





摘要
近年来据报道肺炎患者肺炎脓肿(PPE)发病率的不明原因增加。本研究调查了从住院治疗严重肺炎的胸腔积液样本中检测到胸腔积液样本中的新分离株的遗传和生物学规范。
在未诊断的胸腔积液样本中应用了病原体发现方案,并导致了三种新的TTMV (TTMV- ly)分离株的鉴定。将分离的TTMV-LY基因组转染到A549和人胚肾293T细胞中,通过实时荧光定量PCR和全基因组扩增检测病毒复制情况。进一步用释放的TTMV-LY病毒粒子感染A549细胞,通过多重免疫检测诱导先天免疫应答。
三种TTMV-LY基因组的遗传分析显示了一种经典的基因组组织,但具有已知序列的弱异构(<64%)。我们展示了这一点体外肺泡上皮细胞TTMV-LY的复制与传染性病毒颗粒的有效释放。我们还表明,响应TTMV感染的炎症介质的选择性生产。
本研究报告了复制TTMV-LY的描述与患有PPE住院的儿童的肺炎术,表明病毒在肺炎发病机制中的潜在作用。
介绍
每年从肺炎死于肺炎的约180万儿童[1].广泛的病原体被认为是肺炎的病原体,但14-23%的病例在病原学上仍未得到诊断,尽管不断优化用于检测呼吸道病原体的诊断测试[2,3.].
已经开发了不同的方法,用于鉴定未经证实的,新的和发散病原体。近年来,这些技术导致了呼吸样本中几种病毒,包括人类metapneumovirus,Bocavirus,吴和Ki多瘤,以及扭矩对象病毒(TTV)[4].其中一些已被证实随后作为低呼吸道感染(LRTI)的致病药物,但大多数这些病毒的累及仍然是一个广泛的讨论问题。TTV属于最近创建的病毒族Anelloviridae,由单链圆形DNA病毒组成,例如TTV(Alphatorquevirus.)、转矩teno MIDI病毒(醋克雷维斯)和扭矩teno mini病毒(ttmv)(Betatorquevirus) [5].
在本研究中,我们从法国住院的肺炎性脓胸(PPE)儿童胸膜液中分离并鉴定了3个TTMV (TTMV- ly)新种。这种病理,随着发病率的增加,与高发病率相关,经常需要延长住院时间和侵入性操作[6].因为呼吸道被认为是TTV原发感染和持续复制的场所[7],我们进行了进一步的研究,以评估这些新病毒可能在多大程度上与病理有关。我们在这里报道,TTMV-LY分离在肺泡上皮细胞中积极复制,并能够调节其固有免疫反应。
实验步骤
患者和样品
一项前瞻性研究对2007年4月4日至2009年3月30日期间因个人防护装备住院的28名5个月至14岁儿童(12男16女)进行了研究,这些儿童在法国里昂临终关怀医院的儿科重症监护室接受治疗。本研究经所有参与机构评审委员会批准(CE 06-139),并获得所有家长的知情同意。在入院48小时内,使用一次性粘液提取器获得28例鼻咽吸出物。在置胸引流管时采集25例胸腔积液标本。样本保存在-70°C的冷冻环境中,待进一步研究。
呼吸道病原体核酸检测
使用FTD呼吸道病原体21+分析(Fast Track Diagnostics, Junglinster, Luxembourg)结合AgPath-ID一步RT-PCR试剂盒(Ambion, Austin, TX, amon),在28例鼻腔吸入和25例胸腔积液样本中检测了25种最常见的呼吸道病原体的存在。根据生产厂家推荐,采用NucliSENS®easyMAG平台(bioMérieux, Craponne,法国)(以下命名easyMAG)提取总核酸。
TTMV通过病原体发现和遗传表征分离
5例阴性胸腔积液标本采用a兰德等.[8]并基于随机RT-PCR产物的克隆和测序。“Homebrew”序列分析软件用于从数据库中存放的那些识别新的或发散序列。通过反向后的TTMV-LY-9F和TTMV-LY-8R引物进行反向PCR,对TTMV序列的片段进行TTMV全长基因组扩增的三种样品中的三种样品进行了TTMV全长基因组扩增(表格1).扩增子通过两条链被克隆和测序。使用MEGA5计划(http://megasoftware.net/mega.php),通过Clustalw生成多个对齐(www.clustal.org/omega.),并采用邻居连接法构建系统发育树。预测二级结构采用RNA structure 4.5软件(D.H. Mathews, M. Zuker和D.H. Turner)建模。有关程序的详情请参阅网上补充资料。本研究获得的3个全长基因组分别保存在GenBank数据库中,登录号如下:JX134044,JX134045和JX134046..
细胞培养及TTMV转染或感染
用两种细胞系:人胚胎肾(HEK) 293T细胞系和A549肺泡上皮细胞系(均来自CelluloNet, UMS3444/US8 Biosciences germany -Lyon Sud, Lyon, France)研究TTMV新分离株的复制。HEK293T细胞在DMEM、10%胎牛血清、2 mM谷氨酰胺和抗生素中维持,A549细胞在F12K、10%胎牛血清、2 mM谷氨酰胺和抗生素中维持。将克隆的TTMV DNA线性化生态rv和BAM嗨(罗氏诊断,梅伦,法国)。病毒和载体DNA被COTANSFECTE融入细胞(每10次1μg5使用制造商推荐的JetPEI试剂(polyplus -转染,Illkirch,法国)。对照组包括单独转染载体或单独转染JetPEI的细胞。转染的培养物在37°C和5%的二氧化碳中培养过夜。18 h后,用PBS冲洗3次,然后加入新鲜培养基。用报告质粒优化转染效率。在感染研究中,将健康细胞接种于24孔板,用转染后3天收获的TTMV dna转染细胞上清感染,在37°C和5%二氧化碳中孵育2小时,7 h和24 h。然后用PBS洗涤3次,用新鲜培养基孵育6 h。所有实验都至少进行2次,每次重复。
TTMV-LY复制
所有实验均在规定时间收集上清液,离心后立即用dnase在37℃处理2 h, -80℃冷冻。用easyMAG提取TTMV DNA, 50 μL洗脱。用2 mL NucliSENS裂解缓冲液(bioMérieux)收集细胞,用easyMAG提取核酸,用100 μL洗脱缓冲液洗脱。然后用酶切法消化细胞DNADPN.我或NDE.II,其分别切割输入TTMV的局部甲基化GATC位点和复制TTVMV的非甲基化的GATC-位点(表格1).为了完全消化DNA,降低qPCR中观察到的本底水平,外切酶III (ExoIII;Fermentas, Burlington, Canada),如其他地方所述[9].通过反相PCR检测TTMV的全长基因组。简而言之,反应混合物由5μl核酸组成,每个引物(TTMV-LY-11F和TTMV-LY-10R)(表格1),2mM的脱氧核苷酸三磷酸混合物,并扩大高保真酶混合物(Invitrogen,Darmstadt,德国)。通过Taqman实时PCR测定(Eurogentec,Seraing,Belgium)实现了复制TTMV的定量,所述引物特异性地靶向GATC位点。QPCR测定被优化,如前所序推荐[10].扩增步骤在25μL体积中实现,含有12.5μLIQSupermix(Bio-rad,Hercules,Ca,USA),每种引物和探针()0.3μm(表格1)和5 μL DNA。使用CFX96检测机(BioRad)进行扩增检测。每个反应中都包含未转染细胞的DNA。在转染研究中,复制率计算为转染后检测到的定量复制TTMV数量的比值DPN.i消化到TTMV的总量化量(复制和残留输入TTMV基因组)。
细胞因子/趋化因子释放的测量
从ttmv感染的A549细胞系培养中收集的无细胞上清同时定量人表皮生长因子、eotaxin、粒细胞集落刺激因子(G-CSF)、粒细胞巨噬细胞集落刺激因子(GM-CSF)、干扰素(IFN)-α2、IFN-γ、白细胞介素(IL)-10、IL-12(p40)、IL-12(p70)、IL-13, IL-15, IL-17, IL-1ra, IL-1α, IL-1β, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IP-10,单核细胞趋化蛋白(MCP)-1,巨噬细胞炎症蛋白(MIP)-1α, MIP-1β,肿瘤坏死因子(TNF)-α, TNF-β和血管内皮生长因子(VEGF),使用一个商业化的多重珠状免疫检测试剂盒(Milliplex;Millipore, Billerica, MA, USA)。
结果
鼻腔吸入和胸腔积液阳性样品中呼吸道病原体的患病率
采用灵敏的多重实时分子检测方法对因重症肺炎住院的儿童的鼻吸和胸腔积液样本中主要的呼吸制剂进行鉴定。在鼻吸样本中,发现鼻病毒、副流感病毒和人偏肺病毒是主要的呼吸道病原体(表2.).在80%的胸腔积液样品中发现细菌剂,而20%仍未诊断。
对5份阴性胸腔积液标本采用随机RT-PCR的病原体发现方案,其中3份获得了与TTMV相关的几个短序列。TTMV的特征需要对病毒全长基因组进行扩增和测序。Betatorquevirus-特异性反PCR扩增TTMV-LY基因组。TTMV-LY基因组由TTMV-LY1 2912 bp、-LY2 2979 bp和-LY3 2915 bp组成。采用PCR方法,在每个患者相应的鼻吸样中也检测到胸腔积液样本中发现的TTMV-LY基因组。3例胸腔积液标本仅为TTMV阳性的患者中,男性2例,女性1例。TTMV-LY1、-LY2、-LY3患者年龄分别为48个月、7个月、37个月,临床特征与队列中其他患者相同。按照国际病毒分类委员会的建议,对预测开放阅读框(ORF)1的核苷酸序列进行系统发育分析[5].基于系统发育树(图1),所有三种新的TTMV-LY分离株属于GenoGroup I,与先前识别的TGP96(64%的最大同一性)和PT-TTV8-II(44%的最大识别)分离物。TTMV-LY分离物具有至少三个ORF:分别为TTMV-LY1,-659氨基酸(AA)的一个主要ORF1,分别为TTMV-LY1,-659氨基酸和几种重叠ORF。TTMV-Ly1和-Ly3分别具有两个重叠ORF的91和101AA和97和129AA,而不是TTMV-LY2,其具有100和109AA的ORF,以及113AA的另外的ORF3。这些病毒的另一个特征是高度保守的5'-未结晶区域(UTR),如图所示图2..图2A显示TTMV-LY2在这个5 ' -UTR中有一个显著的插入,形成一个预测的RNA序列二级结构(无花果。2B.).该结构似乎能够形成稳定的环和二聚体结构。
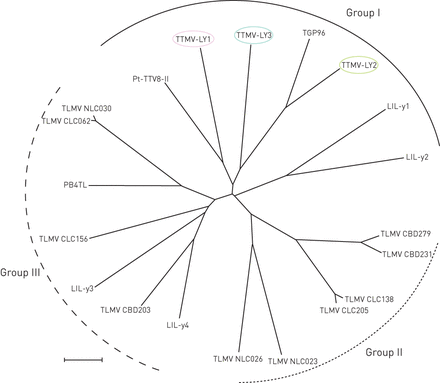
TTMV分离株TTMV- ly1、-LY2、-LY3和TTMV- ly1的开放阅读框-1核苷酸序列的系统发育树Betatorquevirus由邻接法构造。从胸腔积液中分离的TTMV-LY基因组被圈出。标尺= 0.1核苷酸替换每个位置。
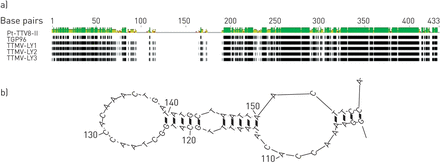
在新定义的系统发育I组核苷酸序列的5 '非翻译区域的多重比对显示,在扭矩teno迷你病毒(TTMV)分离物TTMV- ly2基因组中存在一个重要的插入(a),可以在预测二级结构中形成茎环模式(b)。序列中相同的残基给出黑色背景。残基之间的线表示序列之间的缺失。答:腺嘌呤;C:胞嘧啶;G:鸟嘌呤;T:胸腺嘧啶。
将TTMV-LY基因组转染到细胞中体外复制
在转染线性化病毒基因组后,我们监测:1)新的TTMV分离株在A549和HEK293T细胞中的复制;2)病毒颗粒在上清液中释放,用特定的标准化qPCR方法对新合成的基因组进行定量分析,持续1周。我们最初观察到图3一, TTMV-LY分离株均在两种细胞系中均有活性复制。3个菌株在A549细胞(25%)和HEK293T细胞(10%)中的复制平均率无显著差异。根据动力学实验(无花果。3B.),从第2天到第4天,TTMV-LY的平均复制率有规律地上升,在两种细胞系中均达到35%的复制形式。这种复制率在A549细胞中直到第7天保持相对稳定,但在HEK293T细胞中从第4天开始略有下降。TTMV-LY颗粒的释放与细胞内复制率相似,在转染后第4天达到峰值,A549增加8倍,HEK293T上清增加20倍。
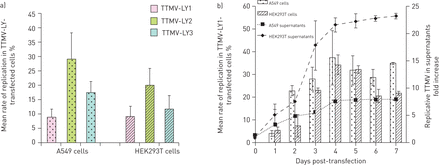
转染后TTMV- ly基因组在人胚肾(HEK) 293T细胞中复制的TTMV比在A549细胞中复制的多。直方图表示平均值±sd用线性化全长基因组转染细胞中TTMV-LY的复制速率。TTMV-LY的复制是A)在第3天后测量或B)在转染后一周内监测。B)中的曲线表明DNASEI(Thermo Sciencific,Illkirch,法国)处理和核酸提取后的培养培养基中TTMV-LY1在细胞的培养基中定量。结果表示为在第1-7天的定量TTMV-LY1与转染后的定量TTMV-LY1的比率0.数据仅针对TTMV-LY1表示,因为三种病毒中的复制曲线是相似的。
此外,我们确保了重复的TTMV-LY检测到图3.有效地与病毒颗粒相对应,而不仅仅是部分DNA片段。因此,TTMV-LY在A549细胞(无花果。4A)及上清液(无花果。4B.)测量。我们观察到三个TTMV- ly分离株存在一个2.9 kb的PCR片段,该片段代表了循环复制的TTMV,并通过测序分析进行了验证(数据未显示)。我们证实细胞中存在圆形全长基因组,并释放到A549细胞的上清中,表明存在病毒颗粒。
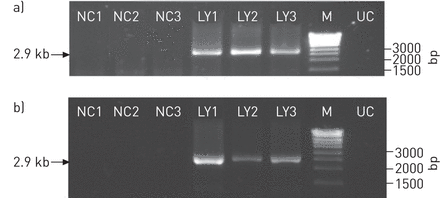
通过背对背引物的PCR检测,在a) A549细胞和b)转染后第3天的细胞上清液中分离的TTMV- ly1、-LY2和-LY3的复制循环基因组。NC1、NC2、NC3:各基因组阴性对照。LY1、LY2和LY3:分别转染TTMV-LY1、-LY2和-LY3基因组的细胞;M: DNA分子量标记;加州大学:未感染的细胞。
TTMV-LY感染颗粒在肺泡上皮细胞中复制
通过定量A549细胞中的TTMV-LY基因组来测量制备重复粒子的能力。在第3天在第3天后释放到转染的A549和HEK293T细胞的上清液中,我们暴露2小时,7小时和24小时的肺细胞。我们注意到,A549感染TTMV-LY的最佳条件与A549和HEK293T转染细胞的上清液分别孵育2小时和7小时后孵育(无花果。5Ab).我们还观察到TTMV A549进入细胞的效率在病毒载量最高时更显著(107个基因组当量)。此外,感染A549细胞上清液后,A549细胞内TTMV-LY的数量比HEK293T细胞内的数量(1.7×106)要多相对5.6×104,提示由肺细胞产生的病毒颗粒可能更具传染性。这一结果被两种细胞系感染后的复制TTMV-LY定量证实(无花果。5C).感染的A549细胞系统地显示出最高病毒滴度,分别具有48倍和167倍,分别增加了HEK293T和A549上清液的感染,相对HEK293T细胞在第3天的22-和44倍的增加。对于所有三种TTMV-LY基因组(数据未显示)制作该观察。
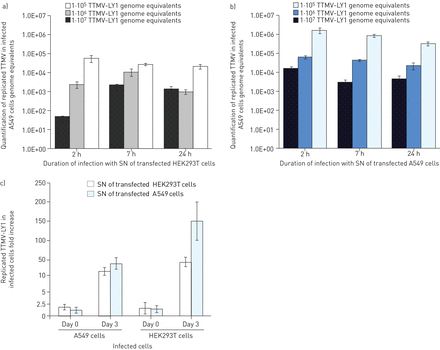
实时荧光定量PCR结果显示,Torque teno mini virus (TTMV)-LY分离物优先感染A549肺泡细胞系,而不是人胚胎肾(HEK)293T细胞系。a)和b)用TTMV-LY1孵育A549细胞2 h、7 h和24 h,三种不同的病毒载量为105, 106, 107在第3天转染后,从转染A549或HEK293T细胞的上清液(Sn)收获的基因组等同物。在培养基暴露细胞中测量的TTMV-LY的量为零。c)在2-H暴露的A549和HEK293T细胞中,在第0天和第3天测量复制的TTMV-LY的量,病毒载荷为1×10的病毒载量7基因组等同物。与暴露于TTMV-LY基因组的细胞上清液的细胞相比,结果表达为折叠增加,没有转染试剂。数据显示为平均值±SD。
每个单独的TTMV-LY以特定的方式调节肺上皮细胞的固有免疫反应
在这项研究中,我们使用了一种基于29-plex珠粒的免疫测定,测量用TTMV-Ly1,-Ly2或Ly3感染的肺细胞上清液中的各种炎症介质的水平。如图所示表3在感染后6小时,TTMV-LY诱导了几种细胞因子和趋化因子的产生,这些细胞因子和趋化因子与未感染细胞相比显著增强。我们观察到TTMV-LY1驱动大量促炎因子(IFN-γ、RANTES、IL-2和IL-12)、趋化因子(MIP-1b)和抗炎因子(IL-10和IL-13)以及生长因子(G-CSF、GM-CSF和IL-7)的产生。另外两个TTMV-LY基因组诱导的应答较弱,TTMV-LY2诱导RANTES、IL-12(p40)、MCP-1、VEGF和IL-13, TTMV-LY3诱导IL-2、IL-12(p40)和MCP-1。三个TTMV-LY分离株感染细胞后仅释放IL-12到上清液中。
讨论
本研究检测了在胸腔积液样本中鉴定的TTMV的新分离物的潜在累及,在儿童严重肺炎的发病机制中。主要研究结果是:1)TTMV-LY可以深入殖民肺部;2)发生有效复制的肺泡上皮细胞,允许TTMV-LY分离物;3)TTMV-LY感染通过诱导炎症介质的产生来调节肺细胞的先天免疫应答。
PPEs是由由细菌病原体感染引起的胸膜腔炎症引起的[11].细菌经常在胸腔液中描述,并且在该研究中也证实了胸腔积液样品的研究,但病毒来源的存在仍然有穷的文件[12,13].本研究是第一个揭示肺炎术中蒽虫病毒的存在。Anelloviruses的特征在于极端遗传多样性,在各种群体中具有高普遍性以及体液中的广泛分布。这种笨蛋,以及没有合适的体外文化系统,在该组病毒调查中受到了妨碍进展[14].
在以往的分子流行病学研究中,仍缺乏明确的证据表明anellovirus与疾病的发病机制和严重程度有关,或证据不足[15,16].在这里,我们发现在因重症肺炎住院的儿童的鼻吸液和胸腔积液中存在完整的TTMV基因组。在本队列中,与中国队列中216例LRTI患儿的鼻吸样本(数据未显示)一样,未观察到疾病严重程度与TTMV存在、患者年龄或性别之间的相关性。然而,与病毒复制相关的TTMV全基因组的存在可能表明其在肺炎发病机制中的作用。
本研究报告说,在严重肺炎的儿童中分离三种新的全长TTMV基因组(TTMV-LY)。作为第一步,研究了这些新分离物的TTMV-LY的生物学性质。我们首先转染三种TTMV-ly病毒的全长基因组,以确定病毒复制是否在HEK293T细胞中进行,如已经证明的[17,18],随后在肺泡上皮A549细胞中,作为II型肺细胞的模型[19].我们首次描述了TTMV,如TTV,在转染其基因组后能够在细胞系中复制,从而导致传染病的产生。我们观察到,肺泡细胞中的病毒感染比在肾细胞中更有效,表明肺细胞中TTMV-LY基因组的更好的热衷于肺细胞和在LRTI发病机制中的作用。我们假设如猪循环系统和乳头瘤病毒,TTMV的潜在致病性可能仅限于某些物种或一组物种[20.,21].实际上,在这项研究中,只有在不同患者中观察到高遗传的TTMV多样性,但在来自同一患者的不同样品中系统地发现了相同的全长基因组。此外,所有三种基因组属于相同的系统发育分支Betatorquevirus(组I)。我们研究了TTMV-LY三个基因组序列的保守的5'UTR基因组序列,以确定是否存在特定的复制模式。没有找到假设的复制起点(_AGT_TTACA),但在TTMV-LY2中检测到在保守的5 ' -UTR中插入200个碱基,形成茎环结构。如前所述,这个二级结构可能作为增强子和启动子元件参与病毒的致病性[22].尽管TTMV-LY2提出了这种二次结构,但与其他TTMV-Lys相比,没有观察到复制产率的增益。然而,这种遗传修饰的效果可以以另一种生物方式表达,响应于与宿主细胞的相互作用,如许多病毒所观察到。
第二步,我们通过测量参与炎症过程的可溶性介质的产生来研究宿主A549细胞对TTMV-LY感染的响应。肺泡II型上皮细胞通过形成肺泡屏障、产生表面活性剂和修复受损的I型上皮来维持肺泡的完整性[23].此外,它们是肺固有免疫的组成部分,通过细胞因子分泌来强化树突状细胞和肺泡巨噬细胞的功能[24,25].在A549细胞中未观察到细胞病变效应,但TTMV-LY感染诱导了复杂的固有免疫应答,由三种分离株不同的调节。因此,TTMV-LY感染不仅导致促炎细胞因子的产生,如前所述,包括IFN-γ [26]但还测量了显着水平的IL-10或IL-13抗炎细胞因子,证实TTMV-LY可以改变先天的免疫平衡,如TTV已经证明[27].不同分离物的TTMV的不同分离物可能因其他病毒感染观察到的方式而异的TTMV的产生不足令人惊讶[28,29,可能反映了它们被不同受体识别和/或通过不同分子途径相互作用。IL-12似乎在对TTMV-LY感染的反应中发挥了核心作用。这种细胞因子,增强活化t细胞和自然杀伤细胞的增殖和细胞毒性,被认为是调节宿主防御许多细胞内病原体的关键因素[30.].我们可能假设TTMV-LY介导的肺炎可能是由于IL-12的丰富分泌,尽管需要进一步的研究来证明这一点。
总之,这些结果表明TTMV-LY分离物可以侵入肺泡细胞,在那里它们在呼吸道中复制和诱导先天免疫不平衡。对于每种新发现的病原体的需要,需要进一步研究TTMV作为呼吸系统疾病中的呼吸系统疾病中的治疗剂的研究。这些调查允许我们了解在PPE的情况下,向我们了解扩大到炎症病毒的研究范围,以提高这种病理学的诊断效率。
致谢
作者感谢所有参与本研究的患者,以及中国的国际科技合作计划(2010年度福布33270),支持中国的一部分研究。我们特别感谢UMS3444 / US8 Biosciences Gerland-Lyon Sud(法国里昂)为其设施(流式细胞术和Cellulonet)以及其员工的可用性的贡献。
脚注
本文提供了补充材料www.www.qdcxjkg.com
支持声明:该研究得到了Mérieux(Lyon,法国)的支持。中国国际科技合作方案(2010年汇款33270)支持了中国研究的一部分。UMS3444 / US8 Biosciences Gerland-Lyon Sud(Lyon,法国)贡献设施(流量细胞计量和Cellulonet)和工作人员。
利益冲突:无声明。
本文于2016年4月修改,以纠正许可证信息中的错误。
- 已收到2012年7月12日。
- 接受2012年9月26日。
- ©2013年
ERJ开放文章是根据(188滚球软件创意共享归因非商业许可证3.0)











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}