





摘要
气道上皮的异常生长和气道壁的所得到的增厚可能产生气道口径的变窄,从而有助于支气管中的慢性阻塞性肺病(COPD)的恶化。β2肾上腺素能激动剂已广泛用于COPD的治疗,但其对气道上皮细胞生长的影响尚不清楚。
研究了三种人气道上皮细胞系的生长情况体外.在无血清培养基中暴露沙丁胺醇可使16人支气管上皮(16-HBE)细胞和NCI-H292细胞中3‐(4,5-二甲基噻唑- 2 -基)‐2,5‐二苯四唑溴化还原和细胞内脱氧核糖核酸(DNA)含量增加,但在A549细胞中没有。沙丁胺醇对16-HBE细胞的促生长作用与10%胎牛血清等效,且被普萘洛尔和环磷酸腺苷(cAMP)拮抗剂rp -腺苷3′,5′-环磷酸硫代三乙基铵盐(Rp-cAMPS)抑制。同样,福斯柯林和8 -溴腺苷3 ',5 ' -环磷酸(8 - Br-cAMP)也能促进细胞生长和DNA合成。Western blot分析显示,沙丁胺醇、福斯考林和8 - Br-cAMP均诱导了丝裂原活化蛋白(MAP)激酶磷酸化形式的表达,而沙丁胺醇诱导的磷酸化被普萘洛尔、rp - camp和MAP激酶抑制剂PD98059抑制。
这些结果表明β在某些气道上皮细胞中具有刺激作用2肾上腺素能受体和随后产生的环磷酸腺苷可能上调细胞生长,可能通过激活丝裂原激活的蛋白激酶级联。
本研究部分由日本教育部,科学和文化部的赠款号06670243得到支持。
丝裂原活化蛋白(MAP)是一个42和44 kDa的胞质丝氨酸-苏氨酸激酶家族1那2已被鉴定为许多细胞类型的生长和增殖反应的重要调节成分3..越来越多的证据表明,地图激酶可以通过各种信号激活,包括生长因子4.,Phorbol酯5.和加利福尼亚州2+6..此外,与Gi和Gq蛋白相关的细胞表面受体的刺激已被证明能有效激活MAP激酶7.那8.,但Gs蛋白偶联受体对酶活性的影响尚不清楚。
β-肾上腺素能受体是GS蛋白偶联的受体,并刺激细胞内环状腺苷一磷酸盐(营地)的合成。吸入β.2- 一种肾上腺素依赖者激动剂已被广泛用于治疗慢性阻塞性肺疾病和哮喘,并且很可能在高浓度下暴露于这些药物。最近已经证明CAMP刺激了听觉受体上皮细胞的增殖9..然而,营养依赖性信号通路在气道上皮细胞增殖反应中的作用是未知的。因此,目前的作者研究了三种人类气道上皮细胞系体外,为了确定是否β2- 肾上腺素能激动剂影响呼吸道上皮细胞的生长,并评估阵营介导的MAP激酶级联的阵营介导的刺激。
材料和方法
细胞生长
人支气管上皮(HBE)细胞系16-HBE来自心血管研究所(University of California, San Francisco, CA, USA)。人气道黏液表皮样癌细胞系NCI-H292和肺癌上皮细胞系A549购自美国型培养馆藏(Rockville, MD, USA)。细胞培养于含10%胎牛血清(FBS)的Ham's F12培养基中,100 U·mL-1青霉素,100μg·ml-1链霉素和100μg·ml-1两性霉素B在二氧化碳中(CO2)孵化器(5%CO2:37°C时,95%空气)。在融合生长后,将细胞用0.25%胰蛋白酶/ 0.02%乙二胺四乙酸(EDTA)溶液分离以进行通过。在初步实验中,以这种方式制备的细胞保持相同的形态≥15通道。
用含10%胎牛血清的Ham's F12培养液100µL,密度为1×104.在96孔微量滴定板中每平底井的细胞,并生长为分组。用磷酸盐缓冲盐水(PBS)洗涤细胞三次后,通过在没有FBS的Ham F12培养基中孵育72小时,通过孵育来阻止细胞生长。然后将培养基替换为无血清HUM的F12培养基,含沙丁胺醇(1×10-7M) (Sigma Chemical Co., St Louis, MO, USA),细胞进一步培养。对照组在无血清培养基中进行细胞培养。使用MTT染色技术,在暴露于沙丁胺醇后的指定时间评估上皮细胞的生长,该技术依赖于活细胞对四唑盐3‐(4,5-二甲基噻唑‐2 -基)-2,5-二苯基四唑溴化(MTT) (Sigma化学公司)的特定代谢还原10.MTT(100µL的10µg·mL-1每孔加入PBS), 37℃孵育4 h。从孔中取出培养基和MTT,连续加入25µL的Sorenson's甘氨酸缓冲液(0.1 M甘氨酸和0.1 M NaCl用0.1 N NaOH平衡到pH为10.5)和200µL二甲亚砜。使用免疫吸附分析平板阅读器(450型;Bio-Rad Laboratories Ltd, Watford, UK)对抗试剂空白(IE。无细胞),测试波长570 nm,参考波长630 nm。
为了确认沙丁胺醇对细胞增殖的影响是由β-肾上腺素能受体介导的,并且在含有Salbutamol(1×10的无血清培养基中,将16-HBE细胞培养16-HBE细胞(1×10-7β肾上腺素能受体拮抗剂普萘洛尔(1×10)-5 M) (Sigma Chemical Co.) or the cAMP antagonist Rp-adenosine 3′,5′-cyclic monophosphorothioate triethylammonium salt (Rp-cAMPS) (1×10-4M), MTT测定(德国不来梅BIOLOG生命科学研究所)。为了评估camp依赖的信号转导通路的贡献,forskolin对细胞生长的影响(1×10-7M) (Sigma化学公司),腺苷环化酶和8 -溴腺苷3 ',5 ' -环磷酸一磷酸(8 - Br-cAMP)的刺激剂(Sigma化学公司)(1×10-7M)是cAMP的稳定类似物,用类似的方法进行了研究。在评价这些药物的效力时,将其与培养48小时后10%胎牛血清诱导的反应进行比较。
脱氧核糖核酸合成
用Rago法测定培养的上皮细胞脱氧核糖核酸(DNA)含量等等。11.简而言之,将细胞培养在含有Salbutamol的无血清HU12培养基(1×10-7米)。每孔加入100µL蒸馏水,孵育1 h。为了破坏细胞膜,将细胞冷冻在−80℃,并在室温下解冻。Hoechst 33258原液1份(200µg·mL-1(Sigma Chemical Co.)使用前用TNE缓冲液(10 mM Tris, 2 M NaCl, pH 7.4, 1 mM EDTA)稀释10倍,每孔加入100µL稀释后的Hoechst 33258。荧光用分光荧光计(Ultrospec Plus;Phamacia LKB Biochrom Ltd, Cambridge, UK)的激发波长和发射波长分别为350 nm和460 nm。此外,暴露于forskolin (1×10-7米),8 Br-cAMP应承担(1×10-7和沙丁胺醇(1×10-7M)加上心得安(1×10-5米)。
为了确定MAP激酶级联和cAMP在沙丁胺醇诱导的DNA合成中的作用,16-HBE细胞在含沙丁胺醇的无血清培养基中培养(1×10-7PD98059(1、30和100µM) (New England Biolabs, Inc., Beverly, MA, USA)是MAP激酶的特异性抑制剂12,或与RP-CAMP(1×10-4 M), and the DNA contents were measured 48 h later. In this experiment, to block MAP kinase cascade, a MAP kinase-kinase inhibitor was used because a specific MAP kinase inhibitor was not available.
Western Blotting.
细胞裂解物在含10µg·mL的放射免疫沉淀(RIPA)缓冲液中溶解(0.15 M NaCl, 50 mM Tris-Cl pH 7.4, 0.5% Nonidet P40 (NP40)和0.1%十二烷基硫酸钠(SDS))-1Leupeptin,1mM苯基甲基磺酰氟化物,10μg·mL-1抑肽酶和1mm钒酸钠,然后通过sds -聚丙烯酰胺凝胶电泳分离,转移到聚偏二氟乙烯膜上,并用针对MAP激酶和磷酸化MAP激酶的抗体(1:10 000)探测(New England Biolabs, Inc.)。使用增强化学发光法(SuperSignal, Pierce, Rockford, IL, USA)观察一抗(1:25 500)。
统计分析
所有值均表示为平均值±sem。采用单因素方差分析和Newman Keuls多重比较检验确定组间差异有统计学意义,p<0.05被认为是显著的。
结果
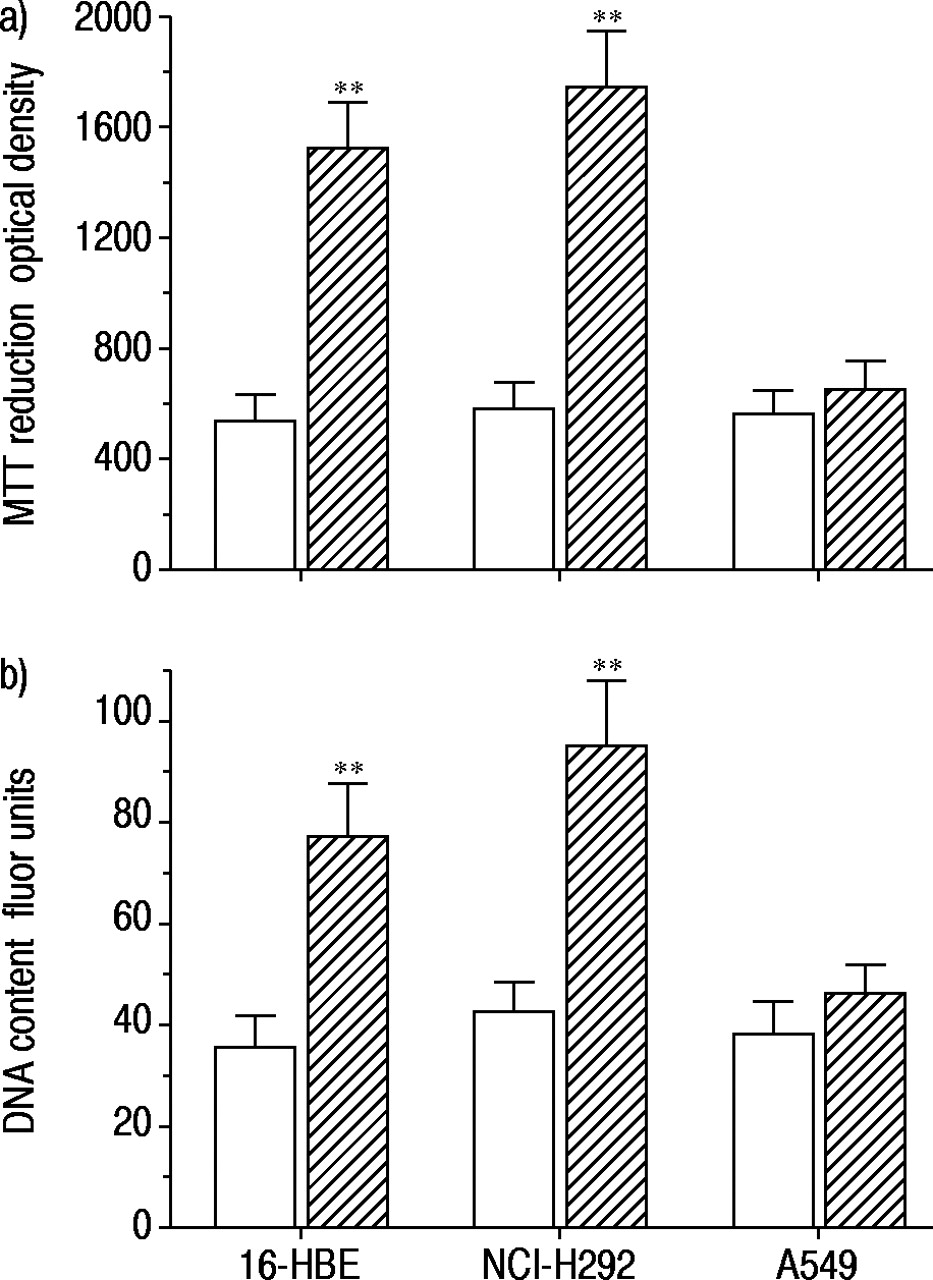
沙丁胺醇孵育(1×10-7 M) for 48 h in serum-free medium caused increases in both MTT reduction and intracellular DNA content in 16-HBE cells and NCI-H292 cells, but not in A549 cells (fig. 1⇓).因此,在上皮细胞系中,Salbutamol的生长促进作用不同,并且使用16-HBE细胞进行随后的实验。
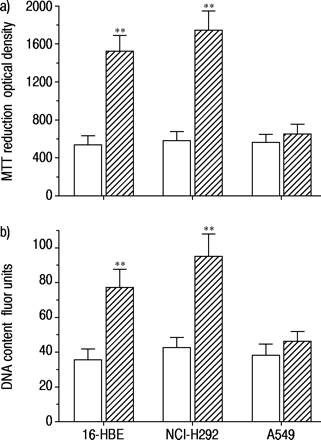
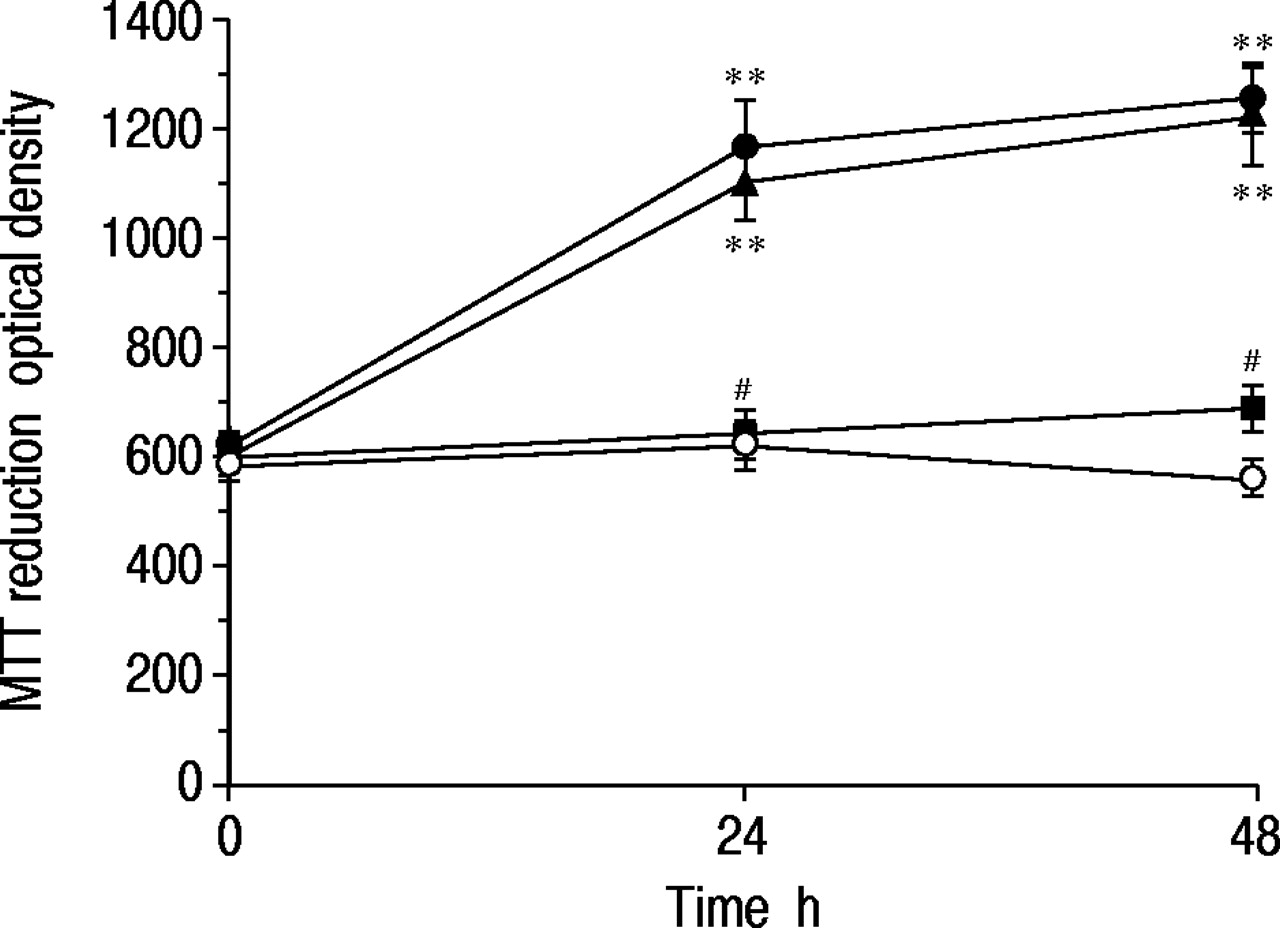
a)沙丁胺醇对3‐(4,5-二甲基噻唑-2 -基)-2,5-二苯四唑溴化(MTT)还原的影响;b) 16人支气管上皮(16-HBE)细胞、NCI-H292细胞和A549细胞内脱氧核糖核酸(DNA)含量的影响。数据分别用MTT法测定的光密度和用Hoechst 33258测定的荧光单元表示。在无血清培养基中培养72小时以抑制细胞生长后,沙丁胺醇(1×10-7 M, ),培养48 h。对照实验中未添加任何药物(□)。数据显示为平均值±sem;16-HBE细胞n=5,其他细胞n=6。**: p<0.01,与对照组差异显著。
),培养48 h。对照实验中未添加任何药物(□)。数据显示为平均值±sem;16-HBE细胞n=5,其他细胞n=6。**: p<0.01,与对照组差异显著。
如图2所示⇓而16-HBE细胞在Ham's F12培养基中培养72 h后,在没有FBS的情况下,48 h后在无血清培养基中没有生长。然而,添加沙丁胺醇(1×10-7与对照组(不含沙丁胺醇)相比,培养24 h时MTT值为197±20% (p<0.01, n=10),培养48 h时MTT值为233±27% (p<0.01, n=10)。沙丁胺醇对细胞生长的影响与10%胎牛血清相似。当细胞在含有沙丁胺醇(1×10)的无血清培养基中培养时-7M)加上心得安(1×10-5M)时,增长的响应完全消失。为了确定cAMP在上皮细胞增殖中的作用,我们测定了cAMP产生剂的作用。
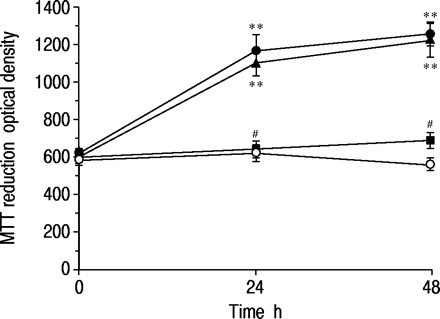
3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑鎓 - 溴化物(MTT)的时间过程响应Salbutamol(1×10)的16人支气管上皮(16-HBE)细胞的减少(1×10-7 M, •), salbutamol (1×10-7M)加上心得安(1×10-5▪),10%胎牛血清(▴)。在无血清培养基中培养72小时以抑制细胞生长后,加入每种药物,细胞培养至48小时。对照组不添加任何药物(〇)。数据显示为平均值±sem;每点N =10。**:P <0.01,与控制值显着不同;#:P <0.01,与单独的Salbutamol的相应值显着不同。
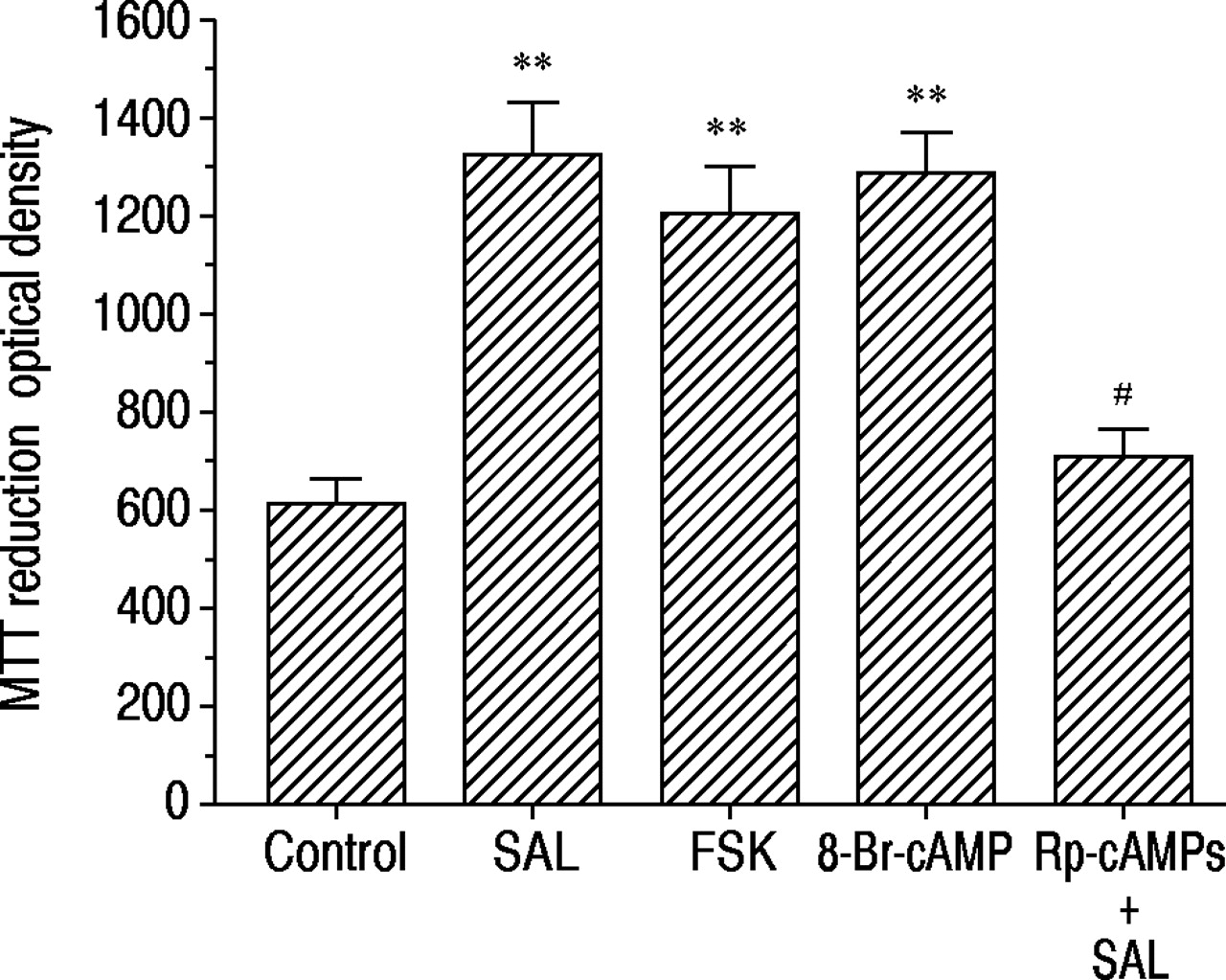
如图3所示⇓,用福斯柯林孵育48小时(1×10-7 M) or 8‐Br-cAMP (1×10-7 M) in the serum-free medium increased MTT reduction to the same degree as with salbutamol (1×10-7米)。通过与RP-CAMP的共同孵育(1×10,Salbutamol诱导的MTT降低的增加减少了86±10%(p <0.01,n = 8)(1×10-4米)。
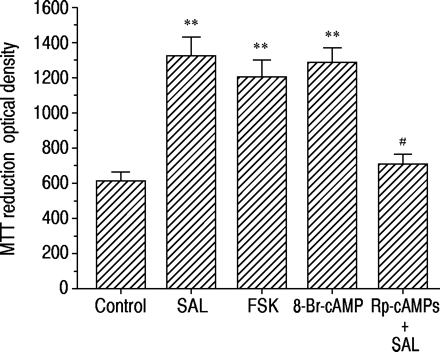
salbutamol (SAL), forskolin (FSK)和8 -溴腺苷3 ',5 ' -环单磷酸腺苷(8 - Br-cAMP)对3 -(4,5-二甲基噻唑-2 -基)-2,5-二苯四唑溴化(MTT)减少16人支气管上皮细胞(16-HBE)的影响,以及rp -腺苷3 ',5 ' -环单磷酸硫代三乙基铵盐(Rp-cAMPS)的影响(1×10)-4 M) on salbutamol-induced MTT reduction. The response was assessed 48 h after incubation in serum-free medium alone (control) or containing each agonist at 1×10-7 M. Data are shown as mean±sem; n=8 for each column. **: p<0.01, significantly different from control values;#: p<0.01,与单用沙丁胺醇有显著差异。
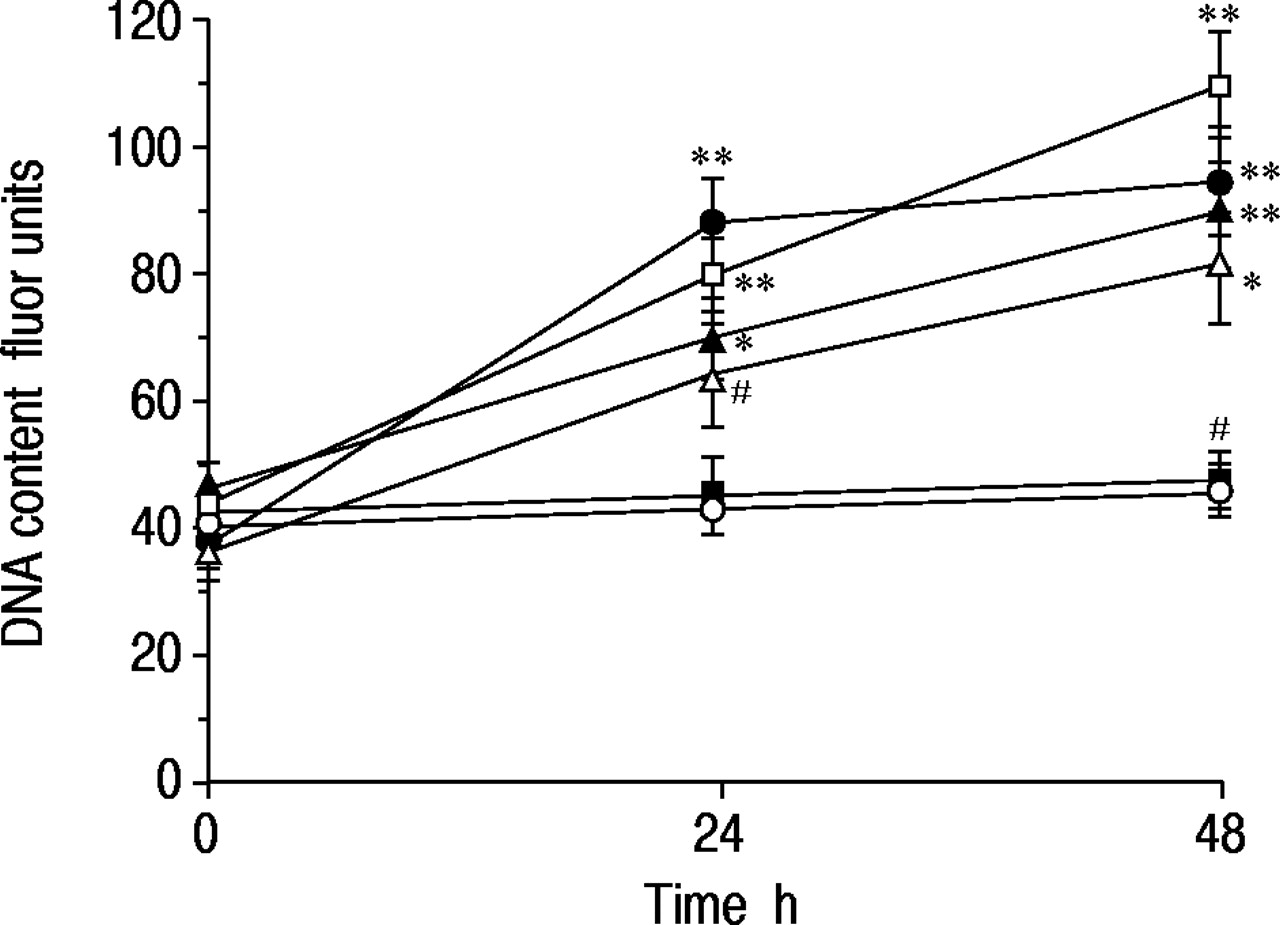
在48小时培养期间,在无血清培养基中没有在无血清培养基中改变16-HBE细胞的DNA含量,但在加入Salbutamol后,以时间依赖的方式增加(1×10-7 M) to the medium (fig. 4⇓).与MTT实验的发现一样,沙丁胺醇对DNA合成的影响与福斯考林、8‐Br-cAMP和10%胎牛血清一样有效,并且在普萘洛尔的存在下被消除。
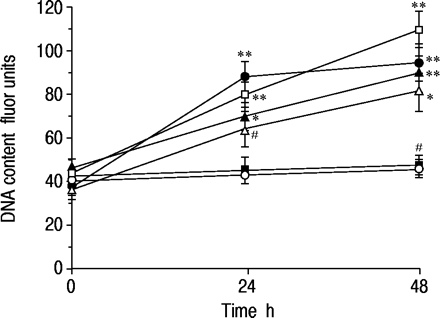
沙丁胺醇对16人支气管上皮细胞(16-HBE)脱氧核糖核酸(DNA)含量反应的时间过程(1×10-7 M, •), salbutamol (1×10-7M)加上心得安(1×10-5 M, ▪), forskolin (1×10-7M,□),8‐溴腺苷3 ',5 ' -环磷酸(8‐Br-cAMP) (1×10-7 M, ▵), and 10% foetal bovine serum (▴). After a 72‐h incubation in serum-free medium to arrest cell growth, each drug was added and the cells were cultured for up to 48 h. In the control experiment, no drug was added (○). Data are shown as mean±sem; n=10 for each point. *: p<0.05, **: p<0.01, significantly different from control values;#:P <0.01,与单独的Salbutamol的相应值显着不同。
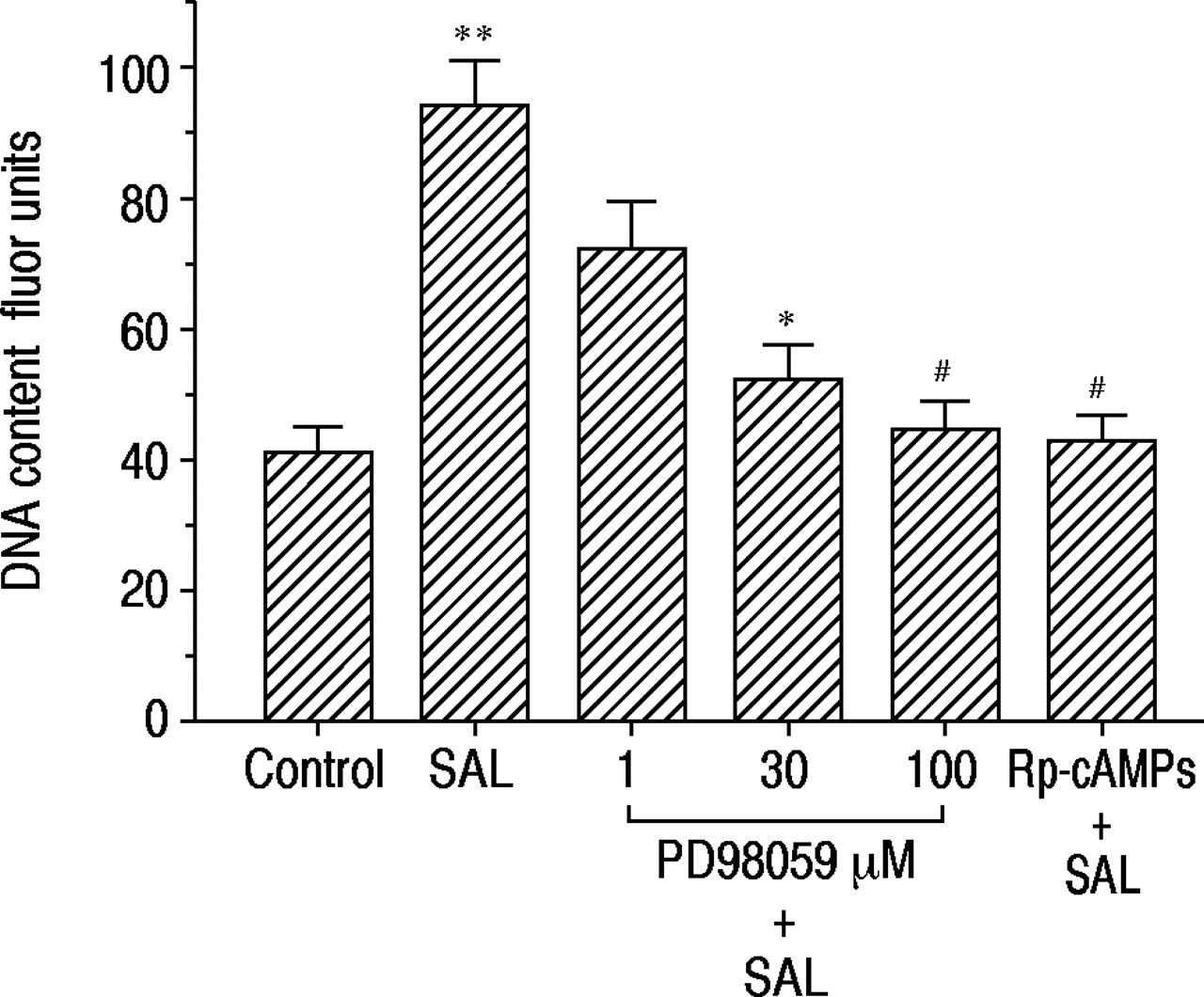
为了评估MAP激酶级联反应和cAMP在沙丁胺醇诱导的DNA合成中的作用,我们检测了MAP激酶激酶抑制剂PD98059和rp -cAMP的作用。如图5所示⇓PD98059和rp - camp以浓度依赖性方式抑制沙丁胺醇诱导的DNA含量增加。
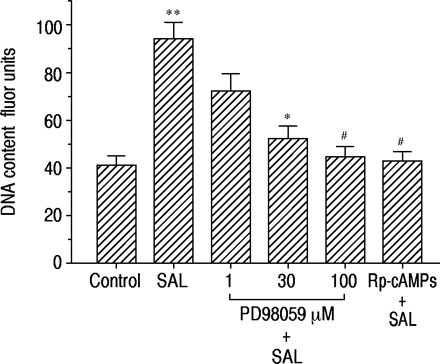
PD98059和RP-腺苷3',5'-环状二磷酸三甲酯三甲基铵盐(RP-CAMP)对16-人支气管上皮(16-HBE)细胞中的脱氧核糖核酸(DNA)合成的影响。在单独的无血清培养基中(对照)或含有沙丁醇莫酚(Sal,1×10)孵育后48小时测定DNA含量48小时。-7M)在没有和存在不同浓度的PD98059或1×10-4 M Rp-cAMPS. Data are shown as mean±sem; n=9 for each point. *: p<0.05,#: p<0.01,与单用沙丁胺醇差异有统计学意义;**: p<0.01,与对照组差异显著。
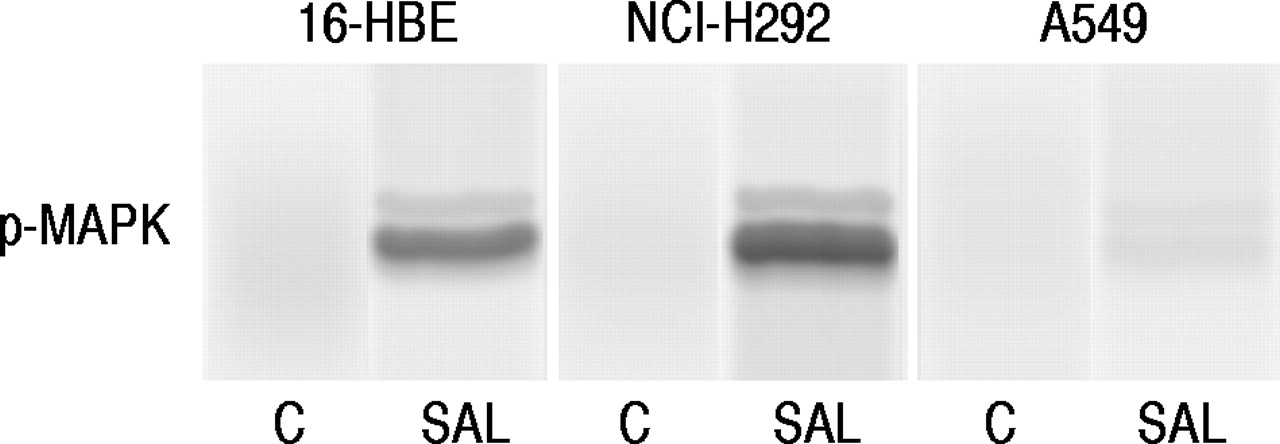
Western blot分析表明沙丁胺醇(1×10-7M)使MAP激酶在16-HBE细胞和NCI-H292细胞中磷酸化,但在A549细胞中影响不大(图6)⇓).
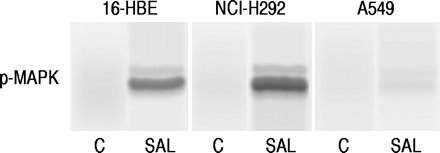
在5分钟暴露于单独的无血清培养基后,在16-人支气管上皮(16-HBE)细胞(NCI-H292细胞和A549细胞中,在16-人支气管上皮(16-HBE)细胞(NCI-H292细胞和A549细胞中)的蛋白质印迹(c)或含沙丁胺醇(SAL,1×10-7米)。
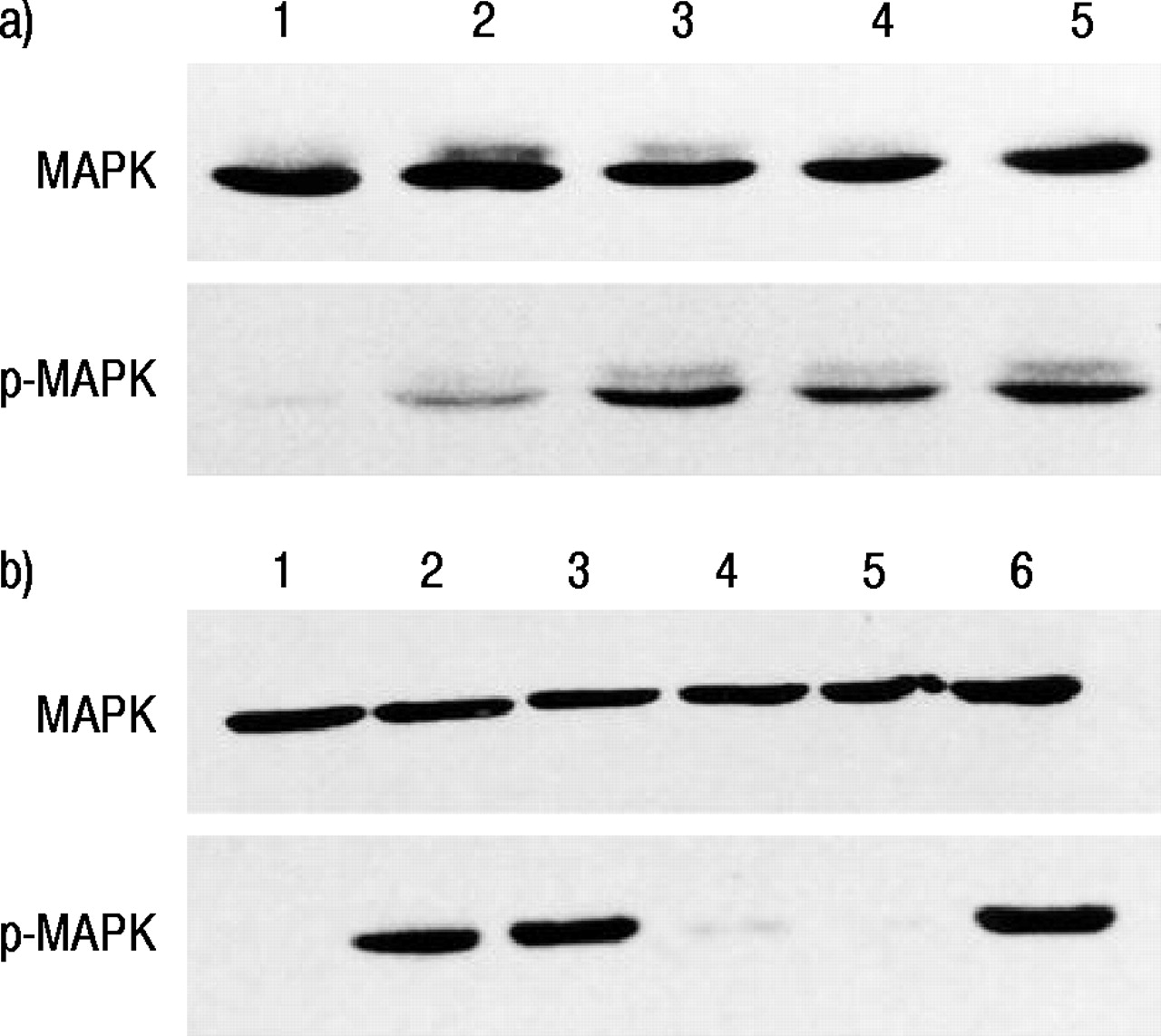
在16-HBE细胞中,MAP激酶被沙丁胺醇、福斯科林和8 - Br-cAMP诱导活化,沙丁胺醇诱导的磷酸化MAP激酶表达被普萘洛尔极大地抑制(图7)⇓).此外,通过PD98059和RP-CAMP抑制由Salbutamol产生的MAP激酶的磷酸化。
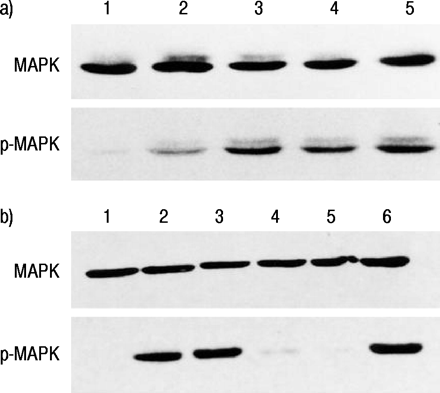
对16人支气管上皮细胞(16-HBE)中丝裂原活化蛋白(MAP)激酶(MAPK)和磷酸化MAP激酶(p‐MAPK)在无血清培养基(对照)或含各种药物暴露5分钟后进行Western blotting。a)环腺苷单磷酸(cAMP)生成剂和β -肾上腺素受体拮抗剂对沙丁胺醇诱导的MAPK磷酸化的影响。巷1:控制;第2道:沙丁胺醇+心得安;巷3:舒喘灵;巷4:forskolin;第五道:10%胎牛血清(FBS)。b) MAP激酶激酶抑制剂和cAMP拮抗剂对沙丁胺醇诱导的MAPK磷酸化的影响。巷1:控制;巷2:舒喘灵; lane 3: 8‐bromoadenosine 3′,5′-cyclic monophosphate (8‐Br-cAMP); lane 4: salbutamol plus PD98059; lane 5: salbutamol plus Rp-adenosine 3′,5′-cyclic monophosphorothioate triethylammonium salt (Rp-cAMPS); lane 6: 10% FBS.
讨论
本研究表明沙丁胺醇,β2- 肾上腺素能受体激动剂刺激人气道上皮细胞系的增殖。目前的作者发现,在16-HBE和NCI-H292细胞(但不是A549细胞)中,暴露于Sal丁醇酰胺,即使在FBS的培养基中,MTT降低和细胞内DNA含量也会导致MTT降低和细胞内DNA含量增加。被取代以抑制细胞生长。16-hbe细胞中的这种效果是通过刺激β的介导的2‐肾上腺素能受体,因为与普萘洛尔共孵育可抑制沙丁胺醇诱导的细胞生长和DNA合成。此外,cAMP生成剂,包括腺苷酸环化酶刺激剂、福斯考林和稳定的cAMP类似物8‐Br-cAMP,也显示出促进生长的效果。在本实验中,沙丁胺醇与其他药物的DNA合成时间过程不同;对沙丁胺醇的反应在前24小时达到平台期,而福斯柯林和8‐Br-cAMP似乎在整个48小时内诱导DNA含量的持续增加。这种差异的原因尚不清楚,但可能是由于β的脱敏2沙丁胺醇‐肾上腺素能受体。此外,作者发现沙丁胺醇诱导的MTT还原和DNA合成受到cAMP拮抗剂rp -cAMP的抑制。总之,cAMP的形成可能代表了某些气道上皮细胞系的有丝分裂信号。
cAMP在调节细胞生长中的作用是有争议的,可能是因为细胞类型和实验条件的不同。以往的研究表明,细胞内cAMP含量的升高可能与抑制人和豚鼠气道平滑肌细胞的生长有关13那14.相反,在大鼠腮腺细胞中15,狗甲状腺细胞16,老鼠心肌细胞17,已显示营地内含量的增加刺激细胞生长。此外,根据最近对耳蜗感觉上皮的研究,Forskolin和8-Br-camp刺激细胞生长,蛋白激酶A的抑制剂可以阻断的效果a9..这些发现与目前的结果一致,并提示camp依赖的信号通路在上皮细胞的增殖中具有刺激作用。但本实验仅采用气道上皮细胞株,不同细胞株间沙丁胺醇的作用不同。因此,这种信号通路是否也存在于人气道上皮原代培养中仍有待确定。
MAP激酶积极参与将细胞表面受体检测到的胞外刺激转化为胞内信号,导致细胞生长和分化所需基因的表达18.例如,有充足的证据表明GI和GQ蛋白偶联受体的刺激效果效果均激活映射激酶7.那8..关于Gs蛋白偶联受体刺激的作用,异丙肾上腺素已被证明可以激活和抑制心肌细胞的酶活性17和cos-7细胞19, 分别。在本研究中,在16-HBE和NCI-H292细胞中观察到磷酸化地图激酶的诱导,但与沙丁胺醇孵育后,不是A549细胞。在16-HBE细胞中,其他营养产生剂也引起了MAP激酶的激活,并且通过RP-CAMP取消了Salbutamol诱导的MAP激酶磷酸化。这些结果表明,在某些人气道上皮细胞中,可能存在由CAMP介导的GS诱导的刺激信号。
有丝裂丝糖苷活化蛋白激酶活化的主要途径通常需要RA,RAF-1和丝裂原活化蛋白激酶激酶的顺序活化20.那21.本文作者发现了丝裂原活化蛋白激酶的特异性抑制剂PD9805912,抑制沙丁胺醇诱导的丝裂原活化蛋白激酶磷酸化,并以浓度依赖性方式抑制沙丁胺醇诱导的脱氧核糖核酸合成。因此,丝裂原活化蛋白激酶激酶的活化可能参与了观察到的16人支气管上皮细胞的生长。然而,在丝裂原激活蛋白激酶级联反应中,环腺苷单磷酸依赖途径和定位于丝裂原激活蛋白激酶激酶近端的最初激活分子之间的联系仍然未知,需要进一步研究。
致谢
作者谨此感谢M. Shino和Y. Sugimura的技术援助。
- 收到了2001年8月17日。
- 公认2002年2月1日。
- ©ERS期刊有限公司










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}