





文摘
肺结核是增加。目前的治疗方案要求至少6个月,因为潜在的或固定相生物很难杀死。这样的方案不能实现完全的遵从性,“直接观察治疗短期课程”(点)是比预期减少的影响。这个令人担忧的情况加重合并感染人类免疫缺陷病毒(HIV),和耐药菌株的增加。
我们需要新的见解,导致更多的快速治疗和免疫疗法,和更可靠的疫苗。
最近的见解来自:理解之间的关系结核分枝杆菌和巨噬细胞;承认分枝杆菌肽的多个T细胞类型,脂质和糖脂;干扰素γ应承担的重要作用(IFNγ)和白介素12 (IL 12)应承担的人类分枝杆菌感染了基因有缺陷的孩子;定量的存在和重要性Th2淋巴细胞激活人类结核病;当地的角色转换的不活跃的可的松活跃的皮质醇病变;也认识到一些有效的预防性疫苗immumotherapeutics而另一些没有。从长远来看最近的测序结核分枝杆菌基因组将导致进一步发展。
在短期内,有效的免疫疗法仍是最容易突破的管理肺结核。实际推进的类型,将从基因组测序结果讨论了大胆的,但尚无法预测。
结核病是一个全球性的紧急情况。世界上三分之一的人口感染,尽管只有5 - 10%发展活动性疾病第一次接触后几年1,这还导致了一个巨大的负荷,每年有八百万新发病例,三百万人死亡。此外,发展到疾病的比例正在增加。肺结核是第一个激活的继发感染人类免疫缺陷病毒(HIV)阳性的个体2。此外贫困的压力、营养不良和战争,增加的速度复活原因稍后讨论。即使在发达国家,如英国,疾病分布在大城市比较贫困的分布3。与此同时医疗保健系统的故障导致不完整的情况和接触者追踪,不完整的治疗和增加耐药性。在世界的某些地方,许多可用的药物被伪造或过时了4。在许多地区,现有的治疗可能是弊大于利,作为耐药不完整的治疗方案选择。耐多药结核病正在以惊人的速度蔓延,入侵西欧从东部块爱沙尼亚等国。有更多的肺结核病例在1999年人类历史上前所未有的。
六个月的治疗方案的问题
目前未能控制结核病的一个重要原因是,即使是最好的可用的治疗必须持续至少6个月。这个治疗方案是不现实的命题在大多数发展中国家,甚至在富裕国家的内陆城市,因为病人觉得几周后,停止服用这些药物。世界卫生组织(世卫组织)现在承认,短期直接观察治疗(点),病人的监督在采取每一剂量的治疗,帮助但不解决问题5。
持续的杆菌和潜伏性感染
有两个相关的原因要求6月方案。第一个是显而易见的,经常讨论。化疗杀死绝大多数细菌在几天内,但也有亚种群的“持续程序”6。目前尚不清楚这些生物是否在真正的固定相7或者仅仅是复制非常缓慢。也不清楚他们所在的地方。大多数作者认为他们在旧病变或网站纤维化或钙化,氧气可用性可能很低。然而,在一个被遗忘的论文发表于1927年,欧派和阿伦森8报告说,80%的结核性损伤已经无菌原发感染后5年,而活细菌可以在宏观上发现正常的肺组织。灭滴灵,一种药物,应该积极在厌氧条件下,不活跃在潜伏结核感染小鼠模型,意味着活的生物体也坚持氧化网站在这个物种9。
不仅持续程序生物治疗造成问题,但他们也构成一个重要的感染源。他们可以持续个人的生活的其余部分10,至少在较低或中度肺结核风土性的国家,很多情况下肺结核由于潜伏期感染11- - - - - -13。
保护与免疫病理
需要长期治疗的其他原因通常是被忽视的。大多数结核病患者有坏死的应对模式结核分枝杆菌科赫率先指出,类似的现象14因为豚鼠。有压倒性的证据表明科赫现象并不是一个最佳的保护性免疫的相关结核病。事实上preimmunisation动物所以他们以前科赫现象与致命的挑战结核分枝杆菌结果在一个清晰的和可再生的增加对疾病的敏感性,而nonimmunised控制15。这和其他方面的科赫现象在后面详细讨论。其相关性在这一点上,这种不恰当的模式迅速反应可能不正确的治疗。因此如果停在3个月的化疗,复发率很高16即使化疗是一个最佳rifampicin-containing实现痰消极之前3个月,尽管事实上,很少有生物生活在病人的组织。
因此,我们需要了解保护性免疫和科赫现象之间的差异,并确定哪些响应模式存在的因素。学习的最终目标是取代的病理反应中保护一个非常早期的治疗。
对结核病的免疫力
免疫小鼠肺结核
抗体
普遍认为,结核病抗体areirrelevant免疫力。这种假设可能为时过早。老鼠缺乏B细胞似乎反应通常感染17。然而,在另一个小鼠模型中至少有一个单克隆抗体被发现显著的保护18。这件事已经详尽的回顾和显然需要重新调查19。而很明显,大部分的抗体形成是无关紧要的,它仍然是可能的,中和抗体特定药物活性成分的生物将发挥重要作用。不幸的是这种活性成分的识别的生物仍处于起步阶段。
1型反应的至关重要的作用
能力tomanipulate与中和抗体或基因敲除小鼠的免疫系统提供了强有力的证据表明,在这个物种,免疫肺结核与1型反应。比起现场T辅助(Th)应承担的1或Th2细胞CD8的一致行动+细胞,和许多其他类型的细胞包括巨噬细胞、B细胞和基质细胞。集体产生两个细胞因子释放的模式被称为1型(由白介素2 (IL 2)应承担的,应承担白介素12 (IL 12)应承担的,应承担和干扰素γ(IFNγ))和2型(由白介素(IL)应承担的高4在5日和13)20.,21。“1型”一词用于优先Th1当它目的是指整体模式中所有细胞类型的细胞因子释放受感染的网站,而不是仅仅由CD4+辅助T cellsthat Mosmann包含在最初的计划22。
破坏的主要组织相容性复合体(MHC)二类基因或基因的α/ββ链的T细胞受体23导致CD4的不足+α/βT细胞,使老鼠敏感甚至无毒杆菌Calmette Guerin (BCG)。基因的中断IFNγ让老鼠非常容易结核分枝杆菌(在3周内死亡),这样老鼠甚至许多周后死于与波士顿咨询公司的挑战24- - - - - -26。IL 18基因敲除(KO)应承担的老鼠也更敏感,也许是因为IL 18有助于IFNγ表达式的感应27。1型通路的主要诱导物是IL 12。细胞因子的确切作用取决于鼠标应变28,但IL 12 KO小鼠更容易肺结核29日。
2型反应的有害作用
这些数据强调1型反应的至关重要的作用。同意这一点,其他数据表明2型反应不仅无法保护小鼠,但会严重破坏1型反应的疗效。如果2型应对疲弱共享分枝杆菌抗原是故意诱导挑战之前,老鼠发现明显更容易比nonimmunised结核病控制的动物30.。同样,在肺的Balb / c小鼠模型(肺结核)结核感染,IL 4应承担的出现在肺部病变(所看到的免疫组织化学和反向transcriptase-polymerase连锁反应(rt - pcr)同时暂时和空间领域的出现肺炎和坏死,导致快速临床恶化和死亡31日。声称这些观察并不反驳,进入IL 4基因的老鼠,目前没有证据表明增加抵抗感染32。首先,这种老鼠并不缺乏2型细胞因子活动因为地理IL 13可以代替许多破坏了基因的功能。其次,2型反应的有害作用最明显的进步阶段的疾病,尤其是在60天之后30.,31日。2型的角色在免疫病理反应是稍后讨论。
T细胞介导免疫结核病人
分枝杆菌抗原的反应模式
一些确认的必要性1型反应的人,和老鼠一样,出现在比较患者和健康的联系。例如,病人产生相对更多的抗体,而正常的接触产生相对更强的30 kda抗原的T细胞反应结核分枝杆菌。此外,患者的细胞释放更少的IFNγ和更多的IL 10的抗原35。同样,从BCG-immunised对象T细胞反应moreto 16 kdaα的晶状体蛋白的蛋白质结核分枝杆菌高于肺结核患者的T细胞,相比之下,有高水平的抗体36。这种类型的研究结果表明,1型反应是保护,如老鼠。研究血清细胞因子的浓度是不提供信息的肺结核37。
遗传易感性肺结核;自然基因淘汰赛
常规肺结核患者的遗传研究表明,多态性基因编码natural-resistance-associated巨噬细胞蛋白(NRAMP1), IL 1基因簇,应承担的维生素D受体和mannose-binding凝集素与易感性有关38。NRAMP1仍不确定的功能和审查39。维生素D受体多态性的相关性增加时也缺乏维生素D的古吉拉特语人口在伦敦40。然而对疾病易感性的影响很小,到目前为止,这种类型的研究将小的免疫机制。
另一方面明确的证据表明,1型响应是至关重要的免疫结核病研究的另一位来自儿童1型细胞因子系统的遗传缺陷。接种卡介苗,无毒的导数有机体负责forbovine肺结核,偶尔引起播散性感染。IFNγR1基因的基因在这样一个孩子,有一个单核苷酸缺失导致N附近建立一个过早终止密码子的终点站41。另一项研究42涉及四个孩子来自同一个小镇在马耳他,谁提出与分枝杆菌感染传播。分枝杆菌物种隔离分枝杆菌fortuitum,鸟型分支杆菌(2株)和分枝杆菌chelonei。一个孩子也有长期的沙门氏菌病。这些孩子有一个单核苷酸替换(C)而不是删除42。它允许正常水平的表达的信使核糖核酸(mRNA),但同样引入了过早的终止密码子。
IL 12应承担的受体缺陷也被发现在健康人分枝杆菌感染。与儿童IFNγR不足,这些患者能够成熟granulomata形式,但他们的自然杀伤(NK)细胞和T细胞分泌小IFNγ。因此,IL 12应承担的高依赖IFNγ分泌人类似乎至关重要的分枝杆菌感染的控制,尽管成熟肉芽肿的形成43,44。
参与免疫T细胞类型
除了传统的CD4细胞+α/β二类MHC-restricted T细胞,其他几个T细胞也参与反应的分枝杆菌。
CD8+T淋巴细胞
实验,涉及过继转移,在体外细胞耗竭,基因淘汰赛(如。β2量微球蛋白缺陷动物),说明CD8的重要性+细胞在小鼠肺结核的控制23,45,46。保护小鼠接种分枝杆菌热休克蛋白65脱氧核糖核酸(DNA)似乎主要是由CD8介导的+细胞47。在一个在体外系统,这种激活CD8的能力+细胞似乎涉及造成泄漏的吞噬体抗原进入细胞质,因此加入介绍MHC类的传统途径148,49,但另一个新颖的途径也可能参与进来50。haemolysin-like分子实际上是表达的结核分枝杆菌和波士顿咨询集团51和BCG菌株溶血素的表达单核细胞增多性李斯特氏菌了相信这将增加CD8吗+响应52。
这些CD8+细胞是细胞毒性,尽管这种细胞死亡的机制一直存在争议。它被认为最有cytoxic T淋巴细胞(ctl)的行为应承担的溶解感染细胞并允许发布的分枝杆菌被激活,可能杀死他们的未感染的巨噬细胞。然而,现在看来,一些ctl直接杀死结核分枝杆菌通过granulysin granule-associated蛋白质,与穿孔素作用53。另一方面,由CD4细胞溶解+细胞毒性T细胞不减少含有细菌的生存能力54。进展的结核病小鼠缺乏穿孔素并不不同于野生型发展55,56。小鼠CD8的重要作用+细胞在这个阶段可能IFNγ的分泌57,58。最近,tuberculosis-specific CD8+在人类细胞也被确认59,60在这个物种,但是他们的作用同样是不确定的。有CD8+细胞将识别结核病感染细胞和血液中分泌IFNγ患有这种疾病61年,但这似乎并没有导致ofintracellular扩散控制结核分枝杆菌在一个离体的系统用人类细胞62年。
大多数tuberculosis-specific CD8+细胞识别抗原与MHC类我,然而一些目前已知限制其他分子,如CD1(见下文)63年,64年。
CD1限制淋巴细胞
相对nonpolymorphic CD1分子MHC类的家庭我的类,并拥有疏水裂,脂质和糖脂分子结合,允许他们的演示各种CD1-restricted细胞,包括αβ对CD4和CD8 T淋巴细胞-分子(所谓的双重否定T细胞),γδT细胞和某些CD4细胞+,CD8+CD8α/α+和NK淋巴细胞65年,66年。
的确切角色CD1、和CD1-restricted细胞,保护或病理的肺结核,已被证明难以评估,因为老鼠拥有同系物CD1d但没有人类CD1a同系物,b或c。事实上,老鼠缺乏CD1d尚未发现不同于控制结核病的易感性67年,尽管有一个声称中和CD1导致恶化感染的老鼠在非常早的时间点68年。这些发现对人类疾病的相关性是值得怀疑的。人类CD1d尚未出现分枝杆菌抗原,与CD1a不同,b和c可能出现霉菌酸,lipoarabinomannan和其他分枝杆菌细胞壁组件69年- - - - - -71年。
双重否定(CD4 - CD8 -αβT细胞受体(TCR))淋巴细胞能够识别上下文中的分枝杆菌脂质CD1。他们的主要效应机制似乎IFNγ的分泌和CD95 / CD95L交互,并且很少他们重要的分枝杆菌死亡原因64年。这促使建议,他们的作用是消除局部炎症反应的下调的antigen-loaded抗原呈递细胞。许多其他类型的人类CD1-restricted T细胞也产生大量的IFNγ,但似乎能够溶解感染细胞,直接杀死胞内分枝杆菌53,64年,71年。
看来,结核分枝杆菌可以抑制CD1表达人类抗原呈递细胞,免疫反应从而可能逃避这个组件72年。
γδT淋巴细胞
如前面所讨论的一些γδT淋巴细胞识别的脂质和醣脂类上下文中的分枝杆菌产品CD1分子。然而,主要的人类外周血亚型,Vγ9Vδ2,也遍布和分泌细胞因子暴露在蛋白73年- - - - - -75年和其他非蛋白76年,77年抗原来源于结核分枝杆菌。γδ细胞积累在实验性早期病变78年和在体外研究表明对被感染的巨噬细胞细胞毒性79年。小鼠模型表明γδ从高剂量细胞在保护中发挥作用,系统性结核分枝杆菌innocula,但对于保护小不太重要,气溶胶的挑战80年,81年。在后一种情况下,监管角色建议因为老鼠缺乏γδ细胞有更高的初始,然后开发一个更化脓性细菌负担和破坏性的反应,可能与夸张的病理学相关结核病患者低水平的结核分枝杆菌活性γδ细胞82年。
结核患者的外周血γδ细胞的表型出现激活(老年病ICAM 1和MHC II级)83年,但是直到最近一直有争议,他们的总数在病人的血液和支气管肺泡灌洗(BAL)。这在很大程度上是通过快速老年病的示范来解决表面CD95 Lγδ细胞和提示activation-induced细胞死亡,使分析的时机至关重要84年,85年。
T细胞凋亡
在短期内文化、刺激结核分枝杆菌抗原诱发重大γδ细胞凋亡在病人和正常人,通过一种机制涉及Fas (CD95)84年。这可能与诱导Fas配体表达的订婚γδ分枝杆菌抗原的T细胞受体85年。对CD4的影响+T细胞凋亡可能取决于文化的分枝杆菌制备和持续时间。一组,利用肺结核患者的细胞,已经注意到2倍增加CD4 T细胞凋亡诱导活H37Ra 96 h。效果不再出现在相同的帖子还是治疗的患者86年。也有报告说,结核病感染导致增加Fas表达和减少bcl 2应承担的CD4的表达+T细胞。当这样的T细胞受到刺激在体外,他们显示细胞凋亡增加,减少生产IL 2和IL IFNγ但不是4。这表明选择性Th1类细胞凋亡,这可能是一个因素在向Th2开关87年,88年。
2型反应在人类肺结核
这些观察结果,最重要的是,儿童的易感性与受体缺陷IFNγ或者IL 12日提供明确的证据1型细胞因子的重要性,并建议密切与小鼠模型。最近,2型细胞因子的负作用在人类肺结核(TB),再并联小鼠模型,建立了89年,90年。地理表达IL 4,是否通过流式细胞术,或敏感定量rt - pcr如果外周血T细胞91年,增加(图1所示⇓)和与疾病严重程度和空化89年,90年。地理IL 4 mRNA拷贝数也与总免疫球蛋白E (IgE)(图。1⇓)89年和可溶性CD30水平(未公开的数据)。因此尽管实际细胞因子水平确实对Th1和信使rna复制数字高于肺结核Th2细胞因子,主要变化在细胞因子的表达而健康的捐赠者并不如前所述,小Th1细胞因子的表达减少,而是大规模高褶皱区域(80 - 100)增加Th2细胞因子的表达89年。这解决了一个长期争议,值得解释。有一个Th2组件在人类肺结核病人的反应结核分枝杆菌10年前应该被接受,因为没有其他已知的解释特定的IgE抗体的存在92年。有趣的是,其他大部分2型cytokine-dependent抗体,免疫球蛋白检测G4 (IgG4)也增加的病人93年。同样,免疫组织化学显示IL 4量表达应承担的肺结核患者的淋巴组织细胞(但不是在结节病)患者组织94年。
为何,这个问题一直有争议吗95年,96年吗?首先,IL 4应承担比IFNγ生物活性较低的浓度,并相应降低mRNA拷贝数,所以方法可靠接IFNγ或其mRNA未能检测到生物显著水平的IL 4。其次,试图提高刺激细胞因子表达的细胞在体外不产生一个准确反映了细胞因子平衡在活的有机体内早期、快速生产IFNγ可以抑制Th2细胞因子释放。最后,先前的研究未能考虑到地理的存在IL 4 splice-variant (IL 4δ2应承担的)。这只变种的IL 4应承担IL 4应承担的活动可能是一种抑制剂,地理和总是coexpressed IL 4,大约在同一水平97年。在肺细胞可能表达水平高于IL 4本身。然而,几乎所有研究IL 4应承担的mRNA水平的引物,扩增信使rna用于IL 4和剪接变体。
2型细胞因子的机制转向一个概要文件
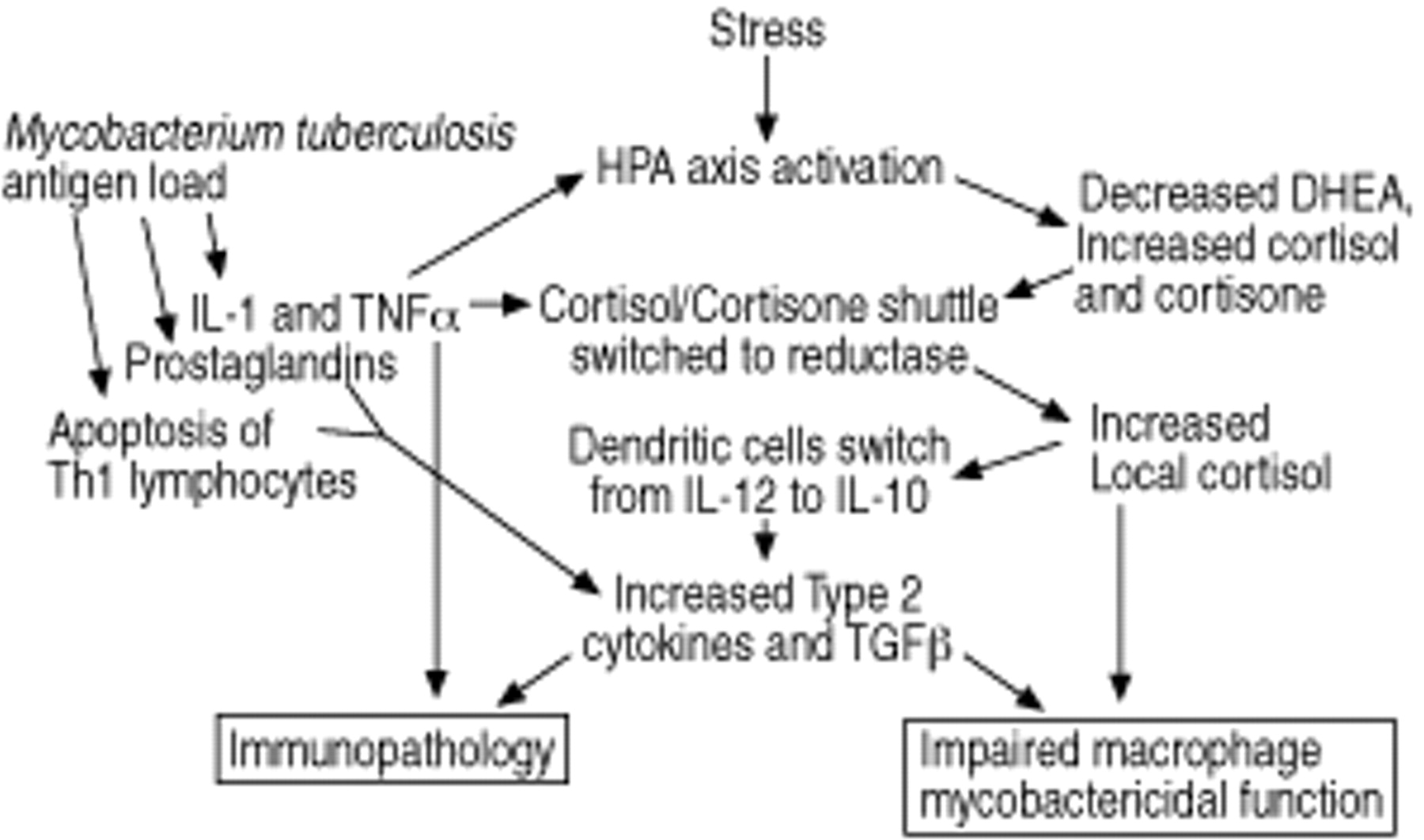
可能是一些相对赤字IFNγ表达式89年和肺结核患者的外周血lymphoproliferation将封存antigen-recognising细胞的淋巴结98年或站点的疾病99年。然而,这并不能完全解释这种大规模上升2型细胞因子的表达89年。那么,细胞因子是这种转变的可能的原因?几个可能性是如图2所示⇓。增加抗原负载可能是一个因素,因为Th1 / Th2平衡是明显与剂量当大量颗粒抗原如分枝杆菌One hundred.或利什曼虫101年。在某些人群中,过度或不适当的接触环境分枝杆菌可能启动一个2型应对至关重要的共同抗原。老鼠这种机制显然是显而易见的,它可以大大增加敏感性,并建议,但不是决定性地证明,在人102年,103年。
在活跃的感染,Th1类T细胞的选择性细胞凋亡可能是一个因素87年,88年,因为可能前列腺素释放104年,105年和增加转化生长因子的分泌β(TGFβ)和IL 10106年。然而,后者可能转向2型配置文件的结果而不是原因。
终于有了强有力的理由表明内分泌与免疫系统相互作用是重要的。维生素D3,皮质醇新陈代谢和脱氢表雄酮水平在内分泌学部分稍后讨论。
肿瘤坏死因子(TNF)的保护作用
这种促炎细胞因子在小鼠疾病防护或不利影响,同样可能是真实的人,稍后讨论与免疫病理反应。它似乎依赖于其他细胞因子的影响。在鼠标,肿瘤坏死因子(TNF)在感染早期保护。肿瘤坏死因子水平升高的早期(3天)对小鼠感染通过气管内的路线,到达第二个峰值在第三周,与成熟的肉芽肿形成107年。肿瘤坏死因子受体基因敲除小鼠屈服更快结核分枝杆菌感染控制老鼠108年和肉芽肿的中断响应和分枝杆菌增加负载而著称结核分枝杆菌来华的老鼠当TNF生物活性被阻塞109年。
巨噬细胞功能和结核分枝杆菌
吸收的分枝杆菌
分枝杆菌被多个通路包括补体受体和甘露糖受体110年,111年。然而,这显然不是故事的全部,因为在体外,结核分枝杆菌可以输入一个各种各样的细胞不表达这些受体112年,113年。准确的吸收方式必须影响随后的细胞内的命运114年。
toll样受体
大部分的初始活化和细胞因子反应的巨噬细胞分枝杆菌可能是由交互toll样受体(通常)。这些都是地理IL 1受体家族的成员,人数有关,分子参与了果蝇的先天微生物耐药机制。内毒素激活细胞CD14和通常的互动,和一些分枝杆菌lipoarabinomannan (LAM)准备工作同样,尽管可能涉及到不同的TLR115年。然而毒性和减毒株结核分枝杆菌可以激活TLR-dependent方式没有膜结合或可溶性CD14要求。TLR2,但不是TLR4可能带来响应从快速增长的分枝杆菌LAM孤立。相比之下,林隔绝结核分枝杆菌或杆菌Calmette-Gue (rin未能诱导TLR-dependent激活。因此,必须有其他组件与TLR可溶性和细胞wall-associated分枝杆菌因素有关。可溶性热稳定和protease-resistant因素被发现调解TLR2依赖激活,而一个热敏感细胞相关分枝杆菌因子介导TLR4-dependent激活116年。脂蛋白可以激活通过TLR,几个结核分枝杆菌以这种方式将推动地理IL 12生产吗117年,也许解释了后者的结果。
Mycobactericidal机制内巨噬细胞
抑制或杀死的结核分枝杆菌很容易诱发小鼠巨噬细胞的接触IFNγ,但这种影响是极其困难的演示令人信服地在人类细胞118年,119年。成功是用人类肺泡灌洗巨噬细胞暴露于TNFα报道在体外120年。这是有可能的,虽然一点也不确定,主要杀伤机制不是一个激活巨噬细胞的直接作用,而是一个事件发生在某些类型的细胞凋亡或杀害themacrophage期间由“注入”granulysin和细胞毒性T细胞穿孔素(参阅CD8部分+效应细胞之前)(图3所示⇓)。
活性氧和氮中间体
如果巨噬细胞做自己杀死结核分枝杆菌这些都是可能造成机制(图。3⇓)。抑制剂的产生一氧化二氮(NO)加重肺结核感染的死亡率,评估细菌负担,和组织病理学121年,122年。的作用机制无是不确定的,因为它有重要的信号和第二信使的功能可能是一样重要的直接毒性的生物123年。此外,KO小鼠不能没有或其他活性氮中间体(RNI)没有增加扩散结核分枝杆菌在肺部感染,直到很晚,但是有增加脾脏的增长。相比之下,KO小鼠,无法使活性氧中间体(ROI), hadincreased肺部细菌的增长。有趣的是,由IFNγ激活巨噬细胞在体外控制扩散结核分枝杆菌是依赖于RNI而不是ROI,所以似乎并行免疫脾肺而不是124年。然而形势依然复杂,在一个模型m . avium感染,摧毁了诱导一氧化氮合酶(间接宾语)基因实际上改善脾清除细菌。这可能是由于这样的事实:在小鼠脾脏,没有水平能达到免疫抑制的水平125年。没有在人的作用仍不清楚。这可能是由适当地激活人类巨噬细胞126年,但从未在小鼠巨噬细胞的大量释放。因此人类细胞释放出来的水平可能留在有益的抗菌范围127年,128年虽然没有普遍的协议的anitmycobacterial相关性机制129年。有证据表明,1,25 (OH)2D3的激活可能参与伊诺人类单核细胞细胞系130年这可以解释animycobacterial这种材料的影响118年。
巨噬细胞凋亡
叶参与人类结核性肺部分被发现含有比noninvolved叶巨噬细胞凋亡131年。然而,大多数工作基于反应的细胞凋亡在肺结核分枝杆菌感染的细胞培养模型。感染致命的结核分枝杆菌减少生存健康的人类肺泡巨噬细胞(heat-killed分枝杆菌相比),和抑制TNFα可以部分逆转132年,133年。细胞包含结核分枝杆菌明显更敏感TNFα杀死的吗113年。
被感染的巨噬细胞的死亡可以被包含的分枝杆菌和死亡联系在一起(图3所示⇓)。然而,有人建议,杆状细菌的数量减少,只有通过感染单核细胞的凋亡,而不是死亡的坏死模式134年,135年。三磷酸腺苷(ATP)诱导细胞凋亡促进杀害的毒性结核分枝杆菌在人类巨噬细胞136年,137年在Fas配体诱导细胞凋亡134年,氢peroxide-induced细胞凋亡也导致mycobactericidal效果138年。除了新鲜的未感染的自体巨噬细胞凋亡的文化m . avium来华的巨噬细胞抑制细菌增长90%。细胞凋亡也防止释放细胞内的组件和隔离分枝杆菌感染的病原体的传播在凋亡的身体139年。
逃避抗菌功能的巨噬细胞
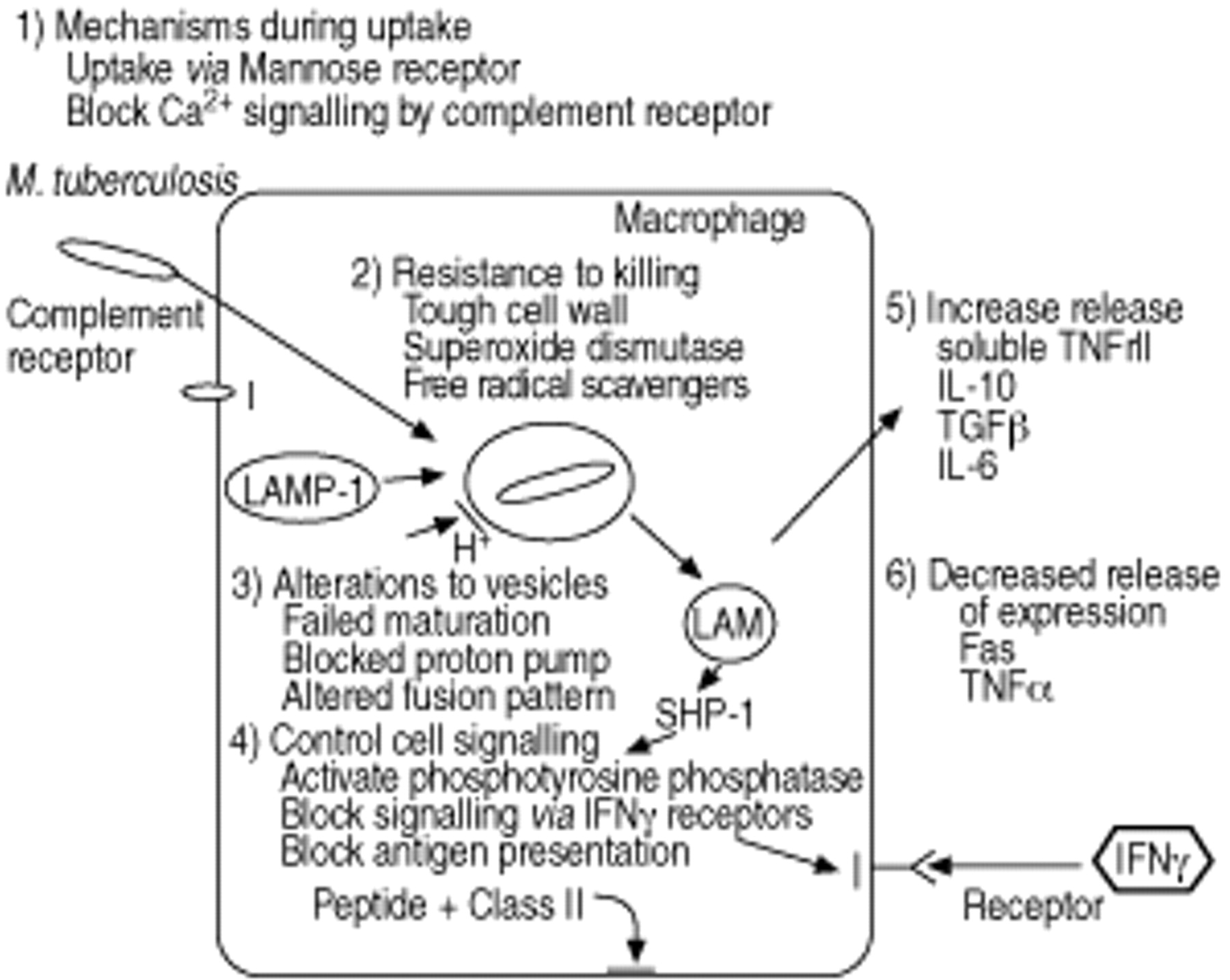
分枝杆菌有各种各样的策略来避免被吞噬细胞(图4所示⇓)140年。结核分枝杆菌将会是通过甘露糖受体无法触发杀人事件。它还能抑制complement-receptor-mediated Ca2 +信号,这可能导致的失败死亡机制141年。分枝杆菌能抑制吞噬体的酸化142年和修改细胞内液泡的贩卖,所以他们表现得像endosomal回收舱的一部分,而不是有毒吞噬溶酶体143年。这些液泡释放大量的LAM插入glycosylphosphatidylinositol细胞膜(GPI)丰富的领域144年。GPI LAM本身就是一个不寻常的多糖结构,能够修改大量的巨噬细胞功能包括IFNγ的反应能力,和现在的抗原的能力了144年)。最后一点可能相关的某种明显的无能的长期mycobacterium-infected巨噬细胞呈现CD4抗原+T细胞145年。林所使用的机制之一可能是由于激活的蛋白质酪氨酸磷酸酶轴马力高1 phosphotyrosine磷酸酶,密切参与细胞信号通路146年。
从被感染的巨噬细胞释放细胞因子变化的模式,这样减少巨噬细胞活化,和T细胞招聘受损(图4所示⇓)。招聘Th1淋巴细胞IL 12应承担的生产,需要由生产的增加抑制TGFβ和IL 10106年,147年,148年,和IL 6释放可能也是一个因素。TGFβ和IL 10还影响巨噬细胞杀菌剂的功能和IL 10应承担有助于增加释放肿瘤坏死因子受体高2块TNFα的激活作用149年。
如前所述,某些类型的细胞凋亡减少所包含的分枝杆菌的生存能力。已经指出,释放可溶性的2型肿瘤坏死因子受体(sTNFRII)毒性菌株引起的结核分枝杆菌可能限制感染肺泡巨噬细胞的凋亡的死亡,他们也减少了Fas表达也可能限制这种途径的凋亡诱导134年。这使得一些研究者假设致病分枝杆菌可能是调节宿主免疫反应减少巨噬细胞凋亡(图4所示⇓)。反对这一假说,是被感染的巨噬细胞也可能抑制mRNA的表达地理bcl 2,抑制细胞凋亡的人131年。可能有机体是优先诱导的细胞凋亡形式离开生物安然无恙。
免疫病理
的毒性结核分枝杆菌
生活结核分枝杆菌本质上是有毒的细胞。例如,人类和小鼠巨噬细胞摄取超过5生物通常死去,而m . avium压力或麻风杆菌可以将细胞内的数字没有杀害他们。有人建议结核分枝杆菌可能会产生脂质产生的毒素相似吗溃疡分枝杆菌(布鲁里溃疡的原因)150年。同样清楚的是,结核分枝杆菌释放的因素大大增加受感染细胞的敏感性TNFα的毒性可能是存在于所有结核性损伤113年,151年。
虽然结核分枝杆菌显然有一些固有的毒性,这并不完全解释疾病的病理学。或结核菌素纯蛋白衍生物(产后抑郁症)都非常无毒在活的有机体内和在体外,但是在准备适当的人类或动物会引起坏死,这显然是由于免疫病理反应。
科赫现象;疾病的响应特性
正如前面介绍的,科赫14指出建立因为豚鼠感染后4 - 6周,皮内挑战整个有机体或本地文化滤液导致坏死,在原来的结核性损伤。类似的现象发生在人类身上。结核菌素试验经常坏死的实验对象是谁,或者,结核性。这不是一个必然的结果,延迟超敏反应结核菌素反应因为坏死不积极对结核菌素皮肤测试时出现了在正常BCG收件人,或类似结节的麻风病人。此外,科赫公司试图利用这一现象治疗肺结核,并发现注入大量的培养滤液(旧结核菌素),皮下注射到肺结核患者,在建立结核性唤起坏死病变在遥远的地点152年。这导致坏死、脱落和“治愈”的病变皮肤结核病(寻常狼疮,通常由牛菌株),但当类似坏死在深在脊柱病变或诱发肺,结果是灾难性的,只是提供了进一步坏死组织中细菌增殖。这种治疗因此抛弃了。
突出显示的错误在科赫公司的想法是在1940年代。因为豚鼠preimmunised时,他们有强大的科赫现象在小剂量的结核菌素反应,他们变得更容易比nonimmunised结核病控制的动物15。这显然被认为只有在挑战进入肺部感染,或深肌内注射,坏死,导致脱落的受感染的组织。
科赫和施瓦茨曼氏反应现象之间的关系
科赫公司发现可溶性细菌物质可能引发坏死在遥远的结核性与施瓦茨曼氏网站有一些相似之处153年进一步加强平行反应和随后的实验。例如,endotoxin-rich材料注射到一个遥远的网站(而不是使用的结核菌素科赫)也将引发结核性坏死病灶154年- - - - - -156年,注射进了旁边的另一个细胞因子触发,胞壁二肽(MDP),导致网站坏死炎症由于完全弗氏佐剂157年。这些观察兼容视图,结核性损伤易受叠加cytokine-mediated损伤。
分枝杆菌病变的cytokine-sensitivity老鼠
还有一种可能性,失败的一个重要的监管作用的γ/δT细胞,这可能导致更大的组织破坏82年。然而,更好的为特征的解释这些发现是增加易感性cytokine-mediated组织坏死组织发生炎症同时由1型和2型细胞因子介导的。这很容易在Balb / c小鼠肺结核158年。在前3周,潜孔网站不敏感的地方注射1μg TNFα重组。这是一段1型反应159年。50天后,慢性疾病的动物进入一个阶段伴随着增加Th2细胞因子生产视为IL 4阳性细胞病变。在这些动物中,潜孔成为TNFα-sensitive网站30.,One hundred.。这对TNF毒性倾向的白介素4应承担的支持使例子nonmycobacterial模型。劳伦斯和同事160年,研究旋毛虫感染小鼠,发现肠病引起的肿瘤坏死因子取决于IL 4。其他小鼠研究也显示,Th2细胞maymediate局部组织炎症IL 4应承担的依赖161年在某些小鼠品系,潜孔引起的Th2细胞与肿瘤坏死因子生产的细胞162年。如果重要组件的科赫现象iscytokine-mediated损害网站的混合型1 / 2型炎症,科赫公司的“治疗”的毒性结核病是容易解释(图。2⇓)152年。
内分泌学
肺结核有几种内分泌和代谢变化可能导致的失败1型反应控制感染,和增加2型细胞因子表达水平。
维生素D3新陈代谢肺结核病变
肺结核患者的巨噬细胞,激活IFNγ,表达一个活跃1α羟化酶,并迅速转化为25 (OH)量的维生素D3对骨化三醇118年,167年。这是一个强有力的现象,导致偶尔泄漏骨化三醇到外围,血钙过多,虽然过去一直很难理解其在疾病中的作用167年。现在看来,这是一个反馈机制,抑制Th1和增强Th2反应,因为骨化三醇抑制生产IFNγ和IL 2和增加生产IL 4和IL 5168年,169年。这也许是与骨化三醇的抑制能力释放IL 12170年。真正的生理相关性这些影响在活的有机体内仍是未经证实的,但小说的合成类似物的骨化三醇用更少的倾向于导致血钙过多使他们测试Th1反应的抑制在活的有机体内模型。这些类似物将延长同种异体移植物存活率,降低环孢菌素一个要求171年。
在1940年代试图与维生素d治疗肺结核患者皮肤结核病(寻常狼疮,通常是由于牛结核分枝杆菌)是处理这种维生素,长期无法愈合肉芽肿病变通常接受坏死决议紧随其后172年。然而,坏死和液化也发生在脊柱病变和肺173年,所以结果是灾难性的科赫的免疫疗法的使用前面描述的152年。这种效应的机理仍然未知,但增加2型细胞因子表达granulomata富含1型细胞因子和TNFα预计将导致坏死性免疫病理(如前所述)。
肾上腺类固醇在肺结核
压力的影响
糖皮质激素导致开关2型细胞因子的生产174年,175年对树突细胞的影响,可能是因为,少分泌IL 12和更多IL 10在他们面前176年,177年,但糖皮质激素也直接把一些2型细胞因子的影响178年和抑制抗巨噬细胞的影响179年,180年。因此不足为奇活化或恶化感染肺结核是敏感的糖皮质激素治疗和hypothalamopituitary肾上腺轴的激活。揭露人类战争和贫困的压力3或牛的交通压力足以引起活化的疾病。诱发疾病的效果更加可控条件下的压力已经证明在老鼠身上181年,182年。
激活hypothalamo-pituitary-adrenal轴(HPA)在感染的老鼠
在老鼠身上感染了致命的结核分枝杆菌气管的路线,有HPA轴的早期活动,相关的启动开关从“纯”1型混合型1 / 2型肺的渗透183年。尽管这个协会是间接的,它可能是重要的治疗与antiglucocorticoid类固醇脱氢表雄酮(DHEA)或androstenediol密切相关,可以推迟,甚至扭转这个开关向2型细胞因子,而皮质甾酮补充剂在生理层面上提高细胞因子的变化183年,184年。
HPA轴在人类肺结核
有很多猜测HPA轴功能的变化在人类肺结核,包括申请一个晚上几乎全部损失的糖皮质激素槽185年和小程度的肾上腺机能不全了挑战与促肾上腺皮质激素(ACTH)186年。最近的研究表明,许多之前报道发现文物,消失如果病人可以适应压力的医院环境前几天测试执行。在这种情况下的昼夜节律是正常的,所以是肾上腺皮质释放的肾上腺激素的反应(CRH)和非常低剂量(即。ACTH的生理)187年。24 h应承担的皮质醇总产量可能正常或略有提高。更严重的疾病,患者的24 h应承担的输出脱氢表雄酮的代谢产物可能减少,针对antiglucocorticoid,和Th1促进类固醇的影响上面所提到的,这可能导致免疫功能障碍188年。然而,最引人注目的和一致的代谢异常是一种变化模式的皮质醇,指示一个大改变的平衡点cortisol-cortisone航天飞机,我们将在下面进行讨论188年。
cortisol-cortisone航天飞机的失调
局部组织的主要机制调节皮质醇水平的互变现象是活跃的皮质醇(11公/羟基)和不活跃的可的松高酮区域(11)。从而有效的皮质醇浓度在不同器官可能非常不同于血清中发现的值。此外,这些酶是监管。地理作为一个例子,颗粒细胞表达11βhsd 1在某个阶段的排卵的周期(luteinising)所以当时可能是敏感的可的松(转换后皮质醇,11βhsd 1)和皮质醇,而在其他时间周期(nonluteinised)细胞表达只有11βhsd 2把应承担的皮质醇不活跃的可的松,所以不会类固醇敏感189年。气相色谱和质谱显示显著超过皮质醇,代谢物的相对于代谢物可的松的24 h尿收集应承担的肺结核患者188年。这种不平衡在治疗恢复正常。研究结果进一步支持了观察肺结核患者更快地转换口服可的松的加载到皮质醇(以等离子体)比控制个人或治愈的肺结核患者190年。
后续研究结核性小鼠和肺结核患者的肺泡灌洗样本的分析和控制,显示异常的不活跃的可的松转换成活跃的皮质醇的病人,是感染肺本身187年,190年。这可能是解释的观测TNFα和IL 1β都增加了表达水平和还原酶活动11βHSD量1细胞系在体外191年。然而,还原酶相对增加的活动也可能是由于减少11βhsd还是2的活动,因为它已经变得很明显,最近这种酶存在于肺192年。进一步enzymological和定量rt - pcr研究是必需的。无论酶,结果是一个当地的皮质醇水平没有明显增加血清皮质醇的测量。这皮质醇过量会导致转向2型细胞因子表达,失活的抗巨噬细胞的影响,增加IL 10和TGFβ增加,所以它可以占很多人类疾病的变化。
疫苗接种
BCG接种疫苗是非常安全的193年。然而,保护效果从80%变化在不同人群保护不保护了194年)。卡介苗是大多数预防结核性脑膜炎,在某些环境中,进步的主要疾病,但它是不那么有效活化或再感染。这变化似乎没有由于使用不同批次的波士顿咨询集团,或种群之间的遗传差异。自BCG是动物的“黄金标准”工作,几乎总是比实验疫苗在动物模型中,尽管其不足的人,需要理解这个变化之前进行漫长而昂贵的试验新型疫苗在人是显而易见的。三个假设目前正在接受调查。
环境分枝杆菌的干涉
无处不在的环境腐生菌的作用已经探索了许多年102年。明显减少BCG的功效可能会因为环境腐生菌本身保护,或因为他们启动有害的反应模式(如。2型),因此阻塞BCG的功效。这两个效应被认为是发生在不同的国家102年,很容易证明实验的影响30.。腐生菌之间的共同抗原的至关重要的作用结核分枝杆菌这些假设的核心,后来完全解释。这些机制受到审查和数学建模103年。
并发寄生虫感染
一些作者认为,伴随的寄生虫感染的存在可能会导致系统性偏向Th2反应破坏了卡介苗诱发Th1反应能力分枝杆菌抗原。
疫苗剂量
另一个建议是,波士顿咨询公司将更可靠,如果在一个非常低的使用剂量,因为疫苗的剂量开始唤起一个有害的2型组件可能是一些人比其他人要低得多。这个想法来源于疫苗研究利什曼虫在不同品系小鼠,剂量,唤起1型反应在某些压力太高,所以2型高诱导101年。如果有人来说,标准BCG剂量过高,从理论上讲,这个问题避免通过使用剂量很低。因为BCG活疫苗,它应该仍然是有效的。在每个个体应该增殖的水平mycobactericidal Th1反应诱导195年。在鹿,5×104或5×107保护,但5×10吗8是不那么有效,所以有这个想法显然是有些道理196年。一项研究男人只用在体外参数(IFNγ释放和淋巴细胞增殖)比较低(1.6×105集落形成单位(cfu)和3.2×106(1.6×10 cfu)标准8cfu),或高(3.2×108cfu)剂量,得出的结论是,超过10的剂量8是必要的197年。然而,如果没有保护的研究,这些数据无法解释。这些参数之间的关系和保护仍然是模糊的。在最近的一次研究中,BCG疫苗接种引起的PPD皮肤试验-主题皮肤试验积极性,但没有影响在体外lymphoproliferation或细胞因子的生产198年,皮试反应和保护之间的关系同样模糊。
脱氧核糖核酸疫苗
最近有很多DNA疫苗的兴趣。这种疫苗接种方法常常导致抗原T淋巴细胞分泌IFNγ并显示细胞毒性的潜力,可取的肺结核的因素。这可能是相关的辅助属性nonmethylated CpG序列199年。许多抗原已经在动物模型研究,特别是分泌蛋白200年和热休克蛋白47稍后讨论,但是,这种动物模型的相关性对人类患者难以确定。
使用动物研究试图确定保护性抗原
巨大的努力已经在搜索“保护”抗原,相信特定子集的抗原或抗原表位将被证明是最优目标保护性免疫反应。几乎所有这些研究都涉及测试纯化或重组抗原在结核病小鼠模型,在各种各样的佐剂,或表达牛痘或沙门氏菌201年。其他修改BCG,希望增加其免疫原性和其诱导CD8的能力+细胞的反应52。显而易见,这种方法的有效性是有限的。基本上所有的蛋白质抗原结核分枝杆菌测试小鼠模型将保护如果他们管理的方式诱导分化Th1反应。同样,所有结核抗原尝试,似乎作为DNA疫苗防治肺结核在老鼠身上202年和复杂的实验系统功效,不允许有轻微的变化看作是归因于内在差异“保护”的角色。一些抗原只工作后如果选择最佳的辅助一个反复试验的过程。计检测6是一个例子,说明了这些实验带来的困境。这是一个占主导地位的T细胞目标人类与动物在早期的肺结核203年。但早期T细胞识别计检测6表明,这是一个保护性抗原,或它是一个失败的标志反应和发展疾病?老鼠实验肯定不帮助。计6将会保护老鼠如果使用合适的复杂的佐剂,但保护不太容易实现与其他抗原如hsp65或30 kda组mycolyl转移酶204年。
如何,因此,应该选择抗原的疫苗试验的人,辅助应该使用什么?人类研究非常困难,只有一个子集的成功概率高的抗原可以进行临床试验。一个建议是另一轮的测试在几内亚猪模型。然而,没有理由认为这将推迟决策天以上。
通过人类研究保护抗原的识别
研究反应接触那些没有疾病
克隆从自然PPD-converted个人显示一系列的反应,有些特定的结核病,承认所有分枝杆菌物种测试205年。因此,为了发现一些抗原是否有专门的保护作用,有必要跟进个人最近暴露于肺结核,所以那些之间的任何差异,也不会发展成活动性疾病可以被识别。这些研究进展3地理上不同的非洲国家,通过项目由欧洲共同体。
常见的分枝杆菌抗原和物种特异性抗原表位
多年来它已经知道BCG一样有效的麻风疫苗防痨,虽然麻风杆菌是一个完全不同的物种206年。BCG因此必须能够通过共同的抗原表位。同样有证据表明与环境生物接触,导致分枝杆菌皮肤试验积极性,保护马拉维的人口结核病和麻风病207年。在小鼠,强大的保护或“anti-protective”效应可以与环境分枝杆菌诱导腐生,仅仅通过改变免疫协议,导致1型或2型反应30.。这些影响显然是由于共同的抗原。同样重要的一点是,肺结核病人仍然保持坏死性皮肤试验抗原的积极性结核分枝杆菌本身,已经减少或没有皮试反应环境腐生菌208年保护人群的,而这些做唤起反应。这是一个了不起的悖论,这意味着缺乏应对共同抗原可能与对疾病的易感性。尽管这些事实,但有一个很深的偏见的观点可以介导的保护通过不种特异的抗原表位。这种偏见可以追溯到早期的时代抗体介入疫苗,因为绑定构象表位抗体中和微生物组成的毒素,酶或粘附分子。这些物质往往species-restricted。T细胞没有任何抵消,但承认短肽序列从微生物蛋白质裂解,连同这一事实T细胞不分类,应该足以驱散偏见。种特异性T细胞功能无关的概念。同样重要的是要记住最初制定的克隆选择学说伯内特109年现在是一个过时的概念。远离选择曲目不认可自我的T细胞,胸腺选择曲目基于识别,虽然弱,肽来源于自我的。因此,随后的T细胞能够识别细菌严重偏向保守蛋白质的细菌版本,还存在于男人80年。
热休克蛋白的作用
热休克蛋白(hsp)是重要的这一概念的例子。事实上65 kda热休克蛋白麻风杆菌可以保护小鼠免受结核分枝杆菌209年可以根据其序列DNA疫苗。然而,有一个报告,在几内亚猪应承担至少与hsp可能导致肺部免疫病理、免疫自身免疫的暗示,当触发随后气溶胶感染结核病210年。保护性免疫的重要性守恒的蛋白质,如热休克蛋白已经被别人反复强调211年。不仅是hsp重要目标抗原在他们自己的权利,但是他们也有一个基本的监管职责。因此hsp的可以作为佐剂,用作纯化蛋白,或当他们的编码序列是纳入DNA疫苗,编码,例如,乳头状瘤病毒抗原E7212年。他们可能作为一个危险的信号。这是极大的兴趣,在感染单核细胞增多性李斯特氏菌有增加膜的表达哺乳动物hsp60, hsp65的同系物213年。
未来
免疫疗法
这也许是工人面临的最重要的问题领域的肺结核。正如前面介绍的,点帮助,但未能解决问题,以及耐多药疾病越来越威胁。免疫治疗是唯一的解决方案,这种方法需要以来一直认可的r·科赫的时间214年。
一个潜在的免疫治疗方法是患者细胞因子的直接使用,系统,或由气溶胶。地理IL 2 IFNγIL 12和GM CSF应承担的所有被调查了215年)。他们潜在的角色在治疗中,除了作为一个潜在的辅助药物治疗在抗多种抗菌素的情况下,尚未阐明。
出惊人的结果在小鼠体内的另一种方法是使用脱氢表雄酮或androstenediol非常相似。这些化合物反对的一个子集糖皮质激素的影响,并且可以扭转开关对2型细胞因子在Balb / c小鼠(图5所示⇓),但这并没有测试的人184年。
抗原免疫疗法不太可能通过简单的准备,因为需要有一个免疫调节组件,2型细胞因子的差别,对这些已存在的生产。这或许可以解释为什么抗原准备工作,使用时可以预防结核病疫苗在挑战之前,不可以作为治疗的小鼠模型217年。
有趣的是,唯一的准备工作证明是有效的免疫治疗结核性老鼠被杀死分枝杆菌衰弱(图5⇓)216年,218年最近,分枝杆菌抗原DNA疫苗编码常见219年。
尝试使用环境腐生物免疫疗法,衰弱等的影响,然后研究生物对疾病易感性和BCG接种疫苗的功效102年一个概念,是现在被广泛接受的103年。自衰弱不包括种特异的抗原表位,诱导坏死性科赫现象是避免,而多个共同抗原的保护效果(包括hsp)可以利用。两项研究的单一注射heat-killed衰弱进行了GoodClinical实践(GCP)在人类肺结核患者还接受点。结果是矛盾的220年,221年。
针对不可避免的困难证明效果叠加在点单剂量时,有一个明确的需要多药物耐药性疾病,多剂的试验,用于飞行员non-GCP研究耐药结核病222年。衰弱使用适当的属性,因为,尽管它死亡,不仅唤起1型反应呢223年和细胞毒性CD8+T细胞杀死靶细胞感染结核分枝杆菌224年,但也作为一个强有力的非特异性downregulator预先存在的Th2反应在老鼠225年GCP的研究,在人类哮喘患者226年。它也可以导致艾滋病毒Th1共同分枝杆菌抗原的识别+个人,这可能证明是一个重要的发展中国家的需求227年。
创建可靠的诊断测试
有很多评论的尝试产生一个可靠的血清学检测结核病228年。不存在这样的测试目前由于双重问题无处不在的环境背景可交叉反应的抗体诱发的分枝杆菌(BCG),在一些国家,与个体差异的特定抗原表位的认可。淋巴组织或IFNγ应对某些低分子量文化滤液抗原可能区分BCG接种疫苗和tuberculosis-infected个人229年现场使用,但这样的测试是麻烦了。
基因组序列的剥削
1998年的4411529个碱基对结核分枝杆菌基因组发表230年。在3924个蛋白编码基因中,只有40%的人强烈与其他已知的蛋白质,使绝大多数并没有明确的功能。这个序列已经允许的重大进展231年。一个例子是四个酮化合物合酶的识别系统。这些没有已知的目的,但这些途径的产物可能是脂质产生的毒素类似溃疡分枝杆菌(布鲁里溃疡的原因)150年。其他作者分析了3924个蛋白质编码序列推导的结核分枝杆菌基因组,发现52蛋白质携带一个aminoterminal分泌信号肽,但缺乏额外membrane-anchoring半个。其中52蛋白质,只有7之前报道称分泌蛋白232年。分泌蛋白质是好的候选人保护性抗原。
允许进一步的基因组测序与非病原的致病分枝杆菌的比较,和毒性outbreak-causing菌株(如。“奥什科什”应变)毒性菌株较少。这可能加速识别重要的发病机制。类似的微阵列的发展促进了BCG菌株的基因分型和比较233年。
基因调控的评价,结合分析的基因组数据库,允许识别的基因,或抑制在应对环境和其他刺激。例如基因引起的接触发现了异烟肼234年。
基因组也揭示了一些重复的DNA序列,以前不明,这些可能是有用的分子类型。

)定量逆转录酶聚合酶链反应使用新鲜如果外周血monuclear细胞从tuberculin-positive结核病患者(▪)和健康的捐赠者(□)与年龄、种族和性别,表明尽管信使核糖核酸(mRNA)拷贝数为1型2型细胞因子,细胞因子仍高于肺结核的主要变化是增加2型细胞因子表达(白介素4 (IL 4)应承担的,应承担p = 0.004和白介素13 (IL 13)应承担的由Mann-Whitney测试分别p = 0.0009)。中位数显示;第25和第75百分位数标记为错误。b):地理IL 4 mRNA拷贝数与血清免疫球蛋白E (IgE)应承担的mRNA水平暗示这是翻译成活跃的细胞因子。▪:肺结核病人;▵:健康对照组。改编自89年。

可能的机制,可能导致模型2型细胞因子的过度激活在人类肺结核。这些会削弱mycobactericidal巨噬细胞的功能,与肿瘤坏死因子α(TNFα),导致免疫病理反应。IL:白介素;HPA: hypothalamicpituitary轴;TGFβ:转化生长因子β;脱氢表雄酮:脱氢表雄酮。

多个巨噬细胞机制试图杀死结核分枝杆菌。尚不清楚这是导致死亡的重要机制结核分枝杆菌,尤其是在人类细胞死亡是非常难实现的在体外。候选人路径所示,所有引用和讨论的文本。注意,只有一些诱导细胞凋亡导致减少含有细菌的生存能力。进一步激活必须发生通过toll样受体,但他们的角色在杀人是不清楚。IFNγ:干扰素γ应承担的;TNFα:肿瘤坏死因子α;ATP:三磷酸腺苷;H2O2:过氧化氢。

结核分枝杆菌避免被巨噬细胞。这个数字应该与图3所示⇑。每一个死亡通路如图3所示⇑反对一些机制在图4吗⇓。粗线表示阻塞通路。大部分的数据是自解释的。吸收通过甘露糖受体可能避免引发死亡机制。包含分枝杆菌的液泡内排序/回收endosomal机械的巨噬细胞和未酸化和酸溶酶体融合。但是他们与溶酶体融合激活蛋白(灯检测1)包含核内体和他们释放lipoarabinomannan (LAM)包含囊泡。因此他们的融合模式改变的病原体。地理轴马力1:phosphotyrosine磷酸酶。TNFrII:肿瘤坏死因子rII;IL:白介素;TGFβ:转化生长因子β;TNFα:肿瘤坏死因子α;IFNγ:干扰素γ。

采用免疫分析细胞阳性)地理白介素2 (IL 2)应承担和b)白介素4 (IL 4)应承担的领域的肉芽肿Balb / c小鼠的肺部感染结核分枝杆菌H37Rv,接受60天的表示方案。12 - 18的随机数据字段(3字段从老鼠每4 - 6)。两种免疫治疗方案造成一个增加IL 2阳性细胞,然后降低IL 4阳性细胞。(□)控制感染的老鼠;d60和d90(○)治疗分枝杆菌衰弱1.0毫克;(▪)治疗从d60×3·周−1androstenediol和生理替代剂量的肾上腺酮。误差线控制点是sd。他们是省略了从测试组为了清晰,但在这些群体规模类似。这两个测试组都显著不同控制从88天(即。从启动免疫治疗后28天(p < 0.01)。改编自216年。
脚注
↵本系列之前的文章:1号:核磁共振Hammerschlag。衣原体肺炎和肺。欧元和J2000;16:1001 - 1007。2号:S。Ewig h·谢弗,托雷斯。在社区获得性肺炎的严重程度的评估。欧元和J2000;16:1193 - 1201。3号:石油醚Nicod J C。Pache:豪。真菌感染在移植受者。欧元和J2001;17:133 - 140。4号上午:琼斯,卷多德A.K.韦伯。伯克不过:当前临床问题,环境争议和伦理困境。欧元和J2001;17:295 - 301。
- 收到了2000年6月20日。
- 接受2000年6月27日。
- ©人期刊有限公司
引用
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}