





文摘gydF4y2Ba
背景gydF4y2BaCOVID-19-induced死亡率主要发生在老年患者。在这些患者中几个immunomodulating疗法似乎不那么有益。这些观点背后的生物基质是未知的。本研究的目的是为了获得洞察老化之间的关系,宿主反应,COVID-19患者死亡率。gydF4y2Ba
方法gydF4y2Ba我们确定43生物标志物反映四个病理生理改变的领域:内皮细胞激活和凝固,炎症和器官损伤,细胞因子和趋化因子的释放。我们使用中介分析副ageing-driven改变宿主反应与30天死亡率。生物标志物与老化和死亡率都在重症监护室和外部群组验证。gydF4y2Ba
结果gydF4y2Ba464年普通病房COVID-19患者根据年龄分层几十年。年龄增加30天死亡率是一个独立的危险因素。老化与改变有关的每个主机响应域,以更大的激活内皮和凝固系统和更强的海拔炎症和器官损伤标记,这是独立的与年龄相关的并发症的增加。可溶性肿瘤坏死因子受体1,可溶性在髓细胞触发受体表达1,可溶性thrombomodulin显示相关性最强的老化和解释的一部分ageing-driven增加30天死亡率(分别介导比例:13.0%,12.9%和12.6%)。gydF4y2Ba
结论gydF4y2Ba老龄化与一个强大和广泛的修改主机响应COVID-19和特异性免疫的变化可能有助于在老年患者死亡率增加。这些结果可以提供洞察潜在COVID-19特异性免疫调节的目标。gydF4y2Ba
介绍gydF4y2Ba
从一开始的COVID-19大流行,流行病学数据显示,COVID-19-related死亡率急剧增加与老gydF4y2Ba1gydF4y2Ba- - - - - -gydF4y2Ba3gydF4y2Ba]。在荷兰,50岁及更年轻的个体仅占∼COVID-19死亡病例的1%,而个人年龄≥70∼占89%的这些死亡(gydF4y2Ba4gydF4y2Ba]。这背后的生物基质目前未知的急剧上升。摩根大通并发症调解只有一小部分的死亡率增加gydF4y2Ba2gydF4y2Ba,gydF4y2Ba5gydF4y2Ba]。gydF4y2Ba
COVID-19与炎症标记和细胞因子的浓度增加,凝血障碍,和endotheliopathygydF4y2Ba6gydF4y2Ba,gydF4y2Ba7gydF4y2Ba]。多个随机对照试验(相关的),试图改善这些主机响应干扰报告改善临床结果(gydF4y2Ba8gydF4y2Ba]。然而,在最大相关,两个当前COVID-19住院病人的治疗基础,即地塞米松和叫小最古老的患者(如果有任何有益的效果gydF4y2Ba9gydF4y2Ba- - - - - -gydF4y2Ba11gydF4y2Ba]。一般来说,老龄化与先天和适应性免疫的变化联系在一起;在这种背景下,“inflammageing”是指持续慢性炎症状态,而“免疫衰老”是指逐渐下降的能力产生有效的免疫反应的新抗原(gydF4y2Ba12gydF4y2Ba]。尽管老龄化对免疫系统的影响涉及COVID-19在老年病人的发病机制gydF4y2Ba13gydF4y2Ba,gydF4y2Ba14gydF4y2Ba),研究分析老年人对主机的影响反应干扰COVID-19稀缺。以前的研究集中在有限数量的患者血浆细胞因子水平在不同疾病的严重性,显示出衰老协会增加炎症反应(gydF4y2Ba15gydF4y2Ba- - - - - -gydF4y2Ba18gydF4y2Ba]。老龄化的影响在COVID-19内皮和混凝反应还有待评估。探索age-driven这些关键主机响应域的差异及其与ageing-driven死亡率COVID-19可能为新immunomodulating目标识别奠定基础。gydF4y2Ba
本研究的主要目的是了解关键病理生理年龄和畸变通路之间的关系涉及COVID-19承认综合医院病房的病人(gydF4y2Ba即。gydF4y2Ba非关键生病)。为此,我们决定43生物标志物反映内皮的改变和凝固反应,炎症和器官损伤,细胞因子和趋化因子的释放。我们的次要目标是将主机响应畸变在老年患者中发现与这个年龄段的30天死亡率增加。gydF4y2Ba
方法gydF4y2Ba
研究设计和人口gydF4y2Ba
数据来源于ELDER-BIOME研究(ClinicalTrials.gov标识符gydF4y2BaNCT02928367gydF4y2Ba)和阿姆斯特丹大学医疗中心(阿姆斯特丹联电)COVID-19生物研究(AUMC 2020 _065)(有关详细信息,请参阅补充方法)。这两项研究是由阿姆斯特丹联电伦理委员会批准。ELDER-BIOME在阿姆斯特丹是一个前瞻性研究联电(这两个位置:sccp学术医疗中心(AMC)和医疗中心(VUMC),阿姆斯特丹,荷兰)和Flevo医院,Almere,荷兰gydF4y2Ba7gydF4y2Ba]。阿姆斯特丹联电COVID-19生物研究AMC和VUMC进行,根据临床护理的剩下的血液是等离子体处理存储。书面知情同意了所有的病人或其法定代表人。在AUMC生物研究中,一个附加选项延期同意使用一个可选的方法是实现无行为能力的病人。我们包括患者,18岁以上COVID-19-related症状和积极SARS-CoV-2 PCR或疑似COVID-19 CORADS-CT评分≥4反映了高(4)或(5)非常高怀疑肺介入COVID-19 [gydF4y2Ba19gydF4y2Ba]。包容是2020年2月到2021年9月完成的。普通病房患者样本48 h内承认(或-在一个单独的队列重症监护室(ICU)录取)。患者划分根据四个“波”(见文献[gydF4y2Ba20.gydF4y2Ba),和补充方法)。生物标志物在外部队列数据验证;详细信息请参阅在线补充和裁判gydF4y2Ba21gydF4y2Ba]。gydF4y2Ba图1gydF4y2Ba描述了COVID-19群组的概览。此外,死亡率数据也从普通病房的病人是因为SARS-CoV-2以外的其他病原体引起的社区获得性肺炎;详细信息请参阅在线补充和裁判gydF4y2Ba7gydF4y2Ba]。gydF4y2Ba
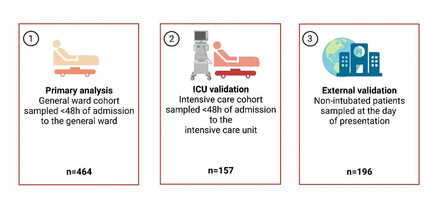
COVID-19群组的概述。gydF4y2Ba描述gydF4y2Ba图1gydF4y2Ba:gydF4y2Ba概述COVID-19测量等离子体生物标志物的军团。外部验证队列是源自一个公开的数据集的non-intubated COVID-19血浆蛋白的病人被Olink测量距离扩展试验(gydF4y2Ba21gydF4y2Ba]。gydF4y2Ba
化验gydF4y2Ba
43主机响应生物标志物以EDTA实际上等离子使用Luminex(研发、美国)和Bio-Plex 200系统(Bio-Rad实验室Inc .,大力神,CA)(见补充资料选择理由)。生物标志物是分层分为四个病理生理领域(见gydF4y2Ba表S1gydF4y2Ba详情)。参考价值从10灌木丛生的,19岁,sex-matched非传染性门诊所控制(ClinicalTrials.gov标识符gydF4y2BaNCT02928367gydF4y2Ba)。gydF4y2Ba
统计数据gydF4y2Ba
患者根据年龄分层几十年:< 50,≥50 - < < 70,≥60岁,≥60 - 70年。这个方法被选为促进可视化和临床解释。30天的生存差异被kaplan meier曲线呈现。生物标志物数据进行对数转换的。整体年龄差异几十年在生物标记在一个主机响应域得到了主成分分析(PCA),如前所述[gydF4y2Ba7gydF4y2Ba]。之间的差异主成分得分(PC)年龄段进行分析,方差分析(方差分析)。PC的相关性分数与衰老在连续的规模是由斯皮尔曼相关测试。gydF4y2Ba
几十岁个体生物标志物水平差异量化使用对冲的gydF4y2BaggydF4y2Ba效果和视觉效果的热图(gydF4y2Ba22gydF4y2Ba]。此外,回归分析中,年龄是模仿作为连续变量。分析了协会的力量使用枪兵的相关性。在一个单独的分析,个人与衰老生物标志物的协会也探索了住进加护病房时。gydF4y2Ba
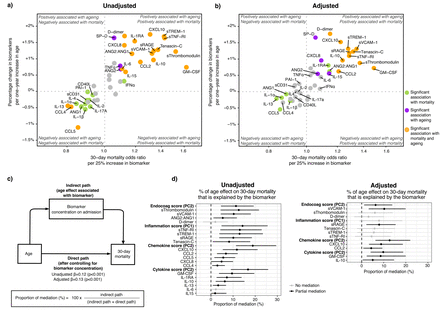
我们调查如果age-driven改变生物标志物浓度与age-driven增加30天死亡率有关中介分析(gydF4y2Ba5gydF4y2Ba,gydF4y2Ba23gydF4y2Ba]。生物标志物或PC分数需要老化和30天死亡率显著相关进入中介分析(见补充假设方法和细节)gydF4y2Ba24gydF4y2Ba]。生物标志物与老化和死亡率也在一个独立的外部验证群必然要196住院non-intubated COVID-19患者队列描述)(见补充方法(gydF4y2Ba图1gydF4y2Ba)[gydF4y2Ba21gydF4y2Ba]。最后,我们在≥70岁的患者进行了聚类分析来评估它们的宿主反应的均匀性,使用病房的方法(gydF4y2Ba25gydF4y2Ba]。此外,我们研究了生物标志物的中介效应是否影响病人的主机响应表型。gydF4y2Ba
回归和中介模型包括未经调整和调整方法。调整模型评估老化协会协变量包含生物标志物浓度与老化或COVID-19保健和包括:人口统计数据(包括医院、性别和包含波),老年性并存病(高血压、糖尿病、恶性肿瘤、免疫抑制和慢性心脏,神经系统,呼吸系统,和肾脏疾病),年龄,入学前和biomarker-related慢性药物(抗血小板和抗凝药物),和COVID-19-related immunomodulating治疗前抽样(皮质类固醇,anti-IL-6,伊马替尼)。当评估协会的老化与死亡,我们使用相同的协变量调整模型中的+反是;这可能会影响死亡率:immunomodulating治疗入院时但经过采样和抗生素的使用和remdesivir(有关详细信息,请参阅补充方法和假设测试)。gydF4y2Ba
细节描述missingness missingness和处理的补充方法和补充Excel文件(表1),gydF4y2Ba表S2和S3gydF4y2Ba。假定值的分析多个测试修正使用Benjamini-Hochberg (BH)过程。一个BH-adjusted p值< 0.05被认为是具有统计学意义。gydF4y2Ba
结果gydF4y2Ba
病人,表现和结果gydF4y2Ba
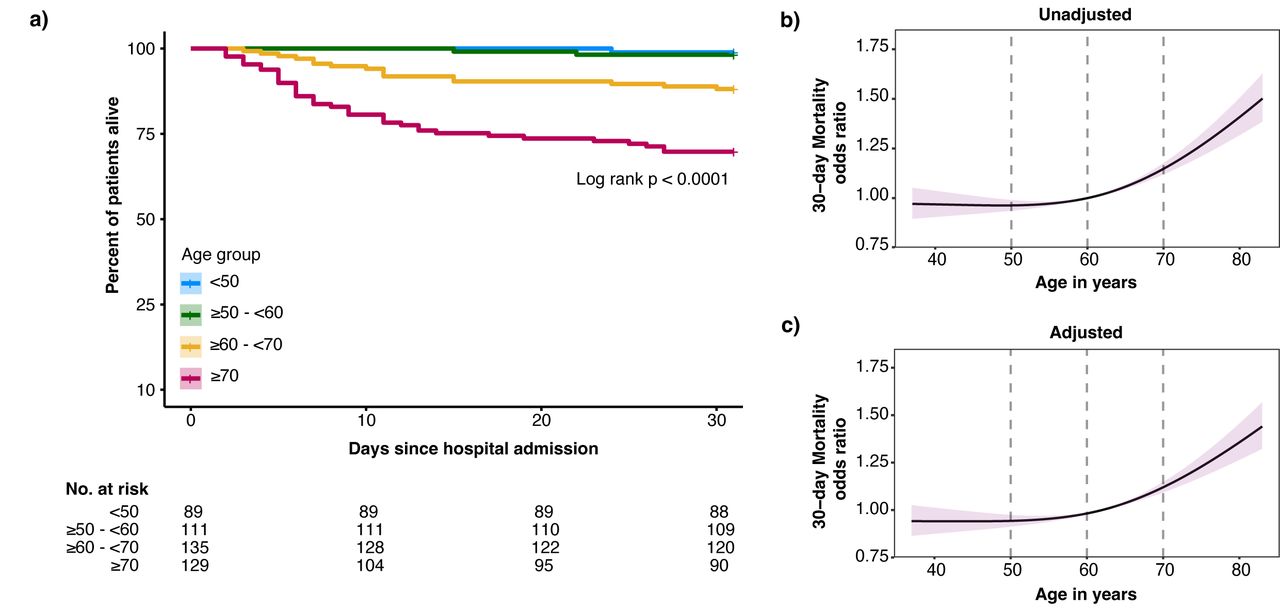
464年COVID-19承认综合医院病房患者包括(gydF4y2Ba表1gydF4y2Ba)。其中,89名患者(19.2%)小于50年在住院,111人(23.9%)≥50 - < 60年,135(29.1%),≥60 - < 70年和129年(27.8%)≥70年。性别之间没有差别年龄分布的几十年。大多数患者登记在第二个和第三个COVID-19浪潮在荷兰,SARS-CoV-2α变异的发生,成为主导。波之间的年龄分布是相似的(gydF4y2Ba表S4gydF4y2Ba)。并发症患者的比例随着年龄增加。虽然入学之前,病人的症状持续的时间没有年龄组之间的差异,老年患者面对更高的疾病严重程度评分(gydF4y2Ba26gydF4y2Ba]。常规实验室值可比年龄组之间除了年龄相关性淋巴细胞数量减少,和年龄相关性增加neutrophil-lymphocyte率和肌酐。COVID-19-related治疗,主要是补充氧气和地塞米松,年龄组之间的可比性(gydF4y2Ba表2gydF4y2Ba)。死亡率随着年龄的增加以非线性的方式(gydF4y2Ba表2gydF4y2Ba和gydF4y2Ba图2gydF4y2Ba)。例如,年龄从60到70的增加是增加30天死亡率(比值比(或)1.15,95%可信区间(CI) 1.12 - -1.18) (gydF4y2Ba图2 bgydF4y2Ba),这是统计独立的,与年龄相关的并发症和COVID-related治疗(调整或1.14 (1.11 - -1.17)(gydF4y2Ba图2 cgydF4y2Ba)。在患者肺炎病原体除了SARS-CoV-2 (gydF4y2Ba表S5gydF4y2Ba),增加一个类似的年龄(从60到70)与30天死亡率几率增加无关(OR: 1.01 (0.996 - -1.025))。此外,老年患者肺炎病原体引起的其他比SARS-CoV-2显示与SARS-CoV-2相比显著降低死亡率;≥60 - < 70 (p < 0.01),≥70 (p < 0.001) (gydF4y2Ba图S1gydF4y2Ba)。gydF4y2Ba
基线特征COVID-19病人入院时普通病房gydF4y2Ba
治疗,疾病COVID-19过程和结果的病人入院后一般病房gydF4y2Ba
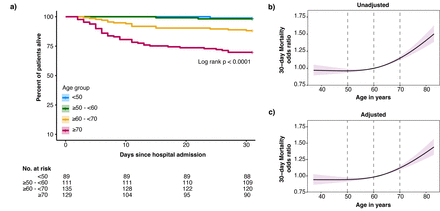
死亡率分析COVID-19一般病房的患者按年龄分层几十年。gydF4y2Ba描述gydF4y2Ba图2gydF4y2Ba:gydF4y2Ba一)kaplan meier情节的患者按年龄分组的分层。b) 30天死亡率的风险随着年龄增长模型作为连续变量。由于年龄和死亡率之间的非线性关系,限制三次样条函数三个内部结默认使用分位数的位置。计算比值比,参考设置为60岁。红色阴影代表了30天死亡率优势比的95%置信区间。b c)面板相同的方法,但是现在的30天死亡率调整优势比人口统计数据(包括医院、性别和包含波),老年性并存病(高血压、糖尿病、恶性肿瘤、免疫抑制和慢性心脏,神经系统,呼吸系统,和肾脏疾病),老年性慢性药物(抗血小板和抗凝药物),和COVID-19-related治疗前后采样(糖皮质激素如地塞米松、anti-IL-6伊马替尼、remdesivir和抗生素)。gydF4y2Ba
协会与畸变在不同的宿主反应域老化gydF4y2Ba
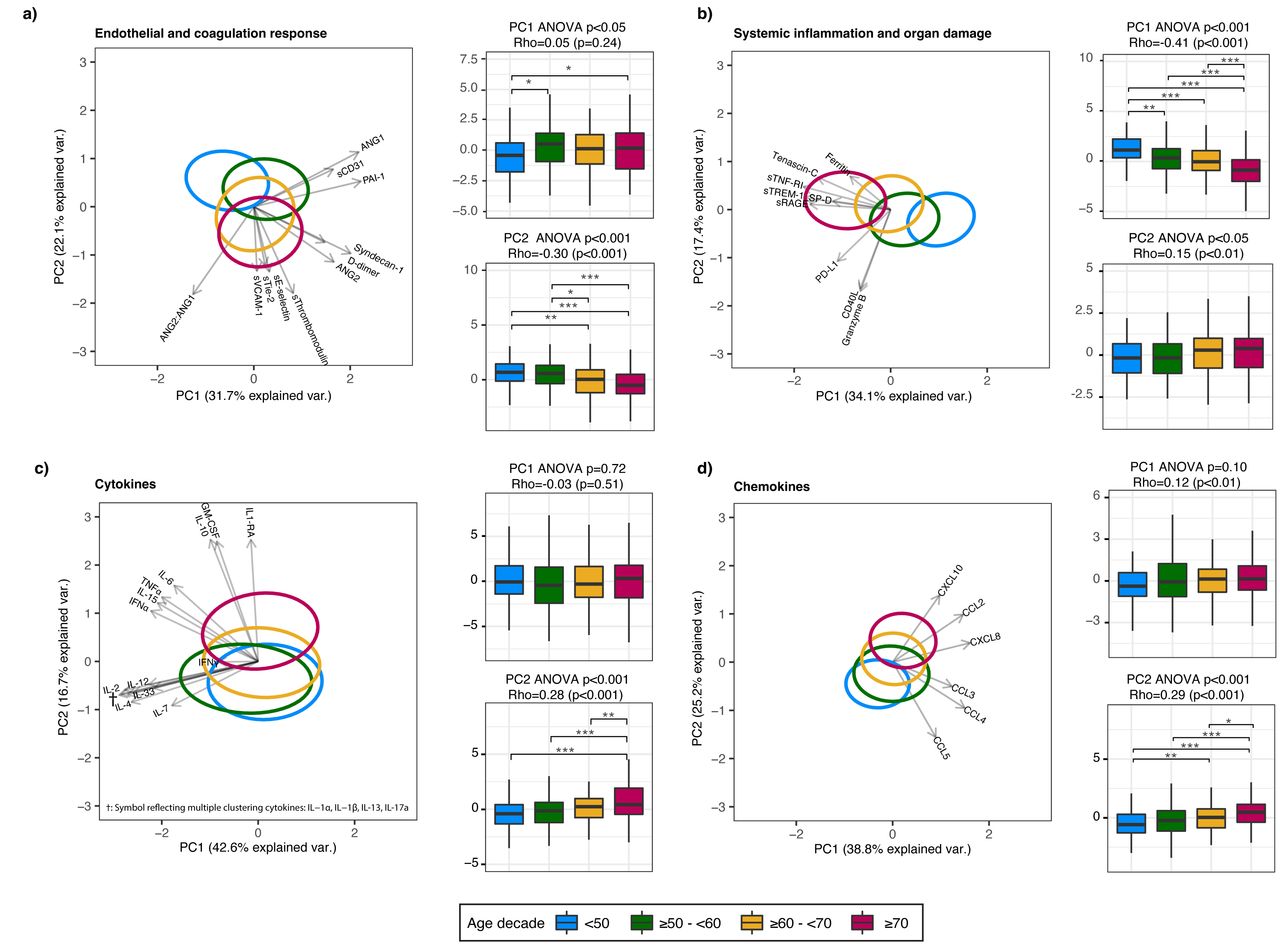
我们确定43主机响应生物标志物在等离子体后48小时内得到承认和分层这些四个病理生理学领域中。首先,我们进行PCA想象整体年龄差异几十年在生物标记在每一个病理生理域(gydF4y2Ba图3gydF4y2Ba)。所有域表明年龄组之间有着显著的不同。观察最严重的老龄化的PC1系统性炎症和器官损伤域(ρ= -0.41),这主要是由等离子体浓度的增加可溶性肿瘤坏死因子受体1 (sTNF-RI),溶在髓细胞触发受体表达1 (sTREM-1)和可溶性受体晚期糖化终产物(sRAGE)在老年患者(gydF4y2Ba图3 bgydF4y2Ba)。第二个最强协会是老化的PC2内皮和凝固域(ρ= 0.30),主要的等离子angiopoietin-2/1比率的增加,sThrombomodulin和gydF4y2BasegydF4y2Ba在老年患者-selectin (gydF4y2Ba图3一gydF4y2Ba)。细胞因子的细胞因子域是由两组特征:一个与老龄化水平增加(gydF4y2Ba图3 cgydF4y2Ba、PC2主要由集落刺激因子(gm - csf)和抗炎细胞因子IL-1RA和il - 10),和一个水平相对相似年龄组(gydF4y2Ba图3 cgydF4y2BaPC1)。趋化因子的混合模式观察领域一些趋化因子与老龄化水平增加(gydF4y2Ba如。gydF4y2Ba,C-X-C主题趋化因子配体10 (CXCL10))而其他减少老化(gydF4y2Ba如。gydF4y2Ba趋化因子配体5主题趋化因子(CCL5)) (gydF4y2Ba图3 dgydF4y2Ba、PC2)。每个生物标志物PC的完成贡献分数描述了gydF4y2Ba表S6gydF4y2Ba。敏感性分析,我们进一步分层患者≥70为病人≥70 - < 80 (n = 86),≥80 (n = 43),没有修改年龄相关性趋势在很大程度上(gydF4y2Ba图S2gydF4y2Ba)。gydF4y2Ba
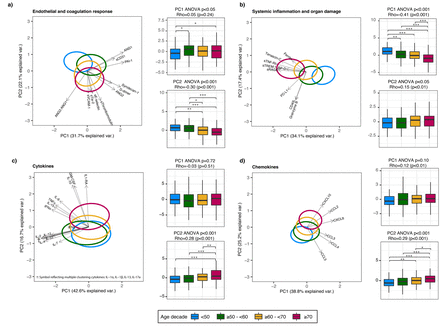
主成分分析主机响应域年龄组之间的差异。gydF4y2Ba描述gydF4y2Ba图3gydF4y2Ba:gydF4y2Ba主成分分析(PCA)的主成分1和2 (PC)绘制每个域。对每个域,x和y轴标签%的总方差域中分别由PC1解释和PC2。每个生物标志物PC的完成贡献分数描述了gydF4y2Ba表S6gydF4y2Ba。椭圆表明中央每个年龄组的10%,彩色编码显示在底部图的一部分。箭头指示的方向(箭头方向)和力量(箭头长度)每个生物标志物和个人电脑之间的相关性。每个PCA旁边情节与1.5箱线图PC1和PC2四分位范围的胡须。本上假定值年龄组之间是通过方差分析:ρ的伴随假定值生成使用枪兵与衰老相关的连续的规模。注意,主成分与老龄化的负面协会可能仍然反映了与生物标志物浓度正相关,所反映的方向箭头。因果测试与图基做了测试。* * * * * p < 0.001, p < 0.01, p < 0.05。缩写:和:检验;sTie-2:可溶性Tie-2;gydF4y2BasegydF4y2Ba-selectin:可溶性E-selectin;sThrombomodulin:可溶性thrombomodulin;sVCAM-1:可溶性血管细胞粘附molecule-1;PAI-1:纤溶酶原激活物inhibitor-1;sCD31:可溶性集群分化31日;sRAGE:晚期糖化终产物的可溶性受体;sTNF-R1:可溶性肿瘤坏死因子受体1;sTREM-1:可溶性触发受体表达髓细胞1;SP-D:表面活性剂蛋白D;CD40L: cd40配体; PD-L1: programmed death-ligand 1; CCL: chemokine C-C motif ligand; CXCL: C-X-C motif chemokine ligand; IL: interleukin; TNF: tumor necrosis factor; GM-CSF: granulocyte-macrophage colony-stimulating factor; IFN: interferon.
协会的老化与单个主机响应生物标志物gydF4y2Ba
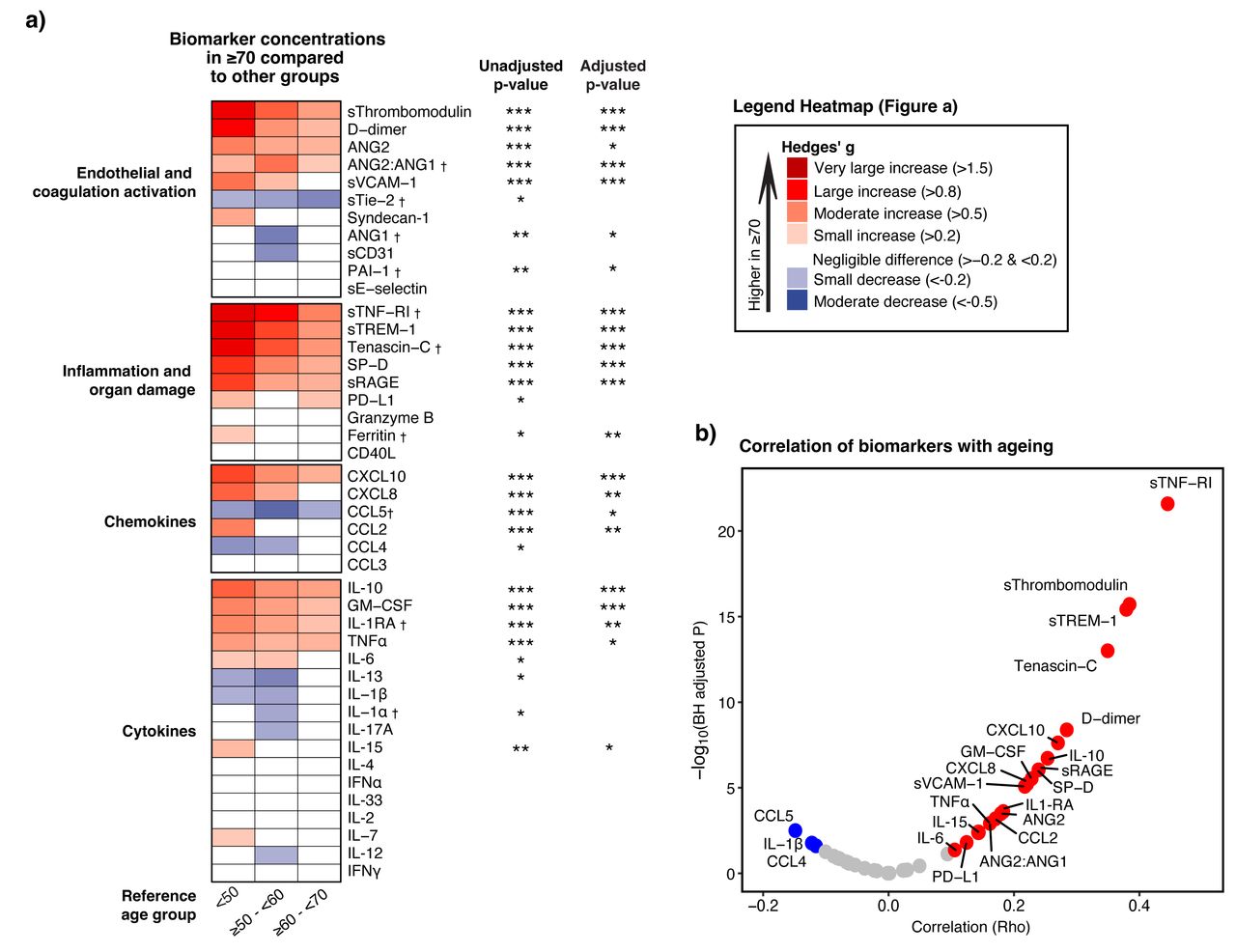
我们下一个比较个人主机响应生物标志物在年龄几十年。43的生物标记物决定的,36之间明显不同病人和控制(补充Excel文件,表2)。gydF4y2Ba图4一gydF4y2Ba显示生物标志物浓度≥70岁的病人比其他年龄组,表示为对冲的g,常用的效应大小测量(gydF4y2Ba22gydF4y2Ba]。这种分析证实,大多数生物标志物反映内皮和凝固的激活,以及那些表明系统性炎症和器官损伤,患者高≥70,而趋化因子和细胞因子表现出混合模式。灵敏度分析中,我们进一步分层患者≥70为病人≥70 - < 80和≥80取得了类似的结果(gydF4y2Ba图S2gydF4y2Ba)。几乎所有生物标志物与衰老有重要的协会在未经调整的模型中,意义是保持人口结构调整后,并存病,COVID-19-related immunomodulating治疗前抽样(有关详细信息,请参阅方法)。规模连续,sTNF-RI、sTREM-1 sThrombomodulin和tenascin-C最强的(积极的)与衰老相关(gydF4y2Ba图4 bgydF4y2Ba)。每个生物标志物的关系的强度和方向与衰老表现出强烈的类似于跨包含波(gydF4y2Ba图S3gydF4y2Ba)。gydF4y2Ba
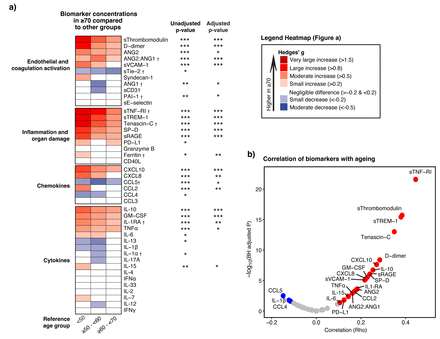
协会主机响应与衰老生物标志物。gydF4y2Ba描述gydF4y2Ba图4gydF4y2Ba:gydF4y2Baa)的热图描绘生物标志物的大小之间的差异(树篱的g)患者≥70岁,另一组。假定值获得从线性(如果线性)或三次样条回归分析(如果非线性)的年龄是模仿作为连续变量。调整后的模型包括人口统计,与年龄相关的并发症、年龄和biomarker-related慢性药物和COVID-19-related immunomodulating治疗在抽样之前,看到细节的方法。红色代表高水平的病人≥70:蓝色表示这个年龄段的低水平。b)火山情节描绘的强度之间的相关生物标志物和老化。红点代表显著正相关,蓝点显著负相关,和灰色点与相关性。调整和未经调整的假定值使用Benjamini-Hochberg多个测试修正(BH)程序测试43生物标志物。* * * * * p < 0.001, p < 0.01, p < 0.05。†生物标志物与非线性连续规模与衰老的关系。缩写:和:检验; sTie-2: soluble Tie-2;segydF4y2Ba-selectin:可溶性E-selectin;sThrombomodulin:可溶性thrombomodulin;sVCAM-1:可溶性血管细胞粘附molecule-1;PAI-1:纤溶酶原激活物inhibitor-1;sCD31:可溶性集群分化31日;sRAGE:晚期糖化终产物的可溶性受体;sTNF-R1:可溶性肿瘤坏死因子受体1;sTREM-1:可溶性触发受体表达髓细胞1;SP-D:表面活性剂蛋白D;CD40L: cd40配体; PD-L1: programmed death-ligand 1; CCL: Chemokine C-C motif ligand; CXCL: C-X-C motif chemokine ligand; IL: interleukin; TNF: tumor necrosis factor; GM-CSF: granulocyte-macrophage colony-stimulating factor; IFN: interferon.
在一个单独的分析,我们评估了年龄分布的生物标志物群入住ICU危重COVID-19病人样本(n = 157,gydF4y2Ba表S8和表S9gydF4y2Ba;补充Excel文件,表3)。例≥70和年轻患者之间的差异很大程度上再现这群危重患者,尤其是相比,最年轻的年龄组(< 50),尽管如此,由于相对较低的样本大小,并不总是达到统计学意义(gydF4y2Ba图S4agydF4y2Ba)。类似于一般的病人的病房里,sTNF-RI, sTREM-1, sThrombomodulin, tenascin-C,可溶性血管细胞粘附molecule-1 (sVCAM-1)与衰老表现出显著的正相关(gydF4y2Ba图S4bgydF4y2Ba)。gydF4y2Ba
协会age-driven主机响应变化与死亡率gydF4y2Ba
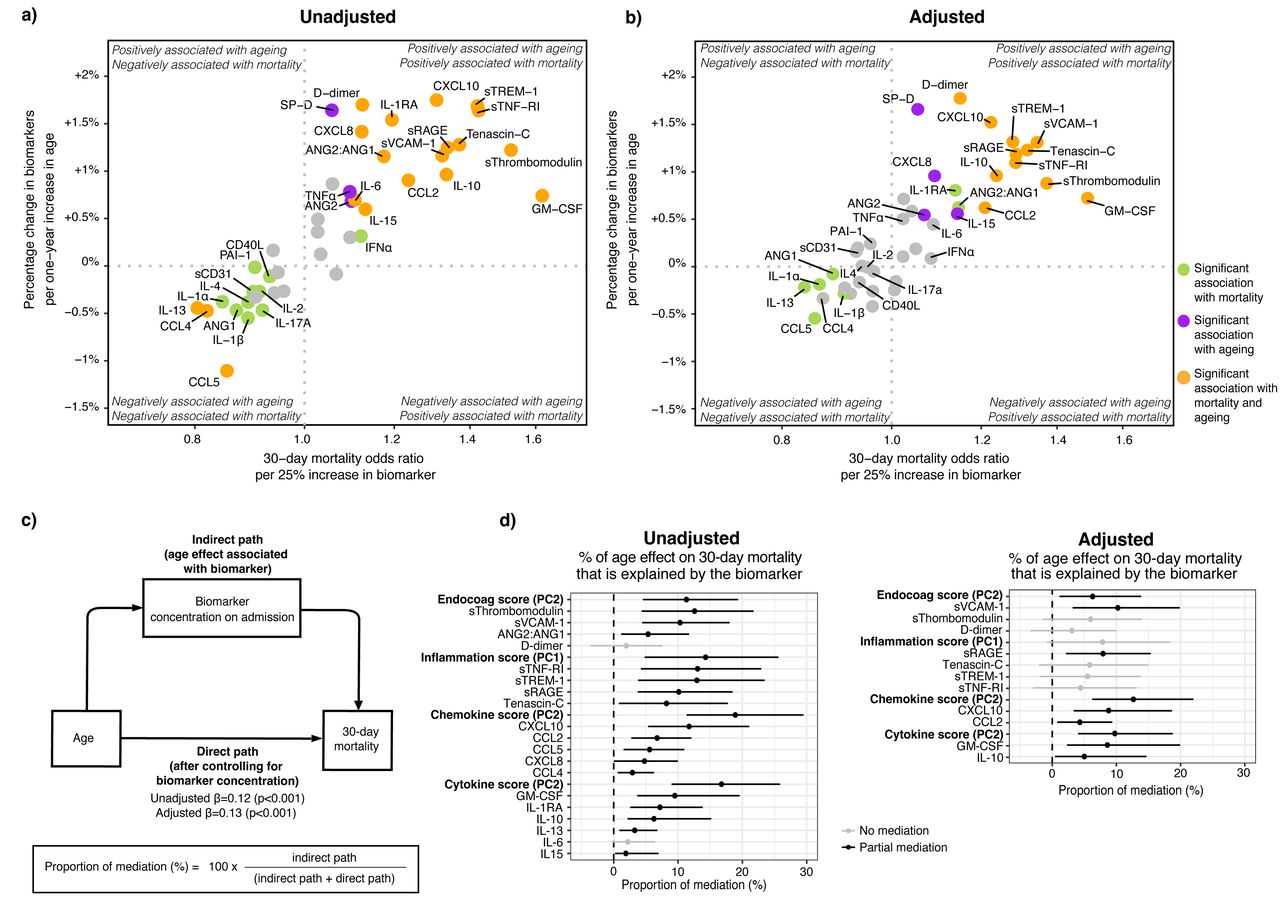
我们使用中介分析评估是否age-driven改变生物标志物浓度与age-driven增加30天死亡率(gydF4y2Ba23gydF4y2Ba]。首先,我们发现生物标志物与老化和调整之前和之后30天死亡率可能的混杂和衰老相关因素(gydF4y2Ba图5一个gydF4y2Ba和b)。然后,我们估计的年龄比例影响死亡率与年龄相关改变的病理生理学领域取得的成绩在PC和个人主机响应生物标记(gydF4y2Ba图5度gydF4y2Ba)。在未经调整的模型(gydF4y2Ba图5 dgydF4y2Ba(左),PC的内皮和凝固(PC2)、系统性炎症和器官损伤(PC1)趋化因子(PC2)和细胞因子域(PC2)解释年龄影响死亡率的重要组成部分。关于个人的生物标志物,摩根大通sThrombomodulin差异sVCAM-1, sTNF-RI, sTREM-1 sRAGE最相关age-driven 30天死亡率的增加。值得注意的是,在外部群体,sThrombomodulin sVCAM-1, sTNF-RI, Tenascin-C显示最严重的老龄化和30天死亡率(gydF4y2Ba图S5gydF4y2Ba);sTREM-1不是测量(gydF4y2Ba21gydF4y2Ba]。在调整后的模型,只有ageing-driven sVCAM-1的变化,sRAGE, CXCL10, gm - csf和il - 10与ageing-driven 30天死亡率增加有关。总的来说,这些结果表明,年龄和死亡率之间的关系中所占的比例是由特定的主机响应差异。gydF4y2Ba
![Overview of COVID-19 cohorts. Description Figure 1: Overview of COVID-19 cohorts in which plasma biomarkers were measured. The external validation cohort was derived from a publicly available data set of non-intubated COVID-19 patients in whom plasma proteins were measured by Olink Proximity Extension Assay [21].](http://www.qdcxjkg.com/content/erj/early/2023/04/06/13993003.00011-2023/F1.large.jpg?width=800&height=600&carousel=1){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
中介分析ageing-associated死亡率和宿主反应生物标记物在进入普通病房。gydF4y2Ba描述gydF4y2Ba图5gydF4y2Ba:gydF4y2Ba象限图。x轴描绘了增加30天死亡率优势比每增加25%的生物标志物来自未经调整逻辑回归的对数转换生物标志物作为解释变量和30天死亡率作为响应变量。y轴显示的百分比变化的生物标志物浓度增加每一时代来自未经调整线性回归分析与对数转换生物标志物作为反应变量。生物标志物在右上角和左下角最有可能与年龄相关性增加30天死亡率相关。是多个测试修正系数的意义使用Benjamini-Hochberg (BH)程序测试43生物标志物。b)面板相同的方法,然而,这两个系数现在调整人口结构(包括医院、性别和包含波),老年性并存病(高血压、糖尿病、恶性肿瘤、免疫抑制和慢性心脏,神经系统,呼吸系统,和肾脏疾病),年龄和biomarker-related慢性药物(抗血小板和抗凝药物),和COVID-19-related治疗前后采样(糖皮质激素如地塞米松、anti-IL-6伊马替尼、remdesivir和抗生素)。冥想的c)图分析。模型包含相同的协变量调整面板b d)未经调整(左)和调整(右)中介分析结果。只有生物标志物和主成分与衰老和30天死亡率显著相关进行分析。置信区间从1000年获得x引导。 The higher the proportion of mediation, the stronger the association of the age-dependent differences in that biomarker and the age-dependent increase in 30-day mortality. The principal components and their contributing biomarker are depicted in图3gydF4y2Ba。每个生物标志物PC的完成贡献分数描述了gydF4y2Ba表S6gydF4y2Ba。缩写:Endocoag得分:内皮和凝固分数;ANG:检验;sTie-2:可溶性Tie-2;gydF4y2BasegydF4y2Ba-selectin:可溶性E-selectin;sThrombomodulin:可溶性thrombomodulin;sVCAM-1:可溶性血管细胞粘附molecule-1;PAI-1:纤溶酶原激活物inhibitor-1;sCD31:可溶性集群分化31日;sRAGE:晚期糖化终产物的可溶性受体;sTNF-R1:可溶性肿瘤坏死因子受体1;sTREM-1:可溶性触发受体表达髓细胞1;SP-D:表面活性剂蛋白D;CD40L: cd40配体; PD-L1: programmed death-ligand 1; CCL: Chemokine C-C motif ligand; CXCL: C-X-C motif chemokine ligand; IL: interleukin; TNF: tumor necrosis factor; GM-CSF: granulocyte-macrophage colony-stimulating factor; IFN: interferon.
考虑到主机响应在老年患者可能不统一,我们在患者≥70进行了聚类分析。我们确定了三个集群(gydF4y2Ba图S6gydF4y2Ba)。集群3岁和30天死亡率最高,高内皮细胞激活,炎症和器官损伤的标记,但低细胞因子浓度(gydF4y2Ba表S10, S11表gydF4y2Ba集群和补充excel文件表4)。1有很高的凝固标记和细胞因子浓度(gydF4y2Ba图S6gydF4y2Ba)。集群2的宿主反应好坏参半。前三调解生物标志物的主要分析(sThrombomodulin sTNF-RI, sTREM-1)是重要的将患者分配给集群3 (gydF4y2Ba表S12gydF4y2Ba)。此外,绝大多数的生物标记和所有电脑成绩的重要介质,主要分析仍然预测病人的死亡率≥70,除了少数例外,独立于指定集群(gydF4y2Ba表向gydF4y2Ba)。gydF4y2Ba
讨论gydF4y2Ba
我们这里报告协会的老化与43个生物标志物反映主机响应干扰四个关键COVID-19的发病机制和病理生理领域相关的这些免疫偏离30天死亡率相关。虽然我们的研究结果证实之前报道血浆il - 6的浓度增加,il - 10, IL-15, TNFαCXCL8老COVID-19患者(gydF4y2Ba15gydF4y2Ba- - - - - -gydF4y2Ba18gydF4y2Ba),我们另外表明,许多细胞因子并不受年龄的影响。更重要的是,我们提供新的证据表明,老龄化与内皮细胞的激活和炎症和器官损伤的凝血系统和海拔COVID-19标记。中介分析表明特定的主机响应变化解释部分对死亡率的影响。我们的研究是最老化之间的关系的综合分析,在COVID-19主机响应和死亡率。gydF4y2Ba
老COVID-19病人显示强烈的等离子体浓度增加几个生物标志物内皮细胞和凝固激活和炎症和器官损伤域。有可能的是,在这些病理生理改变区域至少部分交织在一起的。这些(以及其他所有)分析是在一个未经调整的模型和模型进行调整的变量与衰老有关。虽然这两种分析主要产生了类似的结果,我们认为未经调整的分析更好地反映老年患者的临床,谁(举个例子)本质上有更多的并发症和co-medication比年轻病人。老龄化导致内皮细胞衰老,通过血管炎症和内皮功能障碍为特征的;与年龄增加内皮细胞NFκB活动已经涉及本(gydF4y2Ba27gydF4y2Ba,gydF4y2Ba28gydF4y2Ba]。Endotheliopathy COVID-19的一个关键特性,早期的研究报道升高血浆水平的多种生物标志物指示内皮细胞激活和功能障碍(gydF4y2Ba29日gydF4y2Ba]。目前的结果表明,COVID-19与更深刻的endotheliopathy在老年患者。事实上,病人≥70水平较高的等离子体生物标志物反映内皮细胞激活(sThrombomodulin sVCAM-1),更不安glycocalyx函数(syndecan),和更中断屏障功能(angiopoietin-2)。失去glycocalyx COVID-19[内皮功能障碍的一个重要特性gydF4y2Ba29日gydF4y2Ba,gydF4y2Ba30.gydF4y2Ba]。glycocalyx由蛋白聚糖和粘多糖,抑制免疫激活和提供一个表面抗凝(gydF4y2Ba30.gydF4y2Ba]。因此,一个更不安glycocalyx函数在老年COVID-19病人可能导致系统性hyperinflammation和凝固激活。angiopoietin-2和angiopoietin-2/1比高老COVID-19患者指向同一个方向:angiopoietin-2促进内皮炎症和血管渗漏、高血浆angiopoietin-2与COVID-19(更糟糕的结果gydF4y2Ba31日gydF4y2Ba),和高angiopoietin-2/1比率的强烈与死亡率相关危重患者急性肺损伤(gydF4y2Ba32gydF4y2Ba]。值得注意的是,老年人之间的联系和增加endotheliopathy COVID-19似乎是特定的,因为我们最近报道降低内皮细胞激活和功能障碍老人比年轻人患者脓毒症(gydF4y2Ba33gydF4y2Ba]。gydF4y2Ba
老化是一个独立的危险因素死亡率在这项研究群非关键COVID-19患者,证实了之前的调查,分析人群提供更多样的基线疾病严重程度(gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2gydF4y2Ba,gydF4y2Ba5gydF4y2Ba]。更重要的是,我们表明,年龄相关性增加30天死亡率与特定的与年龄有关的宿主反应的变化。sTNF-RI已被确定为一个生物标志物基础不利”hyperinflammation“亚型急性呼吸窘迫综合征(gydF4y2Ba34gydF4y2Ba]。sTNF-RI水平可能是代理标记细胞相关的激活TNF-RI,可引发肺血管渗漏和中性粒细胞炎症(gydF4y2Ba35gydF4y2Ba- - - - - -gydF4y2Ba37gydF4y2Ba]。在健康个体的封锁TNF-RI导致中性粒细胞减少轮回和内皮损伤在吸入内毒素(gydF4y2Ba35gydF4y2Ba],TNF-RI缺陷小鼠强烈防止肺水肿的形成在急性肺损伤模型gydF4y2Ba37gydF4y2Ba]。总的来说,这些数据表明,针对TNF-RI途径可以改善结果特别是老COVID-19病人。TREM-1是先天免疫细胞的受体放大toll样和NOD-receptors引发的免疫反应gydF4y2Ba38gydF4y2Ba]。浓度的增加sTREM-1反映TREM-1途径活性(gydF4y2Ba39gydF4y2Ba]和与增加细胞因子水平与疾病严重程度患者COVID-19 [gydF4y2Ba40gydF4y2Ba]。在我们的研究中,老化与增强sTREM-1水平,这是一个主要决定因素ageing-driven 30天死亡率的增加。最近完成的一项研究的兴趣,COVID-19病人需要呼吸机支持注入TREM-1通路的抑制剂改善临床结果,包括28天死亡率(数据由赞助商新闻稿;看到裁判(gydF4y2Ba41gydF4y2Ba])。gydF4y2Ba
血浆il - 6与老化和COVID-19死亡率相关。然而,在中介分析,年龄相关性增加30天死亡率没有增加il - 6的浓度,表明il - 6封锁不得老COVID-19患者受益。在协议,叫的有益作用在整个人口COVID-19病人参加一大个随机对照试验没有观察到的最古老的患者(gydF4y2Ba11gydF4y2Ba]。同样,dexamethason——COVID-19其他免疫调节治疗的基石——老病人没有改善的结果(gydF4y2Ba9gydF4y2Ba,gydF4y2Ba10gydF4y2Ba]。老年患者中升高的炎症可能导致类固醇电阻(gydF4y2Ba42gydF4y2Ba),这可能导致较低的响应标准剂量的地塞米松。总的来说,这些发现表明,与年龄有关的变化在主机响应COVID-19影响免疫调制的影响在这个疾病和年龄应该考虑病人的选择。gydF4y2Ba
我们的研究有优势和局限性。我们提供全面的洞察畸变在主机响应途径被认为是重要的发病机制COVID-19人口众多。我们主要分析只包括非关键病人,然而我们提供初步证据表明,老龄化与类似的主机响应变化COVID-19 ICU的患者。我们的研究是纯粹的观察,没有明确的结论可以得出因果关系。尽管如此,通过使用中介分析,我们提供新颖的见解,宿主反应途径有助于ageing-associated死亡率。虽然承认等离子体生物标志物用于指导immunomodulating轨迹(gydF4y2Ba43gydF4y2Ba],他们不包括主人的全光谱响应,我们可能会错过了主机响应变化,没有(还)可见的蛋白质水平。我们之前研究的广泛可用性疫苗和ο变异的主导地位。然而,在疫苗接种覆盖率高的人群,很大一部分住院COVID-19患者仍未接种疫苗(gydF4y2Ba44gydF4y2Ba]。虽然ο变体仍然不成比例地影响老年患者(gydF4y2Ba45gydF4y2Ba),健壮性的ageing-associated变化仍有待验证。gydF4y2Ba
本研究文档老化与实质性的和广泛的跨几个病理生理改变宿主应对COVID-19域。几个生物标志物浓度增加(尤其是sTNF-RI和sTREM-1)与ageing-driven 30天死亡率增加有关。这些结果可能为新在老年患者免疫调节目标奠定基础。gydF4y2Ba
确认gydF4y2Ba
我们感谢芭芭拉Smids-Dierdorp,塔玛拉·德克尔和安妮塔Tuip Luminex化验的性能的技术援助。图形抽象和gydF4y2Ba图1gydF4y2Ba是用BioRender.com。gydF4y2Ba
脚注gydF4y2Ba
1gydF4y2Ba重症监护、阿姆斯特丹联电,阿姆斯特丹,荷兰;gydF4y2Ba
2gydF4y2Ba传染病科,阿姆斯特丹联电,阿姆斯特丹,荷兰;gydF4y2Ba
3gydF4y2Ba实验免疫学,阿姆斯特丹联电,阿姆斯特丹,荷兰;gydF4y2Ba
4gydF4y2Ba神经科学神经学部门,阿姆斯特丹联电,阿姆斯特丹,阿姆斯特丹,荷兰;gydF4y2Ba
5gydF4y2Ba肺学部门,阿姆斯特丹联电,阿姆斯特丹,荷兰;gydF4y2Ba
6gydF4y2Ba美国麻醉学,阿姆斯特丹联电,阿姆斯特丹,荷兰;gydF4y2Ba
7gydF4y2Ba阿姆斯特丹阿姆斯特丹联电生物核心设施,联电,阿姆斯特丹,荷兰;gydF4y2Ba
8gydF4y2Ba部放射,阿姆斯特丹联电,阿姆斯特丹,荷兰;gydF4y2Ba
9gydF4y2Ba医学微生物学、阿姆斯特丹联电,阿姆斯特丹,荷兰;gydF4y2Ba
10gydF4y2Ba内科,阿姆斯特丹联电,阿姆斯特丹,荷兰;gydF4y2Ba
11gydF4y2Ba神经化学实验室,阿姆斯特丹联电,阿姆斯特丹,荷兰;gydF4y2Ba
12gydF4y2Ba美国临床化学,阿姆斯特丹联电,阿姆斯特丹,荷兰;gydF4y2Ba
13gydF4y2Ba临床流行病学、生物统计学和生物信息学,阿姆斯特丹联电,阿姆斯特丹,荷兰。gydF4y2Ba
伦理批准:参与医院的伦理委员会批准了研究收集数据的目的。gydF4y2Ba
数据共享:数据可以共享在合理的提案请求批准后签署了数据访问协议和总合作学习小组。gydF4y2Ba
合作者gydF4y2Ba
阿姆斯特丹联电COVID-19生物gydF4y2Ba
米歇尔•阿格塔米尔gydF4y2Ba2gydF4y2Ba,安妮Geke AlgeragydF4y2Ba1gydF4y2Ba,布伦特AppelmangydF4y2Ba2gydF4y2Ba、地板van BaarlegydF4y2Ba1gydF4y2Ba卡坦,BeudelgydF4y2Ba4gydF4y2BaJan Bogaard伤害,gydF4y2Ba5gydF4y2Ba,Marije BomersgydF4y2Ba2gydF4y2Ba彼得BontagydF4y2Ba5gydF4y2Ba,Lieuwe BosgydF4y2Ba1gydF4y2Ba罗曼斯博塔携手gydF4y2Ba1gydF4y2Ba贾斯汀·德·BrabandergydF4y2Ba2gydF4y2BaGodelieve·德·布莉gydF4y2Ba2gydF4y2BaSanne de熊先生gydF4y2Ba1gydF4y2Ba,玛丽安娜BugianigydF4y2Ba5gydF4y2Ba,以斯帖BullegydF4y2Ba1gydF4y2Ba大卫·t中方gydF4y2Ba1gydF4y2Ba,Osoul ChouchanegydF4y2Ba2gydF4y2Ba,亚历克斯ClohertygydF4y2Ba3gydF4y2Ba,Mirjam迪杰斯特拉gydF4y2Ba12gydF4y2Ba戴夫·a . DongelmansgydF4y2Ba1gydF4y2Ba过w•b西博尔德作品Romein·雅gydF4y2Ba1gydF4y2Ba,保罗ElbersgydF4y2Ba1gydF4y2Ba,卢卡斯FleurengydF4y2Ba1gydF4y2Ba,苏珊娜GeerlingsgydF4y2Ba2gydF4y2Ba西奥GeijtenbeekgydF4y2Ba3gydF4y2Ba,阿尔芒GirbesgydF4y2Ba1gydF4y2Ba,布拉姆GoorhuisgydF4y2Ba2gydF4y2Ba马丁·p·GrobuschgydF4y2Ba2gydF4y2Ba劳拉毫米gydF4y2Ba1gydF4y2Ba,Jorg哈曼gydF4y2Ba7gydF4y2Ba,凡妮莎·哈里斯gydF4y2Ba2gydF4y2Ba罗伯特•HemkegydF4y2Ba8gydF4y2Ba,Sabine m .何曼思gydF4y2Ba2gydF4y2BaLeo HeunksgydF4y2Ba1gydF4y2Ba,马库斯HollmanngydF4y2Ba6gydF4y2Ba、詹妮克角gydF4y2Ba1gydF4y2Ba,Joppe w . HoviusgydF4y2Ba2gydF4y2Ba,卡佳德容gydF4y2Ba2gydF4y2Ba,Menno d·德容gydF4y2Ba9gydF4y2Ba念完,今敏gydF4y2Ba4gydF4y2Ba,Bregje LemkesgydF4y2Ba2gydF4y2Ba,Endry热处理LimgydF4y2Ba1gydF4y2BaNiels van MourikgydF4y2Ba1gydF4y2Ba,Jeaninne NellengydF4y2Ba2gydF4y2Ba以斯帖j . NossentgydF4y2Ba5gydF4y2Ba,Sabine使用橄榄gydF4y2Ba4gydF4y2Ba,Frederique保卢斯gydF4y2Ba1gydF4y2Ba彼得斯,埃德加gydF4y2Ba2gydF4y2Ba,丹人工智能Pina-FuentesgydF4y2Ba4gydF4y2Ba汤姆,van der投票gydF4y2Ba2gydF4y2Ba,Bennedikt PreckelgydF4y2Ba6gydF4y2Ba简•m•普林斯gydF4y2Ba2gydF4y2Ba,Jorinde RaasveldgydF4y2Ba1gydF4y2Ba,汤姆ReijndersgydF4y2Ba2gydF4y2Ba,Maurits C.F.J. de生搬硬套gydF4y2Ba12gydF4y2Ba,米歇尔•SchinkelgydF4y2Ba2gydF4y2Ba马库斯·j·舒尔茨gydF4y2Ba1gydF4y2Ba美联社Femke SchrauwengydF4y2Ba12gydF4y2Ba舒尔曼,亚历克斯gydF4y2Ba2gydF4y2Ba傻瓜,舒尔曼gydF4y2Ba1gydF4y2Ba,金正日SigaloffgydF4y2Ba1gydF4y2Ba玛莲a苗条gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2gydF4y2Ba帕特里克SmeelegydF4y2Ba5gydF4y2Ba,结婚SmitgydF4y2Ba1gydF4y2BaCornelis s . StijnisgydF4y2Ba2gydF4y2Ba,Willemke StilmagydF4y2Ba1gydF4y2Ba,夏洛特TeunissengydF4y2Ba11gydF4y2Ba帕特里克ThoralgydF4y2Ba1gydF4y2BaAnissa M TsonasgydF4y2Ba1gydF4y2BaPieter r TuinmangydF4y2Ba1gydF4y2BaMarc van der ValkgydF4y2Ba2gydF4y2Ba,丹尼斯VeelogydF4y2Ba6gydF4y2Ba,Carolien VollemangydF4y2Ba1gydF4y2Ba,曾经德弗里斯gydF4y2Ba1gydF4y2BaLonneke a VughtgydF4y2Ba1gydF4y2Ba,gydF4y2Ba2gydF4y2Ba米歇尔van VugtgydF4y2Ba2gydF4y2BaDorien武泰,gydF4y2Ba12gydF4y2Baa . H Zwinderman(三星)gydF4y2Ba13gydF4y2BaMatthijs c .这gydF4y2Ba4gydF4y2Ba,w . Joost WiersingagydF4y2Ba2gydF4y2Ba,亚历山大P.J. VlaargydF4y2Ba1gydF4y2BaDiederik van de发现(d.vandebeek@amsterdamumc.nl)gydF4y2Ba4gydF4y2Ba。gydF4y2Ba
资助信息:E.H.A.M.,法学博士。B, C.C.A.v.L.收到欧盟资助的地平线2020研究和创新项目赠款协议没有847786(公平)。H.P.S.支持由荷兰肾脏基金会(每年授予Nr ok009 19日)。超频是兰德斯泰纳基金(LSBR # 1901)。T.D.Y.R.是支持的研究项目“NACTAR,项目“MDR-phage”(批准号16447),这是由荷兰研究委员会(NWO)。WJW Talud基金(左Talud)阿姆斯特丹联电电晕研究基金,荷兰卫生研究与发展组织(ZonMw;TURN-COVID格兰特号10430142110001)。LDJB报告从荷兰肺脏基金会赠款,资助从荷兰肺脏基金会和健康荷兰(公私合作资助),从荷兰肺脏基金会赠款(Dirkje Postma奖),资助的IMI COVID19倡议,从阿姆斯特丹联电奖学金和助学金。本研究的资助者没有参与研究设计、数据收集、数据分析、数据解释或报告的写作。gydF4y2Ba
作者的贡献:EHAM,英航,HPS TvdP导致的概念和设计研究。EHAM,英航、JdB RBEvA CCAvL, OC, ARS, TDYR, tal, AMK, RAD,阿姆斯特丹联电COVID-19生物研究小组负责数据的收集,样品和样品制备。EHAM进行了数据分析。HPS验证了分析方法。EHAM起草了第一个手稿与英国航空公司协商,HPS TvdP。TvdP监督项目并提供资金。所有作者提供知识的输入和修改和批准了最终版本的手稿。gydF4y2Ba
利益冲突:所有作者声明没有利益冲突。gydF4y2Ba
- 收到了gydF4y2Ba2023年1月3日。gydF4y2Ba
- 接受gydF4y2Ba2023年4月10日。gydF4y2Ba
- 版权©2023年作者。gydF4y2Ba
这个版本分布在Creative Commons归因执照的条款4.0。gydF4y2Ba
引用gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba






