





文摘
气道上皮细胞是一个中央效应组织过敏炎症和辅助2-driven上皮细胞(Th)类型的响应,如粘液分泌过多导致气流阻塞过敏性气道疾病。以前的在体外研究也表明,Th2细胞因子作为强有力的调节器的上皮离子运输和液体分泌,但是在活的有机体内影响气道过敏性炎症的离子传输仍然未知。
因此,我们诱导过敏性炎症的气管内的安装来自烟曲霉属真菌提取或interleukin-13老鼠和确定影响离子运输在本地气管和支气管组织。
我们证明过敏炎症增强基底Cl- - - - - -分泌气道地区和抑制上皮Na+通道(钠)介导Na+吸收和增加Ca2 +端依赖Cl- - - - - -在支气管分泌。Allergen-induced改变支气管离子运输与转录水平降低有关α,β,γENaC和信号传感器和催化剂的主要是废除转录(Stat) 6- / -老鼠。
我们的研究表明,Th2-dependent气道炎症产生pro-secretory离子运输表型在活的有机体内,这在很大程度上是Stat6-dependent。这些结果表明Th2-mediated液体分泌可能改善气道表面水化和清除粘液中hypersecreted过敏性呼吸道疾病,如哮喘,并确定上皮Stat6信号作为一个潜在的治疗目标促进粘液水化和气道间隙。
过敏性呼吸道疾病,如哮喘,属于儿童和成人最常见的慢性疾病,但其发病机制仍不完全清楚1。气道上皮细胞提供了重要的保护功能和构成中央效应组织过敏性气道疾病。在呼吸道上皮细胞表面健康的航空公司,作为一个屏障,提供了协同调节的粘液分泌,纤毛跳动和气道表面液体(ASL)体积,功能是必不可少的有效的黏膜纤毛的清除(MCC)和保护的肺部吸入过敏原,病原体和其他有毒物质2- - - - - -4。大量的证据表明,一种辅助细胞(Th) 1/2失衡与Th2-dominant回应吸入过敏原中扮演着一个关键的角色在过敏性气道炎症的发病机制和确定了Th2细胞因子白介素(IL) -13作为这个过程的关键效应分子1,5,6。进一步的研究表明IL-13,通过绑定到一个共同的il - 4 / IL-13受体信号传感器和催化剂的复杂和激活的转录(Stat) 6信号,对气道上皮细胞有直接影响,包括粘液分泌过多,杯状细胞化生及气道高反应性7关键特性,导致气流阻塞在哮喘的临床综合征。
此外,最近的在体外研究表明,IL-13和相关Th2细胞因子il - 4作为强有力的调节器的离子运输在人类支气管上皮细胞培养,识别的另一个直接影响这些Th2细胞因子在气道上皮细胞8,9。积极监管transepithelial离子运输、气道上皮细胞保持液体的薄膜覆盖气道表面和提供了一个低粘度层适当的纤毛跳动和MCC的必要条件3。在这个过程中,amiloride-sensitive上皮Na+通道(钠)是限制钠的吸收+和osmotically耦合流体10,而氯- - - - - -分泌囊性纤维化跨膜电导调节介导的CFTR(检测)11和一个Ca2 +激活的氯- - - - - -通道(中国),最近被确定为跨膜蛋白(TMEM) 1612,13。il - 4和IL-13抑制ENaC-mediated Na+吸收,增加雌性生殖道,CaCC-mediated Cl- - - - - -分泌,从而产生一个pro-secretory离子运输在培养人类支气管上皮细胞表型8,9。这些研究表明,选择Th2细胞因子可能参与美国手语的规定体积粘液促进水合作用和促进MCC在过敏性气道疾病。然而,Th2-driven炎症的作用在调节气道离子传输的复杂在活的有机体内过敏性气道炎症的发病机理仍然未知。
在目前的研究中,我们通过重复诱导小鼠过敏性炎症粘膜接触天然气源性致敏原,来自烟曲霉属真菌提取(Af)14,并研究了影响离子的属性和文本表达ENaC子单元(α,β和γ),雌性生殖道和中国商用飞机有限责任公司候选人TMEM16a刚切除气管组织。我们比较生物属性原生气管和支气管上皮细胞从Af-sensitised vehicle-treated老鼠阐明不同气道过敏性炎症的影响区域沿支气管树。此外,我们比较了Af-induced炎症对生物的影响在野生型(WT)和Stat6属性- / -老鼠,和研究IL-13的直接影响,为了获得机械的见解之间的联系Th2-driven炎症和气道离子运输的监管。这些研究表明首次allergen-induced气道炎症诱发pro-secretory离子运输表型在活的有机体内之间的相互作用,产生新颖的见解Th2-driven炎症和上皮功能复杂的过敏性气道疾病的发病机制,这可能会导致哮喘等呼吸道疾病的新的治疗策略与粘液清除受损有关。
材料和方法
实验动物
所有动物研究都Regierungsprasidium批准,德国卡尔斯鲁厄。雌性BALB / c WT老鼠从查尔斯河实验室(Sulzfeld,德国)。Stat6- / -在BALB / c小鼠的背景15从杰克逊实验室获得(巴尔港,我,美国)。实验动物被Stat6的杂交繁殖+ / -老鼠;后代基因组DNA被PCR基因分型,如前所述15,6-8-week-old Stat6- / -老鼠和WT垃圾伴侣是用于所有实验。老鼠被安置在一个特定的无菌动物设施和免费的食物和水。
过敏原接触
过敏性气道炎症是引起反复呼吸道敏化作用与房颤(美国WA Hollister-Stier实验室、斯波坎)根据先前建立协议14。总之,老鼠犀牛被吸入异氟烷氧含量(3%)和20μL Af (2 mg·毫升−1)溶解在0.9%生理盐水或0.9%氯化钠,被灌输气管内的每周3次,3周的时间。所有终点研究进行24 h后最后一个接触过敏原(图1一个)。
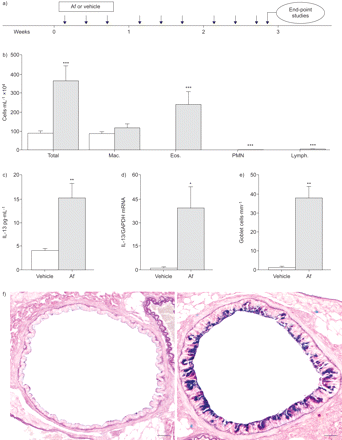
敏化作用引起的过敏性气道炎症来自烟曲霉属真菌提取(Af)的老鼠。与房颤)协议的气管内的敏化作用,b)影响总和微分细胞计数、c)白介素(IL) -13浓度支气管肺泡灌洗,d) IL-13转录水平肺,e)气道杯状细胞计数和f)在小鼠气道组织学(代表气道部分沾阿尔新blue-periodic酸希夫Af-sensitised和vehicle-treated老鼠;比例尺条= 100μm)敏感与房颤或车辆单独一段3周。提出了数据意味着±扫描电镜每组)(n = 5 - 8的老鼠。□:车辆;░:房颤。Mac:巨噬细胞;黎明的女神。:eosinophils; Lymph.: lymphocytes; GAPDH: glyceraldehyde 3-phosphate dehydrogenase. *: p<0.05 compared with vehicle-treated mice; **: p<0.01 compared with vehicle-treated mice; ***: p<0.001 compared with vehicle-treated mice.
IL-13滴注法
IL-13是由重复气管内的滴剂如前所述16。总之,5μg重组小鼠IL-13 (Peprotech,汉堡,德国)溶解在50μL磷酸盐(PBS)含0.1%牛血清白蛋白(low-endotoxin;σ,Steinheim,德国)或同等体积的车辆,仅是由气管内的滴注法每天一次连续三天。终点研究进行24 h后IL-13滴注法。
支气管肺泡灌洗细胞计数和IL-13测量
老鼠深深anesthetised通过结合腹腔注射氯胺酮和甲苯噻嗪(120和16 mg·公斤−1分别被放血,通过减少肾动脉。执行正中,气管插管,左主支气管是结扎和右肺灌洗了PBS如前所述17。支气管肺泡灌洗(BAL)样本离心机和浮在表面的游离是储存在-80°C。总活菌数测定使用血球计和差细胞计数测定沾May-Grunwald-Giemsa cytospin准备17。IL-13 BAL上层清液浓度是衡量ELISA(研发系统,明尼阿波利斯,美国)根据制造商的指示17。
组织学和气道形态测量学
通过左肺切除的正中,immersion-fixed neutral-buffered 10%福尔马林和嵌入在石蜡。肺脾横向水平的近端肺内的主要轴向气道。5-μm部分被切割和沾阿尔新blue-periodic酸希夫(AB-PAS)如前所述17。图像被抓获的ix - 71奥林巴斯显微镜与SIS Colorview我相机设置界面上的(奥林巴斯,汉堡,德国)使用10×目标。杯状细胞通过胞内AB-PAS-positive物质的存在,并数值细胞密度测定每毫米的基底膜使用交互式图像测量工具(细胞•F图像分析软件;奥林巴斯,汉堡,德国),如前所述17。
电致离子传输测量
犀牛的老鼠被放血和气道组织解剖使用立体显微镜如前所述18。总之,气管和主支气管的正中切口暴露颈部前部和胸骨切开术。气道组织被释放从周围的肌肉,血管和结缔组织由钝性剥离原位。接下来,气管和主支气管切除,前部纵向软骨环被割开,pars membranacea被暴露和组织灌注立即安装到micro-Ussing钱伯斯0.8毫米的圆形开放区域2为气管组织和0.5毫米2对支气管组织19。腔内灌注和基底浴的速度连续10毫升·分钟−1,解决以下成分:145毫米氯化钠,0.4毫米KH2阿宝4,1.6毫米K2HPO45毫米dMgCl葡萄糖,1毫米2葡萄糖酸钙,1.3毫米。pH值调整到7.4。实验在37°C在开路情况下如前所述19。总之,transepithelial电压值(Vte)被引用到浆膜的一面。transepithelial电阻(Rte)是由应用间歇(1)当前脉冲(δ我= 0.5μA)和等效短路电流(我sc)是根据欧姆定律来计算的Vte和Rte(我sc=Vte/Rte)。经过平衡的40分钟在我们室,基底我sc决心和阿米洛利(100μM腔的)被添加到抑制电致ENaC-mediated Na吗+吸收。然后,3-isobutyl-1-methylxanthine (IBMX;100μM腔的)和forskolin(1μM腔的)和尿苷5′三磷酸(UTP)(100μM腔的)顺序添加诱导环腺5′一磷酸(营)介导和Ca2 +激活的氯- - - - - -分泌,分别10。此外,我们做了个实验,布美他尼(100μM基底外侧),基底外侧Na的抑制剂+- k+2氯- - - - - -转运蛋白,添加在基础条件下或在阿米洛利阻止transepithelial Cl- - - - - -分泌20.。阿米洛利,IBMX forskolin和布美他尼从σ(Steinheim、德国)和UTP是来自通用电气医疗集团(英国哈特菲尔德)。所有的化学品都可用的最高年级的纯度。
实时rt - pcr
对肺部和气管都被立即存储在RNAlater®(应用生物系统公司,达姆施塔特,德国)。RNA被隔离使用试剂盒试剂(表达载体,卡尔斯鲁厄,德国)。转录表达研究气管组织导管从三个老鼠相同的基因型和治疗组汇集。RNA纯度和数量确定使用NanoDrop ND100分光光度计(PeqLab,埃朗根,德国)和完整性被琼脂糖凝胶电泳验证。互补脱氧核糖核酸是通过逆转录1μg总RNA(上标三世RT;英杰公司、德国卡尔斯鲁厄)。αENaC实时定量rt - PCR,βENaCγENaC,雌性生殖道,TMEM16a IL-13和3 -磷酸甘油醛脱氢酶(GAPDH)上执行应用生物系统公司7500实时PCR系统使用TaqMan普遍PCR大师混合和清点TaqMan基因表达分析根据制造商的指示(应用生物系统公司,达姆施塔特,德国)。相对褶皱的变化目标基因表达测定PCR反应的效率和交叉点之间的偏差Af-sensitised vehicle-treated老鼠,和正常的表达参考GAPDH基因,如前所述17。
统计数据
所有实验都是由一名调查员盲治疗组和老鼠的基因型。数据分析与SigmaStat version 3.1 (Systat软件、Erkrath德国),意味着±扫描电镜。我们使用未配对t进行统计分析- - - - - -测试,Mann-Whitney秩和检验、单因子变异数分析和克鲁斯卡尔-沃利斯是适当的方差分析,p < 0.05,显示统计学意义的截止。
结果
过敏性气道炎症的诱导气管内的敏化作用与房颤老鼠
先前的研究已经表明,il - 4和IL-13作为上皮离子运输的有力调节器在体外8,9;然而,这种监管功能复杂的角色在活的有机体内过敏性气道炎症的发病机制还没有研究。阐明在气道过敏性炎症离子传输的影响在活的有机体内,我们敏感的小鼠天然气源性致敏原,答:来自烟在一段时间内,通过反复气管内的滴剂3周(图1一个)。类似于之前的研究14,这种敏化作用产生一个健壮的协议过敏炎症的特点是气道嗜酸性粒细胞(图1 b),增加了Th2细胞因子的水平IL-13 BAL液(图1 c),肺组织IL-13转录水平增加(图1 d气道),杯状细胞化生及粘液分泌过多(无花果1 e和f)相比,Af-sensitised vehicle-treated老鼠。
过敏性炎症的影响在气管上皮离子运输
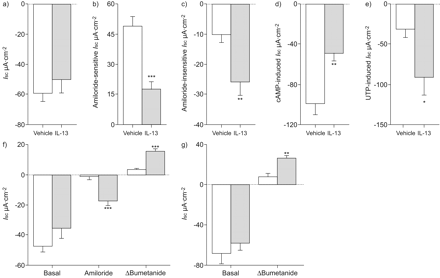
确定allergen-induced炎症的作用在调节气道离子运输在活的有机体内,我们首先研究了它对气管上皮细胞的影响,组织通常用于气道离子运输在老鼠身上的研究10,13,18。刚切除气管组织灌注是安装在micro-Ussing钱伯斯和基底生物属性,amiloride-sensitive Na+吸收,cAMP-mediated UTP-induced Cl- - - - - -分泌比较Af-sensitised和vehicle-treated老鼠。我sc在基础条件下,以及amiloride-sensitive我sc不是不同的气管组织Af-sensitised相比vehicle-treated老鼠(无花果2模拟)。amiloride-insensitive残留我sc在阿米洛利块被过敏性气道炎症显著增加(图2 e),而cAMP-induced (IBMX和forskolin)和Ca2 +激活(UTP) Cl- - - - - -分泌反应没有不同的气管组织Af-sensitised相比vehicle-treated老鼠(无花果2 f和g)。总的来说,这些结果说明过敏炎症amiloride-insensitive增加引起的我sc在本地气管组织。然而,我们的在活的有机体内模型的过敏性气道炎症没有产生调制ENaC-mediated Na+吸收或agonist-induced Cl- - - - - -分泌在本地气管组织。
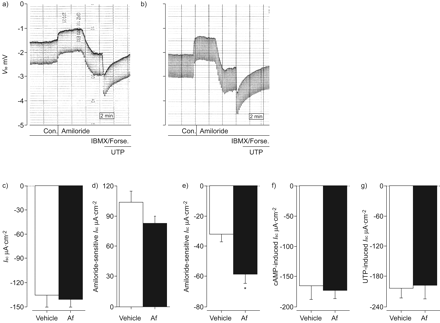
过敏性炎症的影响在本机气管上皮细胞离子运输。a, b)的原始录音阿米洛利的影响,环腺苷5′一磷酸(营)端依赖激活和尿苷5′三磷酸(UTP)端依赖激活transepithelial电压(Vte)和transepithelial电阻(Rte)刚切除气管组织从vehicle-treated和b)来自烟曲霉属真菌提取小鼠(Af)敏感。Rte决心从Vte变形量得到的脉冲电流注入。代表第27 - 31 n =老鼠每组的结果。c g)的总结c)基底等效短路电流(我sc),d) amiloride-sensitive我sc,e) amiloride-insensitive我scf) cAMP-induced我sc和g) UTP-induced我sc在刚切除气管组织从Af-sensitised vehicle-treated老鼠。数据意味着±扫描电镜第27 - 31 (n =老鼠每组)。缺点:控制;IBMX: 3-isobutyl-1-methylxanthine;可能。:forskolin. *: p<0.001 compared with vehicle-treated mice.
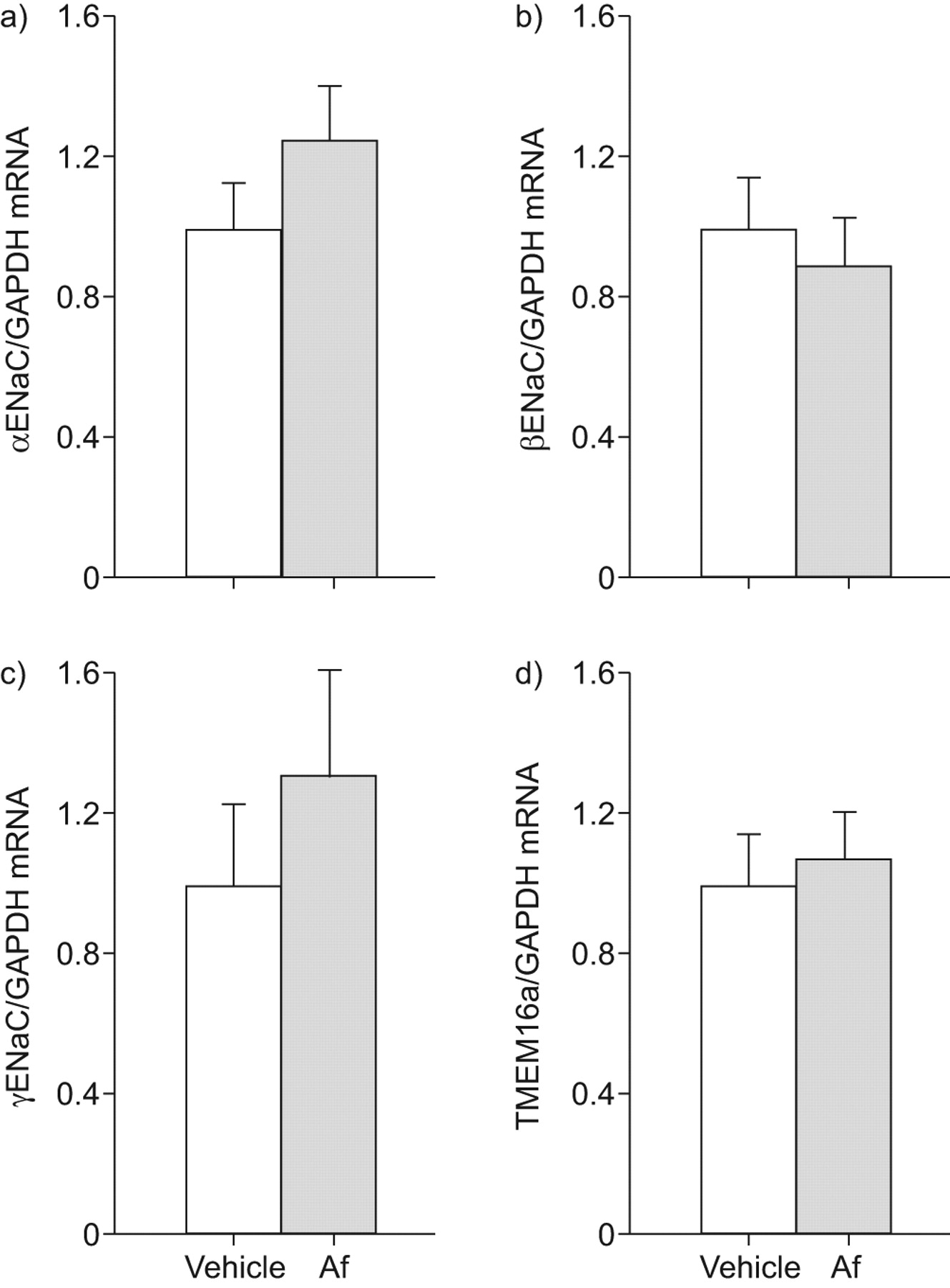
接下来,我们决定Af-induced气道炎症的影响在mRNA转录水平的这些离子通道在气管组织通过实时定量rt - pcr。符合从离子传输测量结果(图2),表达水平αENaC、βENaCγENaC TMEM16a并没有在气管组织区别于Af-sensitised相比vehicle-treated老鼠(图3)。同意之前的功能研究表明,雌性生殖道表示对小鼠气管处于非常低的水平18CFTR,信号检测记录低于检出限在气管组织从两组(数据没有显示)。
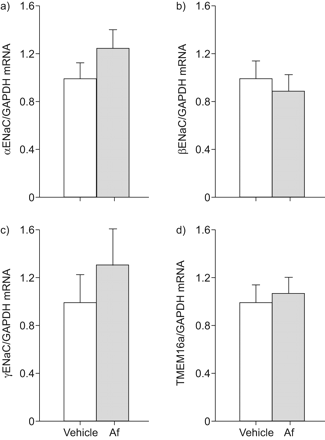
过敏性炎症对表达的影响上皮Na+通道(钠)子单元和跨膜蛋白在本机气管上皮细胞(TMEM) 16。模拟αENaC)转录水平),b)βENaC, c)γENaC和d) TMEM16a刚切除气管组织来自烟曲霉属真菌提取(Af)敏感和vehicle-treated老鼠。数据表示为褶皱变化vehicle-treated老鼠。数据意味着±扫描电镜。n = 5老鼠/组。
过敏炎症诱发支气管pro-secretory离子运输表型
为了解释不同的离子传输调制Th2细胞因子在本机气管组织(我们的研究无花果2和3),前在体外研究人类支气管上皮细胞培养8,9,我们提出,这些差异可能是由于地区差异在离子传输属性和/或监管沿着支气管树。为了验证这个假设,我们扩展研究过敏性气道炎症的房颤模型刚切除支气管组织。
首先,我们比较了生物属性刚切除支气管和气管组织从vehicle-treated WT老鼠来确定区域差异没有过敏炎症。所示表1几个不同的离子传输性质检测支气管与气管上皮细胞。在基准情况下,Rte不是两气道不同区域,而Vte和我sc在支气管与气管组织较低(表1)。总的来说,amiloride-sensitive的绝对值我sc(p < 0.001),剩余amiloride-insensitive我sccAMP-induced (p < 0.001)我sc(p < 0.01)和UTP-induced我sc(p < 0.001)显著降低在支气管与气管组织(表1)。然而,当不同我sc反应表示为基底的一小部分我sc在每一个组织,部分amiloride-sensitive很明显我sc(p < 0.01)以及成倍增加我sccAMP-dependent激活后(p < 0.001)显著较高,而部分amiloride-insensitive我sc(p < 0.01),以及成倍增加我sc为了应对UTP-induced激活(p < 0.05),显著降低支气管与气管组织。气管相比,这些结果表明,1)支气管组织更吸收在正常情况下,2)诱导Cl的相对能力- - - - - -在支气管分泌更大,3)Cl- - - - - -分泌的反应,更大程度上引发了营地受体激动剂,在较小程度上,通过Ca2 +端依赖激活本地支气管与气管上皮细胞。
接下来,我们决定过敏性炎症的影响在支气管离子运输通过比较新鲜的生物属性切除支气管组织从Af-sensitised vehicle-treated老鼠。类似于气管,基底我sc不是被allergen-induced炎症改变(图4 e)。与气管组织相比,amiloride-sensitive我sc显著抑制(p < 0.05)相比,刚从Af-sensitised切除支气管vehicle-treated老鼠(图4 f)。气管组织中观察到,amiloride-insensitive残留我sc是显著增加(p < 0.01)在支气管Af-sensitised老鼠(图4 g)。cAMP-mediated Cl- - - - - -在支气管分泌组织被过敏炎症(减毒图4 h),而UTP-induced Cl- - - - - -分泌反应明显增加(p < 0.05)在本地支气管组织Af-sensitised老鼠相比vehicle-treated控件(图4我)。综上所述,这些研究结果表明,抑制ENaC-mediated Na+吸收和Ca的老年病2 +激活的氯- - - - - -由过敏炎症分泌产生pro-secretory离子运输在支气管表型(图4 gydF4y2Ba)。
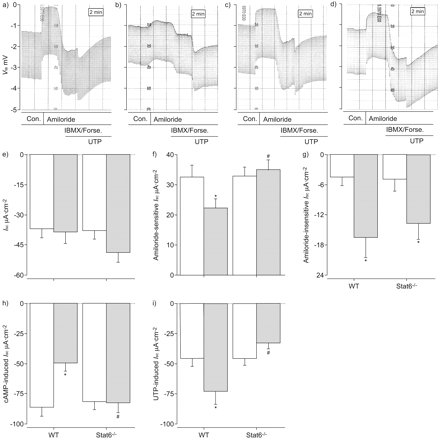
过敏炎症调节离子运输在本地信号传感器和催化剂的支气管上皮细胞转录(Stat) 6-dependent方式。模拟)的原始录音阿米洛利的影响,环腺苷5′一磷酸(营)端依赖激活和尿苷5′三磷酸(UTP)端依赖激活transepithelial电压(Vte)和transepithelial抵抗刚切除支气管组织的对面,c) vehicle-treated和b, d)来自烟曲霉属真菌(Af)敏感的野生型小鼠(WT) (a, b)和Stat6- / -同窝出生(c, d)。总结e)基底等效短路电流(我sc),f) amiloride-sensitive我sc,g) amiloride-insensitive我sch) cAMP-induced我sc和我)UTP-induced我sc在刚切除支气管组织Af-sensitised(░)和vehicle-treated WT老鼠和Stat6 (□)- / -同窝出生的。数据意味着±扫描电镜每组)(n =小鼠年龄在18岁至25岁之间。缺点:控制;IBMX: 3-isobutyl-1-methylxanthine;可能。:forskotin. *: p<0.05 compared with vehicle-treated mice of the same genotype;#:与Af-sensitised相比p < 0.005 WT老鼠。
Th2-mediated ENaC-mediated Na的调制+吸收和Ca2 +激活的氯- - - - - -在支气管分泌Stat6-dependent
先前的研究表明,Stat6中起着重要的作用的规定Th2-mediated过敏性哮喘气道炎症,包括气道嗜酸性粒细胞,粘液分泌过多和气道高反应性7,15,但Stat6信号的作用在调节上皮离子运输还没有被研究过。为了阐明气道的机制调节离子运输过敏性炎症,我们比较反应从WT和Stat6 Af支气管组织中的敏化作用- / -老鼠。所示图4对基底,Stat6击倒没有影响我sc在没有或存在Af-induced炎症(图4 e)。然而,抑制amiloride-sensitive Na+吸收是由过敏引起支气管的炎症在Stat6 WT老鼠完全废除- / -老鼠(图4 f)。虽然Af-induced upregulation amiloride-insensitive我sc观察到在Stat6 WT老鼠没有影响- / -同窝出生的(图4 g),同时减少在营里的反应,以及upregulation UTP-induced Cl- - - - - -观察到支气管分泌组织从Af-sensitised WT老鼠在Stat6完全废除- / -老鼠(无花果4 h和i)。这些结果说明Stat6信号是直接参与调节上皮离子运输过敏性气道疾病。
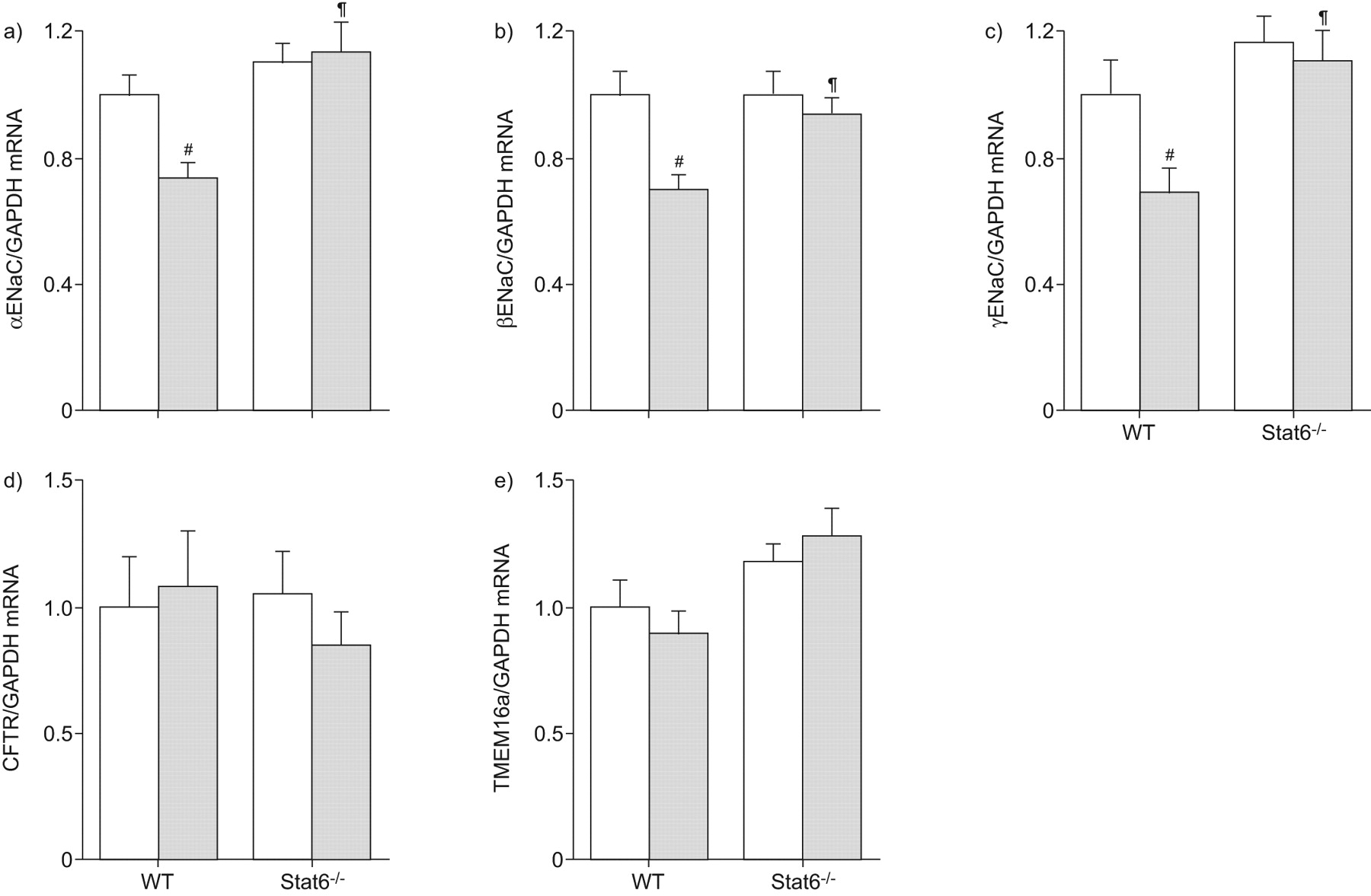
Stat6减少表达α,β和γENaC过敏性气道炎症
因为许多影响Stat6介导通过调节目标基因的表达7,15,21,我们下一个试图确定Stat6-dependent调节离子运输过敏炎症与转录水平的变化即,雌性生殖道和中国商用飞机有限责任公司候选人TMEM16a Af-sensitised小鼠的肺。这些研究在肺组织进行,因为mRNA的收益率从支气管组织太低可靠的量化这些基因的实时rt - pcr。符合离子传输测量结果在支气管组织(图4 f),过敏性炎症αENaC导致表达水平显著降低(p < 0.01),βENaC (p < 0.01)和γENaC (p < 0.05)从WT小鼠肺子单元(图5 a - c)。这种效果是完全废除Af-sensitised Stat6- / -老鼠(图5 a - c)。雌性生殖道和TMEM16a表达从Af-sensitised在肺部保持不变,与vehicle-treated相比,WT老鼠和Stat6转录水平并没有改变- / -老鼠在没有或过敏性炎症的存在(无花果5 d和e)。这些结果表明,Stat6-dependent过敏性气道炎症调节钠的功能是由钠基因的转录调控,而Stat6-dependent cAMP-mediated监管和UTP-induced Cl- - - - - -分泌物是独立的转录调节雌性生殖道或TMEM16a。
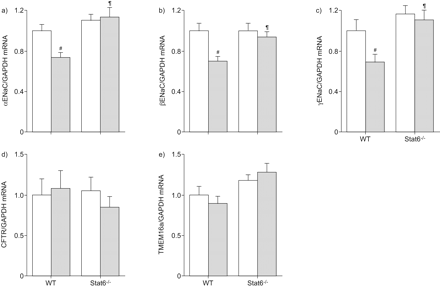
过敏炎症调节肺上皮Na的表情+通道(钠)的子单元,但不是囊性纤维化跨膜电导调节(雌性生殖道)和跨膜蛋白(TMEM) 16,信号传感器和激活的转录(Stat) 6-dependent方式。转录水平的αENaC, b)βENaC, c)γENaC, d)雌性生殖道和e) TMEM16a肺来自烟曲霉属真菌提取(Af)敏感(░)和vehicle-treated(□)野生型小鼠和Stat6 (WT)- / -同窝出生的。数据意味着±扫描电镜褶皱的变化从vehicle-treated WT老鼠(n = 5 - 6小鼠每组)。#:p < 0.05相比vehicle-treated WT老鼠;¶:与Af-sensitised相比p < 0.05 WT老鼠。
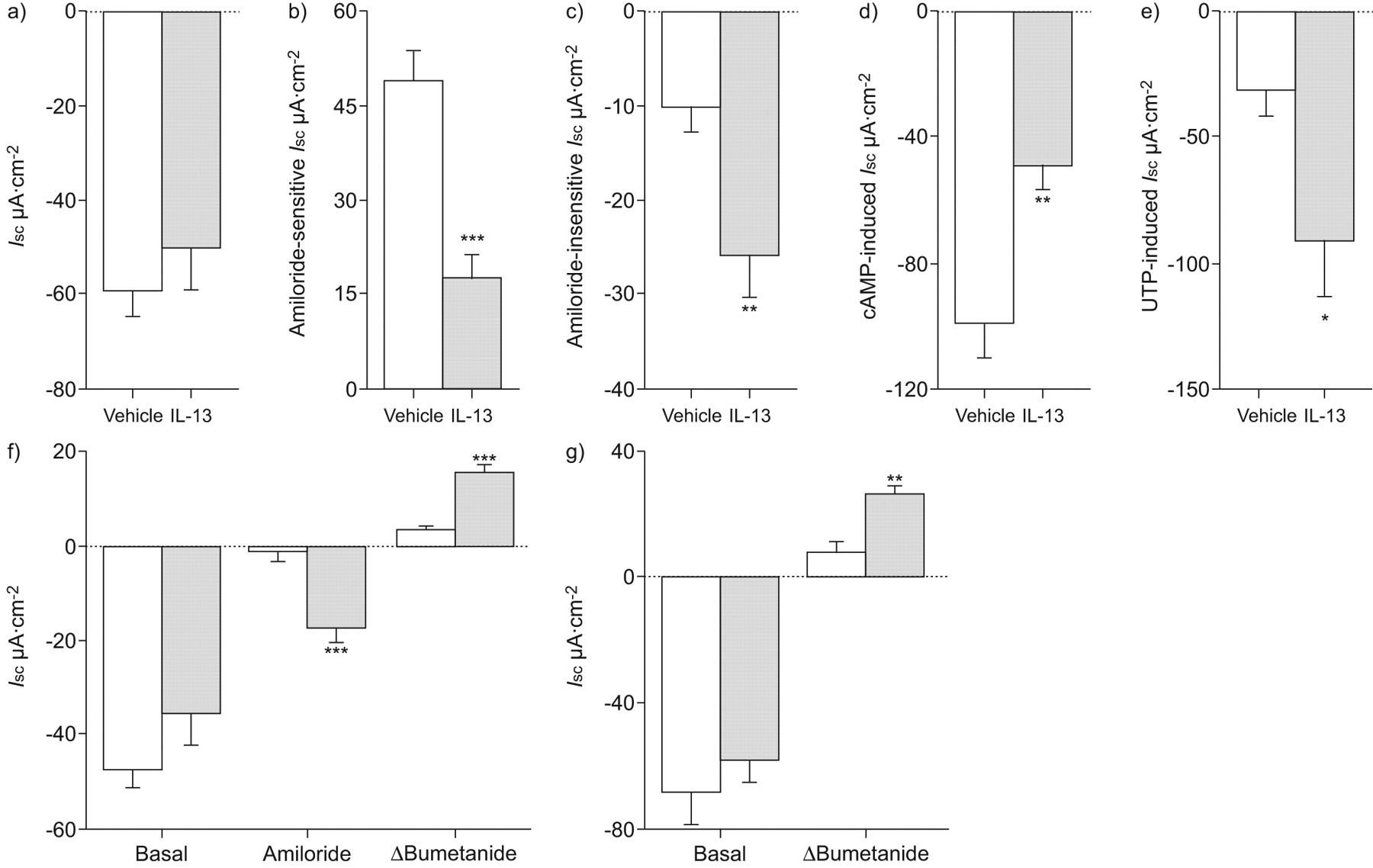
IL-13模仿allergen-induced炎症对气道离子传输的影响
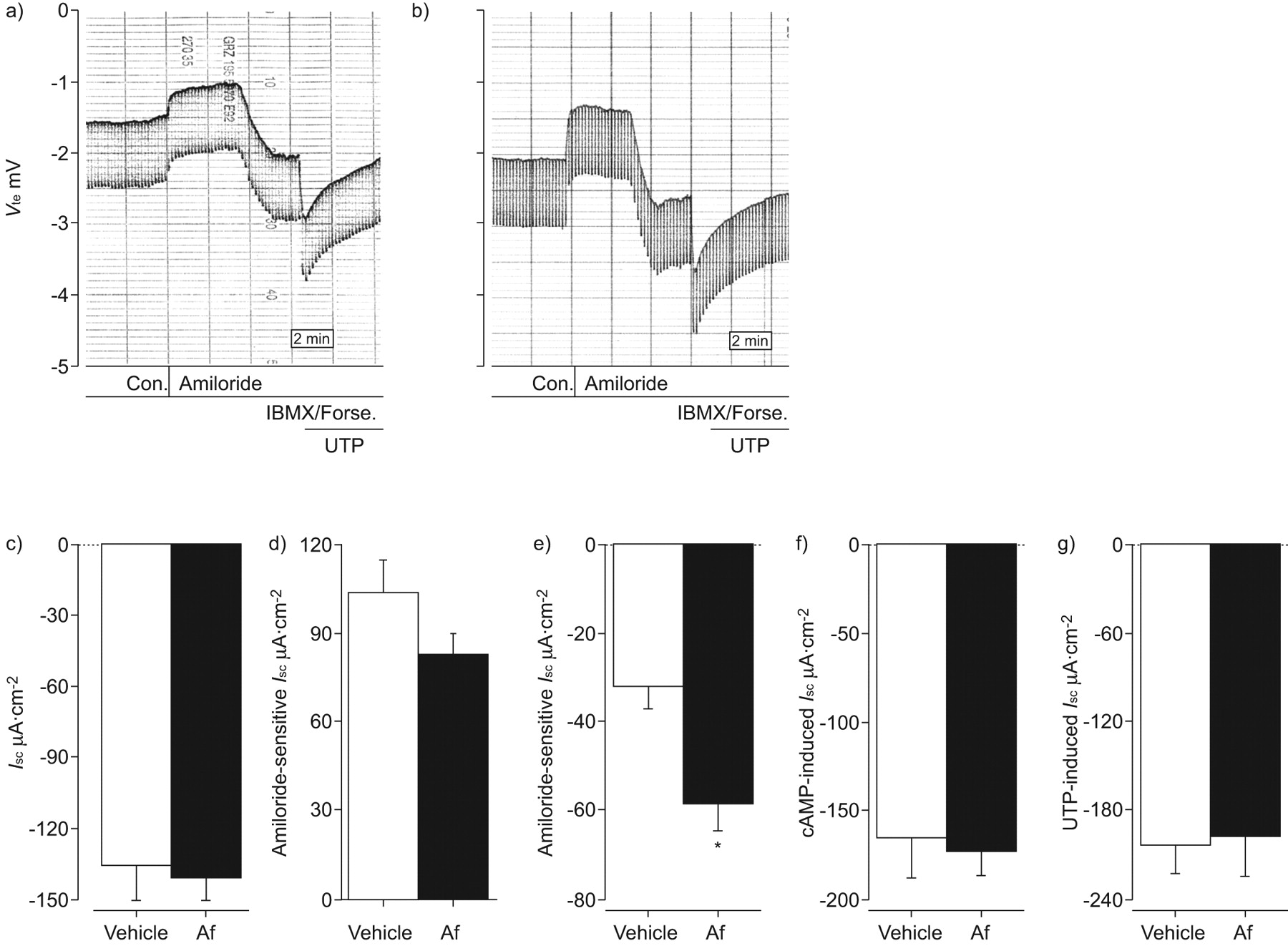
确定Th2细胞因子的作用更直接调制气道离子运输,我们对待WT小鼠气管内的IL-13滴剂,研究了影响生物属性刚切除气管组织。在支气管,类似于Af-induced过敏性炎症,IL-13没有影响我sc在基础条件下;然而,amiloride-sensitive我sc显著降低(p < 0.001)和剩余我sc在阿米洛利的存在显著增加(p < 0.01)与vehicle-treated老鼠相比,IL-13治疗(图6 a - c)。此外,影响IL-13 cAMP-induced我sc和UTP-dependent我sc(无花果6 d和e)也类似于Af-sensitisation (图4 gydF4y2Ba)。在气管,IL-13 amiloride-insensitive产生显著增加我sc(p < 0.05),观察到在房颤模型(数据没有显示)。这些结果表明,Th2细胞因子IL-13扮演着一个重要的角色在调制上皮离子运输复杂在活的有机体内小鼠过敏性气道炎症的发病机制。
白介素(IL) -13模仿allergen-induced炎症对气道离子传输的影响。成年野生型小鼠气管内的对待作品的IL-13(░)或单独车辆(□)和影响离子运输测定新鲜切除支气管上皮细胞。总结的)基底等效短路电流(我sc),b) amiloride-sensitive我scc) amiloride-insensitive我scd)环腺5′-monophospate(营)全身我sc和e)尿苷5′三磷酸(UTP)全身我sc在刚切除支气管IL-13 vehicle-treated老鼠。f, g) transepithelial Cl辅助2-mediated炎症的影响- - - - - -分泌测定通过添加布美他尼在基底条件和阿米洛利的存在。f) Amiloride-insensitive我sc和bumetanide-sensitive我sc(δBumetanide)在支气管从IL-13-treated阿米洛利的存在与vehicle-treated老鼠。g) Bumetanide-sensitive我sc在缺乏阿米洛利IL-13-treated的支气管与vehicle-treated老鼠。数据意味着±扫描电镜每组)(n = 5 - 8的老鼠。*:p < 0.05相比vehicle-treated老鼠;* *:p < 0.01相比vehicle-treated老鼠;* * *:与vehicle-treated老鼠相比p < 0.001。
Th2-mediated气道炎症促进Cl- - - - - -分泌
确定增加amiloride-insensitive的起源我sc观察到Th2-mediated气道炎症、气道组织灌注了布美他尼阻止transepithelial Cl- - - - - -分泌20.。在bumetanide-sensitive阿米洛利的存在,我sc在支气管显著增加(p < 0.001;图6 f)和气管(p < 0.05;数据未显示)组织从IL-13治疗小鼠相比vehicle-treated老鼠。因为阿米洛利hyperpolarises顶端膜电压和Cl可能因此产生驱动力- - - - - -分泌,不存在没有ENaC封锁,我们下个决心布美他尼在基底条件下的影响。在缺乏阿米洛利,布美他尼没有影响基底vehicle-treated小鼠气道离子运输,但抑制基底我sc在支气管显著(p < 0.01;图6克)和导管(p < 0.05;数据未显示)从IL-13治疗老鼠。值得注意的是,bumetanide-sensitive我sc在相同的范围没有和阿米洛利(图6 f和g)。这些结果表明,Th2-mediated炎症诱导基底Cl- - - - - -分泌,导致pro-secretory离子运输在过敏性呼吸道疾病表型。
讨论
本研究的结果1)证明,第一次Th2-mediated过敏小鼠气道炎症,引起粘膜敏化作用与房颤自然高空过敏症或气管内的Th2细胞因子IL-13滴剂,产生调制的气道上皮离子运输,除了嗜酸性粒细胞的炎症反应,杯状细胞化生和粘液分泌过多,和2)确定改变气道离子运输作为额外的异常复杂在活的有机体内过敏性呼吸道疾病的发病机理。
在气管,组织常用的气道离子运输研究老鼠,Af-induced气道炎症产生大量剩余的增加我sc阿米洛利块的存在,但没有影响ENaC-mediated Na+吸收或cAMP-dependent UTP-induced Cl- - - - - -分泌(图2)。在支气管过敏炎症反应是更复杂的图4 gydF4y2Ba)。类似于气管,amiloride-insensitive我sc增加在allergen-sensitised老鼠。此外,ENaC-mediated Na+吸收和cAMP-induced Cl- - - - - -分泌减少,而CaCC-mediated Cl- - - - - -在支气管分泌增加Af-sensitised老鼠相比,控制。气管内的滴注法生产的IL-13气道离子运输的变化相似,证明Th2-dependent过敏性炎症的影响(图6克ydF4y2Ba)。布美他尼、Na的拦截器+- k+2氯- - - - - -转运蛋白,抑制基底Cl- - - - - -吸收和transepithelial Cl- - - - - -运输、阿米洛利预处理组织证明amiloride-insensitive的增加我sc在Th2-mediated炎症反映Cl升高- - - - - -分泌(图6 f)20.。同样,bumetanide-sensitive我sc也没有阿米洛利的增加,表明Th2-mediated气道炎症增加Cl吗- - - - - -分泌基底条件下(图6克)。离子传输的调制引起的过敏性炎症在本机支气管组织很大程度上就像以前观察到的影响在人类支气管上皮细胞刺激后选择Th2细胞因子(il - 4和IL-13)在体外8,9。综上所述,这些研究结果表明,气道离子运输在allergen-driven积极监管、Th2-mediated气道免疫反应在活的有机体内。增加的净效应∼2-3-fold基底Cl- - - - - -气管和支气管分泌组织,一起相互调节气道Na+吸收和CaCC-mediated Cl- - - - - -在支气管分泌(无花果2,4和6)是一个pro-secretory离子运输表型,促进呼吸道表面的液体分泌和水化过敏性气道炎症。
适当的监管的美国手语体积坐标ENaC-mediated Na+吸收和Cl- - - - - -分泌,由雌性生殖道和中国商用飞机有限责任公司,发挥了至关重要的作用在维护正常的MCC,提供一个重要的肺部防御机制3,4。先前的研究患者的损失函数CFTR突变在钠或检测19,22- - - - - -24在小鼠研究显示,airway-specific超表达的βENaC(βENaC-Tg)10,25证明改变气道Na之间的平衡+吸收和Cl- - - - - -分泌产生深远的影响对美国手语体内平衡和黏膜纤毛的功能,并显示pro-secretory离子运输表型加速MCC,而hyperabsorptive表型引起黏膜纤毛的功能障碍和慢性呼吸道疾病。在这种背景下,我们目前的研究结果预测,pro-secretory离子运输表型的诱导Th2-driven炎症改善粘液清除粘液生产过剩的存在在过敏性气道疾病。支持这个假说比较肺表型的βENaC-Tg小鼠过敏性哮喘小鼠模型,以及最近的人体研究。首先,虽然气道粘液堵塞βENaC-Tg鼠标是一个不变的特性10,17,形成粘液栓没有观察到肺Af-sensitised老鼠(图1和数据未显示),或在其他哮喘小鼠模型5- - - - - -7,15,尽管存在显著的杯状细胞化生及粘液分泌过多。其次,人类最近的一项研究表明,MCC在轻度患者显著增加,稳定过敏性哮喘与健康对照组相比26。这些数据与假设是一致的协调增加流体和粘蛋白分泌Th2-driven引起的气道炎症可能会提供一个保护机制,促进粘液间隙,防止气道粘液堵塞在过敏性哮喘等呼吸道疾病。
相反,MCC在急性哮喘患者受损2,27。最近的一项研究表明,过量的血浆蛋白渗出物进入呼吸道的急性哮喘患者抑制protease-dependent黏蛋白退化和建议粘弹性性质的改变可能影响气道粘液间隙28。自分泌的粘蛋白大分子是“干”到气道表面,它们由绑定水化水从美国手语29日,我们推测,旺盛的黏液分泌可能超过Th2-driven离子运输补偿和导致相对美国手语的损耗,这可能会提供一个额外减少MCC在急性哮喘的机制。此外,我们推测,对提高液分泌能力降低过敏性炎症,如。通过基因和/或钠的功能变体,雌性生殖道或其他离子通道参与气道分泌液体,可能是迄今为止未被发现的危险因素的发展黏膜纤毛的功能障碍和严重气道粘液堵塞在致命的哮喘30.。
我们的研究也提供了新机械的见解气道离子传输的调制过敏性炎症。首先,通过比较Af-induced气道炎症的影响在支气管WT和Stat6生物属性- / -老鼠,我们证明pro-secretory表型的重要组件引起的过敏性炎症,即。ENaC-mediated Na+吸收和CaCC-mediated Cl- - - - - -分泌,取决于Stat6信号(图4 gydF4y2Ba),它也扮演着重要的角色在过敏性气道疾病的其他组件,包括嗜酸性粒细胞的炎症反应、粘液分泌过多和气道高反应性7,15,21。因为许多Stat6-dependent哮喘反应调节目标基因表达水平15,21因为以前在人类支气管上皮细胞培养的研究表明,刺激Th2细胞因子il - 4的影响的博亚基的表达,雌性生殖道和TMEM16a9,12过敏性气道炎症的影响相比,我们在记录这些离子通道的表达水平从WT和Stat6航空公司- / -老鼠。这些研究表明,抑制ENaC-mediated Na+吸收在支气管Af-sensitised WT与显著减少小鼠肺转录水平的所有三个博子单元(α,β和γ),这种效果是Stat6-dependent (图5)。最近的证据表明,翻译后调节如。博的蛋白水解分裂细胞外蛋白酶,控制Na中发挥着重要作用+从气道表面吸收31日。虽然我们没有地址转译后的监管可能过敏炎症的影响,我们的研究表明,Stat6-dependent转录调节钠可能提供一种机制,有助于抑制上皮Na+上皮的吸收和转换pro-secretory离子运输表型Th2-driven气道炎症。
相比之下,表达水平的雌性生殖道和TMEM16a没有改变在乙酰甲WT和Stat6- / -老鼠(图5)。鉴于TMEM16A最近被认定为中国商用飞机有限责任公司在人类支气管上皮细胞培养全球基因表达分析在不同搜索基因受il - 412,中国商用飞机有限责任公司功能的鼠标同系物在TMEM16a在最近的一项研究证实- / -老鼠13,令人惊奇的是那个Stat6-dependent UTP-induced Cl的老年病- - - - - -分泌allergen-sensitised老鼠不是TMEM16a与变化相关的表达式在我们的研究(图5)。这些结果表明,中国商用飞机有限责任公司与TMEM16a参与UTP-mediated Cl基因不同- - - - - -分泌。我们推测,缺乏TMEM16a表达的上调,尽管UTP-mediated Cl显著增加- - - - - -分泌引起的过敏性气道炎症的研究(无花果4和5),可能反映了物种之间的差异表达模式不同的中国商用飞机有限责任公司候选人在人类和小鼠气道上皮细胞。另外,调节细胞培养下TMEM16a表达式可能有所不同与在活的有机体内条件12,13。此外,我们表明,基底Cl的增加- - - - - -分泌构成pro-secretory离子传输的一个重要组成部分过敏性炎症引起的表型是Stat6-independent而不是与雌性生殖道或TMEM16a(转录水平的变化无花果4和5)。因此,我们的结果表明,未来的研究需要定义TMEM16a和其他Cl的相对的角色- - - - - -通道候选人在活的有机体内监管pro-secretory离子运输表型的过敏性气道疾病。
比较支气管的离子传输性质与导管在vehicle-treated WT老鼠(表1),以及不同反应allergen-induced炎症(无花果2和4),展示了深刻的差异调节气道离子运输在这两个地区的支气管树。有趣的是,地区差异的概念在小的离子和流体运输与大型航空公司的一项研究支持,而主要人类支气管和细支气管上皮细胞培养32。本机气管的地区差异与支气管上皮细胞可能有关:1)钠子单元和基因表达水平的差异导致基底和agonist-induced Cl- - - - - -分泌;2)当地的介质浓度的差异,如Th2细胞因子(il - 4和Il-13)、细胞外核苷酸和核苷(ATP和腺苷),释放在气道过敏性炎症和作为强有力的调节器离子运输8,9,33- - - - - -35;或3)当地受体密度的差异,尤其是常见的il - 4 / IL-13受体和/或上皮Stat6信号的差异。我们预测,进一步说明这些地区差异在基底的机制和监管气道离子运输可能为当地提供新的见解(来)的监管MCC在应对有害刺激气道不同区域的肺。
总之,我们的研究显示,Th2-mediated allergen-driven气道炎症触发pro-secretory离子运输表型在活的有机体内和表明,气道的积极调制离子运输是一个重要的效应功能复杂在活的有机体内过敏性呼吸道疾病的发病机理。此外,我们证明了这种反应的重要组成部分是由上皮Stat6信号。我们的研究结果表明,流体和黏液分泌增加协调Th2-driven气道炎症可能会提供一个保护机制促进粘液间隙,防止气道粘液堵塞的粘液生产过剩在过敏性呼吸道疾病,并建议,这种上皮反应失调可能提供一个线索严重气道粘液插入致命哮喘。
确认
我们感谢周z(儿科肺学分工和Auergy和囊性纤维化中心,儿科部门三世,海德堡大学海德堡,德国)专家援助与过敏原暴露和有益的讨论。
脚注
支持声明
本研究支持的德意志Forschungsgemeinschaft (DFG马2081/3-2和马2081/4-1)和Ernest-Solvay-Stiftung。p . Anagnostopoulou接到研究生学院的短期奖学金海德堡(德国海德堡)。
感兴趣的语句
没有宣布。
- 收到了2009年11月13日。
- 接受2010年4月12日。
- ©2010人队











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}