





文摘gydF4y2Ba
空气中可吸入颗粒物(PM)可能诱发或加剧中性气道疾病通过触发炎症介质的释放,如科学家趋化因子配体(CXCL) 8,气道上皮细胞。目前还不清楚哪些组件下午开车CXCL8反应,因为大多数候选人都发生在低浓度粉尘。因此,我们假设,不同的点成分可以通过常见的机制诱导CXCL8贡献。gydF4y2Ba
人类支气管上皮细胞(BEAS-2B)暴露在不同的点组件(锌gydF4y2Ba2 +gydF4y2Ba/铁gydF4y2Ba2 +gydF4y2Ba盐、1-nitropyrene、脂多糖和柴油废气/矿物颗粒)。基因表达模式被实时PCR检测到数组中。CXCL8反应被实时PCR和ELISA测定。CXCL8监管评估与抑制剂面板和中和抗体。表皮生长因子受体(EGFR)磷酸化检测免疫沉淀反应和免疫印迹。gydF4y2Ba
Component-induced基因表达主要是与核factor-κB CagydF4y2Ba2 +gydF4y2Ba/蛋白激酶C、磷脂酶C、低密度脂蛋白和促有丝分裂的信号。下午许多抑制剂引起的减毒CXCL8释放所有组件,但在不同区段。然而,EGFR抑制强烈降低CXCL8释放引起的所有测试化合物和所选化合物增加了表皮生长因子受体磷酸化。干扰转化生长因子(TGF) -α或肿瘤坏死factor-α-converting酶(TACE)介导TGF-αectodomain脱落,还减毒CXCL8释放。gydF4y2Ba
不同的点成分诱导CXCL8部分是通过类似的信号途径的相对重要性不同的途径不同。然而,别说话/ TGF-α/表皮生长因子受体信号似乎收敛通路调节气道上皮细胞的先天免疫反应在接触多种空气污染物。gydF4y2Ba
空气中可吸入颗粒物(PM)可能诱发或加剧气道疾病通过肺部炎症反应。气道上皮细胞中发挥着关键作用在这些过程中通过调节先天免疫反应在吸入有毒化合物(gydF4y2Ba1gydF4y2Ba- - - - - -gydF4y2Ba3gydF4y2Ba]。上皮细胞表达的模式识别受体,包括toll样受体(通常),承认守恒的病原体图案和触发主机响应由各种感受器,如细胞因子和趋化因子(gydF4y2Ba4gydF4y2Ba]。我们最近表明,水平的提高neutrophil-selective科学家趋化因子配体(CXCL) 8 /白介素8 (IL)是一个主导炎性反应点和点组件在支气管上皮BEAS-2B细胞(gydF4y2Ba5gydF4y2Ba- - - - - -gydF4y2Ba7gydF4y2Ba]。明显CXCL8水平升高,嗜中性,也特征的气道疾病,如慢性阻塞性肺疾病(COPD),囊性纤维化(CF)和严重的哮喘gydF4y2Ba3gydF4y2Ba,gydF4y2Ba8gydF4y2Ba,gydF4y2Ba9gydF4y2Ba]。中性粒细胞在肺疾病的重要性得到了注意增加中性粒细胞炎症似乎对皮质类固醇治疗,和CXCL8因此被认为是一个潜在的治疗目标gydF4y2Ba9gydF4y2Ba,gydF4y2Ba10gydF4y2Ba]。CXCL8也可能有助于癌症发展通过其促炎效应及其潜在的促有丝分裂的作用,血管生成和motogenic因子(gydF4y2Ba11gydF4y2Ba]。gydF4y2Ba
多个信号通路促进CXCL8监管。在大多数细胞类型,其转录取决于核因子(NF) -κB激活,同时激活蛋白(美联社)1和CCAAT / enhancer-binding蛋白质需要最大的基因表达(gydF4y2Ba12gydF4y2Ba]。最大CXCL8反应进一步激活需要结合三大增殖作用的蛋白激酶(MAPK)级联:细胞外signal-regulated激酶(ERK) 1/2, c-Jun n端激酶(物)和p38 [gydF4y2Ba13gydF4y2Ba,gydF4y2Ba14gydF4y2Ba]。一系列的研究也表明,参与的表皮生长因子受体(EGFR)在CXCL8反应引起的不同的代理gydF4y2Ba15gydF4y2Ba- - - - - -gydF4y2Ba21gydF4y2Ba)、表皮生长因子受体和CXCL8表达式关联CF和严重哮喘患者的气道上皮细胞(gydF4y2Ba22gydF4y2Ba,gydF4y2Ba23gydF4y2Ba]。因此,表皮生长因子受体信号被建议作为一种收敛CXCL8进行监管的途径和其他免疫反应(gydF4y2Ba23gydF4y2Ba]。最近的研究与内生微生物由表皮生长因子受体配体表明CXCL8监管涉及转化生长因子(TGF) -αectodomain metalloprotease脱落的肿瘤坏死factor-α-converting酶(TACE) / ADAM-17 [gydF4y2Ba20.gydF4y2Ba,gydF4y2Ba21gydF4y2Ba,gydF4y2Ba24gydF4y2Ba,gydF4y2Ba25gydF4y2Ba]。下午是否有机或无机组件规范CXCL8通过类似的机制还有待阐明。gydF4y2Ba
机载点是一个复杂的、混合变量包含一些小部件(质量)相当大的基因调控潜力。这些包括木糖醇、金属、多环芳烃等有机化合物,和超细combustion-derived粒子(gydF4y2Ba26gydF4y2Ba]。下午澄清的成分导致肺的不利影响是重要发展的更有效的减排策略改善室外空气质量,同时阐明潜在的机制,这些影响可能改善治疗敏感的群体。特定的难题是,从不同的位置点显示很少的风险估计的变化,尽管尘埃的构成相当大的变化(gydF4y2Ba26gydF4y2Ba]。下午我们提出不同的组件的贡献gydF4y2Ba通过gydF4y2Ba常见的机制诱导CXCL8反应气道上皮细胞。如果几个组件通过一个类似的机制,这就可以解释为什么点可能引起的影响尽管其个人有效成分低浓度,以及为什么点组成的变化并不一定导致相当大的变化的影响。评估这个,我们调查了CXCL8监管BEAS-2B细胞。石英、锌gydF4y2Ba2 +gydF4y2Ba和菲gydF4y2Ba2 +gydF4y2Ba盐,1-nitropyrene (1-NP)和细菌脂多糖(LPS)被选出的模型点的不同组件gydF4y2Ba5gydF4y2Ba]。比较实验与柴油机尾气颗粒(DEPs)也被执行。gydF4y2Ba
材料和方法gydF4y2Ba
试剂gydF4y2Ba
石英(MIN-U-SIL®5)请提供美国硅公司(美国马里兰州弗雷德里克)。DEPs被F.R. Cassee请提供(国家公共卫生和环境研究所,Bilthoven荷兰)。ZnClgydF4y2Ba2gydF4y2Ba,FeSOgydF4y2Ba4gydF4y2Ba1-NP,有限合伙人,以叔(2-aminophenoxy)乙烷-gydF4y2BaNgydF4y2Ba,gydF4y2BaNgydF4y2Ba,gydF4y2BaNgydF4y2Ba′,gydF4y2BaNgydF4y2Ba′-tetraacetic酸tetrakis (acetoxymethyl酯)(BAPTA-AM)从Sigma-Aldrich(圣路易斯,密苏里州,美国)。SB202190, PD98059 SP600125蛋白磷酸酶2 AG1478, Go6976, rottlerin, GF109203, H-8渥曼青霉素,AG490,维拉帕米,支W7、姜黄素、肿瘤坏死factor-α蛋白酶抑制剂(TAPI) 1和TGF-α中和抗体都从Calbiochem (La Jolla、钙、美国)。表皮生长因子受体中和抗体(LA1)从微孔(美国Billerica的)。所有其他化学物质从商业来源购买可用的最高纯度。gydF4y2Ba
细胞培养和曝光gydF4y2Ba
BEAS-2B细胞腺病毒12-simian病毒40 hybrid-transformed人类支气管上皮细胞系,是从美国购买的组织文化类型集合(罗克维尔市,医学博士,美国)和生长在无血清LHC-9 near-confluence介质(Biosource、打击士气、钙、美国)37°C在湿润的气氛中5%的有限公司gydF4y2Ba2gydF4y2Ba在空气中,前曝光。浓度和孵育时间的测试化合物和抑制剂/中和抗体是基于以前的研究与BEAS-2B或其他肺上皮细胞(gydF4y2Ba5gydF4y2Ba,gydF4y2Ba7gydF4y2Ba,gydF4y2Ba18gydF4y2Ba,gydF4y2Ba19gydF4y2Ba,gydF4y2Ba27gydF4y2Ba- - - - - -gydF4y2Ba31日gydF4y2Ba),或首次放映BEAS-2B细胞(数据未显示)。gydF4y2Ba
基因表达gydF4y2Ba
总RNA是使用绝对孤立的RNA Miniprep工具包(美国Stratagene拉霍亚,CA),和反向转录cDNA PCR系统2400(美国马PerkinElmer,沃尔瑟姆)使用高容量cDNA存档工具包(美国应用生物系统公司,培育城市,CA)。基因表达是筛选利用信号转导PathwayFinder实时PCR数组(SABioscience,弗雷德里克,医学博士,美国),运行在ABI 7500快(应用生物系统公司)和正常化与平均表达式看家基因(gydF4y2BaB2MgydF4y2Ba,gydF4y2BaHPRT1gydF4y2Ba,gydF4y2BaRPL13AgydF4y2Ba,gydF4y2BaGAPDHgydF4y2Ba和gydF4y2BaACTBgydF4y2Ba)。结果表示为一个褶皱变化比未经处理的控制。基因被分组在信号通路根据制造商的建议(SABioscience)。DEP-induced CXCL8表达式用预先设计测量TaqMan基因表达分析和PCR TaqMan通用主混合(应用生物系统公司)。CXCL8表达式(Hs00174103_m1)正常化对18 s核糖体RNA (Hs99999901_s1)。gydF4y2Ba
CXCL8释放gydF4y2Ba
CXCL8细胞上清液中蛋白质含量测定ELISA (Biosource)如前所述gydF4y2Ba5gydF4y2Ba]。吸光度测量使用板读者(TECAN日出;美国凤凰研究产品,海沃德,CA)和麦哲伦软件(版本1.10;凤凰研究产品)。gydF4y2Ba
免疫沉淀反应gydF4y2Ba
表皮生长因子受体免疫沉淀反应是前面描述的(gydF4y2Ba18gydF4y2Ba]。简单地说,在冰冷的PBS细胞被洗了两次,一次冰冷的免疫沉淀反应(IP)缓冲区和细胞溶解在IP缓冲区NP-40为1%。细胞溶解产物在2500×离心机gydF4y2BaggydF4y2Ba10分钟,孵化anti-EGFR抗体(美国细胞信号技术,贝弗利,MA) 1 h和孵化与磁珠在一夜之间。包含0.5%的复合物在IP缓冲洗NP-40沸腾和免疫沉淀反应的2×Laemmli缓冲5分钟。EGFR磷酸化通过免疫印迹检测anti-phosphotyrosine抗体(p -酪氨酸- 100;细胞信号技术)。gydF4y2Ba
统计分析gydF4y2Ba
统计学意义是评估使用棱镜(GraphPad软件公司,圣地亚哥、钙、美国),使用方差分析Bonferroni测试后。gydF4y2Ba
结果gydF4y2Ba
Component-induced基因表达谱gydF4y2Ba
比较的影响结晶二氧化硅粒子,锌gydF4y2Ba2 +gydF4y2Ba、铁gydF4y2Ba2 +gydF4y2Ba1-NP,有限合伙人在BEAS-2B细胞,细胞信号通路的变化考察了84个基因的表达。所示gydF4y2Ba图1gydF4y2Ba26基因调节双重曝光后至少一个以上的化合物,在12个表达下调超过两倍,而控制。结果显示相当大的量变,通常1-NP和二氧化硅反应最强的诱导,以及一些定性的变异类型的基因激活。与之前的研究相一致(gydF4y2Ba5gydF4y2Ba),CXCL8是最敏感的基因。此外,所有化合物诱导显著upregulation CC趋化因子配体(CCL) 20(巨噬细胞炎性protein-3α)、肿瘤坏死因子(TNF) -α,集落刺激因子2(集落刺激因子(gm - csf)), c-Jun(主要AP-1单元)和凋亡基因gydF4y2BaBCL2A1gydF4y2Ba(不可靠由于低基因表达水平在大多数样品)。是根据检查结果以来,只有一个集中在一个时间点的影响,应采取一些谨慎当口译mRNA的数据。然而,相对影响CXCL8 CCL20和TNF-α可比先前的发现(gydF4y2Ba5gydF4y2Ba]。gydF4y2Ba
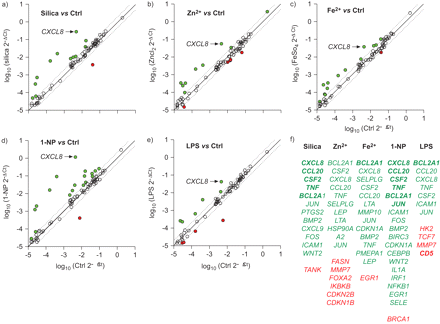
散点图的不同颗粒物组件引起的基因表达在人类支气管上皮细胞(BEAS-2B)。细胞培养是孵化10 h)石英(μg·40厘米gydF4y2Ba−2gydF4y2Ba),b) ZnClgydF4y2Ba2gydF4y2BaFeSO(90μM), c)gydF4y2Ba4gydF4y2Ba(250μM), d) 1-nitropyrene (1-NP)(30μM)或e)脂多糖(LPS)(10μg·毫升gydF4y2Ba−1gydF4y2Ba)。测量84个基因的mRNA水平变化实时PCR数组在材料和方法部分描述。结果表示为日志gydF4y2Ba10gydF4y2Ba(2gydF4y2Ba−ΔCtgydF4y2Ba)−ΔCt是每个基因的周期时间调整后的平均周期阈值(Ct)看家基因,和距离的实线代表了日志gydF4y2Ba10gydF4y2Ba叠化的价值比未经处理的控制(Ctrl)。gydF4y2Ba :两方面的变化;格林:基因调节超过两倍;红色:基因表达下调2倍以上。数据表示的两个独立的实验。f)表——(绿色)或表达下调的基因(红色)超过两个不同的测试化合物。大胆的表明> 10倍改变表达与控制。gydF4y2Ba
:两方面的变化;格林:基因调节超过两倍;红色:基因表达下调2倍以上。数据表示的两个独立的实验。f)表——(绿色)或表达下调的基因(红色)超过两个不同的测试化合物。大胆的表明> 10倍改变表达与控制。gydF4y2Ba
作为第一步确定细胞内信号通路引起的测试化合物,84个基因在集群根据共同的上游调控机制。所示gydF4y2Ba表1gydF4y2Ba的不同的曝光主要诱导upregulation NF-κB有关的基因,CagydF4y2Ba2 +gydF4y2Ba/蛋白激酶C (PK),磷脂酶C (PL),低密度脂蛋白(LDL)和促有丝分裂的途径。gydF4y2Ba
筛选信号通路参与CXCL8监管gydF4y2Ba
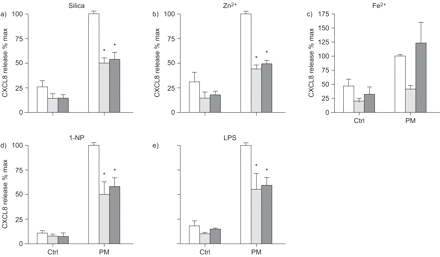
所有测试化合物也诱导显著增加CXCL8蛋白质释放(gydF4y2Ba图2gydF4y2Ba)。进一步检查CXCL8监管的机制,BEAS-2B细胞暴露在不同的测试化合物15抑制剂与电池相结合,针对不同的信号通路(gydF4y2Ba表2gydF4y2Ba)。通常,抑制剂似乎并不影响细胞的生存能力,根据显微镜(细胞密度减少和增加圆角/漂浮细胞),除了CagydF4y2Ba2 +gydF4y2Ba螯合剂BAPTA-AM AG490 Janus激酶(激酶)抑制剂,这有一些毒性(而不是量化)。此外,由于筛查是基于单一浓度和培养时间,结果的解释应该谨慎一些。gydF4y2Ba
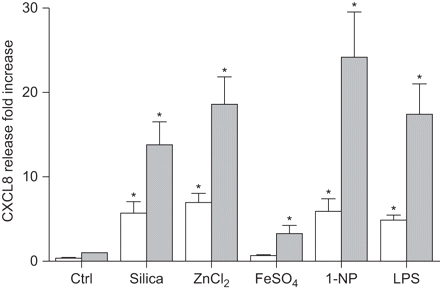
科学家诱导趋化因子配体(CXCL) 8释放后10 h(□)和20 h(▓)暴露于不同的颗粒物组件在人类支气管上皮细胞(BEAS-2B)。细胞培养是孵化与石英(40μg·厘米gydF4y2Ba−2gydF4y2Ba),ZnClgydF4y2Ba2gydF4y2Ba(90μM), FeSOgydF4y2Ba4gydF4y2Ba(250μM), 1-nitropyrene (1-NP)(30μM)或脂多糖(LPS)(10μg·毫升gydF4y2Ba−1gydF4y2Ba)。浮在表面的收获和CXCL8释放ELISA分析,在材料和方法部分描述。数据提出了相对增加(褶皱)比未经处理的控制(Ctrl)。数据意味着±gydF4y2Ba扫描电镜gydF4y2Ba三个或更多独立的实验。*:p < 0.05相比未曝光控制。gydF4y2Ba
而基因表达数据显示激活的CagydF4y2Ba2 +gydF4y2Ba/ PKC和促有丝分裂的信号,抑制屏幕进一步指出,这些途径参与CXCL8监管gydF4y2Ba图3gydF4y2Ba)。总的来说,结果表明,许多相同的抑制剂减少CXCL8释放引起的不同的测试化合物,但相对效果差异很大(gydF4y2Ba图3gydF4y2Ba)。三个主要MAPK通路的抑制剂通常CXCL8释放的有效抑制剂。然而,而p38抑制显著减毒CXCL8表达式由二氧化硅,锌gydF4y2Ba2 +gydF4y2Ba、铁gydF4y2Ba2 +gydF4y2Ba1-NP,只对有限合伙人边际效果观察。此外,ERK和物抑制显著降低铁的影响gydF4y2Ba2 +gydF4y2Ba,1-NP和有限合伙人,但并不影响二氧化硅和锌gydF4y2Ba2 +gydF4y2Ba全身的显著反应。PKCα/β-inhibitor增加背景CXCL8水平控制和提高锌的影响gydF4y2Ba2 +gydF4y2Ba和有限合伙人。同样,JAK2/3抑制剂CXCL8背景水平的增加,表明PKCα/β和JAK2/3在正常情况下抑制CXCL8-release BEAS-2B细胞。然而,PKCα1-NP /β和减毒CXCL8-induction JAK2/3抑制剂。这是否可能是由于非特异性的抑制剂或与能力的影响1-NP诱导CXCL8仍有待澄清。gydF4y2Ba
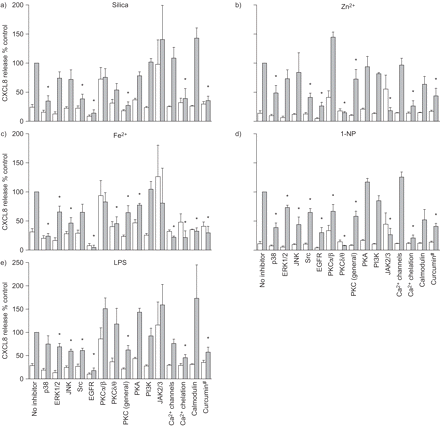
抑制剂屏幕识别途径参与科学家趋化因子配体(CXCL) 8释放引起的不同的颗粒物(PM)组件在人类支气管上皮细胞(BEAS-2B)。细胞培养是预处理的面板的15个不同的针对不同的信号通路抑制剂1 h前20 h孵化)石英(μg·40厘米gydF4y2Ba−2gydF4y2Ba),b) ZnClgydF4y2Ba2gydF4y2BaFeSO(90μM), c)gydF4y2Ba4gydF4y2Ba(250μM), d) 1-nitropyrene (1-NP)(30μM)或e)脂多糖(LPS)(10μg·毫升gydF4y2Ba−1gydF4y2Ba)。给出了使用的抑制剂gydF4y2Ba表2gydF4y2Ba。浮在表面的收获和CXCL8释放ELISA分析,在材料和方法部分描述。结果表示为相对CXCL8释放与点component-exposed细胞在缺乏抑制剂反应(100%)。数据意味着±gydF4y2Ba扫描电镜gydF4y2Ba(n = 3 - 6)。□:控制;▓:接触点组件。兵:细胞外signal-regulated激酶;物:c-Jun n端激酶;表皮生长因子受体:表皮生长因子受体;PK:蛋白激酶;PI3K:磷酸肌醇3-kinase;木菠萝:Janus激酶。gydF4y2Ba#gydF4y2Ba:目标多个通路。*:与控制相比p < 0.05。gydF4y2Ba
尽管一些差异,抑制剂屏幕还显示明显的相似之处:CXCL8释放引起的所有通用PKC抑制剂化合物明显减弱。此外,PKCδ/θ抑制剂rottlerin似乎降低CXCL8水平引起的所有测试化合物除了有限合伙人。另一个一般观察细胞内钙的gydF4y2Ba2 +gydF4y2Ba螯合剂BAPTA-AM或多或少地阻止CXCL8释放化合物,尽管只有铁gydF4y2Ba2 +gydF4y2Ba全身CXCL8降低了抑制剂的CagydF4y2Ba2 +gydF4y2Ba通道和钙调蛋白(gydF4y2Ba图3gydF4y2Ba)。然而,一些谨慎应该采取有关BAPTA-AM的影响,因为它似乎增加细胞毒性(数据没有显示)。姜黄素抗炎化合物,影响多种细胞内目标,包括NF-κB和AP-1 [gydF4y2Ba12gydF4y2Ba,gydF4y2Ba32gydF4y2Ba,gydF4y2Ba33gydF4y2Ba),也显示广泛CXCL8-inhibitory潜力。值得注意的是,表皮生长因子受体抑制剂AG1478或多或少完全封锁了二氧化硅的影响,铁gydF4y2Ba2 +gydF4y2Ba和有限合伙人CXCL8,强烈表达下调锌的影响gydF4y2Ba2 +gydF4y2Ba和1-NP (gydF4y2Ba图3gydF4y2Ba),这表明EGFR的中央监管机构CXCL8 BEAS-2B细胞。gydF4y2Ba
别说话的角色/ TGF-α/表皮生长因子受体途径在CXCL8监管gydF4y2Ba
表皮生长因子受体的作用CXCL8监管进一步评估使用EGFR-neutralising抗体和免疫沉淀反应了EGFR磷酸化(gydF4y2Ba图4gydF4y2Ba)。阻塞表皮生长因子受体配体结合CXCL8感应减少了所有点组件。此外,选择的测试化合物(石英和锌gydF4y2Ba2 +gydF4y2Ba)也诱导表皮生长因子受体磷酸化(gydF4y2Ba图4gydF4y2Ba)。因此,EGFR CXCL8监管的作用似乎涉及到配体绑定和受体激活。在此基础上,我们提出TACE-mediated乳沟pro-TGF-α可能是一个常见的由点组件CXCL8监管机制。因此,TACE抑制剂TAPI-1和TGF-α-neutralising抗体引起的减毒CXCL8释放所有测试化合物,除了铁gydF4y2Ba2 +gydF4y2Ba(gydF4y2Ba图5gydF4y2Ba)。铁的缺乏影响TGF-α中和gydF4y2Ba2 +gydF4y2Ba全身CXCL8令人吃惊,因为AG1478的影响,TAPI-1和EGFR中和。有可能是铁gydF4y2Ba2 +gydF4y2Ba可能引起的影响通过TACE-mediated劈理其他的表皮生长因子受体配体,如amphiregulin。gydF4y2Ba
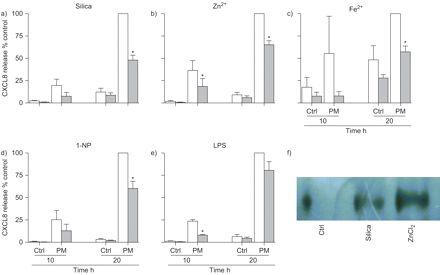
表皮生长因子受体(EGFR)的参与配位体绑定在颗粒物(PM) component-induced科学家趋化因子配体(CXCL) 8释放人类支气管上皮细胞(BEAS-2B)。细胞培养是预处理的anti-EGFR中和抗体1 h前20 h孵化)石英(μg·40厘米gydF4y2Ba−2gydF4y2Ba),b) ZnClgydF4y2Ba2gydF4y2BaFeSO(90μM), c)gydF4y2Ba4gydF4y2Ba(250μM), d) 1-nitropyrene (1-NP)(30μM)或e)脂多糖(LPS)(10μg·毫升gydF4y2Ba−1gydF4y2Ba)。浮在表面的收获和CXCL8释放ELISA分析,在材料和方法部分描述。结果表示为相对CXCL8感应而暴露出缺乏细胞中和抗体在20 h反应(100%)。数据意味着±gydF4y2Ba扫描电镜gydF4y2Ba(n = 3)。□:没有抑制剂;▓:anti-EGFR(10μg·毫升gydF4y2Ba−1gydF4y2Ba)。*:p < 0.05与控制(Ctrl)。f)两个选定的测试化合物对EGFR磷酸化的影响。BEAS-2B细胞孵化与石英(40μg·厘米gydF4y2Ba−2gydF4y2Ba)或ZnClgydF4y2Ba2gydF4y2Ba(90μM) 2 h表皮生长因子受体的免疫沉淀反应之前,在材料和方法部分描述。沉淀蛋白质被dodecylsulfate-polyacrylamide钠凝胶电泳分离和西方的屁股孵化抗体phosphotyrosine探讨表皮生长因子受体激活(n = 2)。gydF4y2Ba
参与肿瘤的坏死factor-α-converting酶(TACE)和转化生长因子(TGF) -α颗粒物(PM) component-induced科学家趋化因子配体(CXCL) 8版本从人类支气管上皮细胞(BEAS-2B)。细胞培养与TACE抑制剂预处理的肿瘤坏死factor-α蛋白酶抑制剂(TAPI) 1或TGF-α-neutralising抗体1 h前20 h孵化)石英(μg·40厘米gydF4y2Ba−2gydF4y2Ba),b) ZnClgydF4y2Ba2gydF4y2BaFeSO(90μM), c)gydF4y2Ba4gydF4y2Ba(250μM), d) 1-nitropyrene (1-NP)(30μM)或e)脂多糖(LPS)(10μg·毫升gydF4y2Ba−1gydF4y2Ba)。浮在表面的收获和CXCL8释放ELISA分析,在材料和方法部分描述。结果表示为相对CXCL8感应而暴露出缺乏细胞抑制剂或中和抗体反应(100%)。数据意味着±gydF4y2Ba扫描电镜gydF4y2Ba(n = 3)。□:没有抑制剂;░:TAPI-1;▒:TGF-α-neutralising抗体。*:p < 0.05与控制(Ctrl)。gydF4y2Ba
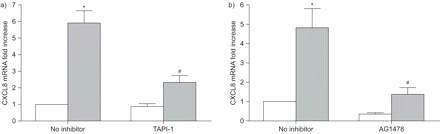
这些结果表明,多个模型化合物共同点的成分刺激BEAS-2B CXCL8释放细胞通过栓塞/ TGF-α/表皮生长因子受体的激活级联。探索更复杂的颗粒是否能通过类似的机制引起影响,细胞治疗DEPs缺席或AG1478和TAPI-1。因为DEPs可能绑定CXCL8蛋白质,从而干扰ELISA检测(gydF4y2Ba34gydF4y2Ba),CXCL8反应测定在mRNA水平。按照影响反应的模型化合物,EGFR和TACE抑制剂减毒DEP-induced CXCL8表达明显(gydF4y2Ba图6gydF4y2Ba)。gydF4y2Ba
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
参与肿瘤的坏死factor-α-converting酶(TACE)和表皮生长因子受体(EGFR)在柴油机排气微粒(DEP)全身的科学家趋化因子配体(CXCL) 8表达在人类支气管上皮细胞(BEAS-2B)。细胞培养是预处理与TACE抑制剂)肿瘤坏死factor-α蛋白酶inhibitor-1 (TAPI-1)或b)的表皮生长因子受体抑制剂AG1478 1 h前4 h与DEPs孵化(100μg·毫升gydF4y2Ba−1gydF4y2Ba)。CXCL8 mRNA水平变化是衡量实时PCR在材料和方法部分描述。结果表示为相对CXCL8感应(褶皱)而未曝光控制正常化后18 s核糖体RNA。数据意味着±gydF4y2Ba扫描电镜gydF4y2Ba(n = 3 - 5)。□:控制;▓:DEPs。*:p < 0.05与控制;gydF4y2Ba#gydF4y2Ba:没有抑制剂相比p < 0.05。gydF4y2Ba
讨论gydF4y2Ba
气道上皮细胞的先天免疫反应重要的宿主防御吸入污染物和病原体,但炎症介质从上皮细胞也可能促进发展和呼吸道疾病的恶化gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2gydF4y2Ba,gydF4y2Ba35gydF4y2Ba]。之前的探索细胞因子/趋化因子表达诱导的不同的点组件BEAS-2B细胞显示石英,锌gydF4y2Ba2 +gydF4y2Ba、铁gydF4y2Ba2 +gydF4y2Ba、1-NP和LPS诱导表达的至少一个neutrophil-specific趋化因子,处于受控,CXCL3, CXCL5 CXCL8。其中,只有CXCL8被所有代理商和诱导中最强烈的调节平均(gydF4y2Ba5gydF4y2Ba]。鉴于其核心作用在中性气道疾病(gydF4y2Ba9gydF4y2Ba,gydF4y2Ba10gydF4y2Ba,gydF4y2Ba36gydF4y2Ba),本研究开始探索的规定CXCL8 BEAS-2B细胞在接触点的组件。gydF4y2Ba
选择的点组件主要诱导基因表达与NF-κB, CagydF4y2Ba2 +gydF4y2Ba/ PKC、PLC、低密度脂蛋白和/或促有丝分裂的信号通路。通常强烈影响CXCL8证实可用数据表明CXCL8表达式是一个非常重要的上皮对吸入的空气污染物,如点。我们还发现显著影响gm - csf,从而防止中性粒细胞凋亡[gydF4y2Ba37gydF4y2Ba,gydF4y2Ba38gydF4y2Ba]。因此,点组件可能会触发的表达不仅epithelium-derived因素激活和招募中性粒细胞,也促进肺中性粒细胞持续生存的因素。gydF4y2Ba
通过应用一组15抑制剂干扰独立的信号通路,我们发现每个组件很可能激活几个信号通路,这大概一致行动CXCL8监管。不同的途径可以通过单独的触发机制,可能在不同浓度的复合问题。例如,多环芳烃可能开始通过与芳基碳氢化合物受体相互作用的影响,或形成活性代谢物和氧和氮自由基(活性氧(ROS) /活性氮物种)(gydF4y2Ba39gydF4y2Ba,gydF4y2Ba40gydF4y2Ba]。有趣的是,有限合伙人,触发它的影响通过TLR2/4激活gydF4y2Ba41gydF4y2Ba),一个相对具体的机制,诱导基因的最低数量的变化。变化的相对影响不同的抑制剂表明,尽管不同的化合物可以通过类似的途径,诱导CXCL8的相对重要性不同的信号通路各不相同。gydF4y2Ba
我们曾观察到表皮生长因子受体抑制大大减少了CXCL8释放引起的两个FgydF4y2Ba- - - - - -gydF4y2Ba和石英A549肺泡上皮细胞(gydF4y2Ba18gydF4y2Ba,gydF4y2Ba19gydF4y2Ba]。符合这一点,抑制剂屏幕发现表皮生长因子受体信号作为CXCL8监管BEAS-2B细胞的共同机制。使用EGFR-neutralising抗体进一步支持这个和牵连EGFR结扎CXCL8的规定。硅接触似乎触发立即磷酸化Src但不是表皮生长因子受体(gydF4y2Ba18gydF4y2Ba]。在这里,我们表明,在后来的跨度为受体被激活,这是惊人的相似,石棉在上皮细胞的影响gydF4y2Ba42gydF4y2Ba),这表明表皮生长因子受体激活,至少在矿物颗粒的情况下,是一个下游的事件而不是触发机制。gydF4y2Ba
表皮生长因子受体配体,如TGF-αamphiregulin,在支气管上皮细胞可以诱导CXCL8,似乎调解香烟烟雾诱发CXCL8通过自分泌环(gydF4y2Ba43gydF4y2Ba- - - - - -gydF4y2Ba45gydF4y2Ba]。这些生长因子产生膜结合pro-forms释放通过metalloproteases ectodomain脱落,主要是别说话/ ADAM-17 [gydF4y2Ba46gydF4y2Ba]。TLR配体(包括有限合伙人),鼻病毒和嗜中性粒细胞弹性蛋白酶诱导CXCL8解除气道上皮细胞通过激活的别说话,乳沟pro-TGF-α和激活的表皮生长因子受体(gydF4y2Ba20.gydF4y2Ba,gydF4y2Ba21gydF4y2Ba,gydF4y2Ba26gydF4y2Ba,gydF4y2Ba47gydF4y2Ba]。我们目前的数据证实,LPS-induced CXCL8反应涉及别说话/ TGF-α/ BEAS-2B细胞表皮生长因子受体信号。此外,化合物的研究扩展了列表使用这个路径CXCL8监管在气道上皮细胞内源性和微生物分子有机和无机空气污染物。我们也观察到,别说话,表皮生长因子受体抑制剂减毒DEP-induced CXCL8表达BEAS-2B细胞,表皮生长因子受体激活之间提供一个可能的机械的联系和增加CXCL8水平的航空公司DEP-exposed人类志愿者(gydF4y2Ba48gydF4y2Ba,gydF4y2Ba49gydF4y2Ba]。gydF4y2Ba
toll样受体——配基的中性粒细胞弹性蛋白酶和曝光,别说话在气道上皮细胞激活似乎涉及ROS生产双重氧化酶(Duox) 1gydF4y2Ba20.gydF4y2Ba,gydF4y2Ba25gydF4y2Ba]。氧化应激在细胞反应中也被认为是一个中央机制点(gydF4y2Ba50gydF4y2Ba]。因此,活性氧化合物形成直接的或间接的gydF4y2Ba通过gydF4y2BaDuox1活化的影响可以解释为什么这样一个多样化的群点化合物可能收敛到相同的途径。值得注意的是,PKCδ/θ牵连的激活Duox1 / ROS /别说话/ TGF-α/表皮生长因子受体级联(gydF4y2Ba20.gydF4y2Ba,gydF4y2Ba51gydF4y2Ba]。根据这一点,我们观察到rottlerin显著降低CXCL8释放引起的锌gydF4y2Ba2 +gydF4y2Ba、铁gydF4y2Ba2 +gydF4y2Ba1-NP,部分由硅(但不明显)。然而,我们的数据不支持的角色PKCδ/θLPS-induced CXCL8,相比别人所提出的(gydF4y2Ba3gydF4y2Ba]。几项研究暗示IL-1α/β也很重要的监管CXCL8通过自分泌信号循环(gydF4y2Ba52gydF4y2Ba- - - - - -gydF4y2Ba54gydF4y2Ba,我们实验室的初步结果显示,IL-1-antagonising药物anakinra / Kineret(可溶性il - 1受体(IL-1R)拮抗剂)压制CXCL8感应通过所有测试化合物除了有限合伙人(未发表的结果)。IL-1Rs通常属于同一家族和使用许多相同的适配器/信号分子,包括MyD88和IL-1R-associated激酶(gydF4y2Ba43gydF4y2Ba]。因此有趣的推测,il - 1可能激活别说话,信号通路诱导由几个测试化合物聚集在il - 1的规定。事实上,IL-1β可能诱发脱落的表皮生长因子受体配体在其他细胞类型和CXCL8甚至可能产生这样的效果,导致自我循环(gydF4y2Ba55gydF4y2Ba]。gydF4y2Ba
从获得的数据外推细胞株有明显的陷阱中被永久地传颂。然而,与正常的人类支气管上皮细胞的研究表明CXCL8监管,别说话/ TGF-α/表皮生长因子受体信号并不局限于细胞系(gydF4y2Ba25gydF4y2Ba]。虽然不能排除其他途径从高危对象可能主要在上皮细胞,表皮生长因子受体表达增加哮喘患者的气道上皮,吸烟者和COPD患者和CF,肺表皮生长因子受体的表达和CXCL8关联严重哮喘患者(gydF4y2Ba3gydF4y2Ba]。因此,肝动脉化疗栓塞的动物模型揭示了组织水平升高在慢性阻塞性肺病gydF4y2Ba56gydF4y2Ba),TACE抑制剂减少中性粒细胞涌入过敏和nonallergic气道炎症(gydF4y2Ba57gydF4y2Ba]。值得注意的是,别说话不仅劈开生长因子参与CXCL8监管,而且蛋白质参与白细胞跨内皮的黏附和轮回,包括血管细胞粘附分子/细胞间粘附molecule-1,gydF4y2BalgydF4y2Ba-selectin和紧密连接分子(gydF4y2Ba58gydF4y2Ba]。因此,别说话的激活可能会触发一系列反应参与促进中性粒细胞炎症的航空公司。此外,也别说话/ TGF-α/表皮生长因子受体信号调节粘蛋白生产,导致恶化的哮喘,慢性阻塞性肺病和CF与嗜中性(gydF4y2Ba3gydF4y2Ba,gydF4y2Ba24gydF4y2Ba,gydF4y2Ba59gydF4y2Ba- - - - - -gydF4y2Ba61年gydF4y2Ba]。gydF4y2Ba
我们目前的数据证实,CXCL8监管是复杂的,涉及到的信号通路,可以想象一致行动引起最后的回应。因此,干扰一个通路可能不足以废除CXCL8反应引起的空气污染物,如点。然而,我们现在发现加强印象,别说话/ TGF-α/表皮生长因子受体是一个重要的收敛通路调节各种吸入引起的先天免疫反应的化合物在气道上皮细胞。它将澄清任何感兴趣的点不同组件之间的协同或其他可吸入刺激物别说话/表皮生长因子受体信号以及现有的炎症或呼吸道疾病是否可以降低阈值水平别说话/表皮生长因子受体激活。gydF4y2Ba
确认gydF4y2Ba
我们感谢大肠Lilleaas和t . Skuland(部门空气污染和噪音,环境医学,挪威公共卫生研究所,奥斯陆,挪威)对整个研究的专业协助。gydF4y2Ba
脚注gydF4y2Ba
支持声明gydF4y2Ba
工作在挪威研究委员会的支持下,通过环境、遗传和健康计划(资助165386年和160863年)。gydF4y2Ba
感兴趣的语句gydF4y2Ba
没有宣布。gydF4y2Ba
- 收到了gydF4y2Ba2010年11月3日。gydF4y2Ba
- 接受gydF4y2Ba2011年4月3日。gydF4y2Ba
- ©2011人队gydF4y2Ba
引用gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba










