





文摘
基因组学和相关的下游技术产生巨大的数据集,为理解提供新的机会和打击传染病和人类遗传疾病。
基因组的方法已经应用于肺结核、传染性发病率和死亡率的主要原因,显著的成功。完整的基因组序列现在可用的三名成员结核分枝杆菌复杂的和相关的细胞内病原体麻风杆菌。
许多预测生成的在网上通过功能分析基因组学已经验证,包括转录组和蛋白质组的研究,导致重要基因的鉴定。知识,后者定义了新的和现有药物及其潜在目标专一性可以通过比较基因组学与评估主机或其他病原体。基因组学也进一步加强结核病疫苗研发通过确定潜在的抗原蛋白,以及提供更好的诊断工具来检测感染。
有一个日益增长的需要新的药物和疫苗来治疗和预防分枝杆菌疾病,尤其是肺结核(TB),改进诊断工具来检测感染更可靠。1990年代看到的广泛出现耐多药菌株结核分枝杆菌(mdr - tb)在发展中国家和工业化国家1,2。耐多药结核病的传播已经记录,尤其是在人类免疫缺陷病毒(HIV)来华,和无法治愈的疾病的幽灵正迅速成为一个现实。尽管人们普遍认为疫苗接种是最可取的方法预防结核病,有大量的证据表明目前的疫苗,卡介苗(BCG),只是对少,有效传播形式的疾病3,4。BCG预防肺结核的效果非常有限,占大部分的疾病负担5和一直未能带来显著的保护发展中国家尽管诱导保护性反应对麻风病在相同的设置6。后者的观察表明,麻风杆菌可能与BCG共享共同的抗原,反过来,与它的近亲结核分枝杆菌。
肺结核药物开发:古代和现代
现用药物的大多数antituberculous是在1950年代开发的产品,源自微生物屏幕自然或化合物库,或偶然导致的结果。链霉素,1943年第一个抗生素的发现,显示治疗感染是由于一些革兰氏阴性细菌,和非常强大的反对后不久结核分枝杆菌。一次偶然的观察7这烟酰胺抑制小鼠的分枝杆菌的生长导致相关化合物的合成和测试,并最终以精美的特定药物异烟肼(INH)8。测试最初进行的活动在体外种植的生物,然后确认在临床试验前感染动物模型进行了有希望的分子。随后,额外的药物开发抑制特定的特性结核分枝杆菌,如生物起源的非凡的细胞壁。理想情况下,抗菌药物显示杀菌活动和目标必不可少的活动。精确定位等功能的一个手段,它从未被应用于结核菌,是分离和描述有条件地致命缺陷的突变体,然后筛选抑制剂能产生同样的效果。这可以通过监测微生物参数,比如增长率,或者通过使用一个在体外分析如果合适的功能测试存在。如今,新药的识别可以从一个理性的结果,假说驱动的方法受基因组学或从化学或组合库的大规模筛选各种自动化的方法。所需的属性的新抗结核的药物包括减少治疗的持续时间,以及活动对潜伏结核感染和耐多药结核菌株9。
基因组学
基因组学、成套的系统研究遗传物质的细胞,通过脱氧核糖核酸(DNA)测序和生物信息学分析,提供了巨大潜力的药物目标抗原的发现和提高新的抗菌药物和疫苗的发展。相对温和,单笔投资的整个补基因病原体可以定义及其在基因组序列的序列与其它生物包括微生物、老鼠和人10,11。结核病研究领域,基因组学最早应用于H37Rv12广泛使用的范式的结核分枝杆菌,然后CDC155113,从美国近期临床分离,然后AF2122/97应变的牛分枝杆菌14负责的结核病流行在牛和獾在英国(表1所示⇓)。基因组序列的结核菌的近亲,麻风杆菌,也已确定15。
基因组学和药物发现目标
所有三个全测序的基因组的成员结核分枝杆菌复杂的包含∼4.4 (+ /−0.1 Mb和港口∼4000基因预测编码蛋白质和基因稳定的核糖核酸(RNA)的物种。在的情况下结核分枝杆菌H37Rv,生物信息学分析导致精确函数的归因∼40%的4000个基因。一些功能的信息进一步推断为20%,但没有学会了剩下的40%12,16。功能信息可用时,它往往使调查人员能够识别潜在的药物靶点的基础上,提出了生物作用或相似已知细菌的药物靶点。然而,现在有了更多的分枝杆菌序列,可以确定哪些基因通常分枝杆菌或局限于一个给定的物种17,18。这些基因编码的功能,如果必要,可以代表小说化疗是非常具体的目标。相比之下,大量的未知功能的基因被发现在许多细菌,这些通常被称为守恒假设基因19,20.。他们广泛的保护无疑是生物意义和其中一些基因随后被发现扮演至关重要的角色。因此,他们代表新靶点为新广谱抗生素19。确保新的抗生素抑制功能局限于细菌,从而减少人类潜在的副作用,现在可以执行在网上人类基因组序列的筛选10,11排除相关的基因或蛋白质的可能性将会发现在主机上。类似的其他病原体的基因组序列的屏幕可以进行提高特异性。许多有吸引力的特性的非常具体的药物,如异烟肼和吡嗪酰胺,避免转移耐药机制,比如那些困扰某些广谱抗生素,减少不必要的副作用,比如无差别破坏肠道菌群。
通过功能基因组学验证药物靶点
有几种不同的方法来确定哪些基因结核分枝杆菌是必不可少的,因此值得进一步研究作为药物开发的目标。这些包括基因淘汰赛,记录分析和蛋白质组的定义。下面是一个简短的描述可用的策略。确定了潜在的候选人,证明基因表达是很重要的,特别是在感染,转录组和蛋白质组学提供了巨大的希望通过允许全球分析承担。所有这些方法大大促进的完整基因组序列的可用性12。
转录组
各种方法已经被用来监控转录组,结节杆菌产生的完整的RNA分子,包括目标和随机方法。后者包括微分表达式定制放大库(贴花)21和选择性捕获的转录序列(苏格兰)22技术,结合聚合酶链反应(PCR)和减法杂交以识别基因表达差异。贴花纸应用于检测时的差异表达与异烟肼对治疗的反应,三过表达基因(iniA, iniB和iniC)的表达式也诱发了乙胺丁醇,另一个药物的靶向细胞信封,和其他治疗影响细胞壁的功能,被发现21。这个建议的作用iniABC可能关键的细胞被膜生物起源,虽然它的功能仍然是未知的。
苏格兰的应用程序来检测基因差异表达在人类主要的巨噬细胞22,建立了几个基因调节。其中有两个可选σ因子(锗硅和叹息),先前与生存压力23、酮化合物合酶(pks2)、异柠檬酸裂解酶(究也被称为icl),一种酶,这种酶已被证明是需要长期坚持的结核分枝杆菌在受感染的老鼠24,mce1B一个基因位点,可以赋予海拉细胞内的入侵和生存能力大肠杆菌25。这些候选人,异柠檬酸裂解酶的晶体结构26,似乎是最有前途的药物目标干预在乙醛酸分流,生化途径仅限于特定的微生物和植物。
DNA微阵列是一种强大的工具为研究基因表达差异,通常由gene-specific探针,固定在固体表面如玻璃,作为杂交模板27。可以用来比较基因表达微阵列在活的有机体内与观察在体外,虽然结果结核分枝杆菌在组织尚未报道。然而,在最优雅的一项研究中,威尔逊et al。28检查的转录响应结核菌异烟肼。超表达的基因编码组件FAS-II脂肪酸合酶系统的检测,如预期从蛋白质组学的研究29日,加上fbpC地理编码大量分泌抗原85 C trehalose-dimycolyl转移酶活性和干涉的最后步骤分枝杆菌细胞壁的合成30.。其他INH-induced基因fadE23和fadE24、编码两个acyl-coenzyme脱氢酶参与脂肪酸β氧化应承担的ahpCalkyl-hydroperoxide还原酶,编码,这可能对异烟肼的中等毒性作用。另外两套调节基因值得进一步的评论:efpA编码一个假定的射流系统,可能在先天耐药中发挥作用28和iniAB,被贴花21上面所讨论的。从这些研究可以看到,微阵列和相关转录组工具是非常有用的对于识别coregulated基因和可以帮助确定集的基因和蛋白质的协同行动。这代表了一个强大的方法来发现额外的药物靶点在同一个通路,从而提高协同效应的可能性。
蛋白质组学
监测基因表达的另一个方法是通过研究蛋白质组,完成的一组蛋白质结节杆菌在不同条件下产生的。最全面的研究蛋白质组的不同菌株结核分枝杆菌复杂的被Jungblut报道et al。31日用二维电泳和质谱分析解决和识别蛋白质,分别。类似的方法已经被应用到文化滤液32安德森和可溶性蛋白质和同事33,34。细节的网站提供这个信息表1中给出⇑。
用两种不同的比较蛋白质组学进行结核分枝杆菌菌株(H37Rv和Erdman)和两株牛分枝杆菌波士顿咨询公司(哥本哈根和芝加哥)。一千八百点对应于分枝杆菌细胞蛋白质和> 200年文化浮在表面的检测,分别确定了其中263和5431日,35。识别蛋白质,一些执行管家功能而其他人参与脂肪酸和糖脂代谢。热休克蛋白质非常丰富,而只有三个点对应于假定的细胞被膜蛋白。
蛋白质组学也可以作为一种工具用于抗原的发现。对比BCG芝加哥和H37Rv显示31变量蛋白质。十三H37Rv在场,但不是在疫苗株,六个被确定的。8蛋白似乎是缺席H37Rv,九个表达下调,一个是过表达对BCG。八的差异是由于流动变异,可能引起的氨基酸变化或转录后修饰。进一步比较两个结核分枝杆菌菌株显示只有轻微的差异蛋白质表达谱在体外31日。这一发现已被证实在一个独立研究中,蛋白质组的菌株H37Rv CDC1551相比36。
最近,二维液相电泳被用来获得几百个分数文化滤液和胞质蛋白,分别。这些都是筛选的能力刺激T细胞反应和30个人应承担的蛋白质被质谱鉴定,其中17小说T细胞抗原37。目前没有可用的信息从巨噬细胞结节杆菌蛋白质组的分离,甚至病变组织,这应该调查集中在未来几年。同样,对蛋白质存在于颗粒细胞的一部分,当一个人认为60%的已知的药物靶点是膜蛋白,很明显,这是一个需要解决的问题。
替代蛋白质组学方法存在协同识别蛋白质相互作用或功能。例子是酵母2台混合动力系统38或串联亲和纯化(TAP)系统39。然而,目前的作者的知识,这些尚未应用于任何重大规模的结节杆菌。蛋白质/蛋白质相互作用也可以预测在网上使用最近开发的算法,比如系统发育分析,在给定的蛋白质的分布目标物种是检查,或通过比较目标细菌序列的多畴的真核蛋白质检测守恒的片段可能表明干预一个共同的途径40- - - - - -42。一系列有趣的预测由这些技术已经被报道43和现在正在实验进行验证。
基因替换
基因能有效灭活结节杆菌通过等位基因交换,利用单倍体或部分二倍体主机,由自杀向量携带有缺陷的基因44或有条件地复制mycobacteriophages45。随机的基因失活可通过传统或“signature-tagged”转座子突变45- - - - - -47。测序的网站插入大量的自然(6110年)或人工(1096年的)转座子提供不必要的基因信息。signature-tagged诱变方法允许中断和不需要不必要的基因的识别在体外但在结核病小鼠模型中起到至关重要的作用46,47。几千signature-tagged突变体测试中两个独立的屏幕,最减毒株包庇转座子基因参与脂质代谢和合成和出口phenolphthiocerol dimycocerosate,特有的蜡仅限于致病性分枝杆菌。消除了不必要的基因让一组> 2000个基因,可能是必要的。然而,考虑到相对较大的基因组的大小结核分枝杆菌,最重要的是,其生成时间长,不太可能批发这些基因的失活等位基因交换将承担。评估重要性的另一个方法是使用反义RNA,但这尚未广泛应用结核分枝杆菌。间接识别重要基因的另一种方法是使用比较分枝杆菌基因组学。
比较基因组学的结核分枝杆菌复杂的
比较基因组学是一个强大的新工具探索微生物的进化和识别基因可能编码新的药物靶点或保护性抗原。基因的多样性结核分枝杆菌复杂的DNA芯片技术研究,促进所有成员共享一个> 99.95%的身份在DNA水平48。检查不同的卡介苗,总共18删除区域(RD1-RD18)发现了几个实验室49- - - - - -51,其中一些特定的菌株。这导致修订BCG的谱系和发现120个基因中存在结核分枝杆菌H37Rv但缺席BCG巴斯德。这种差异可能占疫苗和病原体之间的表型差异。只有一个地区,RD1、缺少BCG菌株,但存在于另一个结核分枝杆菌复杂的成员49,50,52。RD1可能损失可能占的衰减牛分枝杆菌最初由Calmette描述53。
比较基因组学还发现了两个串联重复29和36 kb () DU1 DU2)的染色体牛分枝杆菌BCG巴斯德,这表明该疫苗株部分二倍体58基因54。这些比较基因组的联合调查结果分析表明,有大量不同BCG菌株之间的遗传多样性在今天使用,这可能解释观察到的变化在不同的疫苗试验对结核病。
比较基因组学的各自的成员结核分枝杆菌复杂的揭示了一个基因的存在梯度。人类结核菌,结核分枝杆菌更多的基因m . africanum microti和牛分枝杆菌,因为这些物种失去了遗传物质通过删除事件50,55。基因发生在物种内的高频损失结核分枝杆菌结果的副本之间的同源重组事件6110年侧翼基因直接取向50,55,56。微阵列和Affymetrix芯片的研究已经发现了一个额外的45个基因群的存在,并可能功能,是兼性51,57。从合并后的结果可以得出结论,> 200个基因存在不增长的必要条件结核分枝杆菌复杂的成员主机,但可能影响程度的毒性。
比较分枝杆菌基因组学
一个更强大的方法来减少潜在的新目标结核分枝杆菌更多实实在在的水平可能会发现比较基因组学与麻风杆菌、异常生长缓慢的专性细胞内病原体显示本质上相同的细胞趋向性和宿主范围结核菌。3.27 Mb的基因组麻风杆菌的规模远小于结核分枝杆菌,只有49.5%的1605个蛋白编码基因15。麻风杆菌似乎经历了还原进化,这一过程涉及大量的裁员和基因衰变。大约27%的麻风杆菌基因组包含伪基因,不活跃的阅读框架仍然有结核菌的对应物,其中1114是家喻户晓,而剩下的23.5%的基因组似乎非编码和可能是功能性。还原进化已经被记载在专性细胞内病原体和内共生体,等立克次氏体和Buchnera分别。15。基因逐渐失去了作为他们的功能不再需要在高度专业化的领域,这可能占的异常长时间代麻风杆菌。总之,假设所有的分枝杆菌都起源于一个共同的祖先,麻风杆菌可能已经失去了> 2000个基因在进化和所需的最小基因集合定义的致病性分枝杆菌可能是自然吗15。
当两两比较的基因和蛋白质组麻风结节杆菌12,15,16,581433年进行蛋白质被发现是常见的病原体。切除后的蛋白质与其他原核生物(放线菌除外)和真核生物样品只包含333蛋白质。因为这些致病性分枝杆菌占据人体类似的利基市场,在那里他们遇到相同的生理压力和免疫反应,可想而知,这些基因可能会影响产品高度专业化的功能可以为胞内分枝杆菌的增长至关重要。如果是这种情况,相应的蛋白质或酶可能代表小说药物靶点。发现的333名候选人比较分枝杆菌基因组学可以分为局限于属这些蛋白质分枝杆菌(其中有219),第二组114多肽也发生在链霉菌属或棒状杆菌spp。有关放线菌目王国的成员。它是合理的假设,后者蛋白质放线菌赋予特定的属性,而那些局限于分枝杆菌可能发挥更专业的作用。
目标发现
进一步说明比较分枝杆菌基因组学的有效性识别潜在的重要蛋白质,两个精确的例子将被给予。发生在多个基因重复事件结核分枝杆菌当有限的分歧之后,这似乎在很多生化途径导致广泛的功能冗余12,16,58。事实上,在许多情况下很难预测与确定性的两个重复基因影响特定的功能。这是真的5蛋白(Rv0462、Rv0794c Rv2855, Rv2713, Rv3303c)显示强烈的相似性对数据库搜索各种lipoamide脱氢酶组件丙酮酸脱氢酶复合体的一个重要功能。在最初的分析结核分枝杆菌两种蛋白质基因组,显示这个脱氢酶被称为最强的相似性对数(Rv3303c)和lpdB(Rv0794c),但后续的生化研究的各种基因产物显示真实lipoamide脱氢酶是由Rv0462编码(现在称为lpd),而不是对数或lpdB基因59。检查麻风杆菌基因组序列有助于关注这项工作,因为只有这五个之一结核分枝杆菌相应基因的功能orthologue ML2387 Rv0462,真正的lipoamide脱氢酶。剩余的四个基因存在假基因的形式麻风杆菌基因组15,60。这是强有力的证据比较基因组学的力量。
下面是第二个例子,等待生化确认。Preproteins twin-arginine运输出口,或答,途径通常结合氧化还原代数余子式和褶皱或oligomerise之前穿过细胞膜61年,62年。切除信号肽后,许多这些蛋白质的功能extracytoplasmic电子传递链。认可的专业机械twin-arginine主题63年并把preprotein跨膜是由几个不同的答蛋白质。在大肠杆菌,塔塔和泰特50%是相同的,与TatB分享弱相似62年。所有三个蛋白质预测固定在细胞质膜通过一个N末端疏水α螺旋和细胞质两性分子的螺旋变量地区紧随其后。TatC蛋白预测与六不可或缺的膜蛋白跨膜段。结核分枝杆菌和麻风杆菌都含有明确的识别塔塔,tatB,tatC和tatD因此,基因和必须产生一个功能答系统。
蛋白质组的检查结核分枝杆菌11潜在基质答出口系统认可的基础上他们的信号肽含有潜在twin-arginine图案N的终点站和同源主题,S / TRRXFLK63年(表2⇓)。在广泛的还原的基因组的进化麻风杆菌只有一个相应的基因,ML1190,逃脱了失活。Rv2525c是同源的结核分枝杆菌但没有显示相似其他蛋白质存在于当前序列数据库。240年氨基acid-long前体蛋白编码ML1190 / Rv2525c包含五个组氨酸和一个半胱氨酸残基,对协调二价金属离子可能是重要的。这个编码序列的保护麻风杆菌,在面对大规模的基因丢失,是一个强有力的迹象表明,它必须发挥重要的生物作用。考虑到许多与乙系统其他地方,可能是电子传递。这些间接参数表明,一方面,如果这个函数是必不可少的,ML1190 / Rv2525c基因产物可能代表一种新型药物或目标,另一方面,因为它可能是位于细胞外,它可能是一个重要的哨兵蛋白质抗原。
筛选药物靶点
如果相应的蛋白质有assayable函数,例如激酶活性,它可以用作一个的基础在体外丰富的屏幕识别抑制剂或纯化酶。这种方法的优点是,它通常可以自动或转化为高通量格式方便筛选的大型或复杂的化合物库的合成或天然产品。然而,整个有机体屏幕,涉及重组结核分枝杆菌菌株与记者的活动,如荧光素酶或绿色荧光蛋白64年,65年,常常被认为是合适的,因为它们避免药物渗透问题。一旦一个活跃的药效基因被发现,许多类似物可以合成或组合库中识别孤立更积极的衍生品。他们的力量也可以评估使用记者化验或生化技术,如转录组和蛋白质组分析。coregulated可以这样,基因发现的产品也可以作为潜在的药物靶点,因为他们经常在相同的代谢过程行动自如。识别目标使大量的相应的蛋白质是由基因工程进行进一步的研究。知识的三维结构已知的或潜在的药物靶点也高度理想的用于药物开发,可以通过结构生物学。
结构基因组学
结构基因组学是一个新兴学科,利用高通量克隆技术来生成系统大量的表达式构造相应的大多数,甚至全部的基因存在于微生物基因组(表1所示⇑)。这些构造用于生产大量的标记蛋白,可以净化一个一步强大的亲和层析的方法,从而促进下游结构分析,X射线晶体学、核磁共振(NMR)谱或高分辨率低温电子显微镜。当前经验预测60%的成功率在表达阶段,随后∼20%的纯化蛋白质晶体衍射级。结构基因组学是一种很有前途的药物和药物发现的新方法,因为它容易接口和基于结构的药物设计66年。对于较大的基因组,蛋白质结构基因组学通常集中于大家庭收益最大化,而在药物发现项目的方法通常是假说驱动的。
诊断
活动性疾病的诊断,这在很大程度上依赖于临床专业知识和痰涂片抗酸的细菌的检测,比较基因组学和潜伏性结核病也将受益。潜伏性感染通常是通过监测诊断延迟型超敏反应的程度结核菌素皮内注射后,一个模糊的混合抗原。结核菌素反应是有限的价值在社区BCG接种疫苗,其解释也可能被感染涉及其他分枝杆菌抱愧蒙羞。120个基因的识别结核菌49,50缺席BCG,允许的发展走向一个更具体的测试,可以区分感染和免疫。芯片和蛋白质组学也会广泛应用于监控内生物多样性结核分枝杆菌复杂和帮助确认候选诊断抗原的存在与否。
发现抗原和疫苗策略
了解蛋白质的亚细胞定位是特别有价值的设计新的结核病疫苗,预防和治疗,因为人们普遍认为surface-exposed或分泌蛋白对应的抗原成分在感染免疫系统的第一次遇到67年。图1⇓提出了一种方案,可以找到这样的蛋白质。系统采用生物信息学识别细胞被膜蛋白本土化。这些包括跨膜蛋白疏水性域和脂蛋白N量终端修改的半胱氨酸残基增加脂质组。分泌的蛋白质通过一般的分泌途径68年很容易识别的特征信号肽的metallo-enzymes答分泌的系统61年- - - - - -63年。缺乏信号肽和其他蛋白质从分枝杆菌分泌Sec-independent方式包括那些属于早期分泌抗原(计)量6家庭目标12,58。计检测6是一个强大的T细胞抗原诱发强烈的T辅助1列车类型反应69年并已进行了广泛的研究作为一个潜在的感染诊断试剂70年从波士顿咨询集团以来,基因缺失50,51,53,71年,作为亚单位疫苗的一个组成部分72年。除了高度纯化的蛋白质设想作为亚单位疫苗组件,其他策略正在追求。这些包括核酸疫苗73年,74年建设合理的减毒突变体结核分枝杆菌和改进的BCG菌株75年。所有这些方法提高了基因组序列的可用性12。
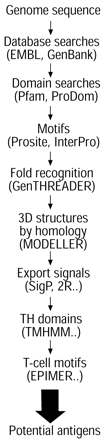
计划使用生物信息学工具来识别潜在的抗原蛋白分析基因组数据。显示在括号中使用的数据库或程序。看到主要的文本为进一步的细节。3 d:三维。
免疫原性的评价在网上和在活的有机体内
比较在网上蛋白质组的分析结核分枝杆菌减少了潜在的亚单位疫苗候选人的数量从∼4000蛋白质< 40潜在出口多肽,它既存在于麻风结节杆菌。这些属于蛋白质的几个家庭总是更大的保守序列结核分枝杆菌比在麻风杆菌。比较基因组分析强调了其潜在的酯酶活化剂检测6蛋白的重要性12,58,连同他们可能分泌机械、和初始免疫描述取得了令人鼓舞的结果76年。建立这些蛋白质的候选人包含潜在的T细胞抗原表位,应承担的差向异构体等强大的新算法77年,可用于进一步精炼的选择。基因组序列的可用性的老鼠和人类10,11将允许潜在crossreactive抗原表位或抗原识别在网上,从而限制可能出现的并发症。最终,方法需要测试候选人的成功保留在细胞和动物模型为肺结核,并建立相应的关联的保护。
确认
作者想给特别感谢分枝杆菌基因组组织巴斯德研究所和桑格中心。这项工作被威康信托基金会支持的巴斯德研究所、欧洲共同体(qlrt qlk2 - ct - 1999 - 01093 - 2001 - 02018),和法语协会拉乌尔Follereau。
- 收到了2002年1月28日。
- 接受2002年3月13日。
- ©人期刊有限公司









{kind=link}
{kind=link}