





文摘
慢性呼吸道疾病是非常普遍的在全球范围内和在可预见的未来还将继续上升。尽管花费了大量的努力近几十年来,新颖和有效的治疗方法的发展缓慢。然而,有越来越多的证据表明,社区的新微生物在体内,人类微生物组,最重要的是参与开发和慢性呼吸道疾病的进展。了解详细的机制之间的相声主机和微生物群的发展是至关重要的微生物——或者host-targeted疗法和预防策略。在这里,我们审查和讨论最近的知识不断倒数宿主和微生物之间的相互作用在健康和呼吸道疾病。此外,我们强调microbiome-based疗法研究的前景和讨论需要采用更全面的方法恢复肺的利基和微生物群落。
文摘
肺的互惠的微生物和宿主之间的相互作用越来越被视为一个重要的健康决定因素。这个相声的复杂性时需要考虑研究疾病和发展未来的新疗法。https://bit.ly/2VKYUfT
介绍
在过去的十年中,我们已经了解到肺部,以前认为是无菌的,事实上港口一个动态的生态系统不同的细菌,真菌和病毒(1]。这种肺部微生物检测健康(2,3),改变在疾病4),预测疾病的结果(5,6和与宿主免疫的变化7,8]。最近的见解,基于对人类的研究(8和老鼠7),是基线肺部免疫的语气,即使在健康,当地微生物环境密切相关(9]。这个假设的“免疫基调”在气道和肺泡至少部分由微生物群是一个激进的背离我们的传统,二分的理解肺免疫:休眠在卫生、激活感染。旁边的肺部炎症环境的变化,大多数肺部疾病,如哮喘(10),慢性阻塞性肺病(11),囊性纤维化12)、特发性肺纤维化(IPF) [13),最近,肺癌(14)在肺与微生物生态失调有关。然而,它是未知的,如果细菌组成的变化驱动疾病发病机理或如果他们而改变的反映的生态位9,15]。因此,至关重要的是要理解底层的分子机制在host-microbiome接口开发了新型靶向治疗或预防方法。
在这里,我们讨论的影响host-microbiome相声对呼吸道健康和疾病,而专注于当地微生物群和主机如何交互的上皮表面。此外,我们审查当前治疗方法,并且提出一个更全面的方法在未来治疗肺部疾病。本文后续的报告和讨论后,人研究研讨会”相声肺微环境:影响理解慢性肺病”(柏林,2018年2月)。
肺粘膜利基
人体的所有外部接口,如肠道、皮肤、生殖和呼吸道,是由不同的微生物菌群(殖民16]。不同的微生物群落不同身体网站由于局部因素(如。氧气、二氧化碳、pH值、营养、主机防御因素,吸入污染物),形状的利基。低的腔为例,胃肠道代表一个低氧,营养丰富的环境,因此通常由厌氧生物丰富的社区;相比之下,皮肤是一个低营养环境直接暴露于氧气,因而通常密集的环境相对稀疏的社区oxygen-tolerant细菌。因此,当地环境是一个关键因素在发展早期的微生物群落的形成,而且在后期17]。
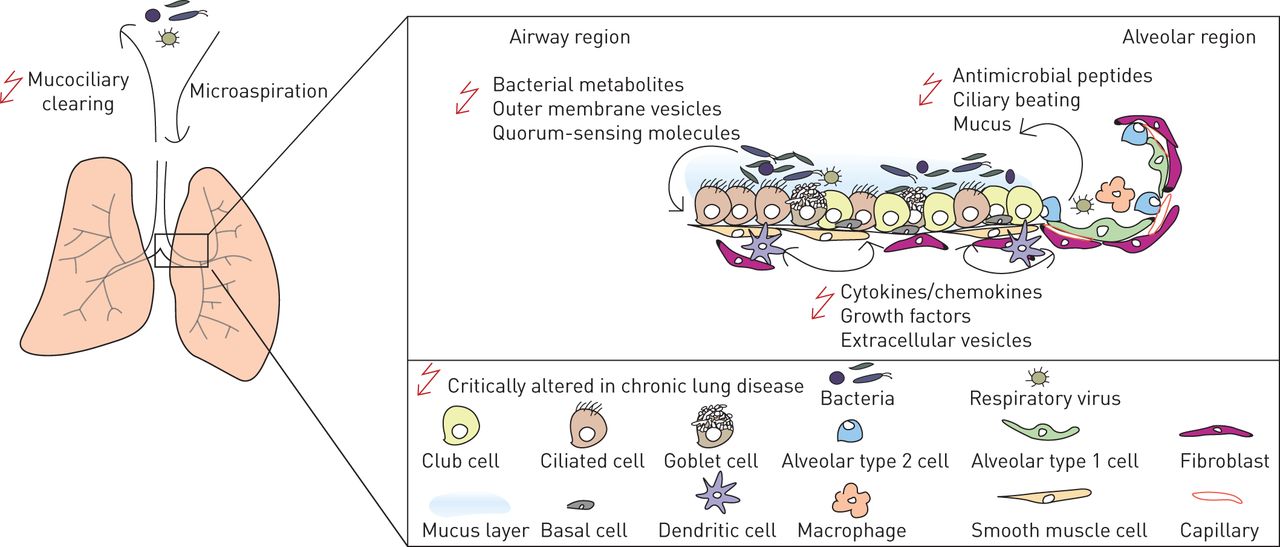
因此,肺微环境创建了一个特殊的生态位微生物不同于身体的其他网站,可能会影响niche-specific殖民化。因此,气道上皮细胞是一个原则的贡献者在塑造这个利基,因为它们是战略定位是第一个接触吸入物质(包括微生物)和宿主组织。各种机制是受雇于气道上皮细胞在宿主防御感染,包括其屏障功能、黏膜纤毛的清除,生产抗菌肽和其他物质,赞成和抗炎介质和传输能力(如。聚合物IgA, IgM从基底上皮细胞通过聚合的顶端免疫球蛋白受体(pIgR)) (18- - - - - -21]。类似的机制很可能导致呼吸道niche-specific社区的形成,和这样的当地条件允许有益微生物群上皮表面存在的关键。这样的参与host-microbiota交互在囊性纤维化病好了。在囊性纤维化,突变功能障碍的囊性纤维化跨膜调节器(雌性生殖道蛋白质导致减少阴离子交换(主要是碳酸氢和氯)在上皮表面,导致表面脱水和粘稠的粘液。从航空公司这种粘液不能容易清除,这有助于解释为什么雌性生殖道突变与变更相关的微生物群,包括频繁的殖民化金黄色葡萄球菌和铜绿假单胞菌(最近了22])。然而,我们仍然缺乏知识的潜在机制利基变化在更复杂的基因肺部疾病,如哮喘和慢性阻塞性肺病。
此外,上皮细胞微生物信号传输环境和免疫系统,让它适当的反应或开发公差(山21,23,24]。
各种微生物存在传感机制包括模式识别受体,如toll样受体(20.),和其他进程不是介导通过经典受体介导的信号,如。集成的压力反应(25]。在传感微生物、环境或内源性挑战,上皮细胞山积极响应通过增加国防分子,如抗菌肽,细胞因子和趋化因子。这些增强防御呼吸道病原体,同时改变复杂微生物群落的生态位,表明生态位的属性遇到的微生物由于环境因素的变化。解剖上的差异以及差异等呼吸道上皮细胞组成形状这一生态位。尽管形状的上皮细胞导致当地条件微生物群,上皮细胞暴露于微生物及其产品标记对其功能的影响。因此,人类上皮细胞和微生物群发展互动互利,应对病原体和容忍无害的物质26]。这个概念很重要,了解吸入环境诱因调节免疫力,因为微生物组成为应对环境因素,如农场(27)和地球成因学的(earth-derived)尘埃28]。然而,上皮细胞如何集成这些微生物和nonmicrobial信号调整反应是不完全理解。
为了从环境中传递信号,上皮细胞与其他肺细胞类型使用复杂的通信系统。气道上皮细胞(皮肤和肠道和)扮演关键角色在发射信号从微生物和环境刺激直接指示抗原递呈树突状细胞免疫对炎症或公差24]。上皮细胞相互作用不仅与其他免疫细胞,如巨噬细胞、中性粒细胞,先天淋巴细胞和t细胞,而且还与结构细胞如成纤维细胞、气道平滑肌细胞和内皮细胞通过大量不同的机制(图1)[18,20.,24]。这个数组的交互环境诱因、微生物和肺细胞上皮细胞可以编排宿主防御和免疫力,维护上皮完整性和调解病理气道重构(图1)[21]。
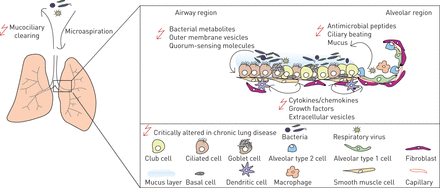
Host-microbiome相声的肺微环境。肺微环境由不同的细胞类型,这取决于proximal-to-distal气道树中的位置。上皮细胞层在较大的航空公司不断暴露于各种不同的微生物的微生物群。而后者影响主机的组成等因素消除通过黏膜纤毛的清除,它还依赖于微生物居民之间的竞争。显然,现在是有一个宿主和微生物环境之间复杂的相互作用。因此,细菌代谢物或外膜囊泡可以影响主人的地位,在抗菌肽或细胞因子可以塑造微生物群的构成。几乎所有的这些单因素改变的慢性呼吸道疾病,至关重要的是欣赏和评估该系统的复杂性在将来的研究中。
建立和维护肺的微生物群
肺部微生物群的建立可能发生在生命的最初几天或几周内。虽然早期高调报道“胎盘微生物组”的存在可能会影响prepartum肺发展(29日),后续控制研究未能发现细菌信号不同于污染DNA存在于消极的控制(30.]。新生鼠肺微生物群的检测极限(以下通过定量PCR),增加总负担在接下来的几天或几周内31日,32]。在人类婴儿,呼吸道微生物的组成似乎成熟可预测,特征明显的模式在生命的第一年(32]。除了肺的特殊生态特征,早期呼吸道微生物群影响交货方式(阴道与剖腹产)的方法喂养(母乳与瓶),曝光(兄弟姐妹、日托出席)32]。挑逗,早期呼吸系统有可能预测后续对呼吸道感染的易感性(32,33],显示角色的当地微生物免疫内稳态和抵抗病原体。相比之下,儿童时期接触微生物多样性增加促进平衡发展的免疫力,预防炎症反应过敏和哮喘发展(34,35]。这种保护在一定程度上是与不同的农场尘埃,在体外增加上皮屏障功能和抗病毒防御(27]。
连接建立后,肺部微生物的组成是由三个生态因子:移民、消除和社区成员的相对增长率(图1)[36]。在卫生、肺社区稀疏和动态,很大程度上取决于移民之间的平衡(通过microaspiration [3])和消除(通过咳嗽、黏膜纤毛的清除和免疫防御系统)。很少有证据支持的存在居民肺部健康的细菌繁殖和生存选择压力(1,4,9]。然而,瞬态和动态社区中发现健康在很大程度上是可行的37],发挥免疫检测影响宪法的下呼吸道7,8]。
建立一个多样化的呼吸道微生物菌丛是受很多因素的影响。之间有一个调整平衡共生的公差或“有益的”细菌上皮表面,和活跃的发展对病原体的免疫反应。值得注意的是,这种平衡是由主机和microbe-derived信号。突然之间的转变,这严格控制平衡,如特定病原体或结果,例如,病毒诱导气道上皮损伤破坏的障碍,诱发局部免疫反应,可以有不利影响宿主和微生物,因此可能会导致呼吸道疾病的发病机制。
环境在塑造肺部微生物的作用
环境影响肺部疾病的发展中扮演关键的角色。这可能是由于直接作用于宿主上皮屏障和免疫反应,但大多数已知的危险因素,如吸烟、空气污染(38,39),病毒感染和饮食也直接影响微生物群(11,40- - - - - -42]。因此,两者的结合可能是疾病发展的关键。
例如,研究最多的一个吸入毒素,香烟烟雾,据说有不利影响宿主和微生物群落。长期吸烟暴露会导致基线增加呼吸道的炎症反应和上皮改造对杯状细胞增生和纤毛和纤毛活动的减少19),可以减少提供的抗菌防御呼吸道上皮细胞(43]。在COPD患者,它已被证明是与减少肺部细菌和增加有关流感嗜血杆菌和链球菌引起的肺炎(44),但在“健康”吸烟者迄今最大的研究没有发现明显吸烟对肺部的影响微生物(45]。因此,它是有趣的推测香烟主要影响主机系统,随着时间的推移和疾病(COPD)易感个体的发展也会影响微生物。
环境影响呼吸道疾病的另一个例子是饮食,对微生物群和健康有着深刻的影响,甚至在短期内(46]。膳食纤维的摄入诱发类似肺部和肠道有益微生物群变化(47]。虽然低纤维的饮食改变肠道微生物群落在多个代小鼠(48),高纤维的饮食有好处在怀孕的老鼠和抑制过敏性气道疾病(AAD)在母亲和他们的后代49]。这也突出了相声肠道和肺之间的重要性。高纤维饮食诱导生产短链脂肪酸的肠道细菌,运输系统的肺,发挥抗炎行动和改善AAD小鼠(47]。相比之下,lipid-rich饮食改变了微生物群,促进代谢炎症(50),一直伴随着premetastatic细分发展在肺癌51]。然而,行动的详细机制仍不清楚。
随着细菌,常见的主要呼吸道病毒,包括呼吸道合胞体病毒、鼻病毒、冠状病毒、流感病毒和腺病毒的呼吸道微生物(52),导致慢性呼吸道疾病的发病机制。这可能源于复杂的病毒与宿主免疫系统的相互作用和微生物群包括其他病原体。这些交互能够影响细菌病原体的流行增加呼吸道上皮细胞粘附分子的表达,损害呼吸道上皮细胞损害屏障功能,损害黏膜纤毛的清除和改变宿主免疫力和肺微环境53- - - - - -55]。有趣的是,有人建议,病毒(噬菌体)诱导一致的和可再生的呼吸系统慢性疾病的微生物的变化,但不健康的状态,并已被证明增加微生物多样性在鼻咽53,56,57]。此外,真菌等曲霉属真菌种虫害导致慢性呼吸道疾病的发病机理的病原体,而且通过激活免疫系统,并可能通过与微生物群的互动,尤其是复合菌群与非结核分枝杆菌(9,58]。
重要的是,没有微生物的某些成员可以有不利影响肺部健康。所示的相对寄生蠕虫的消失,是一个常数我们肠道微生物组的一部分,甚至在化石被发现从喋喋不休的鱼59]。事实上,我们的“现代”免疫系统在连续开发存在寄生虫(60),这也许可以解释他们在免疫重要的监管作用。通常,蠕虫诱导2型免疫反应被认为是在宿主防御这些寄生虫感染。然而,蠕虫感染诱发监管反应,目的是抑制免疫反应针对虫抗原,它允许虫子生活在宿主多年。此外,这种监管反应有一个旁观者效应通过促进过敏原公差(61年]。这是说明了在小鼠模型的研究,各种寄生虫的感染已被证明对过敏性哮喘的发展[提供保护62年]。的范围helminth-induced保护机制包括抑制白介素(IL) -33 (63年在航空公司,调控t细胞的诱导64年,65年),b细胞(66年,67年和巨噬细胞68年]。有趣的是,寄生虫的存在也会影响细菌的组成社区,这可能是重要的保护小鼠过敏性呼吸道疾病(69年]。
利基和微生物改变呼吸道疾病
在疾病、下呼吸道的生态变化显著。气道和肺泡,通常适合细菌繁殖[3),从根本上改变了营养丰富的水肿和粘液的涌入,建立鲜明的氧梯度,促进细菌生长的炎症反应(70年,71年)和本地主机防御障碍(36,72年]。因此,这并不奇怪的横断面研究发现肺改变微生物在肺部疾病,生态区位的改变可能导致不同的微生物群落由于选择压力。肺的构成微生物的相对增长率越来越取决于其成分。
虽然有一些研究在IPF(微生物的作用13)和肺癌(14),大部分知识是基于研究慢性炎症的呼吸道疾病,如哮喘、慢性阻塞性肺病和囊性纤维化。一般来说,患者更易感染和急性加重,再次影响微生物4,73年,74年]。流感嗜血杆菌,莫拉克斯氏菌属复活和肺炎链球菌与哮喘和慢性阻塞性肺病(严重)的发展。他们修改肺部微生物和驱动炎症,氧化应激,症状和急性加重(44,75年- - - - - -80年),这可能会形成一个恶性循环的疾病。复活的和流感嗜血杆菌特别是诱导中性粒细胞炎症,更严重的疾病,和类固醇电阻(79年- - - - - -84年]。此外,在哮喘、呼吸改变微生物概要文件与哮喘相关的表型(85年[]和严重程度86年),反应的过敏原(87年)和治疗(88年]。
囊肿性纤维化和支气管扩张,嗜血杆菌和假单胞菌种虫害日益主导着肺部微生物群(89年,90年]。基因关联研究Phe508del囊性fibrosis-affected纯合个体显示关联的单核苷酸多态性与重要疾病特性相关的细菌殖民化。例如,基因变异的人类白细胞抗原类II基因(暗示作用的t细胞和b细胞免疫)的发病年龄与持久的铜绿假单胞菌感染,表明宿主的免疫系统的作用,在选择一个不同的微生物群(91年]。
像肺部微生物组的细菌组成,抗病毒反应在慢性呼吸疾病改变。在哮喘气道上皮修复,以应对病毒感染异常是由于基因的变化调节上皮屏障功能和修复(92年),气道上皮细胞减少了先天免疫常见的病毒如鼻病毒(79年,93年]。这是由于我和III型干扰素,增加microrna的水平降低(94年,95年]。转化生长因子(TGF) -β哮喘一般细胞因子调节,抑制气道上皮先天免疫反应通过抑制细胞因子的信号(soc) 1和SOCS-3,导致受损的抗病毒免疫(96年]。这种抗病毒免疫受损也可能改变细菌微生物和由细菌导致交叉感染97年]。鼻病毒感染也可以损害肺泡巨噬细胞的吞噬作用的细菌可以导致慢性阻塞性肺病(细菌产物98年]。小鼠的研究表明,influenza-infected上皮细胞祖细胞严重受损的远端肺展出更新能力由于病毒诱导的封锁β-catenin-dependent Fgfr2b信号(99年]。这可能有助于修复受损的远端肺在慢性阻塞性肺病,尽管其参与人类疾病尚不清楚。因此,病毒和细菌感染,无法解决这些问题,可能加剧气道和肺泡修复受损。结合加剧气道和肺泡免疫语气,异常修复机制病原体暴露后脆弱的个体可能代表host-microbiome互动如何导致疾病进展的机制。目前还不清楚如果改变肺部微生物也可以先于肺结构变化和炎症,从而导致疾病的发生。
正如前面所讨论的,肺失调和肺部炎症之间的关系是双向的:无序肺社区引发上皮和腔的炎症,肺微环境的进一步改变生长条件、慢性炎症和保持生态失调(4,36]。炎性肺条件共同发现的结果是炎症反应变形菌门门(4),在小鼠肺部微生物已被证明与肺的天生的细胞因子水平有关IL-1α[7]。此外,改变肠道微生物的可能影响免疫肺的语气,更严重的发展所示实验哮喘在无菌鼠(31日,47]或小鼠干扰微生物由于对早年生活的抗生素治疗(One hundred.,101年]。相比之下,没有微生物群在无菌鼠或由于抗生素治疗最近被证明保护Kras突变的老鼠和p53损失从肺癌发展102年]。研究者称,这种效果是由于IL-1β的感应和IL-23通过微生物群,导致IL-17生产γδ-T细胞和肿瘤发生。
在人类中,慢性阻塞性肺病患者的两到三倍克罗恩氏结肠炎,和50%的患有炎症性肠病也有肺部炎症(103年- - - - - -106年]。胃肠道微生物含量最高的,因此一个交互的肠道微生物群与胃粘膜及其代谢产物影响系统性免疫,这可能反过来影响肺(107年]。因此,改变肠道微生物群和生理学和可能导致呼吸道疾病反之亦然,可能通过gut-lung轴。然而,重要的是要注意无菌繁殖,而且在某种程度上抗生素治疗,会影响这两个的肺部和肠道微生物,所以还不完全清楚这些效果完全可以解释为gut-lung相声或如果他们受到异常的当地microbiota-immune相声。沿着这条线,在小鼠肺免疫的语气已经建议相关肺部细菌比肠道细菌(7]。
相比之下,在老鼠身上,colitis-induced肺部病理学与增加肺部炎症和革兰氏阴性细菌木糖醇,这可能是从肠道(104年]。此外,香烟烟雾诱发COPD模型实验(108年- - - - - -110年)表明,减少肺气体交换和缺氧与肠道缺氧有关,导致结肠改造,细胞死亡,炎症和屏障功能受损111年]。此外,nod样受体的激活肠道细菌生产活性氧增加肺泡巨噬细胞,这表明胃肠道微生物导致肺部氧化应激(112年]。相反,口服应用有益的益生菌(双歧杆菌,和乳酸菌在香烟的)减少气道炎症和肺气肿smoke-exposed老鼠(113年]。
大加拿大的一项研究报道,四个菌属毛螺菌属,韦永氏球菌属,Faecalibacterium和罗思氏菌属减少人类婴儿的粪便,随后患上哮喘,并接种人类粪便微生物补充这四个类群为无菌鼠部分保护他们的F1 AAD实验诱导子代(114年]。因此,有鼓励在小鼠模型的研究强调因果gut-lung轴的呼吸道疾病。然而,人类发现第一个需要验证之前提出这个相声作为治疗呼吸道疾病的特征。
总之,host-microbiome接口,无序社区可能是引起和宿主炎症的影响。然而,考虑到紧密互惠的微生物和宿主之间的相互作用,这可能是不可能确定的真正原因第一个疾病发展的变化,作为东道主和微生物因素一致,紧密地交织在一起。
对新的治疗方法的发展
由于强烈的和动态宿主和微生物之间的相互依赖在当地的利基市场,那么毫无疑问,大多数药物用于临床实践,设计目标主机也会影响微生物。因此,吸入糖皮质激素和质子泵抑制剂影响肺部微生物群(115年- - - - - -117年),以及随后的肺炎风险(118年,119年]。大环内酯物抗生素,广泛有效的慢性肺部疾病,如慢性阻塞性肺病(120年),影响肺部微生物群和宿主免疫力。也许最挑逗,肺部微生物群的基线差异似乎预测病人反应吸入糖皮质激素等治疗(10),这表明肺部细菌的差异可能代表一个尚未开发的“精密医学”的表现型的肺。这将打开新的可能性利用这个重要的相声在治疗干预,但为了做到这一点,我们首先需要提高我们对分子机制的理解。
增加微生物的重要性的证据提出了恢复的概念“病”预防或治疗疾病的微生物使用microbiota-directed疗法或host-targeted疗法,益生菌等代谢物,肺部微生物群移植或维生素D治疗,我们更详细地讨论。
许多临床研究研究了益生菌的功效,据说有利于宿主,防止慢性疾病如哮喘或过敏性鼻炎。然而,这些研究在很大程度上产生了矛盾的结果(121年),这可能是由于这样的事实,使用益生菌没有选择基于潜在的机械效应和可能不是很理想。微生物是一个复杂的生态系统组成的各种不同的居民,受到许多外部(host-derived)因素的影响;因此,添加单一菌株可能不会产生深远的影响。因此,整个微生物的转移通过粪便微生物群移植(FMT)可能是一个更有前途的方法。引入健康的微生物群为患病的主机恢复免疫和生理122年),说明肠道菌群与他们的产品可以调节宿主免疫局部和全身,FMT可以取代疾病与健康的微生物(40,107年]。FMT是成功治疗抗药性显著(90%)艰难梭状芽胞杆菌全身的结肠炎(123年),现在被用作治疗在选定的病人124年,125年]。新的微生物灌输快速和持续至少一个月,表明潜在的困难诱导长期有益微生物的变化通过只有目标微生物。虽然有令人鼓舞的数据,问题仍FMT是否也会影响肺部健康,以及肺部微生物群是否移植是可行的。此外,最近,严重的并发症FMT已报告由于转移耐药细菌(126年]。因此,至关重要的是确定是否这样的方法,到目前为止仅是暂时性的改变微生物,可用于需要长期治疗慢性疾病(肺),配合各种结构性变化,异常黏膜纤毛的间隙,和更多40]。
一起活的细菌,特定的微生物分子如脂多糖(LPS)和肽聚糖可以诱发或调节炎症反应(127年- - - - - -129年]。此外,文化上层的益生菌显示抗炎作用,已被归因于存在分泌免疫调节代谢产物(130年]。例如,某些益生菌文化上层清液双歧杆菌属物种减少2型细胞因子的分泌免疫细胞系和costimulatory分子的表达主要树突状细胞(131年]。一个可能的机制是群体感应。细菌群体感应是一种交流的方式在同一物种的协调效应功能,比如生物膜的形成、孢子形成或毒素分泌。的最佳描述群体感应分子酰基高丝氨酸内酯(ahl分子)132年]。几个ahl分子的目标病原体的宿主干涉增长(133年),反过来利用细菌为了自身利益,调节宿主基因的表达。一些ahl分子可以绑定到不同的苦味受体表达在气道上皮细胞(134年和先天和适应性免疫细胞135年,136年),从而调节障碍和免疫功能。人类最AHL 3-O-C12-HSL,可以激活吞噬细胞吞噬作用增加,粘附受体的表达和趋化性137年,138年),但本身是由呼吸道上皮细胞裂解和灭活(133年]。
的另一个例子bacterial-derived调节器宿主的免疫反应的外膜囊泡(omv) [139年]。omv是球形的双层膜囊泡释放表面革兰氏阴性和革兰氏阳性细菌,并包含父细菌的生物材料,但nonreplicative形式(140年,141年]。证据表明omv的释放为细菌提供了竞争优势当暴露在急性和慢性host-associated压力(142年]。他们可能保护细菌免受先天和适应性免疫反应(143年- - - - - -145年),抗菌肽与抗生素146年,147年]。此外,omv包含因素(如。含铁细胞)协助收购营养缺乏的环境中至关重要的元素,如铁(147年]。除了支持父细菌的生存,omv也可能发挥作用在肺部疾病的进展。已知细菌常与慢性阻塞性肺病急性加重释放omv [148年]。此外,巨噬细胞刺激与omv来源于著名的呼吸道病原体等铜绿假单胞菌,流感嗜血杆菌或复活的释放大量的肿瘤坏死factor-α和il - 6 (148年]。军团菌派生的omv显著增强细菌复制在巨噬细胞149年),无菌铜绿假单胞菌omv已被证明有说服力地诱导小鼠肺部炎症(150年诱发疾病),加强omv的想法发挥活动。此外,omv诱导宽容和hyporesponsiveness,从而促进细菌的坚持和掩饰的巨噬细胞,这可能导致间隙的感染149年,151年]。因此,尽管我们增加知识omv及其潜在作用interkingdom沟通,需要进一步的研究来更好地理解他们的致病性属性和可能的治疗或预防的影响(如。新型疫苗)。
一个有趣的替代FMT或益生菌可能使用免疫调节微生物的代谢产物或omv“有益的”。这种化学定义细菌物质可以在大规模生产在受控条件下,应用于定义(有效)剂量系统或局部,并有可能不利的副作用更少,即。在免疫功能低下的患者,而活细菌(152年]。几项研究已经使用细菌代谢物治疗小鼠AAD的定义。有限合伙人的大肠杆菌O111(153年),细菌多糖(154年),与细菌oligodeoxynucleotides CpG主题(155年),鞭毛蛋白B (156年),短链脂肪酸(47],D-tryptophan [157年)和neutrophil-activating蛋白质幽门螺杆菌(158年所有2型辅助细胞介导的免疫反应抑制气道嗜酸性粒细胞和AAD的小鼠模型。同样地,几个分子寄生虫的寄生虫已经被确认,其中excretory-secretory (ES) -62和半胱氨酸蛋白酶抑制物(AvCystatin)Acanthocheilonema个人简历TGF-β模仿(电报)Heligmosomoides polygyrusalarmin释放抑制剂(HpARI)h . polygyrus和IL-4-inducing原理从血吸虫卵(IPSE)是最有前途的,与影响在过敏性哮喘模型(159年- - - - - -163年]。
然而,有很大的局限性有关细菌或寄生虫的代谢产物用于治疗或预防方法。这些分子有一个非常狭窄的窗口的有益和治疗剂量,可能很难控制在当地组织。此外,行动的详细机制在很大程度上仍未知,阻碍人类研究翻译。最后,单一的代谢物,正如单一菌株,可能不是足够有效的治疗或预防与高度异常复杂的慢性疾病肠道和肺微环境。因此,迫切需要进一步的研究来确定行动的模式,而且单个或组合的功效代谢物发展小说metabolite-based治疗方案。
结合宿主和微生物有针对性的治疗方法
虽然microbiome-based疗法是耐人寻味的,重要的是要恢复肺的生态位。病变肺形成了一个完全不同的生境微生物偏爱不同的微生物菌群。囊肿性纤维化和支气管扩张嗜血杆菌和假单胞菌种虫害日益主导着肺部微生物群,而特定抗生素治疗进一步促进他们的结果89年,90年]。这可以减少竞争的结果从共生的抗生素,摧毁或持续异常的生态位肺囊性纤维化。支持后者,最近的一项研究表明,使用ivacaftor增加患者的雌性生殖道功能G511D突变,从而部分地恢复黏膜纤毛的清除,迅速降低铜绿假单胞菌负担在病人的痰164年]。然而,铜绿假单胞菌没有根除,结果呢铜绿假单胞菌观察治疗后1年,这可能是由于不完全恢复的利基164年]。
因此,它似乎是不可避免的,为了恢复慢性肺疾病的宿主转移平衡,有必要恢复上皮屏障和修复功能。未能实现这种可能导致复发的微生物生态失调。恢复上皮细胞功能可以通过诱导表达抗菌肽通过维生素D治疗(也有抗炎作用)或由farm-dust恢复屏障和抗病毒免疫治疗(27]。
结论和知识空白
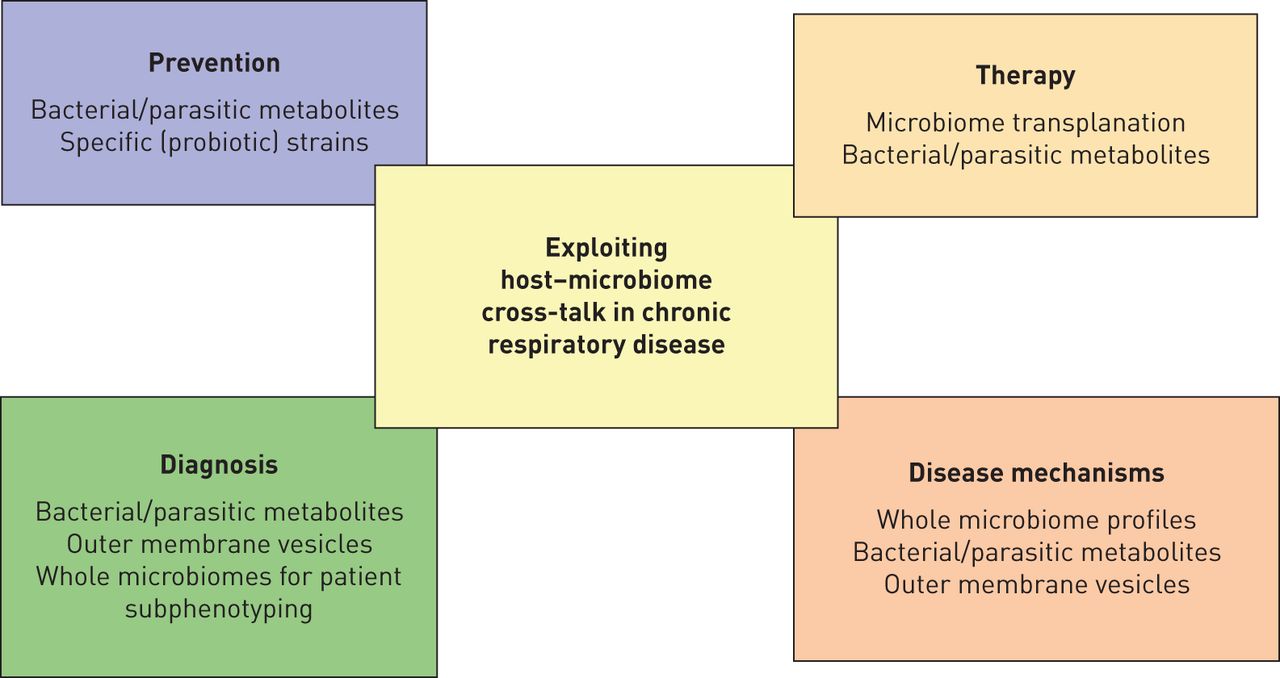
有大量的证据的功能参与慢性呼吸道疾病的微生物,使其未来疗法(一个有趣的目标图2)。针对微生物通过FMT似乎鼓励;然而,这仍然是一个迫切需要更好地理解复杂host-microbiome相声。还有小气道微生物群之间的因果关系信息和疾病发作/进展,除了一些最初的暗示。宿主遗传,转录组和代谢组学将有助于解决这些问题,并提供新范式的有助于疾病的严重程度和发作。虽然现有的治疗影响微生物,可能影响其有效性、安全性、最佳时间点和基于整个微生物剂有效的干预措施,代谢产物或其组合需要阐明在(纵向)的多中心临床试验中。此外,微生物可能是病人subphenotypes和治疗反应的良好指标,应进一步研究在临床样本,disease-relevant先进文化模型和代表在活的有机体内在动物模型(图2)。
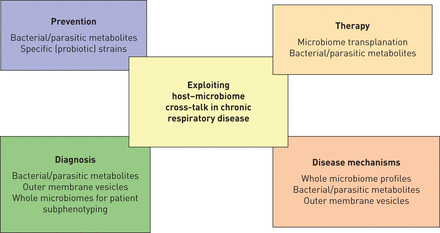
利用机会host-microbiome相声在慢性呼吸道疾病。宿主的相互作用的复杂性在肺微环境研究时应考虑慢性肺部疾病。理解这个相声的详细机制可能会导致新的治疗方法的发展,预防策略或高级诊断工具来区分subphenotypes病人。
可共享的PDF
确认
作者感谢人研究研讨会的参与者“相声肺微环境,影响理解慢性肺疾病”(柏林,2018年2月)的富有成果的讨论导致开发的评审。作者按字母顺序列出,除了Reinoud Gosens, Pieter Hiemstra Sabine Bartel。
脚注
利益冲突:r . Gosens报告从诺华赠款,勃林格殷格翰的发言,Aquilo和基耶西,在提交工作。
利益冲突:注:Hiemstra报告赠款从勃林格殷格翰集团和加拉帕戈斯喷嘴速度外,提交工作。
利益冲突:贝聿铭爱德考克报告个人费用从阿斯利康和Sunovion讲座,从勃林格殷格翰集团资助,在提交工作。
利益冲突:马克投资者没有披露。
利益冲突:r。迪克森没有披露。
利益冲突:下午Hansbro没有披露。
利益冲突:美国Krauss-Etschmann没有披露。
利益冲突:得出史密特没有披露。
利益冲突:F.R.M.斯达森没有披露。
利益冲突:美国Bartel赠款和个人费用咨询委员会工作报告从Bencard过敏GmbH是一家,在提交工作。
支持声明:下午Hansbro由奖学金和助学金资助来自国家卫生和医学研究委员会(# 1079187,1079187),彩虹和悉尼科技大学的基础。Krauss-Etschmann是莱布尼茨的成员竞争2016“肺微生物群在气道上皮细胞和环境”之间的接口和莱布尼茨科学校园“Evolung”。得出支持史密特ZonMW-VIDI格兰特(91714352)从荷兰科学研究组织,第三世史密特和另外Hiemstra财团支持格兰特(# 5115015)从荷兰肺脏基金会(NLF)和由格兰特(# 12017001)从独立加速计划。r . Gosens和另外Hiemstra受到一个财团的支持格兰特(6114009)独立。资金信息,本文已沉积的Crossref资助者注册表。
- 收到了2018年11月30日。
- 接受2020年4月20日。
- 版权©2020人队
这个版本分布在创作共用署名非商业性许可证的条款4.0。











{kind=link}
{kind=link}
{kind=link}
{kind=link}