





文摘
肺上皮细胞促进伤口闭合的分泌各种细胞因子和生长因子。神经生长因子(神经生长因子)气道炎症中描述;然而,它在肺修复中可能扮演的角色还没有检查。
探讨修复神经生长因子的函数,实验进行在体外使用培养的肺泡上皮细胞和在活的有机体内使用naphthalene-induced克拉拉上皮细胞损伤模型。
这两个在体外和在活的有机体内实验显示气道上皮细胞增殖后损伤是依赖于神经生长因子及其受体的表达,tropomyosin-receptor-kinase a .此外,神经生长因子也增强在体外肺泡II型细胞的迁移。在活的有机体内克拉拉细胞中过度表达,转基因小鼠神经生长因子(NGFtg)并没有透露任何增殖或克拉拉细胞表型的改变。然而,克拉拉细胞特定的伤害后,扩散增加NGFtg和受损的神经生长因子的抑制作用。此外,神经生长因子还促进了我胶原蛋白、纤粘连蛋白的表达在体外和在活的有机体内维修期间,明显高于水平测量re-epithelialising NGFtg老鼠。
我们的研究表明,神经生长因子促进肺上皮细胞的增殖在体外肺损伤后,克拉拉细胞的更新在活的有机体内。
成熟健康的气道是一个复杂的,然而脆弱的结构不断暴露吸入环境代理。由于这种紧密联系与外部环境肺很容易受到伤害;因此,损伤和上皮脱落通常描述事件呼吸道感染、哮喘、慢性支气管炎和间质性肺病1。肺有非凡的能力,促进修复通过使用专门的隔间与几个明显的功能细胞。这些细胞类型参与一系列重叠事件恢复呼吸道上皮细胞。在肺的肺泡间隔,肺泡上皮II型细胞(AECII)的假定的祖细胞是肺泡上皮和re-epithelialisation发挥重要作用的肺损伤后肺泡壁2,3。细胞类型中衬气道,nonciliated细支气管细胞(克拉拉细胞)构成主要的上皮细胞类型的许多物种,包括老鼠。克拉拉细胞作为高脚杯的祖细胞和纤毛上皮细胞是主要贡献者气道修复过程4,5。在人类中,克拉拉大大有助于支气管上皮细胞的增殖和细胞已被描述为一个重要的细胞群的维护正常上皮尤其是在远端进行航空公司6。
根据伤口的大小和上皮剥蚀的程度,损坏气道上皮细胞的更新包括迁移和扩散的残余相邻的上皮细胞,未受损区域7。表达的细胞外基质(ECM)临时矩阵中蛋白质增加,和克拉拉细胞纤连蛋白和胶原蛋白等因素我已经描述的吸引和为移民提供坚定点8。
克拉拉细胞特别容易受到损伤是由于其能力将化学物质如萘(NA)转换成有毒的中间体。钠是一种多环芳烃在香烟烟雾和柴油尾气和能够诱导高剂量依赖性和细胞类型,site-selective毒性9,10。NA的毒性是由于这种化合物的代谢细胞色素P450单氧酶有毒中间导致克拉拉细胞肿胀,vacuolisation,剥落的腔气管损伤后24小时开始11。NA在这项研究的使用使我们调查的克拉拉细胞特定的损伤后修复过程在活的有机体内。
受伤后,介质驱动气道上皮修复可能来自许多来源。除了因素释放激活当地的免疫细胞12,上皮细胞也可能自分泌和/或旁分泌生长因子的来源。就是这样一个中介积极气道上皮细胞分泌的神经生长因子(神经生长因子),一位杰出的生成生长因子家族的成员最初描述促进神经元生长和存活13。最近,神经生长因子的重要性和其高亲和力受体(TrkA)被描述的tropomyosin-receptor-kinase炎症与气道功能障碍有关14,15。这样的例子包括神经生长因子表达的激活支气管过敏14、鼻15气道上皮细胞以及上皮神经生长因子升高中介嗜酸性粒细胞浸润16和生存17。神经生长因子在其他组织的上皮细胞也表达了,特别是在激活通过炎症或在再生条件18- - - - - -21。尽管有一些报告显示神经生长因子在伤口愈合的作用22- - - - - -24,其参与肺上皮修复过程迄今为止还没有被调查。
本研究的目的是研究神经生长因子的作用在受伤的气道上皮细胞的修复,我们的研究结果突出的贡献对克拉拉气道上皮细胞的增殖后的肺损伤。
方法
老鼠
6-8-week-old女性C57BL / 6小鼠获得哈伦温克尔曼(Borchen,德国)。与转基因老鼠overexpressing神经生长因子的控制下lung-specific Clara-cell分泌蛋白启动子(NGFtg)25。据“所有动物实验进行实验动物保健和使用指南”的准备和实验动物学会发表的科学26。
伤肺上皮细胞的分析
LA4小鼠肺肺泡上皮细胞(美式文化收藏,罗克维尔市,在美国)播种1×106细胞·好−1和补充F12K营养混合物含有15% FCS(美国Gibco,卡尔斯巴德,CA)培养直到融合到12-well盘子。一夜之间(o / n)血清饥饿后,层从上到下都挠使用200μL吸管提示然后孵化与介质,介质含重组人神经生长因子(β-NGF 100 ng·毫升−1;研发系统,明尼阿波利斯,美国),多克隆抗体阻断神经生长因子(anti-NGF 625 ng·毫升−1;美国新泽西Peprotech,落基山),或免疫球蛋白G (Ig)控制(625 ng·毫升−1;σ,圣路易斯,密苏里州,美国)。所有组中含有1% FCS孵化。媒介与随行补充剂改变每24 h。伤口大小是由光学显微镜以μm受伤后(奥林巴斯,汉堡,德国),每24 h使用细胞ˆ72 h F成像软件程序(软成像系统GmbH,明斯特,德国)。至少3 - 4伤口六LA4细胞层进行了分析。
培养的肺上皮细胞的扩散
主要AECII分离根据协议提供的补充材料。1×105AECII或LA4细胞·−1生长在96孔板在一式三份。AECII种植在介质含有10% FCS其次是介质包含0.1,1或10% FCS 24、48和72 h。AECII扩散细胞进一步评估0.1% FCS包含介质、神经生长因子、anti-NGF或控制上述72 h的免疫球蛋白浓度试验。LA4细胞,20 pmol·μL−1神经生长因子、TrkA或控制核(试剂盒、希尔登,德国)与细胞孵化根据制造商的指示。细胞增殖检测进行使用BrdU (5-bromo-2′-deoxy-uridine)标签设备根据制造商的指示(罗氏,曼海姆,德国)。进一步的细节,包括引物序列,见补充材料。
在体外使用AECII迁移分析
AECII化验了NGF-induced迁移使用24-well修改Boyden室(猎鹰;BD,富兰克林湖,新泽西,美国)使用transwell膜(8-μm孔隙大小)。参议院包含105500年AECIIμL中型和众议院含有500μL中含有游离神经生长因子(100 ng·毫升−1)。细胞培养在37°C 6 h公司为5%2后插入被移除和细胞在低表面与Diff-Quick复染色(戴德,贝林,德国)5分钟。平均5×100字段插入是计算每组5样品使用光学显微镜(奥林巴斯,汉堡,德国)。迁移是表示为迁移细胞/×100场。
在活的有机体内克拉拉细胞损伤模型,并使用NA剥蚀
野生型(WT)或NGFtg控制小鼠腹腔内(i.p。)玉米油(西格玛奥德里奇,慕尼黑,德国)、WT或NGFtg小鼠接受克拉拉细胞剥离得到我。p NA (250 mg·公斤−1德国慕尼黑,西格玛奥德里奇)溶解在玉米油在第0天(图S1a辅料和b)。附加组、WT控制老鼠和WT小鼠接受NA还收到了鼻内anti-NGF (anti-NGF 2.5 s、N 6655;Sigma-Aldrich)或鼻内控制免疫球蛋白(Sigma-Aldrich)在一天的24μg浓度0、3、6和9(图S1a辅料和b)。一套新的老鼠还收到了饮用水中BrdU 1毫克·毫升的浓度−1(水含有1%葡萄糖)从0到每天10(图S1a补充材料)。治疗后1天,10 - 29,支气管肺泡灌洗(BAL)与800年进行了两次μL冰冷的PBS加蛋白酶抑制剂混合(罗斯,卡尔斯鲁厄,德国)和全肺组织学被移除,信使rna和蛋白质分析。
神经生长因子ELISA
神经生长因子ELISA (WI Promega,麦迪逊,美国)是按照制造商的指示执行细胞自由文化上层清液制作六LA4细胞层(有或没有伤口在6、12、24、48和72 h)和平衡液(BALF) WT和NGFtg动物(控制和NA治疗10天,29)。盘子在微型板块阅读autoreader (Tecan、萨尔茨堡、奥地利)在405海里。
免疫印迹分析
总蛋白提取LA4细胞单层膜,AECII和肺组织WT NGFtg控制和NA治疗小鼠使用m /哺乳动物蛋白质提取工具包(热科学、皮尔斯、波恩、德国)。免疫印迹(如前所述)27与神经生长因子、TrkA(1:50 0,圣克鲁斯海德堡,德国),anti-β-actin单克隆抗体(1:40,000;σ)和二级peroxidise-conjugated抗体(1:5000;σ)。免疫反应性的蛋白质乐队与化学发光底物(可视化SuperSignal;热科学,皮尔斯)。乐队与柯达的图像扫描站CF 450和3.5使用柯达1 d成像分析软件。看到更多细节的补充材料。
RNA隔离、实时PCR和分析
5×105AECII·毫升−1被孵化24-well板anti-NGF (625 ng·毫升−1ng),控制免疫球蛋白(625毫升−1ng),或重组人神经生长因子(100毫升−1)24 h。总RNA隔绝AECII或肺组织使用RNeasy迷你包(试剂盒、希尔登,德国)。定量PCR进行使用QuantiTect SYBR®绿色PCR试剂盒(试剂盒)。特定的引物胶原蛋白I,纤连蛋白用于实时PCR。信使rna复制的数字比较,表示相对于鼠标核糖体L32看家基因。看到更多细节的补充材料。
免疫组织化学
免疫组织化学进行肺切片石蜡包埋组织如前所述28。主要抗体浓度;anti-Clara细胞特定蛋白(CCSP)(稀释1:3,000;由j·克鲁格是Justis-Liebig-Universitat、吉森、德国);anti-fibronectin(稀释1:1,000;Biozol、决定、德国),anti-collagen我(1:50 0稀释;Biozol)、羊anti-BrdU(1:10稀释;Biozol)。二级生物素化的抗体(美国向量实验室,伯林盖姆,CA)稀释1:400。部分分析了使用光学显微镜(奥林巴斯欧罗巴GmbH,汉堡,德国)。
克拉拉细胞和BrdU-incorporating增殖细胞的形态学分析
老鼠肺部固定气管滴注法用6%磷酸缓冲多聚甲醛在20厘米液柱的压力。肺容积是由流体位移,系统性的肺组织被均匀随机抽样和根据标准处理方法29日。CCSP——和BrdU-positive细胞形态学分析的是执行先前描述的方法的使用修改后的版本30.。近端和远端航空公司被蒙蔽的方式计算。CCSP-positive细胞含核概要文件和BrdU-positive核资料统计。看到在线补充更多细节。
统计分析
统计学意义的正态分布样本被使用一个未配对t检验分析。对比以上两组是由方差分析的一种方式。的假定值< 0.05被认为是具有统计学意义。所有数值数据表示为±扫描电镜。计算进行了使用图垫棱镜软件,版本3.02。
结果
神经生长因子的生产和TrkA表达式是增加受伤的肺上皮细胞在体外
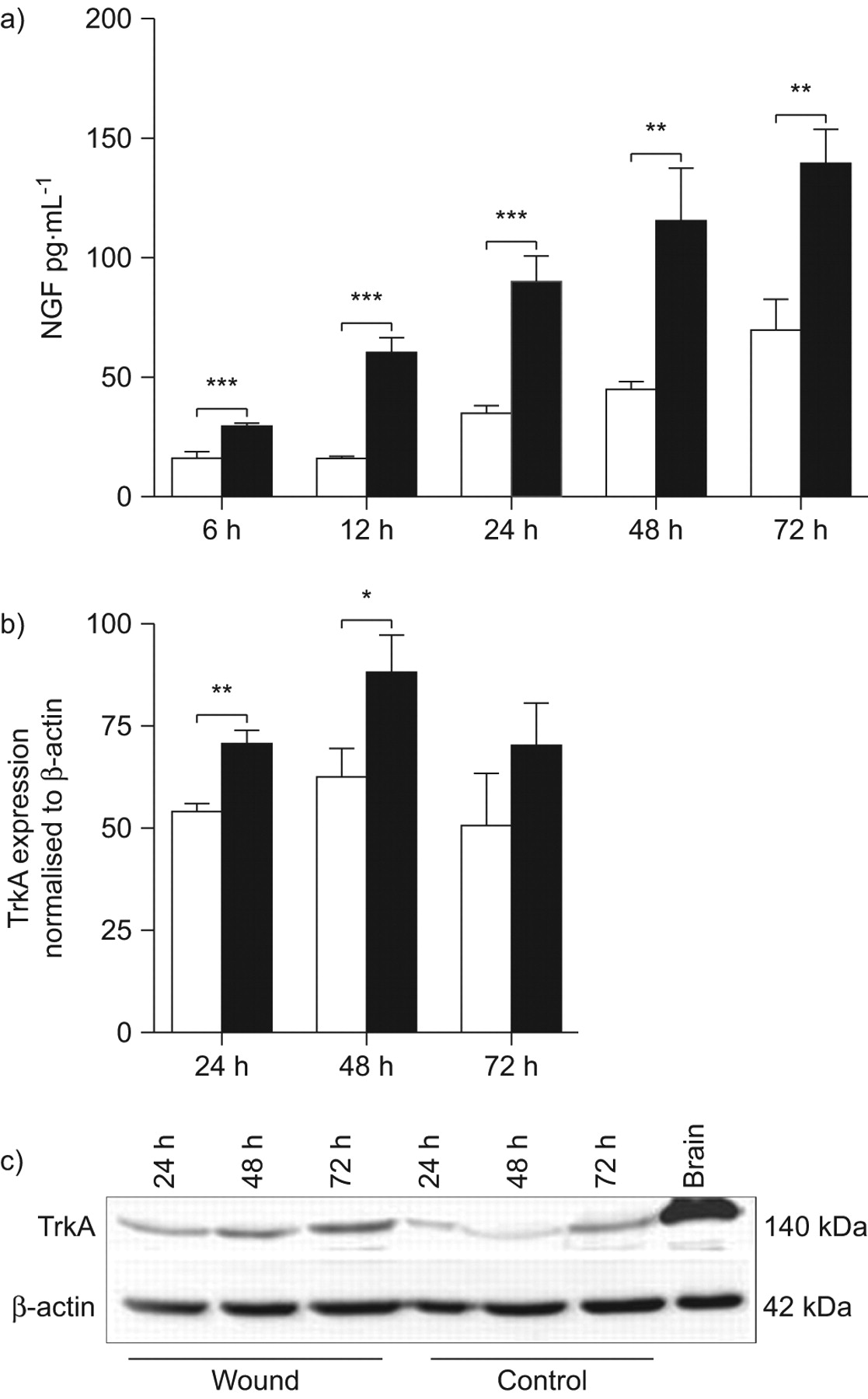
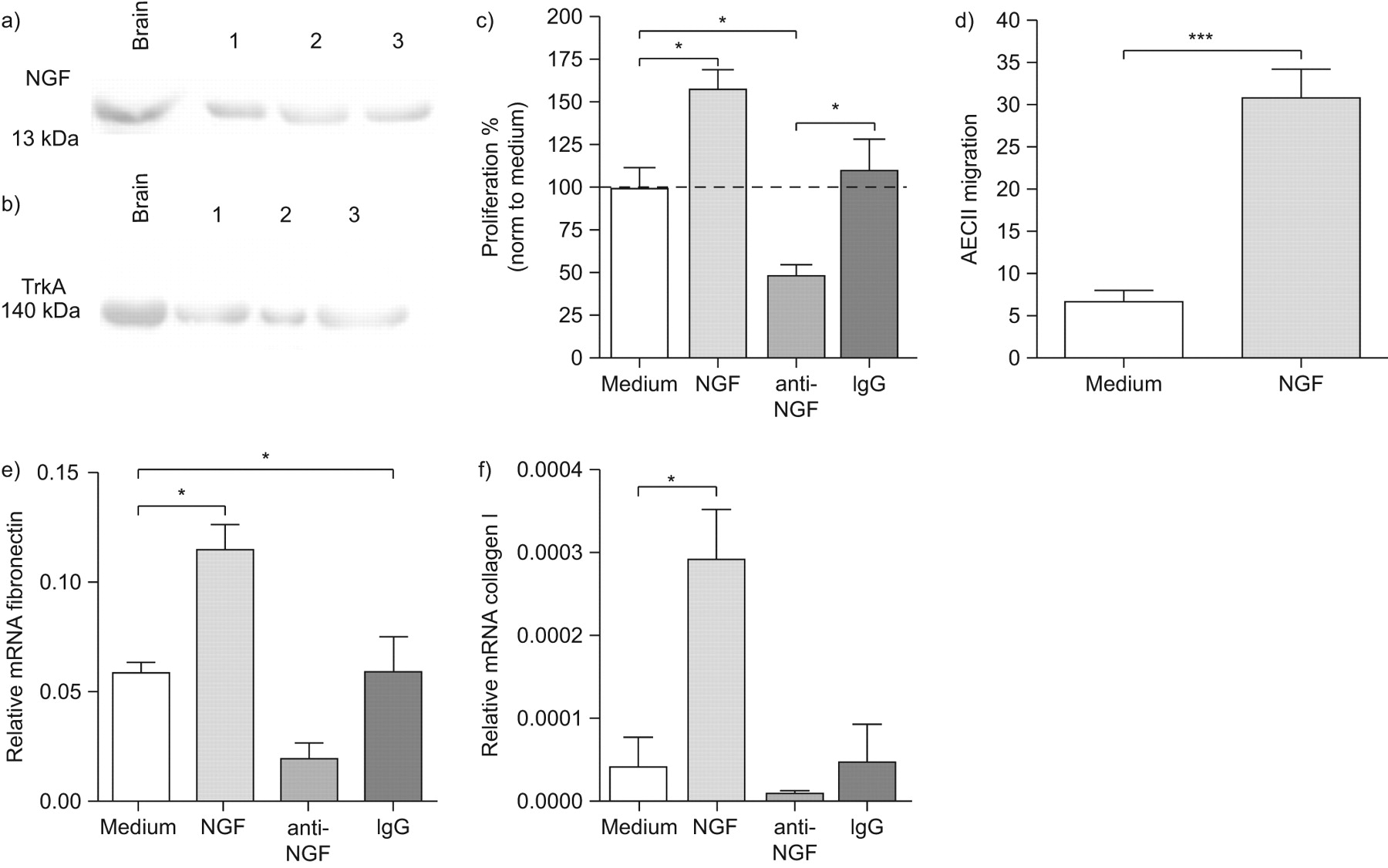
神经生长因子的表达在肺上皮细胞第一次检查在体外伤口使用小鼠LA4细胞分析模型。LA4细胞汇合的单层膜受伤导致增加神经生长因子及其受体的表达,TrkA (图1)。神经生长因子在受伤的上皮细胞蛋白质含量明显高于与nonwounded LA4细胞在所有时间点测量(图1一个)。逐渐增加直到72年观察伤后神经生长因子h,与观察到的最早6 h。神经生长因子的生产还在nonwounded细胞随着时间的增加,与正常细胞增殖相关。相应地,TrkA调节在24和48 h后受伤,出现在量要明显高于nonwounded膜图1 b)。TrkA表达的增加伴随着增加的上层清液中神经生长因子(图1一个)。这些结果表明,自分泌神经生长因子和TrkA诱导肺上皮细胞损伤。
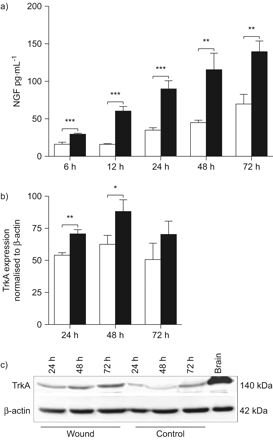
神经生长因子的生产(神经生长因子)和TrkA的表情在体外后LA4细胞单层受伤。)神经生长因子的生产是由ELISA测定在完好无损的控制(□),和受伤(▪)LA4细胞在6、12、24、48和72 h . b)的表达了TrkA量化强度乐队正常化的西方墨点法β-actin控制(□)和LA4单层膜与伤口(▪)24日,48和72 h . c)代表污点的显示表达TrkA (140 kDa)和β-actin (42 kDa)在受伤和控制单层LA4 24日,48和72 h (b)的实验一样。数据提出了均值±sem (n = 6)在每个时间点。*:p < 0.05;* *:p < 0.01;* * *;p < 0.001。
神经生长因子促进伤口关闭和上皮细胞的增殖在体外
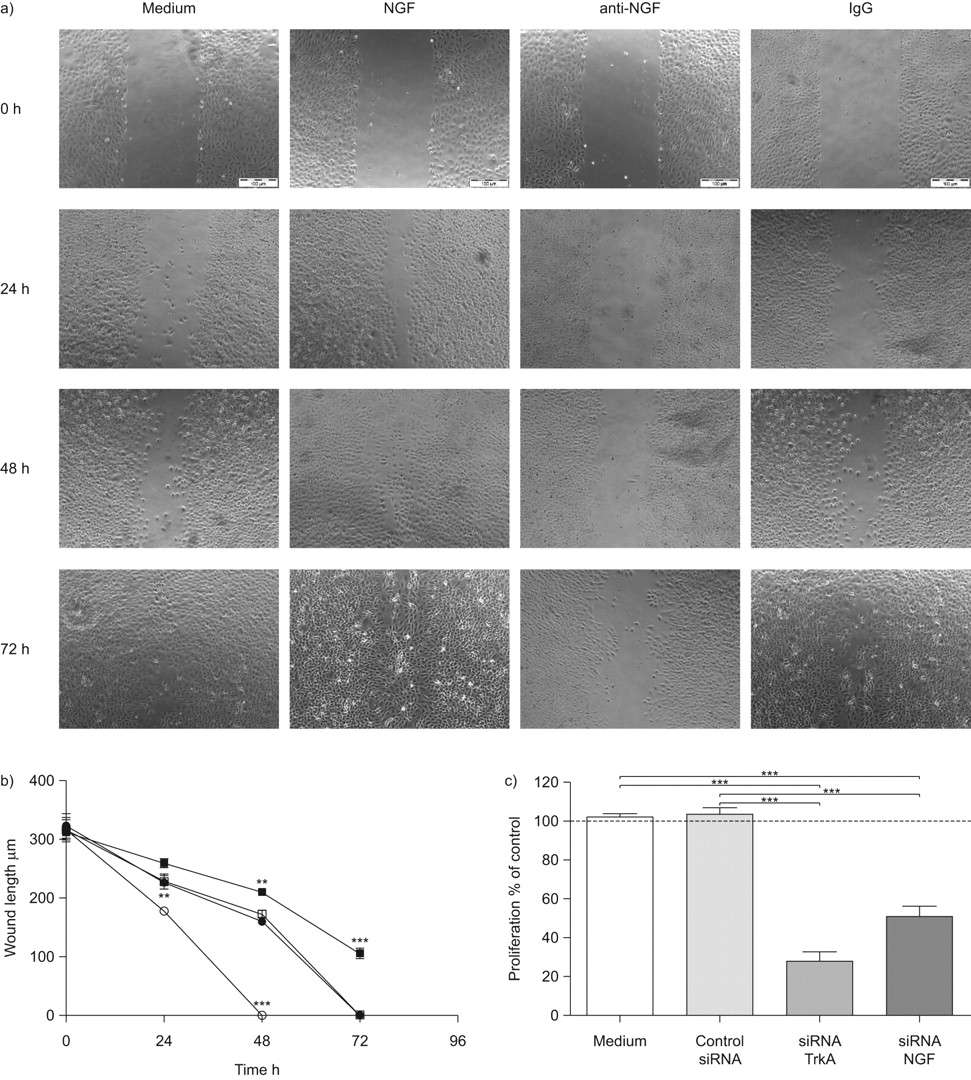
来确定观察到upregulation神经生长因子及其受体的影响关闭伤口,受伤的层的LA4细胞孵化与阻塞神经生长因子抗体(anti-NGF)或外源性神经生长因子。抑制神经生长因子显著延迟关闭伤口,而外源性神经生长因子蛋白显著加速伤口关闭(图2一个受伤和b)。控制层与控制免疫球蛋白抗体或孵化中只有伤口相似的大小(图2一个和b)。在控制层,伤口关闭实现在72 h,而伤口未能关闭anti-NGF治疗组即使在72 h, (图2一个)。相比之下,伤口孵化与神经生长因子在早些时候关闭48 h。因为据报道,神经生长因子的增殖效应是由它的高亲和性受体TrkA,神经生长因子的影响和TrkA肺上皮细胞增殖的规定也被调查在体外。核抑制神经生长因子或TrkA撞倒了信使rna和蛋白质含量显著降低水平相比介质控制(图S2补充材料)。TrkA和神经生长因子的抑制这些siRNAs显著降低LA4上皮细胞的增殖∼75和50%的媒介控制,分别是(图2 c)。这些结果表明,神经生长因子和TrkA重要伤肺上皮细胞的增殖和修复在体外。
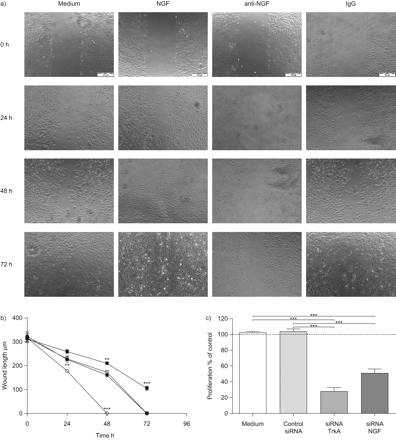
神经生长因子(神经生长因子)调节伤口关闭LA4细胞在体外。a、b)融合性的层的LA4细胞受伤,伤口关闭监控在单层膜的动力学只生长在介质(介质;□),介质与重组神经生长因子(NGF对于○),免疫球蛋白(Ig) G控制(免疫球蛋白、⨐),或与anti-NGF (anti-NGF▪) 0, 24、48和72 h。伤口封被量化测量伤口长度在μm伤口。统计比较NGF-stimulated和anti-NGF-treated组中只有组。c)细胞增殖是由测量的BrdU (5-bromo-2′-deoxy-uridine)增殖LA4细胞的DNA。扩散的速度是相对于基底细胞增殖在中控制BrdU合并为100% (·····)。TrkA(▓)和神经生长因子(┬)相比,被特定的核和细胞生长在转染试验介质(□)和非特异性核(┐)。数据意味着±扫描电镜每个时间点(n = 6)。* * *:p < 0.001。比例尺条= 100μm面板。
神经生长因子介导细胞增殖、迁移和ECM AECII表达式
确定神经生长因子的重要作用后肺上皮细胞系,我们很感兴趣对神经生长因子的影响和TrkA主要AECII。神经生长因子和TrkA被发现被表达在初级AECII (图3和b)。为了测试以初级AECII细胞增殖神经生长因子的作用,我们建立了一个在体外化验。AECII扩散的时间和剂量反应在体外24、48和72 h在0.1%,1%和10% FCS浓度(图S3c补充材料)显示72 h作为检查的最佳时间点扩散。神经生长因子被认为在0.1% FCS浓度显著影响扩散。符合我们之前在体外结果使用LA4细胞,神经生长因子增加AECII的增殖能力,而anti-NGF显著下调扩散与介质和免疫球蛋白控制(图3 c)。我们还在协调迁移研究神经生长因子的作用,这是一个伤口修复的关键事件。神经生长因子被认为强烈诱导迁移AECII在孵化与外源性神经生长因子(图3 d)。此外,ECM的表达因素,如胶原蛋白我和纤连蛋白也被调查在体外在原代细胞培养AECII。纤连蛋白和胶原蛋白的mRNA表达我是显著增加在孵化与外源性神经生长因子(图3 e和f)和神经生长因子的抑制导致纤连蛋白mRNA表达明显降低(图3 e)。这些结果暗示神经生长因子AECII积极促进增殖和迁移,这是伤口修复的关键条件。
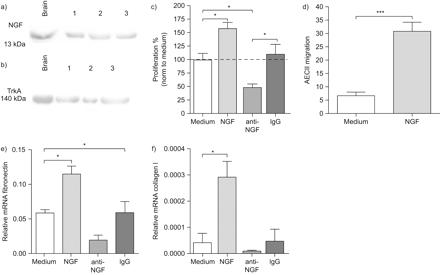
神经生长因子(神经生长因子)调节增殖,迁移和肺泡上皮的细胞外基质mRNA表达II型细胞(AECII)。)神经生长因子和b) TrkA AECII蛋白表达。c) AECII扩散分析72 h后孵化只中等或中等含神经生长因子,anti-NGF或控制免疫球蛋白G (Ig)。d) AECII迁移分析6 h后孵化与介质或重组神经生长因子。迁移是表示为迁移细胞/×100的视野。相对mRNA水平e) AECII细胞纤连蛋白或胶原蛋白f)我孵化24 h只中等或中等含神经生长因子,anti-NGF或控制免疫球蛋白。数据意味着±扫描电镜(n = 6)目标基因拷贝数每册L32(相对mRNA)为每个组。*:p < 0.05;* * *:p < 0.001。
神经生长因子和TrkA增加在气道上皮细胞的修复在活的有机体内
检查神经生长因子在气道上皮修复的作用在活的有机体内,我们采用钠诱导的克拉拉在肺上皮细胞损伤模型。神经生长因子的表达和TrkA决心和29天10点伤害与NA BAL流体和WT和NGFtg动物肺总蛋白(图4)。在未经处理的NGFtg控制老鼠,基线水平的神经生长因子明显高于由于本构下神经生长因子的表达相比,克拉拉细胞specific-promoter WT控制动物(图4)。与NA受伤后,WT动物显示神经生长因子的进步upregulation一直持续到29日(图4)。相反,神经生长因子水平NGFtg动物拒绝在所有时间点比未经处理的钠治疗后NGFtg动物由于钠诱导的神经生长因子过度表达克拉拉细胞损失。然而,神经生长因子在NA NGFtg对待动物的数量相比,在各个时间点上均明显高于NA WT对待动物。神经生长因子的表达在克拉拉细胞没有影响的克拉拉细胞总数的航空公司(数据没有显示)和克拉拉细胞的剥蚀后24 h在NA NA损伤被发现类似治疗WT (WT-NA)和NGFtg (NGFtg-NA)动物(图S4A补充材料)。此外,脱落细胞的数量量化BALF的两组老鼠相当(图S4b补充材料)。
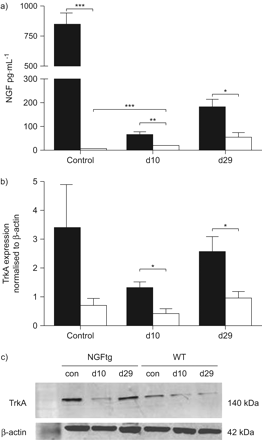
神经生长因子的表达和TrkA(神经生长因子)在活的有机体内克拉拉细胞肺损伤。NGFtg)神经生长因子水平(▪)和野生型(□)动物没有萘(NA)治疗(控制)和钠治疗后第十天,29 (d10, d29)。神经生长因子表达的ELISA测定支气管肺泡灌洗。b) TrkA量化,表示为乐队的表达强度正常化看家基因,β-actin。c)代表污点的显示表达TrkA (140 kDa)和β-actin (40 kDa)控制NGFtg和野生型(WT)动物(控制)和钠治疗后第十天,29 (d10 d29)从实验图如下所示。数据意味着±扫描电镜为每个组(n = 6)的老鼠。* *:*:p < 0.05, p < 0.01, * * *: p < 0.001。
调查的角色TrkA下游目标的神经生长因子修复过程中,其在肺组织的水平量化与此同时。TrkA被发现显著调节10天,29克拉拉细胞损伤后NGFtg动物与WT动物(图4 b)。
这些结果表明,神经生长因子及其受体、TrkA,是调节气道上皮损伤和他们的感应是正在进行的修复过程中维护的。
神经生长因子促进克拉拉细胞的更新
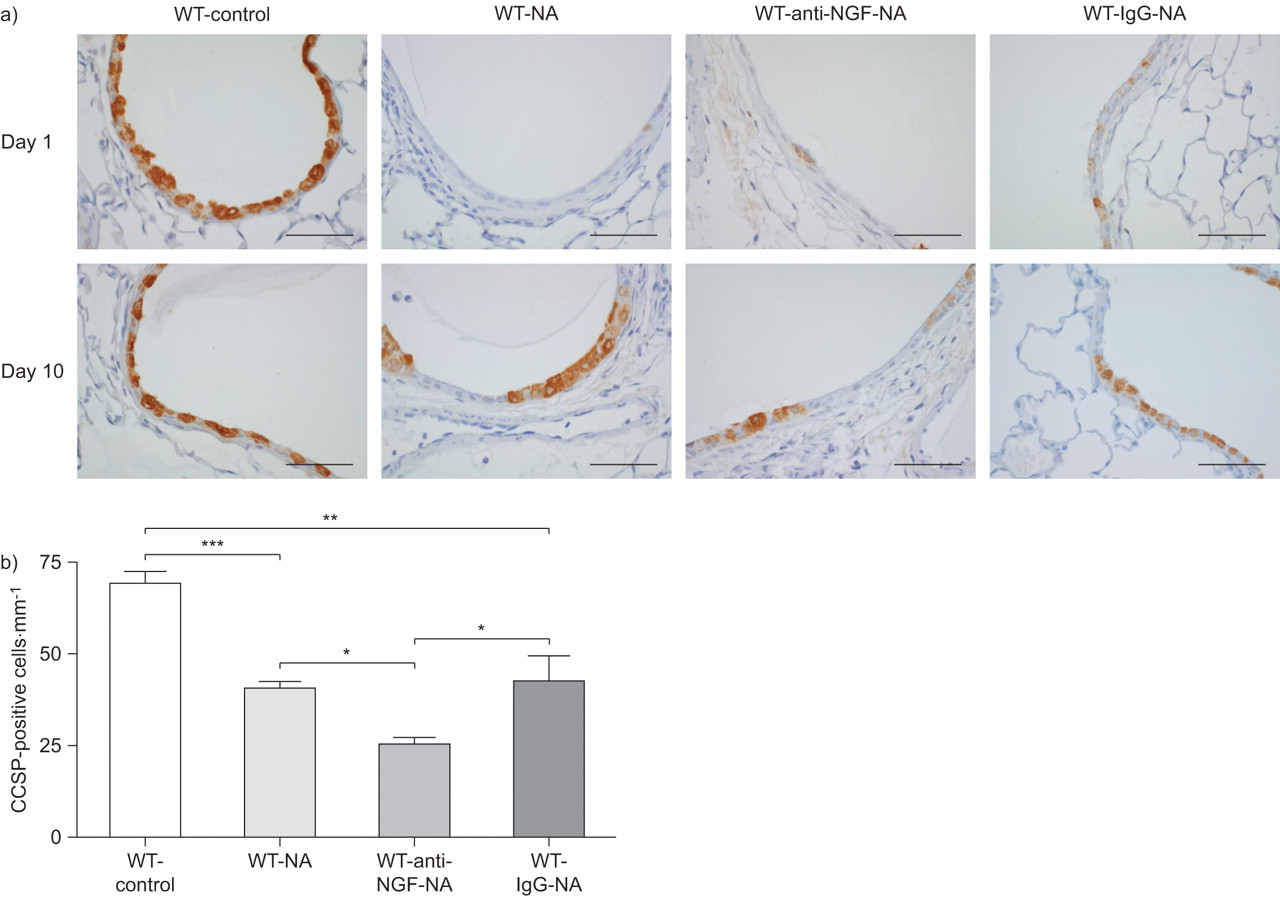
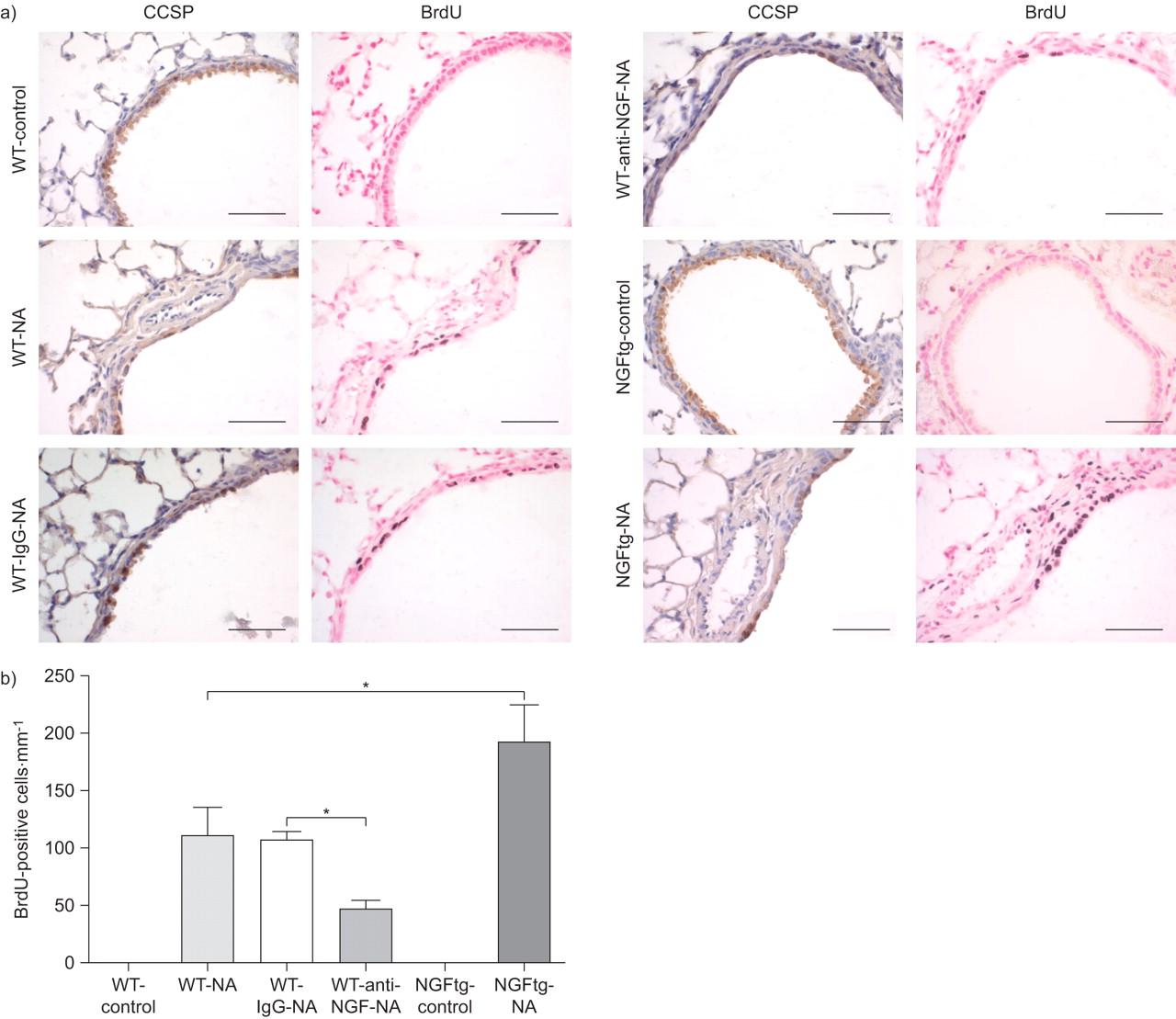
我们下一个测试神经生长因子的作用在克拉拉细胞更新的伤害。克拉拉细胞组成的主要人口衬里上皮细胞小鼠支气管气道。钠治疗后,克拉拉细胞被剥蚀白天1 (图5)。在第十天,克拉拉细胞周围的航空公司已经开始再现航空公司在所有团体治疗NA,量化的CCSP染色(图5 b)。老鼠和NA NA和控制免疫球蛋白治疗显示出类似的克拉拉细胞数量在第十天,与气道的相当大的部分已经由克拉拉细胞排列。相比之下,动物接受NA和中和anti-NGF抗体,显著降低了项CCSP-positive细胞气道在第十天,再现只偶尔在气道上皮细胞(图5)。这些结果表明,神经生长因子促进克拉拉细胞更新在活的有机体内在NA-induced受伤。
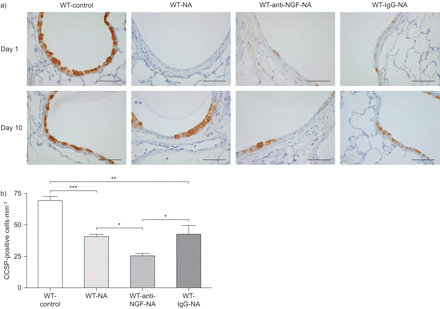
克拉拉细胞更新。玉米油)完整的克拉拉细胞治疗野生动物(WT)控制在每天1和10 (WT-control)和萘(NA)治疗后(WT-NA) anti-nerve生长因子(神经生长因子)(WT-anti-NGF-NA)或控制免疫球蛋白(Ig) G (WT-IgG-NA)抗体。克拉拉细胞被染色的克拉拉细胞分泌蛋白(CCSP)褐色nonciliated柱状细胞对蓝色计数器污渍。酒吧= 50μm规模。b)量化每毫米CCSP-positive细胞的基底膜钠治疗后10天。数据意味着±扫描电镜为每个组(n = 6)的老鼠。* *:*:p < 0.05, p < 0.01, * * *: p < 0.001。
神经生长因子诱导胶原蛋白我和纤连蛋白mRNA表达在NA诱导克拉拉细胞损伤
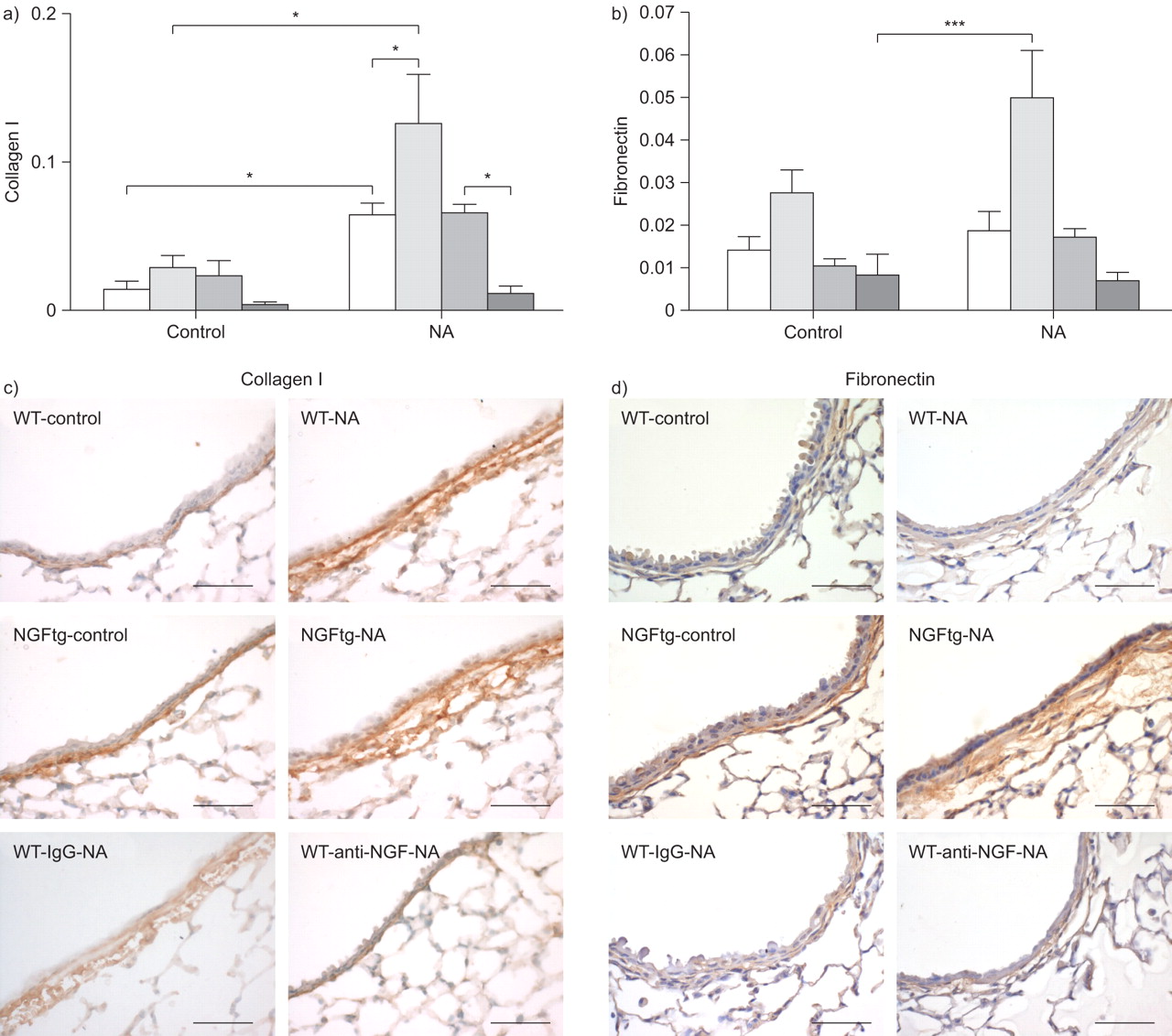
细胞迁移和增殖极度依赖于胶细胞之间的相互作用和基础矩阵。同时,考虑到克拉拉细胞显示优惠我坚持胶原蛋白和纤连蛋白,我们利用NA模型研究神经生长因子的影响在这些基质分子的表达(图6)。事实上,胶原蛋白我和纤连蛋白mRNA水平显著增加在NGFtg动物肺匀浆后第十天NA暴露比未经处理的NGFtg控制动物(图6和b)。此外,anti-NGF减少胶原蛋白的表达的应用我在NA WT对待动物(图6)。免疫组织化学染色证实我胶原蛋白、纤粘连蛋白qPCR数据和更强烈的胶原蛋白我和纤连蛋白染色观察潜在的上皮细胞层控制和NA NGFtg对待动物与各自的控制(图6 c和d)。应用anti-NGF显示相对降低了胶原染色我NA-treated WT动物。这些结果表明,神经生长因子积极增加ECM因素我胶原蛋白、纤粘连蛋白的表达在修复过程中肺。
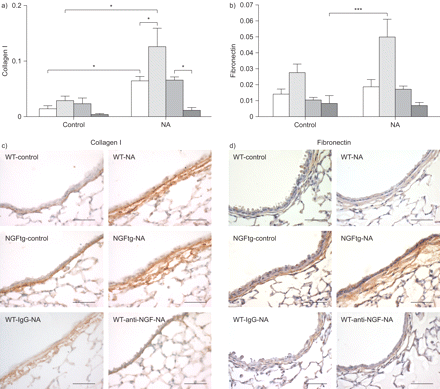
调制我胶原蛋白、纤粘连蛋白的表达在神经生长因子(神经生长因子)。相对的mRNA水平)我和b)纤连蛋白胶原蛋白。数据表示为目标基因的拷贝数每册L32 mRNA(相对)。信使rna分析野生型(WT;□),NGFtg (┐)、WT动物治疗免疫球蛋白(Ig) G(▓)或anti-NGF (┬)。c)胶原蛋白免疫组织化学染色,我和d)纤连蛋白玉米油WT (WT-control)和NGFtg (NGFtg-control)动物和NA-treated WT (WT-NA),与anti-NGF (WT-anti-NGF-NA)或控制免疫球蛋白(WT-IgG-NA)和NA-treated NGFtg (NGFtg-NA)动物分析在第十天。酒吧= 50μm规模。数据意味着±扫描电镜为每个组(n = 6)的老鼠。*:p < 0.05;* * *:p < 0.001。
神经生长因子促进肺克拉拉细胞扩散
后观察神经生长因子促进克拉拉细胞更新以及胶原蛋白的表达我和纤连蛋白表达,我们研究了神经生长因子的直接影响上皮细胞增殖BrdU的政府在活的有机体内(图印地补充材料)。量化BrdU-positive显示缺乏增殖细胞增殖的未经处理的WT和NGFtg控制动物的航空公司,而钠治疗组包含BrdU积极核(图7)。染色并行串行部分和BrdU CCSP显示增殖克拉拉细胞(图7)。损伤和细胞剥蚀因此诱发re-epithelialisation通过克拉拉细胞增殖增加肺。在抑制神经生长因子与anti-NGF抗体,显著降低数量的激增上皮细胞观察,而应用程序的控制免疫球蛋白抗体显示没有影响。符合这些结果,NGFtg小鼠接受NA展出的最高数量的激增上皮细胞(图7 b)。在一起,这些结果显示强大的神经生长因子对克拉拉细胞增殖的影响在修复受伤的肺。
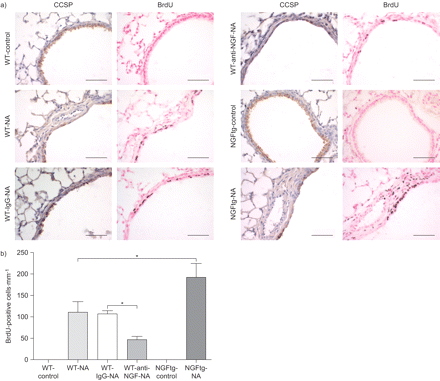
分析扩散BrdU (5-bromo-2′-deoxy-uridine)合并支气管上皮细胞。)串行部分野生型(WT)控制(WT-control)、萘(NA)治疗WT (WT-NA)控制免疫球蛋白(Ig) G (WT-IgG-NA)或anti-nerve生长因子(神经生长因子)(WT-anti-NGF-NA)和NGFtg控制(NGFtg-control)和钠治疗NGFtg (NGFtg-NA)动物彩色克拉拉细胞分泌蛋白(CCSP)和BrdU显示。CCSP-positive克拉拉细胞染色棕色对蓝色污点和BrdU-positive细胞表现出深棕色原子核对核快红计数器污渍。所有的动物BrdU NA显示积极对待核染色。酒吧= 50μm规模。b)量化BrdU积极核资料每毫米的基底膜钠治疗后第十天。数据意味着±扫描电镜为每个组(n = 6)的老鼠。*:p < 0.05。
讨论
神经生长因子的生成和其他家庭成员最初确定为神经元生长和存活的因素,但目前公认的有生物活性的组织外的神经系统31日。众所周知,各种内脏上皮细胞产生神经营养因子调节组织最初猜测的神经支配32和自分泌神经营养因子的功能越来越被描述。例如,神经生长因子的生产增加在炎症过程中伴有上皮损伤哮喘17,33、感染34和结节病35。
先前的报道表明,神经生长因子参与上皮伤口愈合在其他组织。局部应用神经生长因子已被证明加速伤口关闭和re-epithelialisation手术伤口的皮肤36和角膜23。此外,神经生长因子产生强烈的现场受伤的皮肤和增加在新形成的上皮细胞在伤口的边缘而不是周围的表皮角化细胞37。在角膜上皮损伤后观察神经生长因子的增加,角膜上皮细胞神经生长因子及其受体的表达TrkA被观察到38。上皮表面受伤时,正常的反应是移植的受体驱动扩散和生长因子修复39。本研究的目的是确定神经生长因子的作用在促进肺上皮细胞的增殖,尤其是克拉拉细胞组成主要气道上皮细胞群小鼠气道。
在伤口修复上皮神经生长因子的贡献是第一次调查在体外使用一个克隆小鼠肺肺泡II型上皮细胞系(LA4)和原发性鼠AECII。在体外实验与受伤LA4单层细胞显示神经生长因子及其受体蛋白表达增加TrkA受伤。此外,伤口关闭加速补充神经生长因子和抑制神经生长因子抑制。调查主要AECII细胞也表达神经生长因子和TrkA,显示AECII神经生长因子的趋化现象的反应。AECII细胞也回应神经生长因子表达增加胶原蛋白我和纤连蛋白mRNA。最重要的是,在这两种类型的细胞,神经生长因子被认为强烈调解扩散。然后我们调查了前导神经生长因子的函数在活的有机体内利用NA的克拉拉细胞损伤模型。
我们调查了功能性神经生长因子对肺部气道上皮修复的影响在活的有机体内利用克拉拉特异性损伤模型。克拉拉细胞钠介导损伤后的完整更新需要1 - 3个月。上皮损伤的NA模型研究29-day时间评估神经生长因子在肺上皮细胞修复的作用。为此,我们包括NGFtg动物神经生长因子过表达既定下CCSP在克拉拉细胞启动子。NGFtg动物表达神经生长因子水平高出155倍左右与WT老鼠。的存在增加神经生长因子水平在正常情况下没有造成变更在克拉拉细胞的总数或任何细胞表型的变化。此外,WT和NGFtg动物显示类似水平的损伤在第一天证明了相当数量的剥落了克拉拉细胞在老鼠菌株(图辅料S4)。NGF-overexpressing NGFtg动物失去了大部分的克拉拉细胞剥落;然而,剩下的细胞产生的水平明显高于神经生长因子与WT动物相比,这可能是与加速相关上皮修复NGFtg动物。克拉拉细胞损伤后,肺神经生长因子水平增加10天,直到天29日受伤后显著升高WT和NGFtg老鼠,强调神经生长因子的重要性持续肺修复过程。 This is supported by在体外发现与受伤的肺上皮细胞,观察神经生长因子的合成,调节早在伤后6 h和随时间逐步增加。有趣的是,缺乏增殖中观察到的肺NGFtg动物表明,神经生长因子的前导作用仅限于再生情况。因此,克拉拉细胞更新,至少在一定程度上依赖于神经生长因子的前导功能损伤,BrdU公司在活的有机体内在NGFtg增强动物和可以抑制anti-NGF抗体。
神经生长因子的过度NGFtg老鼠的肺之前已经报道过调节肺神经支配和调节气道高反应性25。神经生长因子已被建议作为一个潜在的候选人在哮喘气道重构的发生和报道诱导平滑肌细胞增殖40,41成纤维细胞的迁移和分化42。在这项研究中,神经生长因子似乎也调节胶原蛋白我和纤连蛋白表达,支持的观察在体外在AECII和在活的有机体内NA的克拉拉细胞损伤模型。胶原蛋白我和纤连蛋白mRNA表达显著增加在孵化主要AECII外源性神经生长因子和类似的结果在活的有机体内在较高的神经生长因子水平在转基因动物的肺损伤。基于这些发现,我们表明,神经生长因子可能调节ECM表达其他进程涉及损伤和修复,比如疾病如哮喘气道重构,神经生长因子被牵连。未来调查改造和纤维化可能的神经生长因子的作用机制,将修复愈合过程的线索病理事件。
总之,我们的研究结果提供的证据表明,神经生长因子协调增生性事件在肺上皮细胞的修复,促进克拉拉细胞的更新在活的有机体内。因此,神经生长因子可能被认为是一个重要的自分泌上皮生长因子在支气管气道管理维修。神经营养因子在最初被描述为分子,维持神经元产物和生存,现在有越来越多的证据表明,它们属于一类多功能信号分子,它还可以调节肺上皮细胞的修复和扩散。
确认
我们感谢美国奥肯博,大肠Kotte优秀的技术援助和t . Rausch执行CC10免疫组织化学。
脚注
可以从本文的补充材料www.www.qdcxjkg.com
支持声明
这项工作是支持的,在某种程度上,由德意志Forschungsgemeinschaft (Transregio 22日项目A9)和Stiftung毛皮Pathobiochemie和Molekulare Diagnostik, DGKL。
感兴趣的语句
没有宣布。
- 收到了2008年11月3日。
- 接受2009年11月4日。
- ©2010人队











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}