba30.gydF4y2Ba。在这里,我们表明,这可能是由于一个暂时的失去上皮细胞连接,这可能是由HDM-induced的联接的蛋白质,再分配gydF4y2Ba如。gydF4y2Ba钙粘蛋白和ZO-1以及紧密连接蛋白的蛋白水解降解,随后快速re-localisation这些分子的膜。底层机制:我们的研究结果也提供了洞察HDM诱导表皮生长因子受体激活丝氨酸蛋白酶/ PAR2-dependent态度,这是至关重要的细胞间连接的随后的瞬态干扰。如前所述,这可能涉及到EGFR-induced酪氨酸磷酸化和delocalisation联接的蛋白质,以及减毒的重新分配这些蛋白上皮连接gydF4y2Ba31日gydF4y2Ba- - - - - -gydF4y2Ba35gydF4y2Ba。抑制表皮生长因子受体减少上皮的拆卸连接和随后的迁移/传播,和改进的屏障功能的恢复HDM-induced受伤和受伤。此外,HDM交界完整性通过EGFR-independent机制的影响,导致ZO-1和occludin的乳沟。丝氨酸肽酶活动的抑制作用并没有阻止乳沟TJ分子,建议其他酶的参与活动,包含在HDM (gydF4y2Ba如。gydF4y2Ba几丁质酶)或细胞间蛋白酶的诱导。后者可能导致ZO-1退化,因为这是一个细胞内的过程。感兴趣的注意,甲壳素和微生物葡萄糖结构出现在HDM提取可以激活c型凝集素受体gydF4y2Ba36gydF4y2Ba,导致钙gydF4y2Ba2 +gydF4y2Ba通量gydF4y2Ba37gydF4y2Ba,这可能会导致钙蛋白酶的激活和随后的乳沟交界的蛋白质gydF4y2Ba38gydF4y2Ba。符合这一点,HDM过敏原可以引起CagydF4y2Ba2 +gydF4y2Ba在气道上皮细胞涌入gydF4y2Ba39gydF4y2Ba。虽然需要进一步的调查,我们发现钙离子载体诱导的再分配和裂解ZO-1(数据没有显示)。gydF4y2Ba
鉴于AG1478对钙粘蛋白分布的影响我们推测,持续的表皮生长因子受体活动可能影响上皮屏障功能,因为被认为粘合连接处并且提供适当的组装所需的架构紧密连接。我们已经表明,表皮生长因子受体活动增加/长期的预处理与TGF-β上皮细胞,纤维增生细胞因子水平升高在哮喘气道gydF4y2Ba13gydF4y2Ba。TGF-β似乎通过upregulation EGFR的表达,以及部分预防ligand-induced退化和表皮生长因子受体的内吞作用。结合,TGF-βHDM-provoked EGFR信号增加,导致持续HDM-induced接触损伤。这可能会提供一个可能的解释不充分修复上皮屏障功能的观察到哮喘、HDM暴露可能发生在的存在增加了TGF-β水平gydF4y2Ba13gydF4y2Ba。后者可能与TGF-β启动子多态性与哮喘有关gydF4y2Ba40gydF4y2Ba。值得注意的是,HDM暴露修改的影响gydF4y2BaTGF -gydF4y2Baβ单核苷酸多态性在儿童气道高反应性gydF4y2Ba41gydF4y2Ba。此外,表皮生长因子受体活动增加可能与哮喘相关的异常的表皮生长因子受体活动/表达式在哮喘气道gydF4y2Ba13gydF4y2Ba,gydF4y2Ba15gydF4y2Ba。异常的表皮生长因子受体的活动的重要性,进一步强调了CA-repeat表皮生长因子受体基因多态性,有关存在和哮喘的严重程度gydF4y2Ba42gydF4y2Ba。重要的是,我们的研究表明,HDM诱发哮喘长期粘附delocalisation上皮,表明我们的数据可能确实是哮喘的相关性。这是我们初步的组织学研究结果进一步支持在哮喘病人(数据未发表),我们观察到delocalisation支气管上皮钙粘蛋白的。在未来的研究中,我们旨在执行更扩展比较健康和哮喘。钙粘蛋白的持续交界损失增加/长期的表皮生长因子受体的激活可能有重要的影响,因为它可能导致上皮改造,gydF4y2Ba如。gydF4y2Ba由EMTgydF4y2Ba11gydF4y2Ba。此外,TGF-β和HDM-induced上皮路口的损失可以促进生长因子的表达以及pro-allergic因素gydF4y2Ba1gydF4y2Ba,gydF4y2Ba16gydF4y2Ba气道上皮细胞。此外,中断的障碍可以通过激活促进气道炎症的上皮细胞因子释放炎性细胞在气道腔与受体限于上皮的基底外侧表面。gydF4y2Ba
总之,我们已经表明,HDM诱发临时减少障碍函数,这是持续促进表皮生长因子受体信号时,gydF4y2Ba如。gydF4y2BaTGF-β的存在。因此,尽管促进迁移/传播,增加表皮生长因子受体信号可能会减少上皮的完整性,影响上皮修复和增强的脆弱性HDM的上皮细胞。gydF4y2Ba
确认gydF4y2Ba
我们要感谢美国邮政(实验室的变态反应学和肺疾病,病理生物学和医学部门,格罗宁根大学医学中心,格罗宁根,荷兰)进行酶活性测定螨提取。gydF4y2Ba
脚注gydF4y2Ba
支持声明gydF4y2Ba
这项研究是由来自荷兰哮喘基金会的资助(I.H. Heijink, 3.2.05.039)和肾脏基金会的资助和加拿大的加拿大国家科学与工程研究委员会(a . Kapus)。gydF4y2Ba
感兴趣的语句gydF4y2Ba
没有宣布。gydF4y2Ba
- 收到了gydF4y2Ba2009年8月6日。gydF4y2Ba
- 接受gydF4y2Ba2010年3月15日。gydF4y2Ba
- ©2010人队gydF4y2Ba
引用gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba










