





本文的目的是讨论当前函数的概念基质金属蛋白酶(MMPs),重点是正宗的底物的识别。关注matrilysin (MMP 7)应承担的,它演示了如何一个细胞外蛋白酶可以调节上皮细胞生物学的关键和常见的功能,即抵御微生物,re-epithelialisation损伤和急性炎症反应。利用从急性肺损伤模型,发现matrilysin机制的行动在这些过程中,也就是说,它的作用在促进特定的蛋白质生物过程进行了探讨。
尽管他们专业化服务不同的功能,不同组织的上皮反应类似于由等效损伤和感染和调节炎症机制。通过形成一个屏障和释放抗菌产品,上皮细胞提供对入侵病原体的第一道防线。受伤后,上皮细胞发起一系列程序的协调响应恢复组织的完整性。生产化学引诱物,粘附分子,和其他蛋白质,上皮细胞招募和限制炎性细胞的损伤。虽然看似发散过程,上皮计划调节修复,国防和炎症可能共同演进,特别是对选择性地诱导的基因。毕竟,损伤提供了一个机会感染,这可能导致伤害;这两个事件促炎。因此,许多上皮产品相关的任何这些事件可能是常见的。Matrilysin (MMP 7)应承担的,是一个在国防、蛋白质功能修复,如下所示,炎症。
基质金属蛋白酶占23(目前)相关的一个家庭,然而不同的酶。基质金属蛋白酶分泌或固定在细胞表面1,2,从而限制其催化活性膜蛋白或蛋白分泌途径或细胞外空间内。顾名思义,基质金属蛋白酶被认为是负责营业额和降解结缔组织蛋白,一个函数,它可能是由一些家庭成员。然而,矩阵退化既不是唯一的也不是这些酶的共享功能。近年来一些报告表明,各种基质金属蛋白酶激活各种nonmatrix蛋白质,包括细胞因子、趋化因子、受体和抗菌肽3。因此,基质金属蛋白酶不应仅仅作为矩阵的蛋白酶催化,而是,正如上面提到的,参与调节细胞外酶处理信息和cell-matrix信号事件。
使用基因老鼠允许调查人员发现特定的定义,有时,意想不到的基质金属蛋白酶的功能。基质金属蛋白酶的目标日期,所有,除了地理MT1 MMP没有或只有轻微的表型小鼠的挑战,和一些基质金属蛋白酶似乎在营业额有直接作用或细胞外基质蛋白的降解2,3。这些观察表明,许多基质金属蛋白酶不提供至关重要的功能在开发或体内平衡;许多人甚至不表达的胎儿。相比之下,一旦挑战,MMP基因敲除小鼠揭示应承担的各种表型表明这些酶特定的服务,有时必要的角色在组织修复、血管生成、宿主防御,炎症和肿瘤进展等过程。因此,似乎一些基质金属蛋白酶,至少大部分的被淘汰,已经进化到应对extra-uterine存在的侮辱和压力。
Matrilysin的上皮是由几个noninjured noninflamed组织,如肺、肝和乳房。matrilysin健康上皮的表达表明它的功能在一个共同的稳态过程在上皮细胞中,这似乎是抵御微生物。在老鼠身上,matrilysin激活肠道pro量αdefensins应承担(cryptdins),一个家庭的抗菌肽,并由于缺乏成熟的α高defensins matrilysin-null(垫- / -)小鼠有能力受损肠道病原体4。除了生产的完整的组织,matrilysin表达在受伤的呼吸道和肠道上皮细胞迁移,表明这个MMP也在上皮修复功能。事实上,在野生型(WT)组织,气管的伤口迅速修复,而伤口垫- / -组织显示没有证据表明上皮移植,甚至具有抑制受损。延伸好几天5。事实上,垫- / -小鼠MMP-knockout中最严重的伤口修复缺陷小鼠生成日期2。
作为matrilysin需要关闭黏膜的伤口,这是推断垫- / -小鼠更容易受到严重的组织损伤。毕竟,如果re-epithelialisation受损,然后过度纤维化由于间隙的持续分泌液体,肉芽组织形成和纤维母细胞迁移,将随之而来。这个想法是使用bleomycin-induced的肺纤维化模型进行了测试。博来霉素,气管内的灌输,使快速和广泛的肺泡上皮细胞损伤,其次是严重的,持续的炎症和纤维化。
Matrilysin不是控制老鼠的肺,但表达的诱导肺泡上皮细胞与剥蚀接壤地区后滴剂0.04单位的博来霉素(图1⇓)。与预期相反,蛋白质含量在支气管肺泡灌洗(BAL)和新胶原的沉积是略低于垫- / -小鼠与WT老鼠。进一步探索这个意想不到的反应,老鼠灌输0.02 -16单位争光霉素和生存在未来30天内跟踪(图1 b⇓)。很令人惊讶的是,垫- / -小鼠免受bleomycin-induced杀伤力。与高剂量的博来霉素,大多数WT老鼠死了滴剂后10天,虽然大多数垫- / -小鼠仍然活着(图1 c和d⇓)。
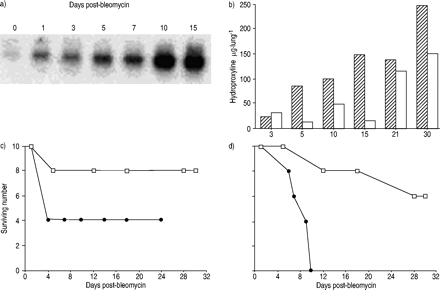
减少纤维化和增强生存matrilysin-null老鼠。)Matrilysin表达式是由逆转录酶聚合酶链反应评估/南部杂交post-instillation 0.04 U博来霉素在不同天。b)总胶原蛋白被羟脯氨酸水平评估总从野生型(└)和肺matrilysin-null(□)小鼠灌输为0.04 U博来霉素。野生型(•)和matrilysin-null(□)灌输与c) 0.04 U和d) 0.16 U博来霉素和生存之后30天。
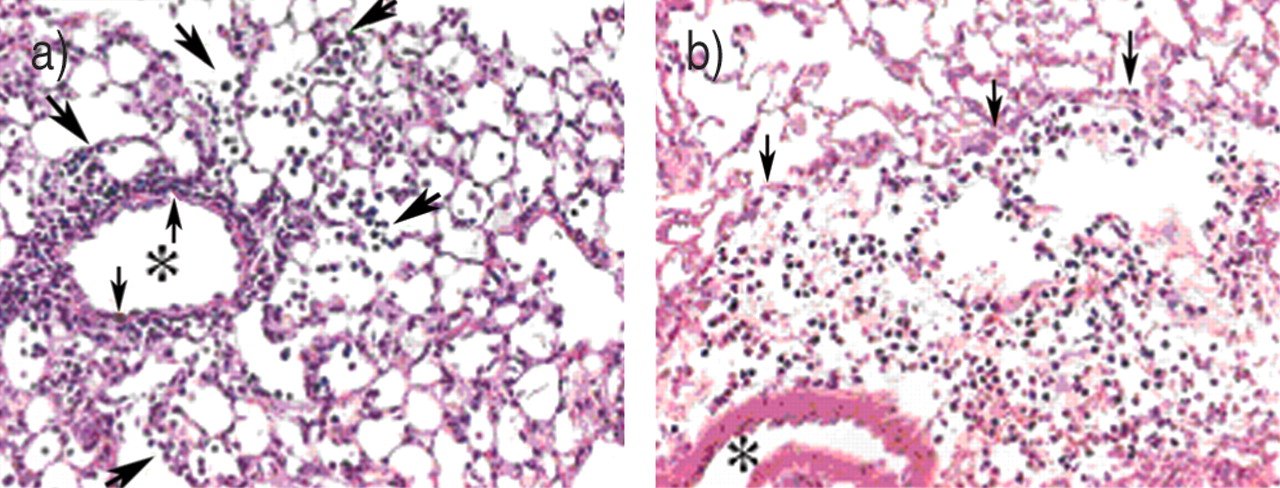
轻微的减少纤维化不是认为导致保护bleomycin-induced杀伤力。相反,它是推断matrilysin调节一种潜在的致命过程损伤的急性期,这个过程决定transepithelial大量中性粒细胞。而一些中性粒细胞从WT老鼠BAL中发现,很少有人从球垫- / -小鼠中恢复过来。然而,总中性粒细胞分离bleomycin-instilled肺组织没有WT和MAT - / -小鼠不同。因此,尽管大约相同数量的中性粒细胞移民到垫- / -和WT肺损伤后不久,这些细胞有能力受损推进从间质进入食管腔室matrilysin的缺失。的确,对中性粒细胞免疫染色显示这些粒细胞被困在肺泡壁或血管周的隔间(图2所示⇓)。因为matrilysin不是表达的小鼠中性粒细胞,内皮或间质细胞,中性粒细胞迁移的缺陷必须是由于一个epithelial-derived过程。
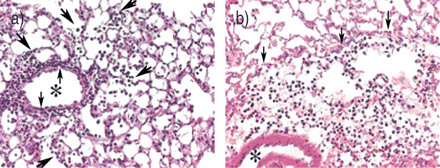
受损matrilysin-null小鼠中性粒细胞迁移。的小静脉腔由星号标记,血管周围和肺泡间隔之间的边界的小箭头,和中性粒细胞迁移到肺泡的大的箭头表示。
基于这些研究结果,推断matrilysin调节的活动epithelial-derived嗜中性粒细胞趋化现象的因素。KC的水平,一个鼠科学家趋化因子、白介素(IL)的功能性同系物的8和一个结构与生长有关的蛋白质的同系物α进行评估。KC是有效的和必要的中性粒细胞化学引诱物,由上皮快速表达,内皮细胞,间质细胞在各种炎症条件,包括肺损伤和感染。
KC水平BAL WT肺的在12 h post-bleomycin和返回控制水平升高了3天。相比之下,KC蛋白质仍处在或接近基线水平BAL bleomycin-treated垫- / -小鼠。基于等效信使核糖核酸水平,减少KC球垫- / -小鼠蛋白质并不是由于表达减少,和血清水平升高表明endothelial-derived KC鼠标菌株本质上是平等的。的意义,而KC蛋白质含量仍然相当恒定在肺提取物WT老鼠,他们积累了明显垫- / -小鼠的肺。
这些数据解释为表明matrilysin作用于蛋白质,促进了KC的分布从腔间质,和作者的集团专注于syndecan量1。IL 8,应承担的KC的人类同系物,需要吸引中性粒细胞硫酸乙酰肝素及其受体结合,CXCR2,高亲和力。Syndecans,四口之家的跨膜蛋白,细胞表面硫酸乙酰肝素的主要来源,可以交互通过与各种各样的蛋白质,碳水化合物链包括趋化因子6。Syndecans剥离损伤和炎症反应,这个过程被认为是其生物活性所必需的。自从syndecan 1是大量表达,其肺上皮细胞的基底表面,这是决定是否参与动员matrilysin衬底的KC。
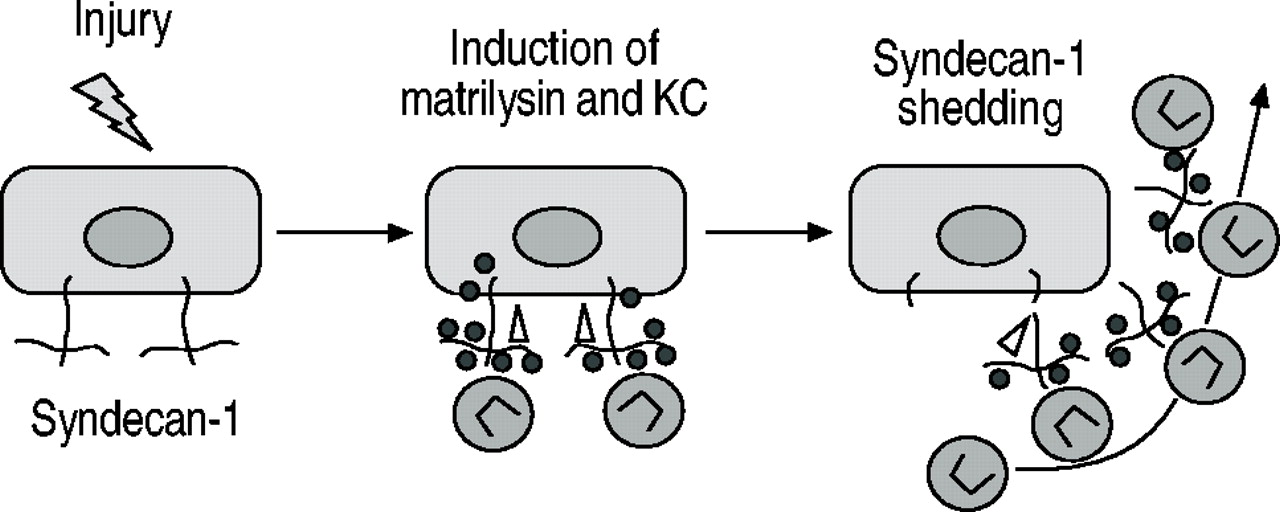
在未经处理的WT和MAT - / -小鼠,syndecan的等效信号量1被认为在进行气道和肺泡,和类似水平的蛋白多糖在洗涤剂中发现提取全肺。在bleomycin-treated WT肺、肺泡的广泛领域缺乏染色syndecan高1略低信号进行航空和显著增加,可溶性syndecan 1 BAL被发现。然而,垫- / -小鼠模式和syndecan 1应承担的染色强度相比并没有改变在控制肺部和可溶性syndecan BAL样本中发现不是1。使用免疫沉淀反应方法,KC被证明是一定会摆脱syndecan量1。此外,如垫- / -小鼠,KC水平和中性粒细胞数量减少BAL syndecan检测1零老鼠相比与WT老鼠。在缺乏syndecan高1或matrilysin KC不是易位到肺泡空间和积累的组织空间。在一起,这些数据表明,KC必将matrilysin-shed syndecan量1,这个协会是需要飞船KC进入肺泡空间(图3人受伤⇓)。
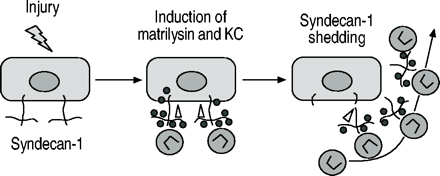
Matrilysin脱落syndecan检测1建立一个梯度指导transepithelial涌入的中性粒细胞趋化因子的趋化作用在受伤的网站。•:KC;▵:matrilysin。
结论
这些数据描述三个上皮组件的一种机制,一个分泌蛋白酶(matrilysin),一个cell-bound蛋白聚糖(syndecan检测1),和趋化因子(KC),行为协调控制和限制炎症的损伤7,8。作者的组织建议受伤,上皮细胞合成、分泌和存款KC(或者IL 8在人类)到预先存在的syndecan 1分子在基底表面。Matrilysin也引起损伤和劈开syndecan检测1核心蛋白质释放ectodomain KC复杂。摆脱复杂的运输,主动或被动地,顶端表面创建一个趋药性的梯度指导中性粒细胞进入肺泡空间⇑⇑⇑。
- ©人期刊有限公司










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}