





摘要
嗜酸性粒细胞作为免疫效应细胞,其主要功能是宿主防御蠕虫和其他微生物病原体的感染,这种持久的观点为关于嗜酸性粒细胞耗尽治疗的安全性的基本问题奠定了基础。如果在哮喘、慢性阻塞性肺病、慢性鼻窦炎伴鼻息肉、嗜酸性肉芽肿伴多血管炎和高嗜酸性粒细胞综合征等情况下,为了减轻组织嗜酸性粒细胞和嗜酸性粒细胞炎症的负面影响,嗜酸性粒细胞显著减少或完全减少,这些患者会容易感染或其他疾病吗?嗜酸性粒细胞谱系被去除的小鼠模型的开发、对天生缺乏嗜酸性粒细胞的患者的观察以及嗜酸性粒细胞消耗药物疗法的研究数据表明,嗜酸性粒细胞的缺乏对健康无害。如本综述所述,迄今为止可获得的证据支持以下结论:即使嗜酸性粒细胞的某些稳态作用可能在受控动物模型和人类中得到证明在体外在此背景下,人类物种的进化似乎提供了足够的免疫冗余,这样就可以在没有嗜酸性粒细胞的情况下身体健康。
摘要
本文综述了小鼠模型和人体临床试验中嗜酸性粒细胞耗竭的累积证据,证实靶向嗜酸性粒细胞治疗嗜酸性粒细胞相关疾病具有治疗价值,而不会对健康产生有害影响https://bit.ly/3biObDI
简介
在模型系统和药理学干预的帮助下,随着科学发现揭示了迄今为止未知的健康和疾病功能,嗜酸性粒细胞正在经历一段复兴时期。简单来说,嗜酸性粒细胞是由表达白细胞介素(IL)-5受体α (IL- 5r α)的祖细胞(图1) [1- - - - - -3.].分化、成熟和从骨髓起始位点迁移的过程涉及一系列特定的转录因子和细胞因子,其中IL-5起着重要作用。一旦进入循环,嗜酸性粒细胞具有有限的停留时间(半衰期8-18小时),它们被趋化因子如eotaxin-1, -2和-3(分别为趋化因子(CC motif)配体CCL11, CCL24和CCL26)招募到外周组织,在那里它们大约存活2-5天[4].虽然关于嗜酸性粒细胞基本生物学和围绕嗜酸性粒细胞发展的过程有很多了解,但居住在外周组织中的嗜酸性粒细胞的作用一直是一个开放的问题和争论的主题。
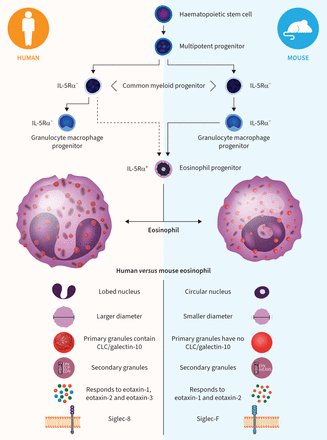
人和小鼠嗜酸性粒细胞的发展及特征[1- - - - - -3.].已经注意到人类成熟嗜酸性粒细胞之间的一些差异与老鼠。人类嗜酸性粒细胞比小鼠嗜酸性粒细胞大,嗜酸性粒细胞染色更强烈,细胞核呈裂片状而不是圆形。人(而非小鼠)嗜酸性粒细胞中的初级颗粒含有Charcot-Leyden晶体(CLC)蛋白/半乳糖凝集素-10。人和小鼠嗜酸性粒细胞次生颗粒的致密核心主要由主要碱性蛋白-1 (MBP-1)组成。人次生颗粒中嗜酸性粒细胞相关rna酶包括嗜酸性粒细胞蛋白X (EPX)、嗜酸性粒细胞阳离子蛋白(ECP)和嗜酸性粒细胞源性神经毒素(EDN)。嗜酸性粒细胞过氧化物酶在人和小鼠嗜酸性粒细胞中都储存在次级颗粒中。人类嗜酸性粒细胞对eotaxin-1 (CCL11), -2 (CCL24)和-3 (CCL26)有反应,而小鼠嗜酸性粒细胞对eotaxin-1和-2有反应。在细胞表面分子差异中,人嗜酸性粒细胞表达Siglec-8,小鼠嗜酸性粒细胞表达Siglec-F。CCL:趋化因子(CC motif)配体;IL-5Rα:白细胞介素5受体α; mEARS: mouse eosinophil associated RNases. Development schematic adapted from Lee等.[1允许。
传统观点认为嗜酸性粒细胞的主要作用是宿主对蠕虫感染的防御[5].嗜酸性粒细胞被认为是在抗原特异性2型(T2)免疫反应诱导的信号级联反应中迁移到感染部位。嗜酸性粒细胞直接与寄生虫相互作用通过酶解作用:一种抗体介导的过程,导致细胞毒性物质(包括颗粒蛋白)的释放,破坏细胞膜并增加其渗透性[6].嗜酸性粒细胞还通过不同于对蠕虫反应的机制参与宿主对病毒、细菌和真菌病原体的防御(在[7- - - - - -9])。然而,正如我们开始理解的那样,对抗感染只是嗜酸性粒细胞故事的一个方面。
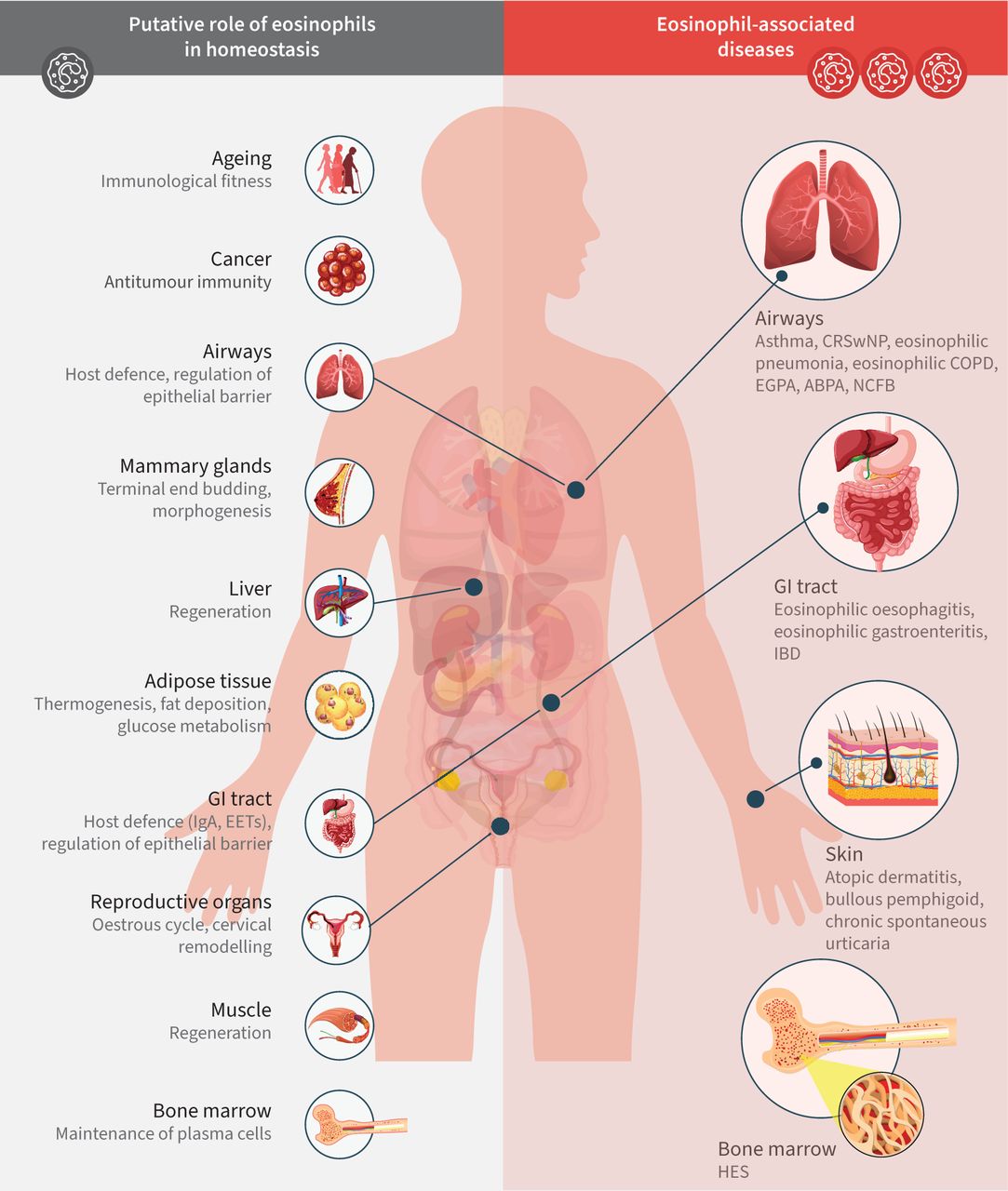
嗜酸性粒细胞遍布全身,主要分布于消化道粘膜,但也分布于脂肪组织、气道、乳腺、胸腺和子宫[10],在缺乏感染挑战的情况下,增加了嗜酸性粒细胞参与宿主防御以外活动的可能性,即。维持体内平衡(图2) [11].嗜酸性粒细胞存在于外周组织的功能意义得到嗜酸性粒细胞缺乏小鼠的支持,在这些小鼠中观察到肠道粘液屏障的缺陷、微生物组的改变和Peyer's斑块大小的差异[12- - - - - -14].对嗜酸性粒细胞缺乏小鼠的研究也表明嗜酸性粒细胞在调节脂肪组织内稳态中的作用[15,16].在小鼠模型中,脂肪组织中嗜酸性粒细胞升高表面上有利于代谢健康,导致产热性和葡萄糖敏感性增强,并减少体重增加,而在稳态中缺乏嗜酸性粒细胞则与代谢健康的负面影响相关,例如产热性脂肪的产生减少[3.,17].然而,有关嗜酸性粒细胞在人体脂肪组织和稳态中的作用的数据尚不清楚[18- - - - - -20.].嗜酸性粒细胞改变的时间和方法的差异可以解释这些相互矛盾的结果。迄今为止,没有证据表明人类嗜酸性粒细胞减少会导致胃肠道问题、脂肪组织变化、体重变化或其他稳态失调[3.,21].
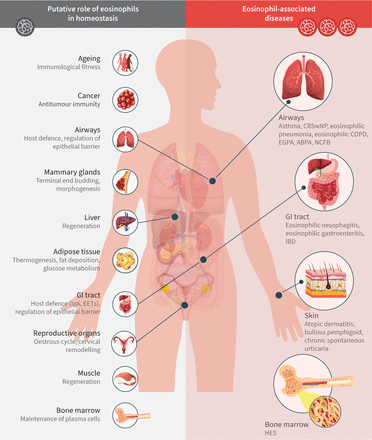
嗜酸性粒细胞在稳态和嗜酸性粒细胞相关疾病中的假定作用。图中所示的嗜酸性粒细胞的稳态作用得到了动物模型和人类数据的支持在体外研究。ABPA:过敏性支气管肺曲菌病;CRSwNP:慢性鼻窦炎伴鼻息肉;EET:细胞外陷阱;EGPA:嗜酸性肉芽肿伴多血管炎;GI:肠胃;HES:高嗜酸性粒细胞综合征;IBD:炎症性肠病;非囊性纤维化支气管扩张。改编自Jackson和Munitz[11].
临床社区在使用抑制嗜酸性粒细胞数量的治疗方法时特别关注的是观察各种肿瘤类型中嗜酸性粒细胞的浸润。目前尚不清楚这一发现对宿主是有利的、不利的还是中性的,而且可能因癌症类型而异。
与嗜酸性粒细胞的保护和稳态功能相反,它们参与疾病的发病机制。嗜酸性粒细胞长期以来一直与功能双重性有关:对寄生虫感染发挥保护作用,但对过敏性疾病的病理有贡献[22,23].作为效应细胞,嗜酸性粒细胞对某些过敏原、蠕虫或其他微生物病原体的抗原引发的免疫级联激活作出反应。由此产生的嗜酸性粒细胞脱肉芽会释放细胞毒性物质,这些物质被认为有利于对抗病原体,但会损害周围的健康组织。这种损害作用是组织特异性的。例如,下气道上皮剥蚀和粘液堵塞在功能上最为重要[24以及上呼吸道的组织水肿。与感染源无关的外周或组织特异性嗜酸性粒细胞增多症和/或嗜酸性粒细胞增多症见于哮喘、慢性阻塞性肺病、慢性鼻窦炎伴鼻息肉(CRSwNP)、嗜酸性肉芽肿伴多血管炎(EGPA)和高嗜酸性粒细胞增多综合征(HES) (图2) [22,23,25,26].由于嗜酸性粒细胞被认为与这些情况下观察到的组织损伤和疾病表现有直接的因果关系,嗜酸性粒细胞耗竭是一种合理的治疗干预手段。事实上,临床试验已经在多种疾病中产生了积极的结果,这些疾病的发病机制被认为与嗜酸性免疫功能障碍有关(表1) [27- - - - - -47]和几种嗜酸性粒细胞靶向疗法已获批准用于临床[48,49].
早期的研究表明,嗜酸性粒细胞具有不同的表型,在小鼠模型的肺中发现了常驻嗜酸性粒细胞和炎症性嗜酸性粒细胞[50].针对IL-5的生物制剂被认为可以靶向并消耗炎性嗜酸性粒细胞,同时保留常驻嗜酸性粒细胞。来自最近一项针对过敏原挑战小鼠的研究的证据与之前的研究相矛盾,表明抗il -5治疗可以减少所有嗜酸性粒细胞的肺种群,而不管其表型如何[51].鉴于此,这些新的数据也表明嗜酸性表型可能存在于一个连续体上,而不是作为不同的细胞亚型存在。
专注于嗜酸性粒细胞的致病方面,忽略了它们假定的保护作用,并提出了一个重要的问题:嗜酸性粒细胞衰竭的潜在好处会被功能丧失所抵消吗?在这篇综述中,我们通过深入研究嗜酸性粒细胞耗竭影响的当前知识状态来解决这个问题,从嗜酸性粒细胞谱系消融的小鼠模型中获得见解,与嗜酸性粒细胞缺失相关的自然发生的情况,以及来自药物治疗诱导的人类嗜酸性粒细胞减少/耗竭的临床数据。尽管嗜酸性粒细胞涉及健康和稳态的多个方面,但为了确定嗜酸性粒细胞减少的益处和风险,我们主要关注感染和恶性肿瘤,因为这些领域存在临床问题,而且缺乏任何直接的人类数据支持嗜酸性粒细胞在生理学的其他方面发挥关键作用。
方法
通过查询MEDLINE数据库,对2000年至2021年发表的使用术语“嗜酸性粒细胞耗尽”、“嗜酸性粒细胞谱系消除”或“嗜酸性粒细胞”和“PHIL”、“ΔdblGATA”、“gada -1缺失”、“benralizumab”、“mepolizumab”、“reslizumab”、“dexpramipexole”、“伊马替尼”、“糖皮质激素”或“系统性皮质类固醇”的英语文章进行了相关研究。通过搜索“benralizumab”、“mepolizumab”或“reslizumab”,并结合“长期”、“安全”、“感染”、“寄生虫”、“恶性肿瘤”、“癌症”、“2019冠状病毒病(COVID-19)”或“怀孕”,可以确定描述嗜酸粒细胞耗尽疗法安全性的出版物。本综述根据主题相关性选择了相关研究进行讨论。通过搜索策略确定的文章所引用的出版物酌情列入。
预测嗜酸性粒细胞耗竭的后果
寄生虫和机会性感染
对嗜酸性粒细胞耗竭的关注的基础是基于这些细胞在对抗寄生虫和其他病原体感染方面的假定功能。对嗜酸性粒细胞重要性的支持来自于对血液和组织嗜酸性粒细胞计数增加的观察,以应对蠕虫感染[5].来自在体外研究和动物模型也表明嗜酸性粒细胞具有抗病毒特性,但这些发现对人类免疫反应的适用性和相关性尚未确定[52].尽管如此,如果嗜酸性粒细胞的主要功能是宿主对寄生虫和其他感染的防御,减少或耗尽嗜酸性粒细胞将增加感染风险。
除了机制的基本原理之外,当通过传染病风险的镜头观察时,有临床理由对治疗性嗜酸性粒细胞耗竭持怀疑态度。机会性感染卡式肺肺炎、带状疱疹和结核病是已知与糖皮质激素相关的危害[53],一种影响嗜酸性粒细胞生成、存活和功能的药物[54].糖皮质激素还与过度感染综合征的发展有关,这是一种可能危及生命的情况,在这种情况下,蠕虫自身感染加速,导致蠕虫负担过重[55].然而,当讨论与糖皮质激素相关的免疫功能和感染风险时,一个重要的警告是嗜酸性粒细胞抑制只是其广泛免疫抑制作用的一个方面[53].糖皮质激素改变多种免疫细胞类型的分化、激活和/或功能,包括嗜酸性粒细胞、巨噬细胞、中性粒细胞、淋巴细胞和树突状细胞[53].因此,关于糖皮质激素感染风险的观察不能仅仅归因于它们对嗜酸性粒细胞的作用。
比较和对比小鼠和人类嗜酸性粒细胞
小鼠和人类嗜酸性粒细胞有许多共同特征,包括从造血干细胞到IL-5Rα的相似发育途径+嗜酸性粒细胞祖细胞和最终成熟嗜酸性粒细胞[1- - - - - -3.].在小鼠中,粒细胞-巨噬细胞祖细胞(GCPs)成为嗜酸性粒细胞祖细胞,而在人类中,GCPs不分化为嗜酸性粒细胞。虽然成熟的人类嗜酸性粒细胞和小鼠嗜酸性粒细胞在结构和外观上大体相似,但仍有一些显著差异(图1).这些差异是否与嗜酸性粒细胞在健康或疾病中的行为有关,或在嗜酸性粒细胞通过对其他免疫介质的作用而耗损的情况下重要,仍有待确定。然而,我们有理由假设,本文所描述的至少一些与物种相关的差异,支持了为什么一些实验小鼠模型预测的嗜酸性粒细胞耗竭的理论后果没有在人类中观察到。
嗜酸性粒细胞耗竭的小鼠模型
到目前为止,由于在21世纪初创建了两个小鼠模型ΔdblGATA和PHIL,对嗜酸性粒细胞在健康和疾病中的作用有了未知的认识[59,60].在ΔdblGATA和PHIL小鼠的总体健康状况中,以及在各种实验条件下对这些动物的后续研究中,都观察到了嗜酸性粒细胞耗竭的后果(或缺乏)。表2) [59- - - - - -77].
第一个模型ΔdblGATA是通过删除位于GATA-1(红系转录因子)启动子区域的高亲和力双GATA结合位点而生成的,该位点被认为参与GATA-1表达的自动调节[59].GATA-1是一种在几种造血细胞系中表达的转录因子,包括红细胞、巨核细胞、肥大细胞和嗜酸性粒细胞[78,79].在体外研究表明,GATA-1的表达水平与谱系规格有关:低浓度的GATA-1蛋白不能在禽骨髓单核细胞细胞系中诱导成熟的表型,而中等浓度的GATA-1蛋白可导致嗜酸性粒细胞的形成,较高浓度的GATA-1蛋白可产生成血栓细胞和可能的成红细胞细胞[78].在一个单独的转基因小鼠模型中观察到,发育过程中完全缺乏GATA-1会导致胚胎死亡[79].纯合子(雌性)/半合子(雄性)ΔdblGATA小鼠缺乏嗜酸性粒细胞,但红细胞、巨核细胞和肥大细胞不受影响或仅受到轻微扰动[59].ΔdblGATA小鼠是可行的和可育的,没有明显的缺陷,除了缺乏嗜酸性粒细胞。
PHIL小鼠是通过引入一种转基因结构来创建的,其中嗜酸性粒细胞过氧化物酶启动子用于驱动白喉毒素a的表达,导致嗜酸性粒细胞谱系性细胞死亡[60].PHIL小鼠缺乏嗜酸性粒细胞,但没有其他造血来源的细胞。就像ΔdblGATA小鼠一样,PHIL小鼠是可繁殖的,没有明显的健康问题,寿命正常。这些小鼠的过敏原挑战表明,嗜酸性粒细胞有助于肺粘液积聚,是气道高反应性所必需的,这两者都是哮喘的典型特征。
嗜酸性粒细胞耗竭
嗜酸性粒细胞减少的后果可以通过两种途径来研究:1)通过评估自然发生的遗传性或后天嗜酸性粒细胞缺乏病例的影响,或2)通过减少/减少嗜酸性粒细胞计数通过医学治疗。自然发生的嗜酸性粒细胞缺乏在人类中并不常见[80].据Gleich等.[80],通常发生在其他免疫系统缺陷的情况下(如。嗜碱性粒细胞缺失)和/或γ球蛋白血症。因此,在这些患者中可以得出的关于嗜酸性粒细胞减少的结论受到其他免疫系统异常共存的限制。少数患者有过敏性疾病及嗜酸性粒细胞缺失,但无其他免疫系统异常的证据[80].这些患者的临床病程在大多数情况下是不显着的,除了过敏性疾病的存在。
多种目前可用的药物治疗可减少血液和组织中的嗜酸性粒细胞计数。这些药物包括抗il -5单克隆抗体mepolizumab和reslizumab,抗il - 5r单克隆抗体benralizumab,酪氨酸激酶抑制剂伊马替尼,口服治疗右普拉克索和皮质类固醇。嗜酸性粒细胞的减少程度因药剂而异,与它们作用机制的差异一致(表1).使用右普拉克索和伊马替尼减少嗜酸性粒细胞取决于个体,一些患者表现出高水平的嗜酸性粒细胞抑制,而另一些患者仅表现出中等或中等效果[43- - - - - -45].非专为嗜酸性粒细胞设计的疗法也容易影响多种免疫细胞类型,如降低嗜碱性粒细胞计数,在某些情况下,右普拉克索可降低中性粒细胞计数[81]或糖皮质激素对巨噬细胞、中性粒细胞、淋巴细胞和树突状细胞的分化、激活、凋亡和/或功能的改变[53].
总的来说,抗il- 5r α单抗benralizumab的嗜酸性粒细胞特异性耗损最为迅速和深刻[27,28].Benralizumab已在哮喘患者中进行了研究[82- - - - - -86], copd [29,87], CRSwNP [30.], egpa [31]和HES [32],积累了数千个患者年的暴露数据。在临床试验和实际研究中,使用贝那利珠单抗治疗可导致血液嗜酸性粒细胞完全或接近完全耗尽[27- - - - - -32,88,89].直接比较严重嗜酸性哮喘患者和基线血嗜酸性粒细胞计数>300个细胞·µL−1,以达到血液嗜酸性粒细胞计数减少50%的时间来衡量,贝那利珠单抗起效迅速(1.7±0.7 h),与强的松(2.5±0.3 h)相似,但明显快于美波珠单抗(25.8±14.3 h;p < 0.001) (28].治疗后30天,血嗜酸性粒细胞几何平均值为8±2.8个细胞·µL−1为92±1.7细胞·µL−1在mepolizumab治疗组中(治疗组间比较p=0.002)。Benralizumab也显著减少了组织嗜酸性粒细胞[27,32,33].在嗜酸性哮喘成人患者中,贝那利珠单抗可使气道嗜酸性粒细胞计数中位数降低95.8% [27].在一项针对HES和胃肠道组织嗜酸性粒细胞增多症患者的研究中,使用benralizumab治疗后,胃肠道各节段的嗜酸性粒细胞完全消失[33].鉴于这些观察结果,接受benralizumab治疗的患者是关于嗜酸性粒细胞减少对人类影响的有价值的数据来源。
嗜酸性粒细胞减少对寄生虫感染易感性的影响
来自小鼠模型的证据
PHIL和ΔdblGATA小鼠的易感性和免疫反应已经使用各种蠕虫进行了测试,包括那些自然感染啮齿动物的蠕虫(例:旋毛虫)和没有的(例:stercoralis圆线虫) (表2).总的来说,缺乏嗜酸性粒细胞的小鼠对蠕虫感染表现出适当的原发性和继发性反应。在ΔdblGATA和PHIL小鼠中嗜酸性粒细胞的缺失对蠕虫感染的测量没有影响曼氏裂体吸虫[61].虫量和虫卵沉积美国曼感染嗜酸性粒细胞谱系切除小鼠与野生型对照小鼠相似。肝脏肉芽肿的形成仅在细胞类型组成上有所不同,在ΔdblGATA和PHIL小鼠中明显缺乏嗜酸性粒细胞。同样,PHIL小鼠和野生型对照组的初级和次级免疫反应具有可比性美国stercoralis[62].然而,在中性粒细胞消耗抗体处理的PHIL小鼠中,幼虫的杀灭受到阻碍[62].
这些发现表明嗜酸性粒细胞和中性粒细胞对蠕虫感染的反应具有内在的冗余性[90].蠕虫是众多病原体之一,可能是免疫系统多样化的驱动因素,这是在感染中生存的重要组成部分。在组织中,蠕虫可能容易受到各种先天免疫应答者的攻击,包括中性粒细胞、嗜酸性粒细胞、嗜碱性粒细胞、巨噬细胞和血小板,每种免疫细胞类型的作用取决于所涉及的组织类型和蠕虫种类。这可能会造成一定程度的免疫冗余。一般来说,嗜酸性粒细胞似乎有助于和增强免疫反应,而不是作为中心成分,因此不是杀灭蠕虫幼虫所绝对需要的。
对感染的反应与象皮病马来,与野生型对照相比,PHIL小鼠在初次感染期间微丝蚴存活时间更长,寄生虫诱导的IgE反应增强,杯状细胞粘液分泌增加,但PHIL小鼠的气道生理变化减少[63].ΔdblGATA小鼠感染Nippostrongylus取代巴西橡胶树与野生型对照相比,有更大的蠕虫负担和卵子沉积,对继发肺部感染的抵抗力较低;然而,初次感染和继发感染的蠕虫排出不受影响[64].同样,在ΔdblGATA小鼠中,成虫清除率未受影响t . spiralis感染,但与野生型对照相比,骨骼肌对幼虫积累的抵抗力降低[65].在ΔdblGATA感染的小鼠中也报告了更大的微丝血症和更高的成虫计数Litomosoides倍与野生型对照相比[66].
有趣的是,来自ΔdblGATA和PHIL小鼠的实验数据表明,嗜酸性粒细胞可能支持寄生虫的生长和生存[67,68].Fabre等.[67报告说t . spiralis与野生型对照相比,在ΔdblGATA和PHIL小鼠感染后15 - 22天内,浸润骨骼肌细胞的幼虫大量死亡,这表明嗜酸性粒细胞是蠕虫存活和维持慢性感染所必需的。在后续研究中,Gebreselassie等.[68]观察到嗜酸性粒细胞从t . spiralis-感染的IL-5转基因小鼠(过度表达IL-5)t . spiralis-感染ΔdblGATA和PHIL小鼠改善了幼虫的生长和存活。这些数据与传统观点形成鲜明对比,传统观点认为嗜酸性粒细胞对抗而不是促进蠕虫感染,这可能表明某些蠕虫已经进化到利用它们促进的嗜酸性环境作为一种生存机制。
对这些结果的解释必须考虑到模型系统的注意事项。首先,用于评估嗜酸性粒细胞减少小鼠免疫反应的蠕虫并不都是通常感染啮齿动物的病原体。例如,老鼠不是天然的宿主美国stercoralis尽管这种寄生虫可以感染小鼠,但它并没有完成正常的感染周期[91].值得注意的是,嗜酸性粒细胞似乎支持蠕虫生长和生存的实验确实涉及一种天然的啮齿动物病原体(t . spiralis).其次,菌株背景效应解释了ΔdblGATA小鼠和PHIL小鼠之间的一些发现差异[92],并提示遗传变异对感染反应的影响。第三,令科学界感到沮丧的是,在老鼠身上观察到的结果并不总是在人类身上得到复制。
来自临床试验的数据
迄今为止,专门减少嗜酸性粒细胞计数的生物疗法与蠕虫感染的增加没有关联(表3) [48,82,85,93- - - - - -99].在benralizumab、mepolizumab和reslizumab的安慰剂对照和长期研究中,未报告蠕虫感染病例。考虑到影响程度,贝那利珠单抗可完全或接近完全耗尽循环嗜酸性粒细胞[27- - - - - -32,提供了最好的机会来检查嗜酸性粒细胞耗竭在人类中的安全性。在患者人群中,研究表明,使用benralizumab治疗不会增加寄生虫感染或机会性感染的风险,即使在寄生虫感染更常见的地理区域,如南美、非洲和东南亚[48].事实上,到目前为止,文献中还没有报道过一例benralizumab治疗期间的蠕虫感染病例。然而,事实是,过度感染综合征是一个公认的并发症,全身皮质类固醇使用的个体感染美国stercoralis[One hundred.表明广泛抑制T2炎症有能力显著影响抗蠕虫免疫,即使更有针对性的抗嗜酸性粒细胞治疗没有。有趣的是,唯一一项抗t2生物制剂的3期试验是在儿童中重度哮喘的情况下使用抗il - 4r单抗dupilumab,该试验强调了寄生虫感染率的数值增加。在本试验中,dupilumab组记录了7例寄生虫感染(2.6%),而安慰剂组为0例[101].在机制上,这与实验感染IL-4/ il -13缺陷小鼠的结果一致n取代巴西橡胶树,显示出蠕虫排出能力受损,并且发现先天细胞产生的IL-4/IL-13是在这种情况下对抗感染的有效T2免疫反应所必需和足够的[102].
嗜酸性粒细胞减少对非蠕虫感染易感性的影响
小鼠模型研究
多项研究评估了ΔdblGATA和PHIL小鼠对病毒、原生动物和细菌的反应(表2).病毒和原生动物病原体的结果通常显示,嗜酸性粒细胞减少小鼠与野生型对照之间没有差异[69- - - - - -71].关于嗜酸性粒细胞减少对细菌定植的影响,有报道称嗜酸性粒细胞缺乏小鼠的胃肠道微生物群组成与野生型对照存在差异[12- - - - - -14].这一发现的意义尚不清楚,因为这些差异与对健康的有害影响无关。14].在实验感染模型中使用幽门螺杆菌或枸橼酸杆菌属rodentium,与野生型对照相比,PHIL小鼠表现出更强的炎症和t辅助因子1反应,然而嗜酸性粒细胞耗损是否增加或减少胃肠道定植在细菌病原体之间存在差异[72].当嗜酸性粒细胞缺乏小鼠和野生型对照受到挑战时,没有观察到定殖的差异博代氏杆菌属bronchiseptica[73].总的来说,稀疏和不一致的数据不足以支持嗜酸性粒细胞在宿主防御病毒、原生动物和细菌病原体中的关键作用。
嗜酸性粒细胞缺乏小鼠对真菌感染的反应已在支气管肺曲霉菌病的背景下被广泛研究。在侵袭性曲霉病的嗜中性粒细胞减少小鼠模型中,嗜酸性粒细胞缺乏小鼠与野生型小鼠相比,真菌负担和死亡率降低[74].随后的一项研究表明,在嗜酸性粒细胞缺乏的中性粒细胞减少小鼠中,卡普菌素诱导的肺曲霉菌病不那么严重,真菌负担也较低与野生型对照,表明嗜酸性粒细胞在这种情况下不利于真菌清除[75].在非嗜中性粒细胞减少小鼠中,嗜酸性粒细胞缺乏与鼻内接种的T2炎症反应减弱有关来自烟曲霉属真菌[76,77].这些数据表明嗜酸性粒细胞参与了对真菌感染的反应,但它们的缺失实际上可能是有益的。
来自临床试验的数据
嗜酸性粒细胞消耗、抗il- 5/ 5r疗法与非蠕虫感染(包括原生动物、细菌或真菌病原体)的风险增加无关(表3).在哮喘患者中,由于使用吸入性皮质类固醇,肺炎风险已经增加,在长期研究中,肺炎发病率很低[48,82,93- - - - - -97,99].此外,来自MELTEMI长期扩展及其前身研究的综合分析数据显示,更大的贝那利单抗暴露不会增加严重感染风险[85].
然而,有相互矛盾的数据表明嗜酸性粒细胞可能在宿主抗病毒反应中起作用。MATERIAL研究评估了接受mepolizumab或安慰剂的轻度哮喘患者(n=37)的实验性鼻病毒诱导免疫反应,报告了mepolizumab治疗的患者感染后第7天病毒载量增加[103].尽管推断存在抗il -5驱动的限制病毒复制机制的损伤,但研究人员没有在接种后的任何较早时间测量病毒载量。重要的是,已知鼻病毒复制在哮喘患者接种后的第2-3天达到峰值,这使得孤立的第7天测量非常难以解释[104].值得注意的是,尽管可以合理地假设大多数个体每年经历一次或多次上呼吸道病毒感染(哮喘加重的主要原因),但综合MELTEMI分析显示,在接受贝那利单抗的患者中,≥75%的患者每年无哮喘加重,59%的患者在整个综合分析期间(长达5年)无哮喘加重。这些观察结果与模式识别受体toll样受体7(已知在感知鼻病毒中很重要)被嗜酸性炎症抑制,导致抗病毒干扰素反应受损的发现相一致[105].
有趣的是,有数据表明,哮喘患者,尤其是重症哮喘患者的嗜酸性粒细胞,与呼吸道病毒的结合和灭活能力较差[106];因此,在这种情况下,来自这些患者的嗜酸性粒细胞可能在宿主防御中发挥较小的作用。然而,由于哮喘患者中的嗜酸性粒细胞已经被启动,它们可能具有增强的细胞毒性作用,从而导致病毒诱导的哮喘加重[106].
在机会性感染中,带状疱疹是唯一在抗il -5/ 5r治疗的临床试验中特别注意到的病原体(表3).带状疱疹感染并不常见(≤2%),机会性感染的风险似乎并没有随着治疗时间的推移而增加[48,82,85,93- - - - - -99].在贝那利珠单抗临床试验中,未观察到机会性感染风险增加[48].3期数据分析显示,接受benralizumab治疗的患者病毒感染率相似与安慰剂(48,87].虽然在一个病例报告中描述了benralizumab治疗后的弥散性带状疱疹感染[107],在临床试验中未观察到感染率增加[48,85,94]或上市后监察[108].病例报告中描述的患者还接受泼尼松龙(未指明剂量)和环磷酰胺[107],这将有助于感染易感性。
生物疗法与COVID-19
在COVID-19大流行早期,住院患者中报告了血液中嗜酸性粒细胞减少的情况。这一点的意义一直存在争议;在COVID-19大流行期间,是否维持嗜酸性粒细胞消耗疗法是临床社区使用这些疗法的最初讨论主题[109].低发病率患者入院时出现COVID-19体征和症状的可能性更大,住院时间和病程更长与正常的嗜酸性粒细胞计数标志着嗜酸性粒细胞减少作为一个假定的预后指标[109].然而,在以色列进行的一项全国性队列研究发现,在严重急性呼吸综合征冠状病毒2 (SARS-CoV-2)检测呈阳性的哮喘患者人群中,使用生物疗法与中重度COVID-19风险增加、中重度COVID-19综合终点或全因90天死亡率无关(n=8242) [110].生物疗法和全身糖皮质激素均与SARS-CoV-2感染风险增加无关,但近期慢性全身糖皮质激素使用与中重度COVID-19风险增加以及中重度COVID-19复合终点或全因90天死亡风险增加相关。对英国24名确诊为SARS-CoV-2感染的成年人(占总研究人群的9%)进行的单中心分析显示,在使用benralizumab治疗严重嗜酸性粒细胞性哮喘时,没有出现严重的感染结局(即。重症监护病房入院、使用机械通气或死亡的要求)[111].总的来说,75%的患者有轻度感染,不需要住院;然而,这可能被低估了,因为研究人群中的其他患者可能经历了无症状感染,或者如果他们只经历了非常轻微的症状,就没有进行验证性PCR检测。
有趣的是,benralizumab成功用于治疗3例COVID-19患者的嗜酸性粒细胞增多和全身症状(DRESS)药物疹[112,113].所有3例患者对糖皮质激素治疗无反应,但在使用benralizumab后,嗜酸性粒细胞计数迅速下降,DRESS临床改善。这些发现引发了对贝那利珠单抗在COVID-19背景下治疗DRESS的探索[114].
嗜酸性粒细胞减少与恶性肿瘤的风险
小鼠模型研究
近年来,由于观察到在许多肿瘤类型中都检测到嗜酸性粒细胞浸润,嗜酸性粒细胞在肿瘤发展中的作用一直是广泛研究和兴趣的领域[57].一些吸引嗜酸性粒细胞到炎症或感染部位的相同因素可能会吸引嗜酸性粒细胞到肿瘤微环境(TME)。嗜酸性粒细胞在TME中的作用是有争议的,并可能因功能异质性而复杂化。对嗜酸性粒细胞分泌的细胞因子和其他因子的研究已明确表明它们具有抗肿瘤作用或刺激肿瘤进展[56,57].这些因素中是否有许多是由TME内的嗜酸性粒细胞产生的仍有待确定。
没有证据表明,嗜酸性粒细胞谱系切除小鼠自然增加了发生癌症或其他恶性肿瘤的风险,ΔdblGATA和PHIL转基因小鼠都不容易发生自发恶性肿瘤[80].然而,有证据表明,嗜酸性粒细胞在实验诱导肿瘤的小鼠中发挥肿瘤监测作用。年代imson等.[115]报道了具有BALB/c遗传背景的ΔdblGATA小鼠在接受甲基胆蒽治疗后,与同基因野生型对照相比,显示出纤维肉瘤发展增加。在IL-5和CCL11 (eotaxin-1)基因敲除导致嗜酸性粒细胞缺乏的小鼠中观察到类似的结果(il5 / ccl11- / -老鼠)。有趣的是,年代tathopoulos等.[116]发现将Lewis肺癌(LLC)或小鼠大肠腺癌(MC38)细胞胸腔内注射到il -5缺乏的(il5- / -C57BL/6小鼠减少恶性胸腔积液的发生与同基因野生型对照采用同样的处理。提供外源性IL-5到il5- / -引入LLC或MC38细胞后,小鼠恢复了MPE形成能力。随后的实验表明,在实验诱导的肿瘤模型中,小鼠IL-5的缺乏可以防止肺转移的发生[117].在这些模型中,IL-5敲除小鼠肺部嗜酸性粒细胞浸润大大减少与野生型控制。正如在其他实验系统中观察到的那样,嗜酸性粒细胞、嗜酸性粒细胞和嗜酸性粒细胞募集在癌症和TME中的作用是复杂的,值得进一步研究[56,57].
来自临床试验的数据
感染信号应该在3期临床试验的时间范围内变得明显,与此相反,恶性肿瘤发病率的增加可能只在较长时间内变得明显。迄今为止,抗il -5/ 5r疗法对恶性肿瘤风险没有影响[48,82,85,93- - - - - -99];在benralizumab、mepolizumab和reslizumab的长期研究中,新发恶性肿瘤的频率与基于研究人群特征的预期基本一致。另外,两项非常大规模(约3000例患者)的benralizumab安慰剂对照COPD试验提供了额外的保证,其中包括本质上癌症风险较高的人群(平均年龄65岁;三分之一的活跃吸烟者)[87].在这些研究中,使用benralizumab治疗的患者恶性肿瘤的发生率在数值上较低与安慰剂。总体而言,长期使用benralizumab治疗期间恶性肿瘤的发生率较低(<1%),在受影响的器官或组织类型方面没有表现出模式,与患者群体的预期一致(表3) [48,82,85,93,94].
虽然与监测、流行病学和最终结果(SEER;https://seer.cancer.gov)在一项针对HES患者的mepolizumab研究中观察到的数据,作者注意到HES患者的基线恶性肿瘤风险可能高于普通人群(从中提取SEER数据)[98].持续的抗il -5/ 5r和其他生物疗法的长期监测登记仍然很重要,特别是对于罕见和生长缓慢的恶性肿瘤。
嗜酸性粒细胞减少和妊娠
在怀孕期间接触减少嗜酸性粒细胞计数的生物疗法的经验是有限的。临床前研究表明,在非人类灵长类动物中,以超治疗剂量给药贝那利珠单抗在整个妊娠期不会导致不良后遗症或有致畸作用[108,118].在新生儿中观察到嗜酸性粒细胞抑制,除一名受试者外,所有受试者在产后6个月时逐渐恢复到正常计数。其他白细胞、血红蛋白浓度、血小板计数和抗体反应的测量结果与对照组相似。
最近的病例报告描述了一名患者在整个怀孕期间接受benralizumab治疗HES [118].病人的婴儿很健康,生长发育正常,没有特异反应性疾病的迹象。直到7个月大,孩子的外周嗜酸性粒细胞仍未检测到;在1年的评估中,嗜酸性粒细胞表型正常。在4、6、9和12个月接种肺炎球菌结合疫苗后,出现了适当的免疫应答。迄今为止的证据表明,治疗性嗜酸性粒细胞减少没有不良影响在子宫内;然而,目前可用的数据的局限性妨碍了一个真正的确定的影响,嗜酸性粒细胞耗竭在怀孕期间在这个时候。
结论
尽管传统观点认为嗜酸性粒细胞对蠕虫和可能的其他微生物病原体具有保护作用,但来自动物和人类的数据,无论是个体还是总体,都没有表明没有嗜酸性粒细胞的生活有有害影响。缺乏嗜酸性粒细胞的小鼠表现出活力和健康,没有感染或恶性肿瘤的倾向。虽然在某些嗜酸性粒细胞谱系切除小鼠实验感染模型中观察到蠕虫感染过程或免疫反应的一些改变,但对蠕虫感染的整体防御能力似乎是完整的。此外,新出现的数据表明,嗜酸性粒细胞可能是蠕虫感染的庇护所,而不是阻碍,这可能是生存进化压力的结果。在人类中,嗜酸性粒细胞的近乎完全耗尽并没有显示出对健康有不良影响,增加对寄生虫或机会性感染的易感性,或增加恶性肿瘤的风险。这些发现至少表明,在关键的宿主防御机制中似乎存在足够的固有冗余,以防止嗜酸性粒细胞的丢失破坏正常的免疫功能。因此,根据目前的数据,专门针对嗜酸性粒细胞的治疗策略的好处似乎不会被任何意外的免疫或关键稳态功能障碍所抵消,这些疗法应该仍然处于治疗嗜酸性粒细胞介导的疾病的前沿。
可共享的PDF
确认
写作和编辑支持,包括在作者的指导和指导下准备手稿草稿,并纳入作者反馈和手稿提交,由Crystal Murcia (CiTRUS Health Group, Philadelphia, PA, USA)提供,阿斯利康(AstraZeneca, Cambridge, UK)资助。
脚注
利益冲突:D.J. Jackson报告了阿斯利康、赛诺菲、诺华、基耶西制药、勃林格殷格翰和葛兰素史克的顾问委员会和演讲费。I.D. Pavord报告在阿斯利康、勃林格殷格翰、Aerocrine AB、Almirall、诺华、梯瓦、基耶西、赛诺菲/Regeneron、美纳里尼和GSK赞助的会议上发言的荣誉;阿斯利康(AstraZeneca)、GSK、赛诺菲(Sanofi/Regeneron)和梯瓦(Teva)为组织教育活动支付的费用;参加Genentech, Sanofi/Regeneron, AstraZeneca, Boehringer Ingelheim, GSK, Novartis, Teva, Merck, Circassia, Chiesi和Knopp顾问小组的酬金;GSK支付支持FDA批准会议的费用;赞助参加勃林格殷格翰、葛兰素史克、阿斯利康、梯瓦和基耶西的国际科学会议。
支持声明:本研究得到了阿斯利康的支持。本文的资助信息已存入交叉参考基金注册.
- 收到了2022年6月16日。
- 接受2022年7月25日。
- 版权所有©作者2023。
本版本根据知识共享署名非商业许可4.0的条款发布。为商业复制权利和权限联系权限在}{ersnet.org











![Development and characteristics of human and mouse eosinophils [1–3]. Several differences have been noted between mature eosinophils from humans versus mice. Human eosinophils are larger than mouse eosinophils, stain more intensely with eosin and have a lobed rather than a circular nucleus. Primary granules in human (but not mouse) eosinophils contain Charcot–Leyden crystal (CLC) protein/galectin-10. The dense core of secondary granules in both human and mouse eosinophils is predominantly composed of major basic protein-1 (MBP-1). Eosinophil-associated RNases in human secondary granules include eosinophil protein X (EPX), eosinophil cationic protein (ECP) and eosinophil-derived neurotoxin (EDN). Eosinophil peroxidase is stored in secondary granules in both human and mouse eosinophils. Human eosinophils respond to eotaxin-1 (CCL11), -2 (CCL24) and -3 (CCL26), whereas mouse eosinophils respond to eotaxin-1 and -2. Among cell surface molecule differences, human eosinophils express Siglec-8 and mouse eosinophils express Siglec-F. CCL: chemokine (CC motif) ligand; IL-5Rα: interleukin-5 receptor α; mEARS: mouse eosinophil associated RNases. Development schematic adapted from Lee et al. [1] with permssion.](http://www.qdcxjkg.com/content/erj/61/1/2201217/F1.large.jpg?width=800&height=600&carousel=1){kind=link}
{kind=link}
![Putative role for eosinophils in homeostasis and eosinophil-associated diseases. Homeostatic roles for eosinophils illustrated in this figure are supported by data from animal models and human in vitro studies. ABPA: allergic bronchopulmonary aspergillosis; CRSwNP: chronic rhinosinusitis with nasal polyps; EET: extracellular trap; EGPA: eosinophilic granulomatosis with polyangiitis; GI: gastrointestinal; HES: hypereosinophilic syndrome; IBD: inflammatory bowel disease; NCFB: non-cystic fibrosis bronchiectasis. Adapted from Jackson and Munitz [11].](http://www.qdcxjkg.com/content/erj/61/1/2201217/F2.large.jpg?width=800&height=600&carousel=1){kind=link}
{kind=link}