





摘要
由吸气中阻塞引起的皮层电位由许多受体引起,其中许多受体可能在上呼吸道内。它们的确切性质尚不清楚。本研究的目的是通过研究局部上呼吸道麻醉对呼吸相关诱发电位的影响来提高这方面的知识。
呼吸相关诱发电位是通过在吸气中期闭塞后脑电图(EEG)周期的平均来描述的(C3.- cZ;C4- cZ)。共有21名健康志愿者(13名男性,22-52岁)在口腔呼吸期间、局部上呼吸道麻醉(利多卡因)前后进行了研究。此外,15名受试者在麻醉和不麻醉的情况下进行了鼻呼吸研究。六名受试者在吸入l -薄荷醇的同时进行了研究。
所有受试者均有典型电位,其组成部分具有正常的振幅和潜伏期。在口呼吸或鼻呼吸过程中,呼吸路径和上呼吸道麻醉对吸气阻塞的脑电图反应没有定性或定量的改变。l -薄荷醇没有效果。
对局部麻醉敏感的上呼吸道感受器不太可能对吸气中阻塞诱发电位有显著贡献。相反,更深层的受体,如关节和肌肉受体,可能对这些电位起主导作用。
各种呼吸刺激可以唤起人的皮层电位。这些方法包括直接刺激肋间肌1或者膈神经刺激2,3.。气道内突然的压力变化似乎特别容易引起皮质反应。这些可以诱导:1)在吸气开始时阻塞口腔气道4或者在此期间5;2)呼气时气道阻塞6;3)吸气阻力的突然施加7;4)吸气时对气道施加负压脉冲8或过期9。
对呼吸事件反应的皮层活动的研究是提高呼吸系统与大脑之间关系的生理学知识的有力手段。进一步来说,这种类型的研究可以通过建立相应的神经生理学底物来更好地理解某些呼吸感觉的机制。Knafelc的研究中给出了这种观点的例子et al。10将呼吸相关诱发电位的某些特征与机械负荷的量级估计联系起来,或在达文波特的研究中et al。11在哮喘儿童中,确定呼吸相关诱发电位早期成分的缺乏与威胁生命的哮喘病史之间的关联。在这个框架内,了解呼吸相关诱发电位的决定因素是一个重要的先决条件。
虽然皮质对施加在气道开口处的压力变化的反应可以反映来自呼吸系统各个组成部分的传入通路的激活,但有几行证据指向上呼吸道是其主要决定因素之一。事实上,Daubenspecket al。12他描述了通过喉罩(绕过大部分上呼吸道,但不包括喉传入)施加刺激时,早期和晚期皮层活动的主要变化。在患有吸气肌泵功能障碍的气管切开患者中,Donzel-Raynaudet al。13研究表明,在吸气过程中对气管进行封闭,抑制了在对口腔进行封闭时始终存在的反应。
上呼吸道含有几种类型的感受器,包括刺激性感受器,以及流量、压力和“驱动”感受器14,15,感知呼吸系统的主要状态的变化。这些受体的位置从粘膜或粘膜下(如。由流量变化激活的冷感受器,或喉刺激感受器)到肌内和关节(如。大量上呼吸道肌肉或关节感受器的纺锤和肌腱器官)。皮质对气道压力变化的反应是否涉及这些受体阵列的刺激或优先的一类尚不清楚。本研究旨在为这一问题提供一些答案。其目的是利用上呼吸道感受器对局部麻醉的不同敏感性14,15为了更好地理解它们各自对吸气中期闭塞后皮层反应的贡献。
材料与方法
研究对象及研究条件
健康志愿者21例(男13例,女8例,年龄22-52岁,体重指数19-30 kg·m−2)在完成法国人类生物医学研究法律程序后参加了这项研究。他们收到了关于这项研究的详细信息,并给出了书面同意。
研究对象被半仰卧在舒适的躺椅上,背部、颈部和头部都得到了舒适的支撑。他们被要求放松,但要睁着眼睛,以避免入睡或产生慢脑电波的风险。他们戴着耳塞和耳机,听自己选择的安静的音乐片段,以掩盖听觉线索。呼吸回路被一个金属框架牢牢地固定在他们的嘴巴上,以尽量减少由于面部肌肉活动造成的人工制品。
测量和程序
呼吸器
在涉及口呼吸的实验中(见后文),受试者戴着鼻夹,通过连接在加热气度表上的法兰口呼吸室内空气(Hans-Rudolf 3700;Hans-Rudolf, Kansas City, MO, USA)合并±2 cmH2O线性压差传感器(Validyne, Northridge, CA, USA)用于测量通气流量,并连接到非再呼吸阀(Hans-Rudolf 2600;(Hans-Rudolf 9340闭塞阀和9330压缩机;(完)。口压(P米)从吹口的侧端口测量(DP 15-32传感器,±50 cmH2O;Validyne, Northridge, CA, USA)。值得注意的是,电路的吸气部分还配备了一个小储液器,可以在其中沉积l -薄荷醇晶体(见下文)。在涉及鼻子呼吸的实验中(见下文),使用了相同的设备,但受试者通过密封的鼻罩呼吸(Comfort Classic;呼吸器,南特,法国),它与气压计相连,其中面罩压力(P面具)是可以衡量的。受试者被要求紧闭嘴巴。
呼吸相关诱发电位
表面电极放置在大脑皮层体感区CZC3.C4以国际10-20制为基础。C3.和C4参考答案CZ分别记录左、右活动。电极阻抗维持在5 kΩ以下。呼吸相关电位由吸入中封闭400-500 ms诱发5,每三到六次呼吸随机进行一次(与本作者先前的研究一样,选择吸气中闭塞13,16,因为它们所产生的电位比起搏-吸气闭塞所引起的电位“更尖锐”5)。信号在0.5 Hz - 500 Hz带宽上以1 kHz采样(Neuropack Sigma®;Nihon Kohden, Tokyo, Japan),根据气流轨迹确定吸气开始前100 ms和吸气开始后2 s进行数字化,并存储在Apple Macintosh计算机上用于后续分析(PowerLab®;AD仪器,Hastings,英国)。数据分析离线进行。对某一特定审判的个人陈述从计算机内存中被召回并显示在屏幕上。遮挡灵感然后选择使用P米信号。仅在存在稳定的脑电图(EEG)信号基线和没有明显异常事故的情况下,才保留给定的阻塞呼吸进行平均。此外,“对照”试验是通过平均相同数量的未封闭呼吸获得的。为了确保信号的即时重现性,在每种研究条件下系统地对80次阻塞呼吸进行两次单独的平均,并将所得到的痕迹叠加。然后对所有轨迹进行整体平均。
根据Davenport法测定呼吸相关诱发电位第一阳性(P1)、第一阴性(N1)、第二阳性(P2)和第二阴性(N2)分量的峰值潜伏期et al。17,从一个“零”点确定的平均值P米线的交点P米基线与第二线通过最陡峭的部分P米摇摆(MatLab5.3®;数学工作,纳蒂克,马萨诸塞州,美国)。从基线到峰值测量所有分量的振幅。
上呼吸道麻醉
5%的盐酸利多卡因溶液被喷在鼻腔和口腔,直到他们被麻醉。受试者被要求吞下麻醉剂以达到后咽部的麻醉。如有必要,给予额外的多卡因直至抑制呕吐反射,然后进一步从口咽后部尽可能远地喷射。达到这一结果所需的利多卡因总剂量从未超过400毫克。所有受试者在手术结束时均报告出现吞咽困难。局部敏感性在局部麻醉下进行的两次试验之间进行测试(见后文)。为了做到这一点,受试者被问及他们的口咽部感觉是否发生了变化,以及最初报告的吞咽困难是否仍然存在。此外,他们还用抹刀探查咽后部,并询问受试者是否有触感或恶心感。如果这些问题的答案都是“是”,就会给予额外的利多卡因。
实验设计
每个实验阶段包括由两个独立的吸气闭塞系列组成的基线评估,然后是由两个独立的吸气闭塞系列组成的“测试”评估。共进行了三组实验,具体如下。1)上呼吸道麻醉对口呼吸时吸气阻塞诱发皮层电位的影响。所有21名受试者都参与了本次研究。在6例病例中,在麻醉后90分钟洗脱后再次记录。2)上呼吸道麻醉对鼻呼吸时吸气阻塞诱发皮质电位的影响。15名受试者参加了本研究,该研究在单独的一天进行。3)冷感受器刺激对口呼吸时吸气闭锁诱发皮层电位的影响。在六名受试者中,吸入l -薄荷醇(300毫克的l -薄荷醇晶体沉积在连接呼吸回路吸气肢体的储层中)来刺激上呼吸道流量感受器。根据相同的设计和程序,在另一天进行了特定的实验。 The effect of L-menthol was considered complete 5 min after the beginning of inhalation18。
此外,在18名受试者的口呼吸和15名受试者的鼻呼吸期间,评估了上呼吸道麻醉对吸入l -薄荷醇相关感觉的影响。
统计分析
数据以均数±表示sd。在检查数据集的正态性和同方差后,使用配对t检验对电位的潜伏期和振幅进行从右到左的比较,以及与口腔和鼻腔呼吸路径获得的结果之间的比较,以及基线和测试条件之间的比较。在进行冲蚀测量的六名受试者中,三种条件用Tukey的线性方差分析进行比较事后测试。当I型错误的概率<5%时,差异被认为是显著的。
结果
典型的呼吸相关诱发电位(图1⇓–⇓3.⇓)在基线时所有受试者双侧均存在。
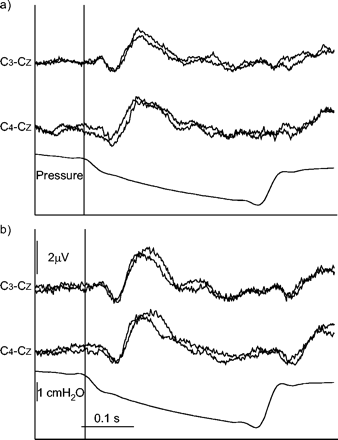
对15名受试者在a)口呼吸和b)鼻呼吸过程中进行吸气中封闭所引起的呼吸相关诱发电位进行比较。C3.- cZ和C4- cZ对应于在所有受试者中获得的吸气闭塞相关脑电图周期的整体平均。每一个C3.- cZ和C4- cZ在a)和b)中推导,两条迹线中的一条对应于基线时记录的电位,另一条对应于上气道麻醉后记录的电位(无显著差异)。鼻呼吸时记录的电位略高于口呼吸时记录的电位,但潜伏期和振幅无显著差异。垂直线表示零点(见材料和方法)。
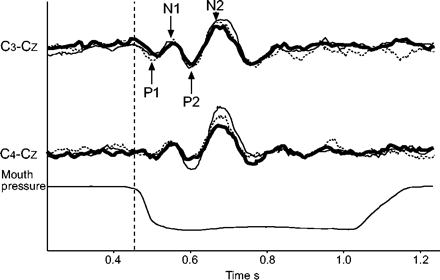
在一个受试者(——)前、(▒)中和(····)后观察到的呼吸相关诱发电位的例子。C中的每条轨迹3.- cZ和C4- cZ对应于从80次吸气中闭塞获得的两个独立的重叠痕迹的整体平均。垂直线表示零点(见材料和方法)。通常,电位开始于第一个积极成分(P1;40-60毫秒后开始与负荷相关的口压变化),被认为反映传入信息的皮层到达。之后的分量,典型的是负的、正的和再次负的(分别是N1、P2和N2)可能在不同程度上反映了感觉信息的认知加工,就像刺激后约300 ms出现的进一步分量一样。
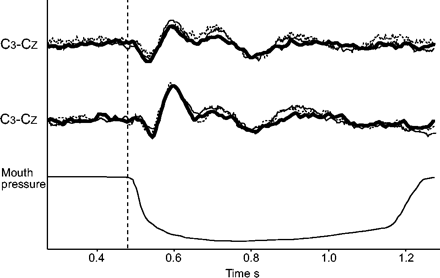
一名受试者吸入l -薄荷醇前(——)、期间(▒)和之后(····)呼吸相关诱发电位的例子。C中的每条轨迹3.- cZ和C4- cZ对应于从80次吸气中闭塞获得的两个独立的重叠痕迹的整体平均。垂直线表示零点(见材料和方法)。
在基线条件下,各组分的平均潜伏期和振幅均在正常范围内(表1列前)⇓和2⇓)。从右到左没有显著差异。在口鼻呼吸期间测量的潜伏期和振幅彼此之间也没有显著差异(图1)⇑)。
上气道麻醉(21名受试者)并没有减少呼吸相关诱发电位及其连续成分的出现次数。它不影响成分的延迟,也不影响它们的振幅(图1)⇑和2⇑;表1⇑和2⇑)。这是真实的任何一方,在嘴和鼻子呼吸。在这样测试的六名受试者中,洗脱数据与基线和麻醉数据没有差异(图2)⇑)。
同样,吸入l -薄荷醇(6名受试者)并没有减少呼吸相关诱发电位及其连续成分的出现次数。它不影响组件的延迟,也不影响它们的振幅(图3)⇑;表3⇓和图4⇓)。在接受测试的6名受试者中,洗脱数据与基线和麻醉数据没有差异(图3)⇑)。
在鼻呼吸和口呼吸过程中,上呼吸道麻醉一致地消除了与吸入l -薄荷醇有关的冷样感觉。
讨论
目前的研究表明,上呼吸道麻醉不影响正常人体中吸气阻塞引起的皮质电位。这一观察结果可用于推断与呼吸相关的诱发电位的来源,但要记住,相应的假设可能不适用于其他类别的个体。
初步考虑
首先,必须承认,上呼吸道麻醉对呼吸相关诱发电位的影响的缺失可能意味着上呼吸道对呼吸相关诱发电位的贡献是不重要的,或者是完全多余的。在各种类型的吸气肌泵功能障碍患者中,唐泽-雷诺et al。13表明上呼吸道对呼吸相关诱发电位的早期成分的发生至关重要。然而,目前缺乏健康受试者的可比信息,因此不能完全排除donzel -雷诺的观察结果et al。13在研究的患者中,这是由于胸腔和通气肌的传入血流缺乏或异常。在健康人或其他疾病中,调节机制可能不同。然而,值得注意的是Daubenspecket al。12当使用喉罩绕过大部分上气道(喉部下部除外)时,观察到由上气道压力变化引起的全球场功率的重要下降。总而言之,本文作者认为将以下讨论建立在上呼吸道对呼吸相关诱发电位的重要贡献这一事实上是合理的,但也意识到可能涉及其他机制。
鼻呼吸和l -薄荷醇的作用
毫无价值的是,本文作者没有观察到在基线条件和麻醉条件下,口呼吸和鼻呼吸引起的吸气中阻塞电位之间存在显著差异。这表明这种类型的电位可以在一种或另一种呼吸路线中交替研究。这一新信息对未来的研究可能很重要,因为鼻子呼吸是人类休息时的正常呼吸方式。
吸入l -薄荷醇并不影响六名研究对象的呼吸相关诱发电位(表3)⇑和图4⇑)。由于“冷感受器”实际上是流感受器,这在吸气闭塞范式中并不意外。l -薄荷醇对受体的强烈激活也可能导致它们的感觉通路饱和。最后,这个子组的规模可能不足以检测出差异。
上呼吸道感受器和局部麻醉
上呼吸道的呼吸感觉神经支配最为复杂14,15,19,20.有很大的种间差异。鼻传入终末器官由许多游离神经末梢组成,分布于鼻上皮细胞内及其下方15。局部麻醉显著影响它们的活动21。鼻压敏感受器存在于几种物种中。局部麻醉也容易减弱它们的活性19。咽部的呼吸感受器很少,而且咽部不被认为是维持呼吸道通畅的重要反射部位22。然而,咽壁的肌肉含有骨骼肌特有的感受器,就像腭和舌的肌肉一样15。喉区神经分布密集。它具有流动感受器、大量的透壁压力感受器和“驱动”感受器,这些感受器由喉内肌的收缩或通过气管或两者传递的喉头被动运动所刺激15,23。前两类患者对局部麻醉敏感15,24,而这只是“驱动受体”的部分情况。23,25需要肌肉麻痹和气管固定才能消音。这些受体可能包括韧带受体、关节受体和肌肉受体,在咽部和喉部肌肉中非常丰富。已知局部麻醉后喉肌感受器伸展反应的传入活动持续存在26。
结果解释
除了前面“初步考虑”一节中提出的问题外,麻醉剂未能到达所有相关受体也可以用来解释所呈现的结果。事实上,尽管鼻腔和咽部受体很可能被正确麻醉,但目前的作者并没有在喉下部进行视觉控制的利多卡因应用。然而,部分喉麻醉可能是由于所有受试者都注意到吞咽困难,以及麻醉后口腔呼吸时持续注意到的l -薄荷醇诱导的冷感消失。目前为止,阻止部分气道受体暴露于给定的刺激确实显著地改变了相应的电位12即使不完全麻醉也会对呼吸相关诱发电位产生一定影响。由此,本文作者认为,局部利多卡因对受试者呼吸相关诱发电位完全没有影响,这表明由吸气阻塞激活的受体对局部麻醉不敏感(值得注意的是,已有研究表明,对局部麻醉敏感的上气道受体对与体积变化相关的呼吸感觉不是必需的27)。推测,这表明与呼吸相关诱发电位相关的上气道受体可能属于前面描述的“驱动受体”(关节和肌肉上气道受体)。尽管在人类身上进行实验可能非常困难,但专门设计来测试这一假设的研究是有必要的。
根据之前的讨论,这使得冷感受器和压力感受器的贡献不太可能(这对于冷感受器来说并不奇怪,因为吸气中期阻塞产生的刺激发生在没有流动的情况下;在这个框架中,通过吸入l -薄荷脑来改变喉流受体的基线活动,并没有引起呼吸相关诱发电位特征的明显变化)。综上所述,作者认为在吸气中期闭锁后参与呼吸相关诱发电位的上呼吸道受体可能主要属于“驱动受体”的范畴。如果这是正确的,关节和肌肉气道受体似乎是检测和处理由大脑皮层刺激引起的机械变化的重要决定因素,这种现象对控制上呼吸道通畅至关重要。
- 收到了二零零四年十二月八日。
- 接受2005年8月8日。
- ©ERS期刊有限公司










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}