





摘要gydF4y2Ba
肺动脉高压(PAH)是一种破坏性的肺血管疾病,常导致右心衰和死亡。目前的治疗干预策略只能减缓疾病进展。异常低氧诱导因子(HIF)2α的稳定性和功能在肺动脉高压(PH)的发生和发展中的作用是近二十年来备受关注的一个领域。gydF4y2Ba
在这里,我们确定了一种新型HIF2α抑制剂(PT2567)对PH疾病起始和进展的影响,使用两种PH临床前模型进行血流动力学测量,然后收集心脏、肺和血液进行病理、基因表达和生化分析。特发性PAH患者的血外生长内皮细胞被用来检测hif2 α抑制对内皮功能的影响。gydF4y2Ba
在su5416/缺氧预防和干预模型中,HIF2a的全面抑制降低了肺血管血流动力学和肺血管重塑。PT2567干预降低了肺和心脏组织中ph相关靶基因的表达,并恢复了血浆亚硝酸盐浓度。用PT2567治疗暴露于野百合碱的啮齿动物减少了对心血管血流动力学的影响,并提高了生存优势。gydF4y2Ba在体外gydF4y2Ba研究发现,人肺动脉内皮细胞中HIF2α信号的缺失抑制了与炎症相关的靶基因,PT2567降低了特发性PAH患者血外生长内皮细胞的高增殖表型和精氨酸酶活性过度活跃。这些数据表明,用口服生物可用化合物靶向HIF2α异二聚体可能为PAH提供一种新的治疗方法。未来的研究需要确定HIF在异质性多环芳烃人群中的作用。gydF4y2Ba
摘要gydF4y2Ba
多环芳烃是一种无法治愈的使人衰弱的疾病。对新的变革性疗法的需求尚未得到满足。通过抑制ARNT异二聚体靶向HIF2α功能可减少动物中与已确定的PH疾病相关的许多临床症状。gydF4y2Bahttps://bit.ly/3jHK8PSgydF4y2Ba
简介gydF4y2Ba
肺部的氧气交换需要通气和灌注之间的良好匹配。虽然肺暴露在最高的局部氧张力的身体,有生理和病理条件,导致局部低氧可用性(缺氧)。这种局部缺氧被肺血管系统特有的、独特的氧感知能力抵消,其特征是对氧张力下降的深刻血管收缩反应[gydF4y2Ba1gydF4y2Ba].局部肺血管收缩刺激灌注的动态变化,以帮助最大限度地捕获氧气;然而,长时间暴露在缺氧环境中会引发强有力的刺激,导致肺血管重塑,这是特发性肺纤维化的一个标志[gydF4y2Ba2gydF4y2Ba]、慢性阻塞性肺病[gydF4y2Ba3.gydF4y2Ba]及肺动脉高压[gydF4y2Ba4gydF4y2Ba].在肺动脉高压(PAH)中,内皮细胞和平滑肌细胞增殖、凋亡和代谢的改变导致血管重构和阻塞。由此产生的表型逐渐降低肺血管的可塑性、同一性和顺应性,从而增加肺血管阻力(PVR)和右心室(RV)的背压,导致右心室肥大,最终导致右心衰和死亡[gydF4y2Ba2gydF4y2Ba].目前,多环芳烃还没有治愈方法,所有现有的治疗方法只能减缓疾病进展,对死亡率影响有限。因此,开发新的变革性疗法的临床需求尚未得到满足。gydF4y2Ba
这种对慢性缺氧的程序性反应在一定程度上是由缺氧诱导因子(hif)调控的,hif属于一组基本螺旋-环-螺旋- per - arnt - sim (PAS)蛋白,其功能是响应氧气和其他应激的转录因子[gydF4y2Ba5gydF4y2Ba].hif作为异二聚体,由组成型表达的β-HIF-1b/ARNT亚基和氧调节的α-亚基(包括HIF1α和HIF2α)组成。每个亚基包含两个PAS结构域(PAS- a和PAS- b),有助于α-和β-异源二聚体复合物的稳定性。HIFα活性在翻译后由氧、脯氨酸羟化酶(PHDs)和泛素蛋白酶体系统的协同作用控制。在常氧条件下,博士羟基化HIFα,使泛素蛋白酶体系统结合并快速降解蛋白质。在缺氧条件下,不活跃的PHDs允许HIFα蛋白积累并与ARNT二聚,形成活跃的转录因子复合物。此外,HIFα的稳定性和功能可能受到异常活跃的细胞因子、生长因子、代谢物和PHD表达降低的影响,独立于氧气,导致伪缺氧表型[gydF4y2Ba6gydF4y2Ba- - - - - -gydF4y2Ba9gydF4y2Ba].这两种HIFα调控途径都有助于特发性PAH的发生和发展[gydF4y2Ba10gydF4y2Ba,gydF4y2Ba11gydF4y2Ba].gydF4y2Ba
HIF1α和HIF2α亚型在肺动脉高压(PH)中已被广泛研究。第一个直接证据来自于半合子小鼠gydF4y2BaHIF1gydF4y2Baα(gydF4y2Ba12gydF4y2Ba]或gydF4y2BaHIF2gydF4y2Baα(gydF4y2Ba13gydF4y2Ba].在这些模型中,慢性缺氧暴露后的肺部疾病进展显著延迟。随后的研究确定了肺血管系统中HIF2α的组织特异性表达,并发现HIF2α在内皮中高度表达[gydF4y2Ba14gydF4y2Ba,gydF4y2Ba15gydF4y2Ba].我们报道了肺内皮细胞HIF2α的遗传消融可以阻止与慢性缺氧诱导的PH相关的肺血管重塑的启动和发展[gydF4y2Ba16gydF4y2Ba].现在有几个小组报道了内皮细胞的丢失gydF4y2BaPHD2gydF4y2Ba随着闭塞性血管病变和严重ph值的发展,导致HIF2α的异常稳定性gydF4y2BaPHD2gydF4y2Ba而且gydF4y2BaHIF2gydF4y2Baα在该PH模型中也抑制了表型,提供了几乎完全的PH保护[gydF4y2Ba8gydF4y2Ba,gydF4y2Ba9gydF4y2Ba].这些小鼠基因操纵研究证实了HIF2α是驱动PH的主要HIFα亚型。此外,患有HIF2α功能获得性突变的患者或小鼠具有与PH相关的肺血流动力学升高,进一步证明了HIF2α的异常稳定性可引发这种疾病[gydF4y2Ba17gydF4y2Ba,gydF4y2Ba18gydF4y2Ba].尽管在揭示HIF2α在PH中的作用方面已经取得了实质性进展,但确定和开发合适的小分子抑制剂作为PH治疗的潜在候选物却花了近20年的时间。gydF4y2Ba
作为一种通过蛋白-蛋白相互作用激活基因表达的转录因子,HIF2α通常被认为是难以处理的小分子抑制。然而,SgydF4y2BacheuermanngydF4y2Baet al。gydF4y2Ba[gydF4y2Ba19gydF4y2Ba]导致发现HIF2α PAS-B结构域的内核具有疏水腔,可以结合变构破坏其二聚到ARNT的小分子,从而阻断转录活性。独立,ZgydF4y2Ba其次是音麦gydF4y2Baet al。gydF4y2Ba[gydF4y2Ba20.gydF4y2Ba]使用细胞筛选方法鉴定HIF2α抑制剂,结果发现了化合物76 (C76)。C76通过与铁调节蛋白结合抑制HIF2α的翻译。虽然后一种方法导致一种分子已用于少量临床前模型,但它只显示出微摩尔效力gydF4y2Ba在体外gydF4y2Ba,并没有被证明是口服生物可利用的。相比之下,前一种方法导致了一系列高选择性、口服生物可利用性的HIF2α抑制剂。这些直接的HIF2α抑制剂在啮齿动物癌症模型中是有效的,重要的是,在转移性透明细胞肾细胞癌(ccRCC)患者中表现出抗肿瘤活性,同时具有良好的安全性和耐受性[gydF4y2Ba21gydF4y2Ba- - - - - -gydF4y2Ba25gydF4y2Ba].因此,鉴于HIF2α是PH条件下肺血管重构的有效刺激因素,我们评估了一种直接的HIF2α抑制剂PT2567 [gydF4y2Ba25gydF4y2Ba]用于临床前PH模型。我们对啮齿类动物su5416/缺氧模型进行了血流动力学分析和生化分析,同时采用预防和干预策略,并将载药、PT2567或西地那非作为临床相关的比较化合物。在这里,我们证明了抑制HIF2α通过逆转肺血管重塑、减少循环促炎因子和恢复血浆亚硝酸盐水平来降低与严重PH发展相关的血流动力学参数,这表明抑制这一关键途径可能为PH治疗提供一种有前途的新策略。gydF4y2Ba
方法gydF4y2Ba
动物gydF4y2Ba
该研究由剑桥大学动物福利和伦理审查机构(AWERB)内政部项目许可证70/8850进行伦理审查后,受英国《动物(科学程序)1986年法》2012年修订条例的监管。所有动物都被安置在12小时/12小时的明暗循环中gydF4y2Ba没有限制地gydF4y2Ba标准的食物和水。功率计算确定了在80%功率,5%显著性水平下,每组检测25%效应变化所需的大鼠数量。su5416/缺氧预防研究的组数计算为n=10, su5416/缺氧和单野葡萄碱(MCT)干预研究的组数计算为n=15和n=14gydF4y2Ba
Sugen5416/缺氧大鼠PH模型gydF4y2Ba
雄性sd大鼠(~ 150-200 g;Charles River, Wilmington, MA, USA)给予su5416 (20 mg·kg)单次皮下注射gydF4y2Ba−1gydF4y2Ba;Tocris, Bristol, UK)在载液中(0.5%羧基甲基纤维素钠,0.4%聚山楂酸80,1%苄醇5%二甲基亚砜(DMSO) 5%聚乙二醇400;所有Sigma, Gillingham,英国),立即放入10%的氧气(OgydF4y2Ba2gydF4y2Ba并在缺氧状态下维持3周。根据治疗策略,被分配到预防方案的大鼠被随机分配到四个治疗组,并接受PT2567 (100 mg·kggydF4y2Ba−1gydF4y2Ba或300毫克·千克gydF4y2Ba−1gydF4y2Ba)口服灌胃每日1次或西地那非(30 mg·kg)gydF4y2Ba−1gydF4y2Ba)在低氧暴露期间,每天两次。干预组大鼠适应常氧24 h后,随机分为三组,使用PT2567 (100 mg·kggydF4y2Ba−1gydF4y2Ba口服灌胃,每日2次,西地那非(30 mg·kg)gydF4y2Ba−1gydF4y2Ba)口服灌胃,每日两次,持续3周。如前所述,测量心脏回波、右心室收缩压和右心室肥厚[gydF4y2Ba26gydF4y2Ba- - - - - -gydF4y2Ba28gydF4y2Ba].使用Vevo 3100成像系统测量心脏回波参数;Fujifilm VisualSonics公司,多伦多,加拿大)。获得左心室胸骨旁长轴视图,以显示左心室、主动脉和二尖瓣小叶。肺动脉可见,彩色多普勒确认正确的位置在最大流速内的脉冲波多普勒。测量肺动脉峰值流速,并测定右心室流出道速度时间间隔(RVOT-VTI)。测量肺加速时间(PA-AT,脉搏波多普勒记录血流开始到峰值速度的时间)和右心室射血时间(PA-ET,肺血流开始到结束的时间)。获得一个根尖四腔视图来量化血流谱和表征峰值三尖瓣反流速度(TRV)(如果存在)。使用公式PVR=10×TRV/RVOT-VTI计算PVR的估计值(伍兹单位(WU))。所有超声心动图均由同一个人进行,分析离线进行,该个人对治疗组不可见。 Afterwards, heart, lung and blood were taken for histological, gene expression and biochemical analysis.
MCT大鼠PH模型gydF4y2Ba
雄性(Sprague Dawley)大鼠(100-125克);Charles River)被随机分配到四组并注射gydF4y2Ba南卡罗来纳州。gydF4y2Ba对照组(0.9%生理盐水)为2 mL·kggydF4y2Ba−1gydF4y2Ba或40毫克·千克gydF4y2Ba−1gydF4y2Ba(2毫升·公斤gydF4y2Ba−1gydF4y2Ba) MCT (phl89251;),如前所述[gydF4y2Ba26gydF4y2Ba].mct或载体注射后14天,动物接受基线心脏回波检查(Vevo 3100成像系统),以确定疾病严重程度。超声心动图由同一个人进行,治疗组不可见。所有分析都是离线进行的。动物处理组如下:第1组:载体非疾病控制(0.9%生理盐水中的羧甲基纤维素);第2组:车体疾病控制(羧甲基纤维素0.9%生理盐水);第三组:hif2 α抑制剂干预(PT2567 100 mg·kggydF4y2Ba−1gydF4y2Ba)每日两次灌胃;第4组:西地那非干预(30 mg·kggydF4y2Ba−1gydF4y2Ba)口服灌胃,每日两次,连续两周。在给药期结束后,所有动物都进行心脏超声心动图检查,如前所述。gydF4y2Ba
肺血管形态测量gydF4y2Ba
肺充气固定gydF4y2Ba通过gydF4y2Ba用4%的多聚甲醛在15-20厘米恒定的流体压力下浸泡气管5分钟。对预防方案的肺进行弹性蛋白/伊红染色,对干预方案的肺进行苏木精和伊红染色,弹性Van Gieson (EVG)染色以评估组织学(MSD/BDH, lutworth,英国)。组织学检查结果由一名独立的盲病理学家评估,他对肺切片的血管周围/血管炎症、血管周围纤维化和小动脉平滑肌肥大进行评分。结果记录为0(正常),1(最小),2(轻度),3(中度),4(标记)。肺小动脉平滑肌肥大的评分基于肌壁的厚度和发现的程度(受影响的小动脉的表观数量)。此外,对来自干预方案的一系列肺切片进行抗平滑肌α-肌动蛋白(α-SM肌动蛋白;DakoCytomation, Ely, UK)、von Willebrand因子和抗髓过氧化物酶(MPO) (DakoCytomation)和抗ki67 (Abcam, Cambridge, UK)来评估小肺动脉(≤50 μm)的肌肉化程度、细胞增殖和MPO阳性细胞。抗体染色使用3-3 '盐酸二氨基联苯底物(DakoCytomation),并用Carrazzi haematoxylin (Bios, Shelmersdale, UK)进行反染色。使用Image J软件(MediaCybernetics, Bethesda, MD, USA)测量血管内侧厚度。组织样本由盲病理学家独立编码和量化。gydF4y2Ba
血液学的分析gydF4y2Ba
抗凝血使用Vet abc血液学分析仪(Horiba,京都,日本)根据制造商的说明进行分析。抗凝全血1500倍离心后分离血浆,进行细胞因子和生长因子谱分析gydF4y2BaggydF4y2Ba5分钟后冷冻至- 80°C。gydF4y2Ba
亚硝酸盐分析gydF4y2Ba
血液样本被离心分离血浆,并通过一个带有10 kda截止过滤器的柱。根据制造商的说明,使用NOA 280i (Sievers, GE Healthcare, Chicago, IL, USA)分析所有样品的亚硝酸盐含量。gydF4y2Ba
RNA分析gydF4y2Ba
使用tri试剂(Sigma)从组织中分离总RNA,然后使用RNeasy柱试剂盒(Qiagen, Venlo,荷兰)进行RNA清理和DNase消化。根据制造商说明书,用1 μg总RNA使用Superscript IV (Invitrogen, Waltham, MA, USA)进行第一链合成。采用定量(q)PCR (One-Step Plus Real-Time PCR System;Life Technologies, Waltham, MA, USA),并在SYBR-Green主混合(Roche, Basel, Switzerland)和Qiagen的相关引物中扩增。相对基因表达量校正为管家基因β-actin和BgydF4y2Ba2gydF4y2BaM。gydF4y2Ba
颈动脉体组织学gydF4y2Ba
颈动脉分叉解剖,用4%多聚甲醛(Santa Cruz Biotechnology, Dallas, TX, USA)固定2小时,冷冻保存(30%蔗糖在PBS中),冷冻切片(10 μm厚;Bright Instruments,卢顿,英国)。酪氨酸羟化酶(TH),采用兔抗TH (1:5000;Novus Biologicals,阿宾顿,英国;NB300-109)一抗和Alexa-Fluor 568偶联抗兔IgG (1:500;ThermoFisher, Waltham, MA, USA)。gydF4y2Ba
颈动脉体面积测量采用显微照片(Leica DM-RB, Wetzlar, Gernany),使用Fiji Software从整个颈动脉体中间隔60µm的切片中截取[gydF4y2Ba29gydF4y2Ba].颈动脉体容量根据Cavalieri原理估算,如前所述[gydF4y2Ba30.gydF4y2Ba].组织样本由盲法研究者编码和量化。gydF4y2Ba
PT2567大鼠药代动力学gydF4y2Ba
PT2567悬浮于甲基纤维素Tween-80中(0.5%甲基纤维素,0.5% Tween-80在水中),大鼠(每剂量水平n=3)灌胃。分别于给药后0.25、0.5、1、2、4、8、12和24 h采集血样。血浆PT2567浓度采用Pharsight WinNonlin (Certara, Princeton, NJ, USA)非隔室法测定。采用非区室模型对数据进行分析。gydF4y2Ba
血外生长内皮细胞分离培养gydF4y2Ba
根据剑桥研究伦理委员会批准的方案(REF:11/EE/0297),从先前诊断为PAH的参与者的血液中分离出血液生长内皮细胞(BOECs),或从Addenbrooke's教学医院剑桥大学医院NHS基金会信托(Cambridge, UK)的正常健康志愿者的血液中分离出来。gydF4y2Ba表1gydF4y2Ba包含PAH患者的特征和健康志愿者的信息。通过Ficoll密度梯度离心从60 mL静脉血中分离出单个核细胞,并在内皮选择性培养基(EGM2;Lonza Biologics,巴塞尔,瑞士)补充10%胚胎干细胞合格的胎牛血清和额外的生长因子(EGM2子弹试剂盒;Lonza生物制剂)。2-3周后出现boec,汇合时产生。gydF4y2Ba
肺动脉高压(PAH)患者特征的血液外生长内皮细胞的分离gydF4y2Ba
boec最多培养5代。对于BOEC低氧暴露实验,细胞被转移到Baker Ruskinn低氧室(brigend, UK),预低氧介质被转移到每个孔中,有或没有抑制剂。细胞在1% O浓度下培养gydF4y2Ba2gydF4y2Ba对于实验结果中所示的时间gydF4y2Ba
细胞培养gydF4y2Ba
786-O和Hep3B细胞株购自ATCC (American Type Culture Collection, Manassas, VA, USA)。细胞在含10%胎牛血清的DMEM中培养。人肺动脉内皮细胞(hPAECs)购自PromoCell公司(Heidelberg, Germany),在内皮细胞生长培养基中培养。PT2567处理细胞镀成六孔板。加入溶解在DMSO中的PT2567,当培养物与最终浓度为>0.1%的DMSO达到汇合时。对于缺氧处理的细胞,细胞培养基暴露于1% OgydF4y2Ba2gydF4y2Ba, 5%的二氧化碳(罗斯金)12小时后转移到细胞。在加入PT2567后,细胞在治疗期间保持在缺氧状态下。gydF4y2Ba
精氨酸酶活性测定gydF4y2Ba
boec是按照前面描述的那样准备的。尿素产量以蛋白质浓度归一化[gydF4y2Ba31gydF4y2Ba].gydF4y2Ba
可拆卸的实验gydF4y2Ba
来自两个对照组志愿者的对照hPAECs和人boec分别使用含有三种不同的短发夹(sh) rna的慢病毒颗粒转导,分别靶向人HIF-1α和HIF-2α mRNA。shRNA序列根据以下标准从RNAi联盟(TRC)中选择:shRNA似乎具有较少的潜在脱靶结合位点,并由MISSION shRNA库(Sigma-Aldrich)验证。选择的单个克隆id为TRCN0000003808 (HIF1α)、TRCN0000003806 (HIF2α)和TRCN0000342501。每个shRNA的寡核苷酸被退火并克隆到pLKO中。1质粒遵循TRC建议。通过测序检查阳性菌落。为了产生慢病毒载体,lentii - x 293T细胞(Clontech, Saint-Germain-en-Laye, France)与单个pLKO共转染。1、pCMV-dR8.91和pMD2。Gplasmids using Lipofectamine 2000 (Thermo-Fisher Scientific) according to the manufacturer's protocol. Lentiviral particles were collected 48 h after transfection and used to transduce hPAECs and BOECs overnight. Next, fresh media was added and the cells were incubated in normoxia for 2 days then in 1% O2gydF4y2Ba的气氛。gydF4y2Ba
扩散分析gydF4y2Ba
boec被镀在24孔板中,每孔2万个细胞。细胞接受含/不含PT2567的完整EGM-2MV培养基。台盼蓝排除后第0、2、4、6天计数细胞。gydF4y2Ba
BOEC内皮网络形成检测gydF4y2Ba
来自PAH患者和健康志愿者的boec增长到90%合流。将boec转移到对照培养基或含有1 μM PT2567的培养基中,然后在Matrigel基质涂层的12孔板中播种约75000个细胞。治疗后20小时,使用相位对比显微镜(徕卡MZ16)拍摄图像。使用Image J软件(MediaCybernetics, Bethesda, MD, USA)确定管道网络长度和网络环路。gydF4y2Ba
HIF2α和ARNT的共免疫沉淀gydF4y2Ba
6孔板细胞在1 mL细胞裂解液(Tris-HCl 20 mmol·L)中裂解gydF4y2Ba−1gydF4y2Ba, pH 7.5, Triton X-100 1%, NaCl 150 mmol·LgydF4y2Ba−1gydF4y2Ba,甘油5%,EDTA 1 mmol·LgydF4y2Ba−1gydF4y2Ba,二硫苏糖醇1 mmol·LgydF4y2Ba−1gydF4y2Ba每10 mL罗氏蛋白酶抑制剂全片1片)。小鼠单抗抗人ARNT (sc-55526;Santa Cruz Biotechnology)或1 μg抗HIF2α抗体(Ab199;Abcam)和50 μL Protein AG Beads(50%浆液为裂解缓冲液;Santa Cruz Biotechnology)被添加到清除的细胞裂解液中。试管在4°C旋转16小时。经冷裂解缓冲液洗涤后,用SDS凝胶电泳分离珠结合蛋白,并用特异性抗体进行Western blotting。gydF4y2Ba
等温滴定量热法gydF4y2Ba
如前所述,人HIF2α-B的表达和纯化[gydF4y2Ba19gydF4y2Ba].大鼠HIF2α与人HIF2α- b仅相差3个氨基酸。因此,将人HIF2α-B的表达载体(T262L、I 265V和I326V)在这些残基上进行突变,得到大鼠HIF2α-B。PT2567和PAS-B结构域之间的结合亲和力是在iTC200系统上使用等温滴定量热法测定的(GE Healthcare)。PAS-B 0.5 mmol·LgydF4y2Ba−1gydF4y2Ba滴定至26 μmol·LgydF4y2Ba−1gydF4y2Ba或25 μmol·LgydF4y2Ba−1gydF4y2Ba在含有20 mmol·L的缓冲液的细胞中,分别含有人或大鼠的PT2567gydF4y2Ba−1gydF4y2BaTris-HCl, pH 8.0, 150 mmol·LgydF4y2Ba−1gydF4y2BaKCl和1% DMSO。gydF4y2Ba
统计分析gydF4y2Ba
所有数据均为均数±gydF4y2BasdgydF4y2Ban个独立实验,除非另有说明。采用单因素方差分析和Tukey多元比较检验评估组间差异。所有数据均为正态分布。如果不是正态分布,除非另有说明,非参数Kruskal-Wallis检验与Dunn的多重比较检验一起使用。采用log-rank (Mantel-Cox)检验对MCT死亡率数据进行统计分析。使用GraphPad Prism进行分析(Mac OS版本7.0 c;圣地亚哥,CA,美国)。p值<0.05被认为是显著的。gydF4y2Ba
结果gydF4y2Ba
HIF2α/ARNT二聚体中断gydF4y2Ba
我们首先用等温滴定量热法测定PT2567对人和大鼠HIF2α PAS-B结构域的结合亲和力。这种HIF2α PAS-B结构域相互作用的解离常数对人和大鼠分别为30 nM和29 nM (gydF4y2Ba补充图S1a及bgydF4y2Ba).接下来,我们在786-O细胞中通过共免疫沉淀试验评估了HIF2α/ARNT相互作用/二聚体的破坏。选择这个ccRCC细胞系,因为它在常氧条件下组成性地表达活性HIF2α,而没有HIF1αgydF4y2BaHIF1gydF4y2Baα基因。最初,我们在PT2567处理的786-O细胞的裂解液中免疫沉淀ARNT。免疫印迹显示,在PT2567处理后,ARNT蛋白与HIF2α的共沉淀以浓度依赖的方式减少,表明HIF2α/ARNT二聚体的形成被破坏(gydF4y2Ba补充图S1cgydF4y2Ba).gydF4y2Ba
抑制HIF2α转录gydF4y2Ba
我们通过qPCR分析PT2567对hif2 α依赖靶基因转录激活的影响。PT2567处理786-O细胞显著降低mRNA的表达gydF4y2BaGLUT1gydF4y2Ba,gydF4y2Ba促红细胞生成素gydF4y2Ba,gydF4y2BaCCND1gydF4y2Ba,gydF4y2BaPAI1gydF4y2Ba而且gydF4y2BaVEGFAgydF4y2Ba,以浓度依赖的方式(gydF4y2Ba补充图S2a-egydF4y2Ba).我们进一步评估了PT2567在Hep3B肝癌细胞中的特异性。缺氧导致该细胞系中HIF1α和HIF2α蛋白的积累。PT2567处理Hep3B细胞仅降低缺氧诱导的HIF2α靶基因表达;gydF4y2Ba促红细胞生成素gydF4y2Ba而且gydF4y2BaPAI1gydF4y2Ba,对HIF1α靶基因表达无影响;gydF4y2Ba的基因gydF4y2Ba或gydF4y2BaPGK1gydF4y2Ba(gydF4y2Ba补充图S2f-igydF4y2Ba).这些重要数据证实了PT2567的靶点特异性,仅破坏HIF1α/ARNT相互作用,而不影响HIF1α/ARNT的转录活性。gydF4y2Ba
PT2567在hpaec中的验证gydF4y2Ba
接下来,我们评估了PT2567在原发性hPAECs中的活性和特异性。我们最初通过短发夹敲低这些转录因子(shRNA-HIF1α或HIF2α),然后定时暴露于缺氧,确认了这些细胞中的HIF1α和HIF2α靶基因(gydF4y2Ba补充图S3a-fgydF4y2Ba).我们证实了HIF2α对PAI-1的近乎完全抑制和对VEGF和GLUT1 mRNA表达的部分抑制gydF4y2Ba−−/gydF4y2Ba对HIF1α靶LDHA mRNA表达无影响的PAECs。用PT2567处理hPAECs显著降低缺氧诱导的HIF2α靶基因和ph相关基因的表达:gydF4y2BaGLUT1gydF4y2Ba,gydF4y2BaVEGFgydF4y2Ba,gydF4y2BaCXCL12gydF4y2Ba,gydF4y2Ba趋化因子受体CXCR4gydF4y2Ba,gydF4y2BaICAM1gydF4y2Ba,gydF4y2Ba希利gydF4y2Ba,gydF4y2BaPAI1gydF4y2Ba而且gydF4y2BaAPLNgydF4y2Ba,并恢复gydF4y2BaID1gydF4y2Ba以浓度依赖的方式表达,而不影响HIF1α靶基因转录本gydF4y2BaLDHAgydF4y2Ba而且gydF4y2Ba的基因gydF4y2Ba(gydF4y2Ba补充图S3g-qgydF4y2Ba).这些结果证实PT2567在不影响hPAECs中HIF1α/ARNT转录活性的情况下破坏HIF1α/ARNT相互作用。gydF4y2Ba
PT2567可以阻止PH的产生和发展gydF4y2Ba
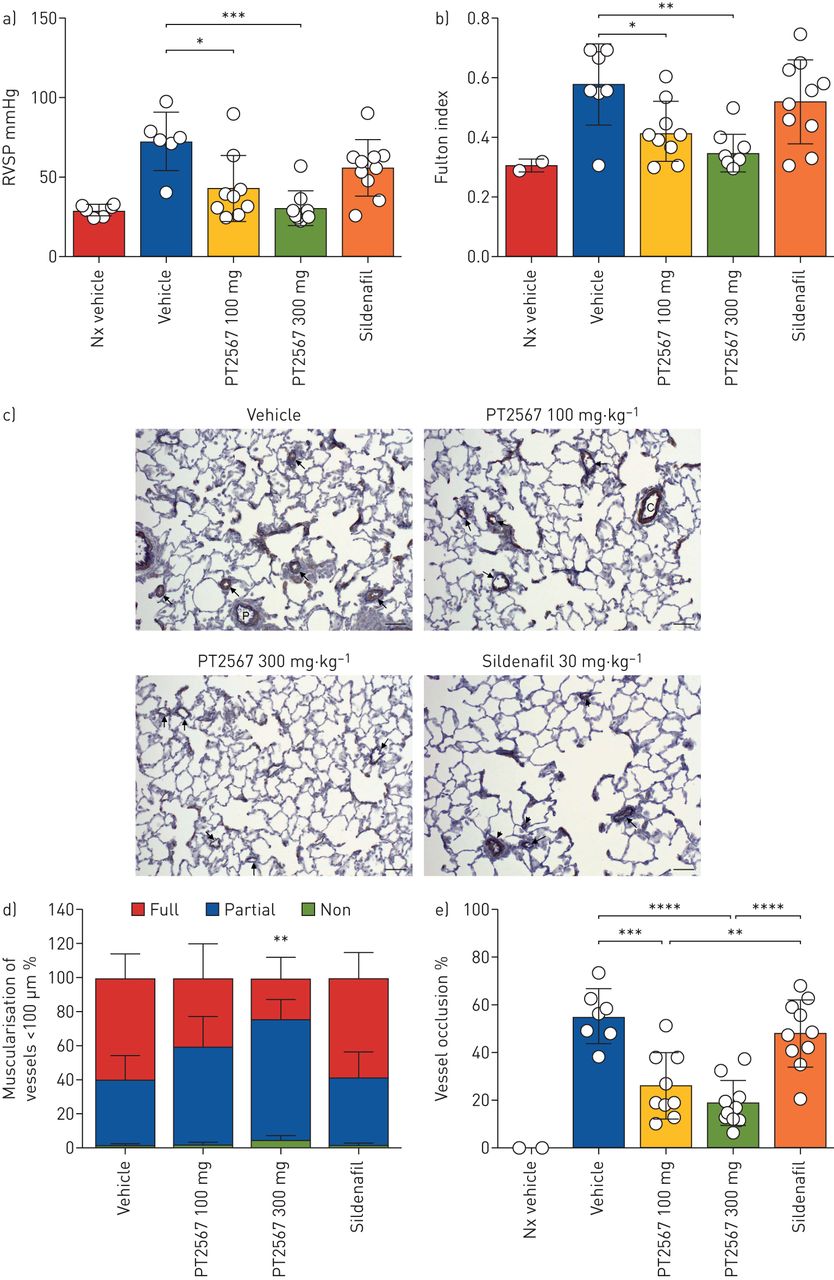
接下来,我们测定了PT2567的大鼠血浆药动学谱。定时血浆分析均为100 mg·kggydF4y2Ba−1gydF4y2Ba300毫克·千克gydF4y2Ba−1gydF4y2Ba口服灌胃达到良好的暴露(gydF4y2Ba补充图S4agydF4y2Ba).我们使用大鼠su5416/Hx模型初步研究了PT2567对PH发展的影响。大鼠随机分为4个治疗组,接受单剂量gydF4y2Ba南卡罗来纳州。gydF4y2Ba剂量的su5416,然后置于10% O中gydF4y2Ba2gydF4y2Ba4周。大鼠注射PT2567 (100 mg·kggydF4y2Ba−1gydF4y2Ba或300毫克·千克gydF4y2Ba−1gydF4y2Ba)每日1次或西地那非(30 mg·kg)gydF4y2Ba−1gydF4y2Ba)每日两次口服灌胃(gydF4y2Ba补充图S4bgydF4y2Ba).通过测量右心室收缩压(RVSP)来评估肺动脉高压的发展和进展。两组PT2567治疗组的RVSP均显著降低,且呈剂量依赖性(100和300 mg·kg组分别为43.22±20.72 mmHg和30.78±11.07 mmHggydF4y2Ba−1gydF4y2Ba(72.83±18.14 mmHg)。相比之下,西地那非治疗组RVSP仅表现出中度钝化,与对照组无显著差异(56.10±17.55 mmHg) (gydF4y2Ba图1一个gydF4y2Ba).此外,pt2567处理动物右心室肥厚(RVH)的富尔顿指数(RV/LV+隔膜)较低(100和300 mg·kg时分别为0.421±0.100和0.350±0.06)gydF4y2Ba−1gydF4y2Ba对照组(0.578±0.13)(gydF4y2Ba图1 bgydF4y2Ba).西地那非治疗对RVH的保护作用很小(0.5220±0.14)(gydF4y2Ba图1 bgydF4y2Ba).与前面描述的数据一致,RVSP与RVH (rgydF4y2Ba2gydF4y2Ba=0.666, p<0.0001) (gydF4y2Ba补充图S4cgydF4y2Ba).所有治疗组的最终体重均未偏离车辆对照组(gydF4y2Ba补充图S4dgydF4y2Ba).gydF4y2Ba
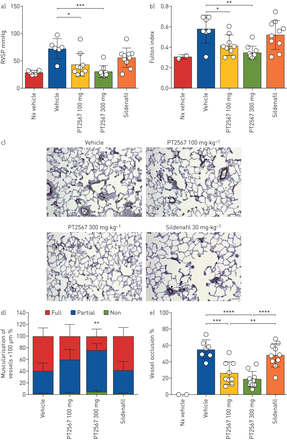
低氧诱导因子(HIF)2α-抑制剂对大鼠SU-5416/Hx预防模型肺动脉高压(PAH)发展的影响a)右心室收缩压(RVSP);b) Sprague Dawley Su5416/Hx大鼠右心室肥厚(RVH)显示富尔顿指数(右心室/左心室+鼻中隔重量比),每天给药1次,PT2567 100 mg·kggydF4y2Ba−1gydF4y2Ba(n=9)或300 mg·kggydF4y2Ba−1gydF4y2Ba(n=9),或30 mg·kggydF4y2Ba−1gydF4y2Ba西地那非(n = 10);C) α-平滑肌肌动蛋白免疫染色的代表性显微照片;箭头指向(载具)完全肌肉化的血管;(PT2567 100 mg·kggydF4y2Ba−1gydF4y2Ba300毫克·千克gydF4y2Ba−1gydF4y2Ba)部分肌肉化血管;(西地那非)短箭头:完全肌肉化,长箭头:远端血管部分肌肉化;比例尺=50 μm;D)肺切面外周肺血管肌化量化的堆叠柱状图(无平滑肌环、平滑肌环、全平滑肌环);E)小动脉血管(>5 μm)闭塞的测定,以完全闭塞的百分比表示。数据以均数±表示gydF4y2Basd。gydF4y2Ba* *: *: p < 0.05, p < 0.001, * * *: p < 0.0001, * * * *: p < 0.00001(单向方差分析)。gydF4y2Ba
肺组织病理学的评估被提出gydF4y2Ba补充表S1agydF4y2Ba.平滑肌肥大伴远端肺血管(≤50 μm)、血管周围炎症和血管周围纤维化评分以灌胃剂组最高(5.42);同时,PT2567 100 mg·kg处理的动物评分呈剂量依赖性下降gydF4y2Ba−1gydF4y2Ba(3.66)和300 mg·kggydF4y2Ba−1gydF4y2Ba(2.55)。西地那非组得分高于pt2567组(4.00)。此外,我们量化了血管肌肉化,并观察到与对照剂或西地那非治疗组相比,PT2567治疗动物中完全肌肉化的小动脉比例下降,同时非肌肉化的小动脉比例增加(gydF4y2Ba图1 cgydF4y2Bad和gydF4y2Ba补充图S4e-ggydF4y2Ba).gydF4y2Ba
接下来,我们评估小动脉闭塞的程度。血管(≤50 μm)被确定为完全闭塞或腔内开放。我们观察到PT2567- (100 mg·kggydF4y2Ba−1gydF4y2Ba300毫克·千克gydF4y2Ba−1gydF4y2Ba)治疗组与对照剂和西地那非治疗组(gydF4y2Ba图1 egydF4y2Ba).gydF4y2Ba
PT2567干预可降低已确定PH值下的PVRgydF4y2Ba
接下来,我们评估了PT2567 (100 mg·kggydF4y2Ba−1gydF4y2Ba)会影响su5416/缺氧大鼠肺血管功能和PH疾病进展。如gydF4y2Ba补充图S5agydF4y2Ba时,所有大鼠均接受预防方案中所述的相同su5416/缺氧启动过程。从缺氧室取出24小时后评估血流动力学基线。这些数据为该研究的干预组提供了疾病严重程度的参考点。其余大鼠随机分为3组,给予PT2567 (100 mg·kg)对照治疗gydF4y2Ba−1gydF4y2Ba)或西地那非(30 mg·kggydF4y2Ba−1gydF4y2Ba)口服灌胃,每日两次,持续3周。常氧对照组给予载药或PT2567治疗3周。干预组在最后一次灌胃后16-18小时完成血流动力学分析。我们认为,这种时间延迟有助于评估肺血管瘀血,而不是化合物的血管舒张调节特性。我们报告PT2567组的RVSP(64.85±16.86 mmHg, p<0.05)显著低于对照组(89.82±27.92 mmHg) (gydF4y2Ba图2一个gydF4y2Ba).与载体治疗大鼠相比,西地那非(77.29±26.56 mmHg)没有降低RVSP。此外,pt2567处理组的富尔顿指数(RVH)(0.4074±0.079)明显低于单药组(04906±0.098)(gydF4y2Ba图2 bgydF4y2Ba);RVSP与RVH (rgydF4y2Ba2gydF4y2Ba=0.7066, p<0.0001) (gydF4y2Ba补充图S5bgydF4y2Ba).值得注意的是,PT2567治疗显著影响心排血量的恢复(gydF4y2Ba图2 cgydF4y2Ba)与疾病控制或载体处理的PH大鼠相比。心脏超声分析PA-AT/PA-ET比值(gydF4y2Ba图2 dgydF4y2Ba)和三尖瓣返流(TVR) (gydF4y2Ba图2 egydF4y2Ba)提示肺动脉背压降低,有可能恢复舒张功能。我们分析了PA流量-时间曲线中PA流速的收缩中期下降,即PH大鼠模型中的“缺口”。缺口持续时间此前曾有报道,并与PH疾病严重程度有关[gydF4y2Ba32gydF4y2Ba].我们报告了notch持续时间与肺动脉压(估计PAP)之间的相关性(gydF4y2Ba补充图5cgydF4y2Ba),与载体处理大鼠相比,PT2567干预减少了缺口持续时间(gydF4y2Ba补充图5d-egydF4y2Ba).gydF4y2Ba
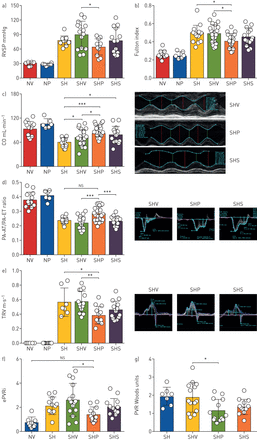
低氧诱导因子(HIF)2α抑制剂降低大鼠Su5416/Hx肺动脉高压(PH)模型肺血管阻力(PVR)评估a)右心室收缩压(RVSP)和b)右心室肥厚(RVH;富尔顿指数);c)大鼠心排血量(CO)。d)肺加速时间(PA-AT)/右心室射血时间(PA-ET)比值的心脏回波分析及代表性显微照片;e)三尖瓣返流(TVR)速度,具有代表性的飞行器控制显微照片,PT2567或西地那非。肺血管阻力计算f)估计- pvr指数;g)伍兹单位。数据以均数±表示gydF4y2BasdgydF4y2Ba.NV:常氧载具(n=8);NP: normoxia / PT2567;SH: Su/Hx-3w (n=12);SHV: Su/Hx-vehicle (n=15);SHP: Su/Hx-PT2567 (n=15);SHS: Su/ hx -西地那非(n=14)。* *: *: p < 0.05, p < 0.001, * * *: p < 0.0001(单向方差分析)。gydF4y2Ba
PVR与肺压和心排血量耦合,被认为能更好地反映PH的病理生理,因为它在很大程度上是外周血管重构的结果。PVR由两种导尿方法计算(估计PVR指数=估计PAP -舒张末期压/心脏指数)[gydF4y2Ba33gydF4y2Ba]和心脏回波(伍兹指数(WU)=10×TVR/RVOT−VTI) (PVR计算中使用的其他参数见gydF4y2Ba补充图S5f-igydF4y2Ba).PT2567干预组PVR指数(1.486±0.59,p<0.05)和Woods指数(1.167±0.615,p<0.05)分别显著低于对照组(2.605±1.370和1.897±0.783)(gydF4y2Ba图2 fgydF4y2Ba抑制HIF2α也使RVOT-VTI恢复到与常氧对照相似的值(gydF4y2Ba补充图S5jgydF4y2Ba),而所有其他su5416/缺氧处理组的RVOT-VTI均显著降低。与常氧对照相比,PT2567或西地那非对左心血压和心率无影响(gydF4y2Ba补充图S5k-lgydF4y2Ba).值得注意的是,恒氧-载药组和恒氧- pt2567组在所有血流动力学分析中没有显示出任何差异。gydF4y2Ba
PT2567干预可减少肺血管重塑gydF4y2Ba
我们评估了一系列膨胀肺的组织学切片对血管重塑的影响。肺切片用EVG染色(gydF4y2Ba图3一gydF4y2Ba)、血管性血友病因子及免疫染色α-平滑肌肌动蛋白(SMA)、细胞增殖标记物Ki67 (gydF4y2Ba图3 egydF4y2Ba)及MPO (gydF4y2Ba补充图S6g-hgydF4y2Ba).肺组织病理学评分见gydF4y2Ba补充表S1bgydF4y2Ba.与su5416/低氧载体组相比,PT2567和西地那非干预组平滑肌肥大、血管周围/血管炎症和血管周围纤维化的发生率均降低。常氧组、su5416/缺氧组、su5416/缺氧组、su5416/缺氧组、su5416/缺氧- pt2567 100 mg·kg组平滑肌肥厚、血管周围/血管炎症、纤维化均分分别为0.66、5.70、8.90、4.70、5.90gydF4y2Ba−1gydF4y2Ba和su5416/hypoxia-sildenafil。gydF4y2Ba
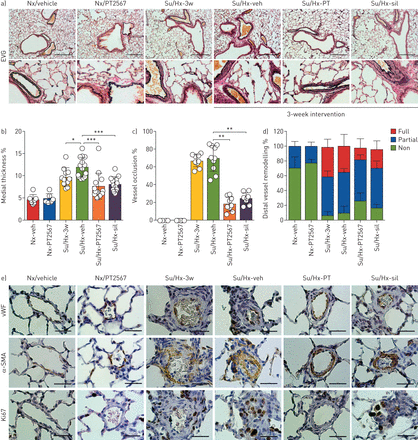
PT2567减少肺血管重塑。a)弹性van Gieson (EVG)染色肺切片代表性显微照片(比例尺=0.30 mm);B)评估肺动脉壁厚占管腔直径的百分比;C)小动脉血管(>50 μm)闭塞的测定,以完全闭塞的百分比表示;D)量化非肌化动脉、部分肌化动脉和完全肌化动脉占肺泡壁动脉和导管动脉总数的百分比;e)血管性血友病因子(vWF)、α-平滑肌肌动蛋白(SMA)和Ki67免疫染色的肺切片代表性显微照片(比例尺=0.30 mm)。数据以均数±表示gydF4y2Basd。gydF4y2BaNx-veh (n = 8)、Nx / PT2567 (n = 6),苏/ Hx-3w (n = 12),苏/ Hx-veh (n = 15),苏/ Hx-PT2567 (n = 11),苏/ Hx-sil (n = 14)。Nx: normoxia;阿明费:车辆;苏/ Hx: su5416 /缺氧;sil:西地那非。* *: *: p < 0.05, p < 0.001, * * *: p < 0.0001(单向方差分析)。gydF4y2Ba
我们评估了较大支气管相关肺血管的内侧增厚作为新内膜病变的衡量标准。大鼠暴露于su5416/缺氧3周后,与常氧对照相比,内侧厚度显著增加,并且在su5416/缺氧对照中注意到进一步的重塑(gydF4y2Ba图3 bgydF4y2Ba).相比之下,PT2567和西地那非干预都显示出相对于su5416/低氧载体的内侧厚度显著降低(gydF4y2Ba图3 bgydF4y2Ba).接下来,我们评估小动脉闭塞的程度。血管(≤50 μm)被确定为完全闭塞或腔内开放。与su5416/缺氧或su5416/缺氧载体相比,PT2567和西地那非干预显著降低了血管闭塞程度(gydF4y2Ba图3 cgydF4y2Ba).此外,来自su5416/hypoxia-vehicle的肺切片显示与肺血管相关的弹性蛋白和α-SMA增加(gydF4y2Ba图3 egydF4y2Ba,gydF4y2Ba补充图S6agydF4y2Ba).这被PT2567干预部分抑制,与su5416/低氧载体组相比,远端血管中α-SMA沉积大幅减少,特别是完全肌肉化的血管(gydF4y2Ba图3 dgydF4y2Ba,gydF4y2Ba补充图S6b-dgydF4y2Ba).增殖标志物(Ki67)的定量表达进一步证实PT2567和西地那非都能减少su5426/缺氧诱导的肺血管增殖(gydF4y2Ba补充图S6egydF4y2Ba)和更具体地说,PAEC增殖(gydF4y2Ba补充图S6fgydF4y2Ba).最后,肺切片中MPO的定量显示,与su5426/缺氧-3w和su5426/缺氧-载体动物相比,PT2567处理大鼠血管周围区域的MPO免疫反应性降低,而血管内区域的MPO免疫反应性没有变化(gydF4y2Ba补充图S6g,hgydF4y2Ba).gydF4y2Ba
综上所述,这些数据表明,PT2567抑制HIF2α功能可以减少肺血管重塑和与PH模型相关的心血管功能的动态变化。gydF4y2Ba
PT2567调节ph相关基因表达并使血浆亚硝酸盐水平正常化gydF4y2Ba
在确定PT2567在肺血管重构方面的差异后,我们接下来研究了全肺样本中与糖酵解、炎症和肺重构相关的已知HIFα靶基因的肺表达谱。在PH模型中,对于HIF2α的整体抑制作用以及对肺多细胞成分的影响,我们知之甚少。考虑到PT2567的全身给药,分析整个肺被认为是有利的。PT2567干预使已知HIF2α靶基因的表达正常化,gydF4y2Baglut1gydF4y2Ba而且gydF4y2Baca9gydF4y2Ba与su5416/低氧车辆控制相比(gydF4y2Ba补充图S7a,bgydF4y2Ba),对HIF1α靶基因表达影响不大,gydF4y2BaldhagydF4y2Ba而且gydF4y2Bapgk1gydF4y2Ba(gydF4y2Ba补充图S7c,dgydF4y2Ba).Su5416/缺氧显著增加炎症靶点基因表达gydF4y2Bacxcl12gydF4y2Ba和受体gydF4y2Ba趋化因子受体cxcr4gydF4y2Ba,以及内皮粘附分子gydF4y2Baicam1gydF4y2Ba而且gydF4y2Ba希利。gydF4y2BaPT2567干预降低了近常氧状态下非疾病动物的基因表达(gydF4y2Ba补充图S7e-hgydF4y2Ba).接下来,我们分析了与PH相关的信号靶点:gydF4y2BaaplngydF4y2Ba,gydF4y2Ba最长gydF4y2Ba,细胞循环gydF4y2Baccnd1gydF4y2Ba,gydF4y2Bapai1gydF4y2Ba和血管活性的gydF4y2Baedn-1gydF4y2Ba都减少了gydF4y2Baid1gydF4y2BaPT2567干预后恢复的表达(gydF4y2Ba补充图S7i-ngydF4y2Ba).西地那非使表达正常化gydF4y2Baglut1gydF4y2Ba,gydF4y2Bacxcl12gydF4y2Ba而且gydF4y2Baid1gydF4y2Ba,但不影响任何其他基因的表达。gydF4y2Ba
考虑到PAH病理生理会增加RVH,通常导致右心衰和死亡,我们接下来研究了结构和应激相关基因在右心衰组织中的表达。PT2567显著降低肌球蛋白重链-7的表达(gydF4y2Bamyh7gydF4y2Ba)和肌动蛋白α -1 (gydF4y2Baacta1gydF4y2Ba)和肌球蛋白轻链-3 (gydF4y2Bamyl3gydF4y2Ba)趋于正常表达(p=0.065非参数分析),与su5416/低氧载体处理大鼠相比(gydF4y2Ba补充图S8a-cgydF4y2Ba).心脏应激基因利钠肽A (gydF4y2BanppagydF4y2Ba)、利钠肽B (gydF4y2BanppbgydF4y2Ba)及膜联蛋白A5 (gydF4y2Baanxa5gydF4y2Ba),而纤维化的目标是1型胶原A1 (gydF4y2Bacol1a1gydF4y2Ba)第三型a1 (gydF4y2Bacol3a1gydF4y2Ba)和金属蛋白酶组织抑制剂2 (gydF4y2Batimp2gydF4y2Ba)在PT2567干预后减少(gydF4y2Ba补充图S8d-igydF4y2Ba);西地那非只降低了gydF4y2Bacol3a1gydF4y2Ba而且gydF4y2Batimp2gydF4y2Ba.gydF4y2Ba
接下来,我们分析了su5416/缺氧大鼠模型中细胞因子、心脏应激标志物和亚硝酸盐的血浆样本。PT2567干预显著降低血浆肿瘤坏死因子(TNF -α)浓度(gydF4y2Ba图4一gydF4y2Ba)和增加免疫抑制白介素-10 (gydF4y2Ba图4 bgydF4y2Ba)与su5416/低氧载体大鼠相比。其他血浆细胞因子包括IL-13, IL-4, IL-5,干扰素-γ, IL-1β和IL-8也被测量(gydF4y2Ba表2gydF4y2Ba),但与su5416/低氧载体对照相比未达到显著性。gydF4y2Ba
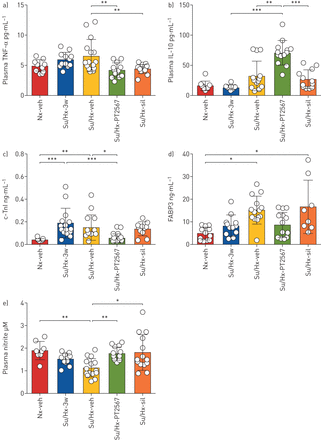
低氧诱导因子(HIF)2α抑制恢复血浆亚硝酸盐水平。a - d)血浆细胞因子和心脏应激因子分析a)肿瘤坏死因子(TNF)-α, b)白细胞介素(IL)-10, (Nx-veh n=8, Su/Hx n=12, Su/Hx-veh n=15, Su/Hx- pt2567 n=11, Su/Hx-西地那非n=14)。c) c-肌钙蛋白- i (c- tni), d)脂肪酸结合蛋白-3 (FABP3) (Nx/Veh n=6, Su/Hx-3w n=6, Su/Hx-veh n=8, Su/Hx-PT2567 n=8, Su/ hx -西地那非n=8)。e)用一氧化氮分析仪(Sievers)测定血浆亚硝酸盐(Nx/veh n=8, Su/Hx-3w n=12, Su/Hx-veh n=15, Su/Hx-PT2567 n=11, Su/ hx -西地那非n=14)。数据以均数±表示gydF4y2Basd。gydF4y2BaNx: normoxia;阿明费:车辆;苏/ Hx: su5416 /缺氧;sil:西地那非。* *: *: p < 0.05, p < 0.001, * * *: p < 0.0001(单向方差分析)。gydF4y2Ba
血浆细胞因子分析gydF4y2Ba
我们还注意到,与su5416/缺氧载体相比,pt2567治疗组血浆心脏应激标志物c-肌钙蛋白- i和脂肪酸结合蛋白-3的正常化(gydF4y2Ba图4 cgydF4y2Bad)。gydF4y2Ba
与PH疾病严重程度一致,我们观察到su5416/缺氧组血浆亚硝酸盐(1.121±0.414 μM)明显低于常氧组(1.889±0.404 μM)。值得注意的是,PT2567和西地那非均能使血浆亚硝酸盐浓度恢复到接近常氧霞-对照水平(分别为1.756±0.294 μM和1.798±0.795 μM) (gydF4y2Ba图4 egydF4y2Ba).gydF4y2Ba
PT2567在正常生理范围内调节血液计数gydF4y2Ba
全血分析显示白细胞计数无变化。然而,PT2567治疗使红细胞计数、血红蛋白水平和红细胞压积百分比在生理范围内降低到较低的参考值。然而,这些读数与车辆控制没有显著差异(gydF4y2Ba补充图S9a-dgydF4y2Ba).gydF4y2Ba
PT2567干预不影响颈动脉体大小gydF4y2Ba
最近的研究表明,HIF2α在慢性缺氧时颈动脉体的发育和生长中至关重要[gydF4y2Ba34gydF4y2Ba,gydF4y2Ba35gydF4y2Ba].鉴于颈动脉体在调节缺氧心肺反应方面的重要性,我们试图确定PT2567治疗是否影响颈动脉体形态及其延伸功能。暴露于su5416/缺氧3周的大鼠颈动脉分叉的组织学分析显示,与nx对照大鼠相比,颈动脉体容量显著增加(gydF4y2Ba补充图S10a,b,fgydF4y2Ba).与对照组相比,PT2567或西地那非治疗大鼠在再充氧期间的典型颈动脉体收缩未受影响(gydF4y2Ba补充图S10c-fgydF4y2Ba).gydF4y2Ba
PT2567干预可提高mct挑战大鼠的存活率gydF4y2Ba
此外,我们还研究了PT2567干预对ph的MCT大鼠模型的影响gydF4y2Ba补充图S11agydF4y2Ba, MCT挑战后14天,大鼠接受PT2567 (100 mg·kggydF4y2Ba−1gydF4y2Ba)或西地那非(30 mg·kggydF4y2Ba−1gydF4y2Ba)两个星期。在MCT挑战后14天以及干预研究结束时(28天)记录心脏回声。PT2567干预的存活率为90%,而西地那非治疗的存活率为77% (gydF4y2Ba补充图S11bgydF4y2Ba).心脏回波显示PT2567干预对PA-AT/PA-ET(比值)、RVOT-VTI和心排血量(gydF4y2Ba补充图S11c-egydF4y2Ba)与用交通工具处理过的动物相比。西地那非治疗的动物在所有心脏回波参数上与载体治疗的动物没有显著差异。gydF4y2Ba
抑制HIF2α可使PAH患者源性BOEC增殖和精氨酸酶活性正常化gydF4y2Ba
boec已被广泛地描述并用作研究模型gydF4y2Ba在体外gydF4y2Ba血管疾病中的内皮功能[gydF4y2Ba36gydF4y2Ba,gydF4y2Ba37gydF4y2Ba].本研究中使用的boec先前通过流式细胞术免疫染色CD133, CD34, VEGFR2 [gydF4y2Ba36gydF4y2Ba]和基因阵列分析,证明它们与PAECs功能和基因表达相似[gydF4y2Ba38gydF4y2Ba].此外,我们之前证明了这些细胞在HIFα表达或缺氧诱导的这些转录因子稳定性方面没有缺陷[gydF4y2Ba16gydF4y2Ba].gydF4y2Ba
首先,我们评估了PT2567 (1 μM)在上述PAH患者和健康志愿者分离的boec中的活性和特异性。选择人qPCR引物来补充su5416/缺氧模型中大鼠全肺基因表达的分析。gydF4y2Ba
来自健康志愿者和PAH患者的boec低氧暴露均增加了HIF1α和HIF2α靶基因(gydF4y2Ba图5一个gydF4y2Ba-g)与我们之前的研究一致。我们还报道了缺氧引起的增加gydF4y2BaGLUT1gydF4y2Ba,gydF4y2BaVEGFgydF4y2Ba而且gydF4y2Ba最长gydF4y2Ba与对照boec相比,PAH boec中表达量更高。PT2567治疗显著降低了缺氧诱导gydF4y2BaGLUT1gydF4y2Ba,gydF4y2BaPAI-1gydF4y2Ba,gydF4y2BaVEGFgydF4y2Ba而且gydF4y2Ba最长gydF4y2Ba而不影响HIF1α靶基因转录gydF4y2BaLDHAgydF4y2Ba,gydF4y2BaPGK1gydF4y2Ba而且gydF4y2Ba的基因gydF4y2Ba在对照和多环芳烃boec中(gydF4y2Ba图5一个gydF4y2Ba- g)。gydF4y2Ba
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
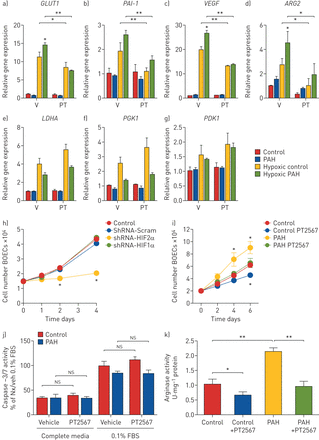
分析人血生长内皮细胞(BOECs)为研究模型gydF4y2Ba在体外gydF4y2Ba肺动脉高压(PAH)的内皮功能。a - g)人boec暴露于缺氧+/−PT2567 (PT)或载体(V),通过定量PCR检测基因表达:a)gydF4y2BaGLUT1gydF4y2Ba, b)gydF4y2BaPAI-1gydF4y2Bac)gydF4y2BaVEGFgydF4y2Bad)gydF4y2Ba最长gydF4y2Ba, e)gydF4y2BaLDHAgydF4y2Baf)gydF4y2BaPGK1gydF4y2Ba, g)gydF4y2Ba的基因gydF4y2Ba.h)低氧诱导因子(HIF α)抑制BOEC增殖。i)控制和PAH BOEC增殖+/−PT2567。j) caspase-3/7活性测定BOEC凋亡。boec分别在含0.1%胎牛血清(FBS) +/−PT2567的完全内皮培养基或基础内皮培养基中培养18 h。数据校正为阳性对照(0.1% FBS对照boec与车辆)百分数平均活动性±gydF4y2Ba扫描电镜gydF4y2Ba.k)来自对照和多环芳烃BOEC +/−PT2567的精氨酸酶活性测定。健康志愿者(n=3)或PAH患者(n=3)的人体boec;数据以均数±表示gydF4y2Ba扫描电镜gydF4y2Ba.*: p<0.05, **: p<0.001(单因素方差分析)。gydF4y2Ba
过度增生是PAH患者分离出的boec的一种有文献记载的表型[gydF4y2Ba36gydF4y2Ba].我们最初在对照组boec中使用短发夹敲除HIF1α或HIF2α的策略,初步评估这些转录因子对增殖的影响。与shRNA-Scramble相比,shRNA-HIF2α显著降低了对照组boec在第2-4天的细胞增殖,而shRNA-HIF1α对增殖几乎没有影响(gydF4y2Ba图5 hgydF4y2Ba).接下来我们研究了PT2567对BOEC增殖的影响。如前所述,与健康志愿者的boec相比,PAH患者的boec在第4天表现出相当高的增殖率,并维持到第6天。PT2567治疗可减少PAH患者和健康志愿者来源的boec的细胞增殖(gydF4y2Ba图5我gydF4y2Ba)而不影响由caspase-3/7活性测定的细胞凋亡速率(gydF4y2Ba图5 jgydF4y2Ba).gydF4y2Ba
我们之前已经在PAH患者分离出的boec中记录了精氨酸酶-2的异常表达和活性[gydF4y2Ba16gydF4y2Ba(人paec不表达arginase-1)。PT2567治疗24小时可抑制PAH患者和健康志愿者BOEC中的精氨酸酶-2酶活性(gydF4y2Ba图5 kgydF4y2Ba).综上所述,这些数据表明,在来自PAH患者的boec中靶向HIF2α,降低了异常的HIF2α靶基因表达,并降低了接近健康对照组的高增殖内皮表型和精氨酸酶-2活性。最后,我们评估了PT2567对BOECs网络形成功能的影响。与对照组相比,PAH患者的BOECs网络更脆弱,连接更短,环路更小[gydF4y2Ba36gydF4y2Ba].这种网络表型在PT2567处理后通过增加管长(gydF4y2Ba补充图S11agydF4y2Ba)和循环大小(gydF4y2Ba补充图S11bgydF4y2Ba)到类似于控制boec的值。载体或PT2567 (1 μM)处理健康对照BOEC后BOEC网络形成的代表性显微照片(gydF4y2Ba补充图S11cgydF4y2Ba)及PAH BOEC (gydF4y2Ba补充图S11dgydF4y2Ba)。gydF4y2Ba
讨论gydF4y2Ba
在这项研究中,我们已经证明了HIF2α信号是PH发展的关键因素,并且在起始阶段用一种新的全局HIF2α抑制剂(PT2567)专门针对这一过程(gydF4y2Ba图1gydF4y2Ba)或作为干预策略(gydF4y2Ba图2gydF4y2Ba)减轻了许多与酸碱度疾病相关的病理。gydF4y2Ba
在整个研究过程中使用的HIF2α抑制剂PT2567直接结合到HIF2α的PAS-B结构域内的一个区域,并破坏HIF2α/ARNT二聚体的形成,这是HIF2α激活所必需的关键事件。我们的gydF4y2Ba在体外gydF4y2Ba研究证实了PT2567对HIF2α的高亲和力和特异性,以及抑制HIF2α靶基因所需的低微摩尔浓度。此外,我们报道了HIF2α信号通路可能与PAH患者的boec中观察到的高增殖和内皮网络表型有关gydF4y2Ba通过gydF4y2BaHIF2α靶基因的表达(gydF4y2BaGLUT1gydF4y2Ba,gydF4y2BaVEGFgydF4y2Ba,gydF4y2Ba黄牌gydF4y2Ba而且gydF4y2Ba最长gydF4y2Ba).gydF4y2Ba
我们的gydF4y2Ba在活的有机体内gydF4y2Ba药动学分析发现成年啮齿动物具有良好的口服生物利用度和血浆稳定性(gydF4y2Ba补充数字S1-S4gydF4y2Ba),其浓度随后用于预防和干预方案。su5416/缺氧预防方案证实了先前在小鼠HIF2α PH操纵模型中描述的几个关键观察结果,但也进一步证实了抑制整体HIF2α活性直接影响PH疾病的引发。在su5416/缺氧模型中,PT2567干预降低了肺血管阻力和血管重构,增加了心功能,并在MCT PH模型中提供了显著的生存优势。在这些研究中,西地那非被用作临床比较,但在预防和干预研究中都只提供了很小的有益效果。我们认为这种差异可能是由于血流动力学分析的时间,在最终剂量后16-18小时完成,因此可能在血管活性窗口之外。gydF4y2Ba
尽管在特发性PAH患者和啮齿动物PH模型的肺血管闭塞性病变中都发现了HIF2α蛋白稳定性的增加,但HIF2α在PH发生和发展中的机制仍不清楚。绝大多数已出版gydF4y2Ba在活的有机体内gydF4y2Ba对HIFα在PH起始和发展中的作用的研究使用了组织特异性基因缺失小鼠模型。尽管这些组织特异性小鼠模型对于确定基因在疾病发展中的贡献是非常有用的,但它们没有考虑到可能影响正常生理或病理反应的潜在发育缺陷,以及特定基因功能如何在多个血管床上不同而影响疾病进展或严重程度。gydF4y2Ba
本研究的主要目的是确定整体抑制HIF2α在预防和恢复肺动脉高压中的影响,并研究该转录因子在调节潜在PH相关靶基因表达时的病理影响,这些靶基因系统地影响血管稳态。gydF4y2Ba
为此,我们研究了啮齿类动物PH模型全肺样本的基因表达谱,以确保包括PH疾病进展所需的所有肺细胞类型。这些数据证实了PT2567的特异性,并显示炎症(gydF4y2Bacxcl12gydF4y2Ba,gydF4y2Ba趋化因子受体cxcr4gydF4y2Ba,gydF4y2Baicam1gydF4y2Ba,gydF4y2Ba希利gydF4y2Ba)及信号传送目标(gydF4y2BaaplngydF4y2Ba,gydF4y2Ba最长gydF4y2Ba,gydF4y2Baccnd1gydF4y2Ba,gydF4y2Bapai1gydF4y2Ba,gydF4y2Baedn1gydF4y2Ba)与PH有关,并附加恢复gydF4y2Baid1gydF4y2Ba基因的表达。虽然西地那非治疗对这些动物血流动力学参数的影响似乎不太明显,但西地那非对全肺基因表达模式的影响与PT2567并无不同。PT2567和西地那非联合或交替治疗的进一步研究可能具有临床相关性。gydF4y2Ba
D最近的研究gydF4y2Ba人工智能gydF4y2Baet al。gydF4y2Ba[gydF4y2Ba39gydF4y2Ba[答案HgydF4y2BaugydF4y2Baet al。gydF4y2Ba[gydF4y2Ba40gydF4y2Ba]为HIF2α在PH中的重要作用提供了进一步的支持,并证实了许多这些炎症和信号靶基因。此外,HgydF4y2BaugydF4y2Baet al。gydF4y2Ba[gydF4y2Ba40gydF4y2Ba]将PT2567用于低气压低氧大鼠PH模型,并报告了肺动脉压和血管重塑的降低。与此一致,我们的研究证实了肺血管平滑肌沉积的减少,并显示了肺血管内皮细胞增殖和mpo阳性细胞(骨髓系)在血管周围区域的减少。gydF4y2Ba
这些发现与先前发表的小鼠组织型特异性HIF2α缺失模型一致,报告内皮血管活性功能降低(gydF4y2Baedn1gydF4y2Ba,gydF4y2Baarg-2gydF4y2Ba,gydF4y2BaaplngydF4y2Ba) [gydF4y2Ba8gydF4y2Ba,gydF4y2Ba9gydF4y2Ba,gydF4y2Ba16gydF4y2Ba],细胞增殖/血管生成(gydF4y2Baccnd1gydF4y2Ba,gydF4y2BavegfgydF4y2Ba) [gydF4y2Ba16gydF4y2Ba]和内皮细胞向间质细胞的转变(gydF4y2Basnai1/2gydF4y2Ba)目标[gydF4y2Ba41gydF4y2Ba].gydF4y2Ba
我们之前的研究表明,在慢性缺氧PH模型中,肺内皮细胞HIF2α的缺失会降低循环的促炎细胞因子和生长因子(IL-6, IL-8, TNF-α, IL-1β, Cxcl12) [gydF4y2Ba16gydF4y2Ba].这些因子的高循环浓度与PAH患者预后不良有关[gydF4y2Ba42gydF4y2Ba].我们报道PT2567干预降低循环TNF-α和增加免疫抑制IL-10血浆浓度。先前的一份报告描述了重组IL-10给mct感染大鼠减少了血流动力学和肺血管重塑,并提供了比车辆对照的生存优势[gydF4y2Ba43gydF4y2Ba].其他小鼠炎症模型将异常的髓系HIF2α表达与低血浆IL-10浓度联系在一起,其中骨髓系HIF2α缺乏的小鼠恢复/增加了血浆IL-10 [gydF4y2Ba44gydF4y2Ba].需要进一步的研究来阐明细胞来源(调节性t细胞、巨噬细胞、b细胞、树突状细胞)和HIFα调节IL-10在PAH中的作用。gydF4y2Ba
一氧化氮(NO)的循环浓度是心血管系统结构完整性和功能的关键。NO生物利用度降低与内皮功能障碍、平滑肌增生有关,并与心血管和多环芳烃疾病严重程度的发展有关[gydF4y2Ba45gydF4y2Ba- - - - - -gydF4y2Ba48gydF4y2Ba].我们发现,在PT2567或西地那非干预后,血浆NO/亚硝酸盐浓度恢复到接近正常/常氧值。这些数据与我们之前在小鼠慢性缺氧PH模型中的研究一致[gydF4y2Ba16gydF4y2Ba],我们描述了hf2 α - arg2轴对ph的发展至关重要。我们发现精氨酸酶表达和活性的增加导致血管NO稳态失调[gydF4y2Ba16gydF4y2Ba].西地那非间接调节内皮功能gydF4y2Ba通过gydF4y2Ba内皮型一氧化氮合成[gydF4y2Ba49gydF4y2Ba然而,西地那非对血浆亚硝酸盐的作用被认为与HIF信号无关gydF4y2Ba
鉴于PT2567的整体HIF2α抑制作用,其他非肺组织也可能间接调节PH疾病的进展。我们报道PT2567干预降低了几个与结构和应激诱导适应相关的RV基因的表达;然而,基于对心血管疾病中HIF2α信号的知之甚少,我们不能完全区分心功能的恢复是PT2567的直接作用还是肺血管功能的间接结果。gydF4y2Ba
HIF1α在PH起始和发展中的作用仍然存在争议。缺乏特异性HIF1α抑制剂阻碍了大鼠su5416/缺氧或MCT模型的潜在干预研究。因此,绝大多数研究使用了组织特异性敲除小鼠模型,这些模型显示出不同的结果,这取决于目标组织和异常激活(PHD或Von Hippel-Lindau)或抑制HIF1α稳定性的方法。在某种程度上,靶向平滑肌细胞中的HIF1α信号已显示出减缓疾病进展的有益作用[gydF4y2Ba50gydF4y2Ba].我们之前研究了肺内皮细胞HIF1α在PH慢性缺氧小鼠模型中的作用。与野生型动物相比,我们没有观察到在7 - 14 - 21天缺氧时间过程中PH疾病的发展有差异。此外,HgydF4y2BaugydF4y2Baet al。gydF4y2Ba[gydF4y2Ba40gydF4y2Ba]最近报道,在PH的慢性缺氧模型中,HIF1α的全面抑制并不支持该转录因子在PH发育中的核心作用。需要进一步的研究来阐明HIF1α或HIF1α/HIF2α的组合是否影响PH的发展。gydF4y2Ba
目前的共识观点支持HIF2α在PAH发展中的作用,最有可能是通过影响肺血管床的多条信号通路,导致血管特性/可塑性/内稳态的逐渐丧失,从而驱动重塑和增加血管阻力。本文中PT2567干预提供的保护程度可能在一定程度上归因于多个PH相关靶基因的调节gydF4y2Baglut1gydF4y2Ba,gydF4y2Bacxcl12gydF4y2Ba,gydF4y2BaaplngydF4y2Ba,gydF4y2Baedn1gydF4y2Ba,gydF4y2Baicam1gydF4y2Ba,gydF4y2Ba希利gydF4y2Ba而且gydF4y2Baccnd1gydF4y2Ba虽然其他已知的多环芳烃目标的贡献包括gydF4y2Babmpr2gydF4y2Ba[gydF4y2Ba26gydF4y2Ba),gydF4y2Bail - 6gydF4y2Ba[gydF4y2Ba51gydF4y2Ba),gydF4y2Basox17gydF4y2Ba[gydF4y2Ba52gydF4y2Ba),gydF4y2Bacav1gydF4y2Ba[gydF4y2Ba53gydF4y2Ba]跨越多个组织和其他器官是最有可能的,不能排除。gydF4y2Ba
近二十年来,异常HIF2α稳定性和功能在PH发展中所起的作用一直是一个备受关注的领域[gydF4y2Ba8gydF4y2Ba,gydF4y2Ba9gydF4y2Ba,gydF4y2Ba12gydF4y2Ba,gydF4y2Ba16gydF4y2Ba,gydF4y2Ba54gydF4y2Ba,gydF4y2Ba55gydF4y2Ba].这些集体研究清楚地阐明了血管HIF2α在PH疾病发生和发展中的关键作用,并支持其作为PH患者治疗的新靶点的评估。然而,不同于统一的遗传背景gydF4y2Ba在活的有机体内gydF4y2Ba多环芳烃是一种复杂的动态疾病,其特点是患者存在相当大的异质性。因此,需要进一步的研究来确定PAH人群对HIF2α抑制剂的总体敏感性,并阐明PT2567与当前治疗策略的联合作用。我们相信,目前的器官芯片技术与来自PAH患者的boec相结合,可以为进一步表征HIF2α在PH中的作用提供一个极好的平台。gydF4y2Ba
总之,在su5416/缺氧的PH啮齿动物模型中,我们的工作提供了对PH启动和发展中HIF2α转录活性的直接抑制的更好理解。需要进一步的研究来确定HIF2α在PH启动所需的因素层次中的位置,以及HIF2α长期抑制不仅影响PH疾病病理,而且影响其他基本生理过程。gydF4y2Ba
补充材料gydF4y2Ba
补充材料gydF4y2Ba
请注意:gydF4y2Ba补充材料不是编辑部编辑的,上传时是作者提供的。gydF4y2Ba
补充数据gydF4y2Baerj - 02061 - 2019 - _supplementary_figuresgydF4y2Ba
补充表gydF4y2Baerj - 02061 - 2019 - _supplementary_tablesgydF4y2Ba
可共享的PDFgydF4y2Ba
脚注gydF4y2Ba
这篇文章有补充资料可从gydF4y2Bawww.qdcxjkg.comgydF4y2Ba
作者贡献:D. Macias, S. Moore, X. Du, H. Tan, E.M. Wallace和A.S. Cowburn设计研究;D. Macias, S. Moore, A. Crosby, M. Southwood, S. Xie, A. Vassallo, A.J.T. Wood, X. Du, H. Tan和A.S. Cowburn进行了研究;D. Macias、S. Moore和A.S. Cowburn分析了数据;马西亚斯、E.M.W和A.S.考伯恩撰写了这篇文章。gydF4y2Ba
利益冲突:D.马西亚斯没有什么可透露的。gydF4y2Ba
利益冲突:S. Moore没有什么可透露的。gydF4y2Ba
利益冲突:A.克罗斯比没有什么可透露的。gydF4y2Ba
利益冲突:M. Southwood没有什么可透露的。gydF4y2Ba
利益冲突:X. Du报告了一种用于治疗肺动脉高压的专利组合物。gydF4y2Ba
利益冲突:H. Tan报告了一种用于治疗肺动脉高压的专利组合物。gydF4y2Ba
利益冲突:S. Xie报告了一种用于治疗肺动脉高压的专利组合物。gydF4y2Ba
利益冲突:A.瓦萨罗没有什么可透露的。gydF4y2Ba
利益冲突:A.J.T.伍德没有什么可透露的。gydF4y2Ba
利益冲突:E.M. Wallace报道了一种用于治疗肺动脉高压的专利组合物。gydF4y2Ba
利益冲突:A.S. Cowburn报告了Peloton Therapeutics公司在研究期间的资助。gydF4y2Ba
支持声明:本研究获得了国家健康研究所剑桥生物医学研究中心、Peloton Therapeutics公司和皇家帕普沃斯医院NHS信托基金的资助。本文的资助信息已存入gydF4y2Ba交叉参考基金注册gydF4y2Ba.gydF4y2Ba
- 收到了gydF4y2Ba2019年10月22日。gydF4y2Ba
- 接受gydF4y2Ba2020年8月26日。gydF4y2Ba
- 版权所有©ERS 2021gydF4y2Ba
本版本根据创作共用授权协议4.0发布。gydF4y2Ba
参考文献gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba










