





文摘
严重的社区和院内肺炎是由嗜肺性军团菌。肺气道和肺泡上皮细胞组成的重要前哨系统机载感染。虽然白介素6 (IL)被称为中央监管机构免疫反应的肺炎、肺广泛的监管不明。
在此,我们证明不同退伍军人菌株诱导延迟il - 6的表达相比,引发了人类的肺上皮细胞。il - 6表达依赖,在早期的时间点,在鞭毛蛋白识别toll样受体(TLR) 5、增殖活性蛋白激酶/细胞外signal-regulated 1和p38激酶激酶(MEK)增殖蛋白激酶(地图),而且,在之后的时间点,iv型分泌系统。以同样的方式,但更迅速,最近描述转录因子IκBζ引起的军团菌核因子感染,绑定(NF) -κB亚基p50——招募白细胞介素6子一起CCAAT-enhancer-binding蛋白质β和磷酸化激活蛋白1亚基cJun。类似地,组蛋白修饰和NF-κB亚基p65 / RelA出现在iκbζ随后在白细胞介素6基因启动子,从而启动基因表达。基因沉默的IκBζ减少军团菌有关的il - 6表达了41%。
总的来说,这些数据显示一系列鞭毛蛋白/ TLR5 -和类型IV-dependent IκBζ表达式,IκBζ/ p50招聘白细胞介素6子,染色质重塑和随后的il - 6转录退伍军人来华的肺上皮细胞。
重症社区获得性肺炎的重要病原体,第二个最常见的病原体检测肺炎在那些承认在工业化国家是重症监护病房嗜肺性军团菌1,2。尽管> 40军团菌物种是已知的,大多数的人类感染所致退伍军人血清组13。退伍军人是一种革兰氏阴性,兼性胞内病原体变形虫的自然和人为水生环境。人类感染后发生气溶胶飞沫吸入被污染的水。承认军团菌由transmembraneous toll样受体(TLR) 2和TLR5和胞质Naip5(可能还有其他未知的受体)似乎激活真核生物的免疫反应4- - - - - -7。
关于退伍军人病机、原生动物的基本结果通过分析感染或巨噬细胞等免疫细胞8,9。然而,尽管军团菌肺上皮细胞内复制有效和最近的研究指向肺上皮细胞作为一种重要的哨兵和先天免疫效应7,10- - - - - -16所知甚少,肺上皮细胞感染的后果军团菌。
白介素6 (IL)浓度在血液和支气管肺泡灌洗液的病人患有肺炎与疾病严重程度呈正相关17- - - - - -19和il - 6启动子多态性与肺炎球菌肺炎的肺外传播20.。研究使用IL-6-deficient小鼠进一步表明肺炎细胞因子il - 6的突出作用21,22。il - 6的影响重要的先天免疫机制,例如,减少中性粒细胞凋亡和增加细胞毒性的能力23,24。它的表达式是由几个转录因子,包括核转录因子(NF) -κB,激活蛋白(美联社)1,和CCAAT-enhancer-binding蛋白(C / EBP)25,26。最近,一个关键的作用诱导核蛋白质IκBζil - 6表达了27。
考虑到il - 6在肺炎的重要作用,我们这里的机制进行了分析军团菌有关的人类肺上皮细胞il - 6表达。退伍军人诱导表达的il - 6 flagellin-TLR5和iv型分泌系统的方式。NF-κB-related表达il - 6表达IκBζ是至关重要的军团菌来华的细胞。军团菌全身的组蛋白修饰和后续招聘的p50和p65 / RelA白细胞介素6和iκbζ基因启动子启动基因转录。在白细胞介素6催化剂,IκBζp50交互军团菌来华的细胞和基因沉默的IκBζ减少军团菌有关的il - 6表达。总的来说,退伍军人鞭毛蛋白激活IκBζ和随后的转录的il - 6的表达在人类肺上皮细胞。
材料和方法
材料
胎牛血清(FCS)和trypsin-EDTA解决方案从生活中获得技术(德国卡尔斯鲁厄)。吡咯烷二硫代氨基甲酸(PDTC)、蛋白酶抑制剂、NP-40,特里同x - 100和Tween-20买来σ化学(德国慕尼黑);从Calbiochem-Merck mg - 132(达姆施塔特,德国)和鞭毛蛋白重组亚历克西斯(德国)勒拉赫市。所有其他化学物质均为分析纯和从商业来源获得。
细胞系
ⅱ型肺泡细胞系A549得到从写明ATCC(罗克维尔市,医学博士,美国)和火腿的F-12培养基培养l谷氨酰胺(PAA实验室、派斯克、奥地利)和10% FCS没有抗生素28。主要的人类小气道上皮细胞(SAEC)获得Clonetics / Cambrex (SAEC系统;美国Cambrex,巴尔的摩,马里兰州)和培养SAEC BulletKit®(Clonetics / Cambrex)根据供应商的指令15。
感染菌株
退伍军人干系人130 b野生型(写明ATCC baa - 74,请提供N.P. Cianciotto,西北大学医学院、芝加哥,美国)29日,JR32野生型30.和JR32δ队伍突变(莱拉·3118,请提供h . Shuman,哥伦比亚大学,纽约,纽约,美国)31日,寇比野生型和寇比δflaA突变体(请提供的k .修,柏林,德国)经常生长在缓冲charcoal-yeast提取(BCYE)琼脂2天在37°C32在上皮细胞中,随后resuspended 37°C。检查了细菌密度确定光密度在660 nm (OD660年),贝克曼分光光度计DU520(贝克曼库尔特,Unterschleissheim,德国)。A549或SAEC (10610)感染5,106和107菌落(CFU)细菌,即。0.1:1-10:1感染复数,每毫升和孵化感染介质(火腿的F-12l谷氨酰胺没有抗生素或SAEC介质)对于一个给定的时间在37°C和5%的公司2。细胞外并不经常使用抗生素杀死细菌。退伍军人上皮细胞培养基上菌株没有明显增长是由串行控制稀释镀BCYE琼脂。对于heat-inactivating退伍军人在95°C,细菌是孵化30分钟14。
il - 6 ELISA
汇合的A549细胞或SAECs表示在湿润的气氛中被感染。孵化后,上层的收集和加工的il - 6量化免疫测定根据制造商的指示(美国研发系统、明尼阿波利斯、MN)。
染色质免疫沉淀反应(芯片)
A549细胞被感染退伍军人显示,然后进行芯片分析如前所述14,28,33,34使用anti-mRNA聚合酶II (Pol II)(在20),anti-p65 (C-20) anti-p50, anti-C / EBPβ(圣克鲁斯生物技术,圣克鲁斯、钙、美国),anti-P-cJun(细胞信号技术,丹弗斯,妈,美国),anti-PSer10/交流Lys14h3, Pan-Ac-H4(美国纽约北部,普莱西德湖)和anti-IκBζ(阿克利抗体GmbH, Hiddenhausen,德国)。的白细胞介素6和iκbζ增强区域放大了PCR使用HotStarTaq聚合酶(试剂盒、希尔登,德国)和特定的引物:白细胞介素6感觉5′-ACAAATTAACTGGAACGCT-3′,反义5′-ATTGGGGGTTGAGACTCTAA-3′iκbζ感觉5′-AGGGGAATGTCCGGGACT-3′,反义5′-TAATGTCTGACCTCGTGGCAA-3′。总输入的PCR扩增DNA在每个样本显示为控制。
Chromatin-IP-IP, DNA被筛选了第一个IP,加ChIP-RIPA缓冲和孵化anti-IκBζ和anti-p50抗体,分别为第二个IP。作为一个控制IP anti-sheep免疫球蛋白(Ig) G (Li-COR)。
免疫印迹
测定IκBζ和cox - 2诱导A549或SAE细胞被感染表示,洗两次,和核(IκBζ,肌动蛋白),膜(TLR5)或完整的细胞提取(ERK2 cox - 2)是收获。细胞细胞溶解在海卫一缓冲区包含NP-40或x - 100进行sds - page,涂抹在硝化纤维膜(Amersham生物科学,弗莱堡,德国)。Immunodetection目标蛋白进行了与特定的抗体:IκBζ(阿克利抗体GmbH, Hiddenhausen,德国)cox - 2(美国圣克鲁斯圣克鲁斯生物技术),随后孵化与二次抗体(IRDye 800 -标记anti-mouse或Cy5.5-labeled anti-rabbit,分别和IRDye 800 -标记抗体)(Li-COR Inc .,糟糕的小礼帽,德国)。同时检测肌动蛋白或ERK2(圣克鲁斯生物技术)通过使用一个奥德赛红外成像系统(Li-COR Inc .)证实蛋白质所述负载相等28,35。
rt - pcr
分析IκBζ,il - 6,引发,粒细胞巨噬细胞集落刺激因子(gm - csf)和GAPDH基因表达在A549和SAECs,总RNA分离RNeasy迷你包(试剂盒)和相对地转录使用moloney小鼠白血病病毒逆转录酶(表达载体,卡尔斯鲁厄,德国)。生成的cDNA IκBζ放大了使用特定引物PCR(转发:5′-TGAATGCACTTCACATGCTG-3′;反向:5′-TTCGTTCTCCAAGTTCCGAGT-3′)、il - 6(转发:5′-TTCTCCACAAGCGCCTTC-3′;反向:5′-TGGACTGCAGGAACTCCTTA-3′),引发(转发:5′-CTAGGACAAGAGCCAGGAAGA-3′;反向:5′-AACCCTCTCTGCACCCAGTTTTC-3′), gm - csf(转发:5′-GTCTCCTGAACCTGAGTAGAGACA-3′;反向:5′-AAGGGGATGACAAGCAGAAAGTCC-3′), GAPDH(转发:5′-CCACCCATGGCAAATTCCATGGCA-3′;反向:5′-TCTAGACGGCAGGCAGGTCAGGTCCACC-3′)。引物都是购自TIB MOLBIOL(德国柏林)。20 - 35放大周期后PCR产品1.5%琼脂糖凝胶上进行分析,与溴化乙锭染色,随后形象化。确认使用等量的RNA在每个实验中,所有样本检查GAPDH mRNA的表达。
RNA干扰
核目标IκBζ(意义:GGAAAAAGGUAAAUACAGCTT;反义:GCUGUAUUUACCUUUUUCCTT)和TLR5(意义:GGAGCAAUUUCCAACUUAUTT;从Ambion反义:AUAAGUUGGAAAUUGCUCCTT)获得。控制non-silencing siRNA(意义:UUCUCCGAACGUGUCACGUTT;反义:CGUGACACGUUCGGAGAATT)从MWG购买(Ebersberg,德国)。通过使用Amaxa A549细胞转染NucleofectorTM根据制造商的协议(NucleofectorTM解决方案V, NucleofectorTM程序G-16;Amaxa、德国科隆)与2μg siRNA / 106细胞。对A549细胞稳定表达非特异性或IκBζ-specific成分,细胞与IκBζnucleofected SureSilencingTMshRNA向量(pGeneClip-Neo)或控制向量(Superarray,弗雷德里克,医学博士,美国),和选择μg·400毫升−1在4周内G418对新霉素抗性。
统计方法
数据意味着±se至少有三个独立的实验。采用配对t效果统计评估。一个假定值<0.05被认为是重要的(如果不是表示,否则,测试执行与控制)。
结果
退伍军人全身的il - 6的表达和IκBζ人类肺上皮细胞
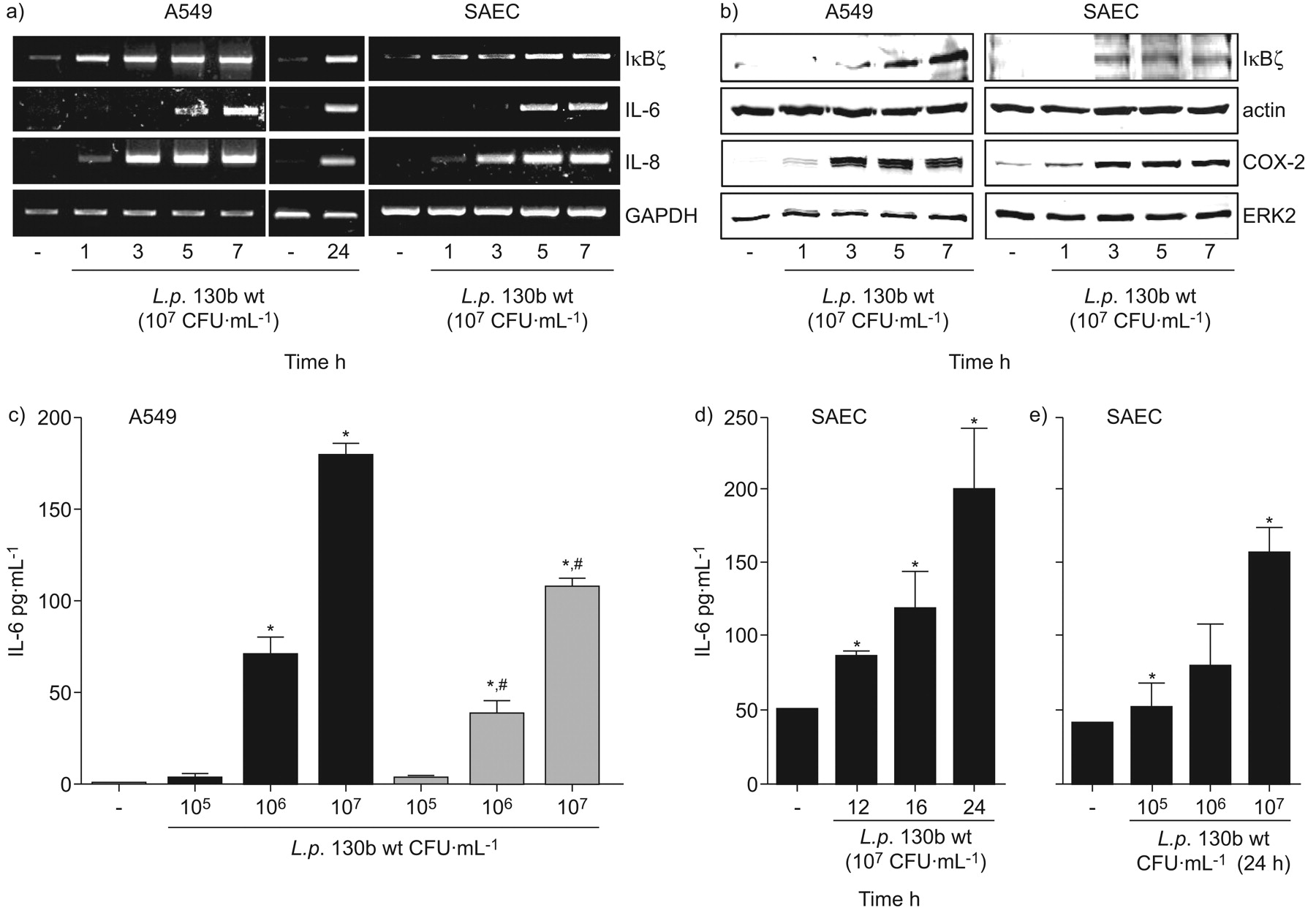
感染人类的肺上皮细胞系A549 (图1一个和图S1A补充材料)退伍军人菌株130 b,应变寇比(图2 a和b)和应变JR32 (图3 a和b)以及主要的人类肺SAEC退伍军人菌株130 b (图1一个补充材料和无花果。印地)诱导il - 6的表达以及引发的信使rna。il - 6蛋白解放所示退伍军人130 b - (图1 c寇比),(图2 c和d), JR32-infected (图3 c)A549和130年b-exposed SAEC (图1 d和e)。Heat-inactivation退伍军人130 b减少,但并没有阻止,解放被感染的A549细胞的il - 6 (图1 c)。
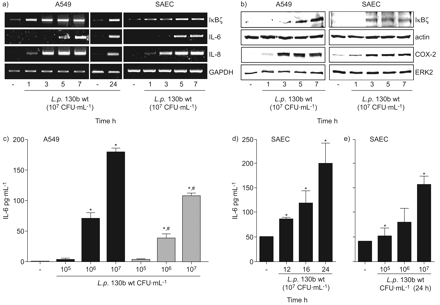
嗜肺性军团菌全身的表达白介素(IL) 6和IκBζ人类肺上皮细胞依赖细菌的生存能力。肺泡上皮细胞(A549)或人类初级小气道上皮细胞(SAEC)被感染退伍军人野生型菌株130 b (107CFU·毫升−1),a, b, d)表示时间或c, e)表示浓度24 h) mRNA水平的IκBζ,il - 6,引发,GAPDH和rt - pcr检测,和b)蛋白表达的IκBζ,cox - 2、肌动蛋白和ERK2被免疫印迹检测。代表凝胶/墨迹图所示的三个独立的实验。(c)肺泡上皮细胞(A549)感染了表示浓度的可行性(▪)或heat-inactivated (▓)退伍军人野生型菌株130 b 24 h和il - 6释放在上层的决定。人类主要SAEC被感染退伍军人野生型菌株130 b (107CFU·毫升−1),d)表示时间或e)表示浓度24 h,并释放il - 6在上层的决定。如果*:p < 0.05与刺激细胞;#:p < 0.05可行与heat-inactivated细菌。
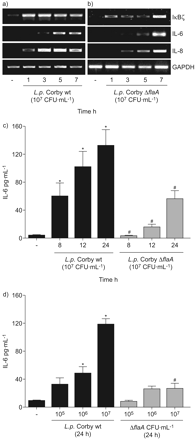
嗜肺性军团菌诱导表达白介素(IL) 6和IκBζ依赖鞭毛蛋白。一)肺泡上皮细胞(A549)被感染退伍军人寇比野生型菌株a)或b)ΔflaA敲出突变(107CFU·毫升−1)表示时间和mRNA表达决心通过rt - pcr。代表三个独立实验的凝胶。A549细胞被感染退伍军人寇比野生型菌株(▪)或ΔflaA敲出突变(▓)(107CFU·毫升−1)c)表示时间或d)表示浓度24 h,并释放il - 6在上层的决定。如果*:p < 0.05与刺激细胞;#:寇比p < 0.05与寇比ΔflaA。
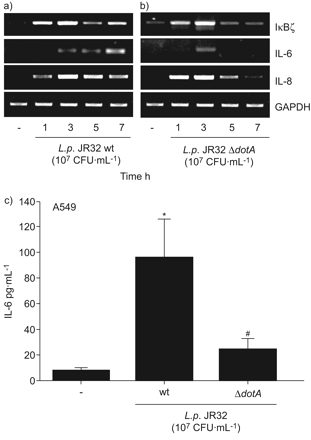
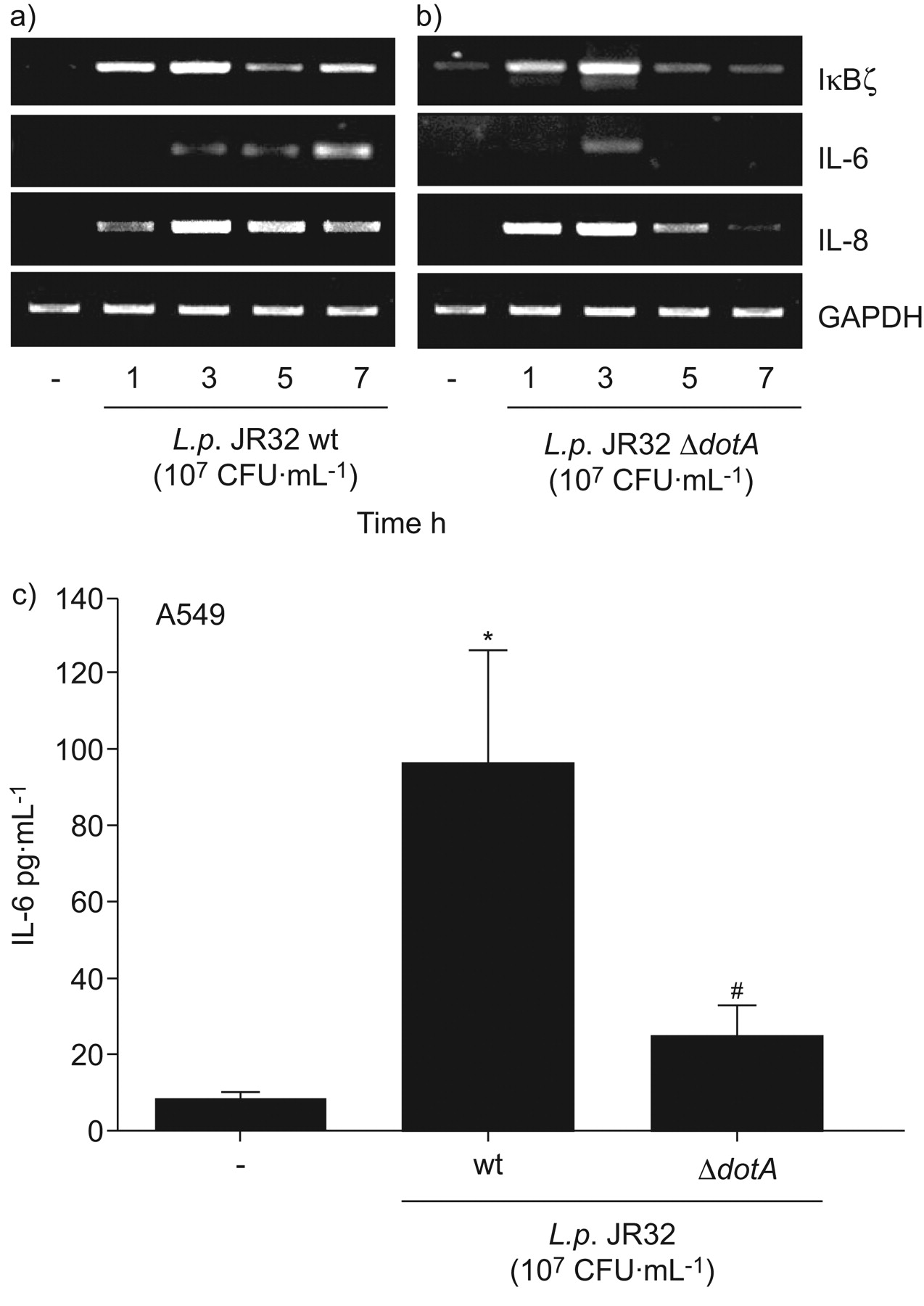
嗜肺性军团菌全身的表达白介素(IL) 6和IκBζ依赖军团菌type-IVB点/ Icm分泌系统。一)肺泡上皮细胞(A549)被感染退伍军人一)野生型菌株JR32或b)ΔdotA敲除突变体(107CFU·毫升−1)表示时间和mRNA表达决心通过rt - pcr。代表三个独立实验的凝胶。c) A549细胞被感染退伍军人野生型菌株JR32或ΔdotA敲出突变(107CFU·毫升−1)24 h和释放il - 6在上层的决定。如果*:p < 0.05与刺激细胞;#:p < 0.05退伍军人JR32与JR32ΔdotA。
观察的mRNA表达il - 6晚于引发,可能作用IκBζ调节蛋白表达的il - 6表达被认为是,和IκBζ表达进行了分析。早期诱导IκBζmRNA (图1一个)和蛋白质(图1 b)观察il - 6 mRNA表达的开始退伍军人130年b-infected A549和SAEC (图1一个在寇比()以及图2一个和b)和JR32-infected (图3和b) A549细胞。信使rna表达IκBζ、il - 6和引发持续24小时(图1一个)。因此,不同的菌株退伍军人诱导,IκBζ和il - 6表达在人类肺上皮细胞在一个特定的时间顺序。
退伍军人全身的il - 6的表达和IκBζ依赖flagellin-related TLR5和激活军团菌type-IVB点/ Icm分泌系统
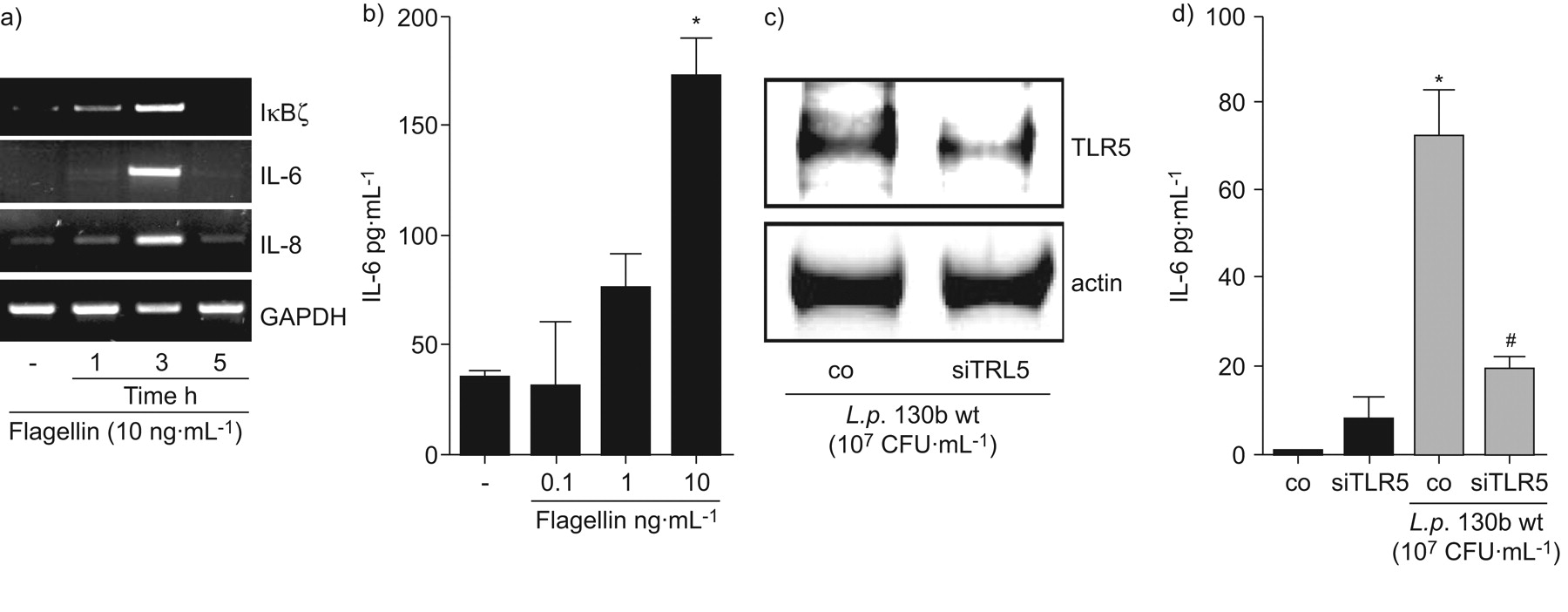
检测军团菌鞭毛蛋白通过TLR5似乎是重要的真核宿主细胞的激活5,14。的确,军团菌缺乏鞭毛蛋白(FlaA)更有效的对mRNA的感应IκBζ和il - 6以及引发的另一个重要的促炎细胞因子在肺炎36,特别是在早期的时间点(图2 b)。此外,解放的il - 6蛋白在细胞感染flagellin-deficient显著降低军团菌与野生型相比,细菌(图2 c和d)。出版以前,FlaA-deficient军团菌复制两次以及野生型细菌在肺上皮细胞24小时但不改变细胞的死亡是由乳酸脱氢酶释放(数据没有显示和Vinzinget al。37)。增加重组鞭毛蛋白细胞培养证明足以诱发IκBζmRNA的表达,il - 6以及引发的人类肺上皮细胞(图4)。最后,补充体内重组鞭毛蛋白诱导il - 6蛋白分泌(图4 b)。抑制内源性TLR5表达的特定核(图4摄氏度)显著降低flagellin-related白介素解放A549细胞(图4 d)。总的来说,这些数据表明TLR5-related检测的关键作用军团菌鞭毛蛋白IκBζ和il - 6的表达。
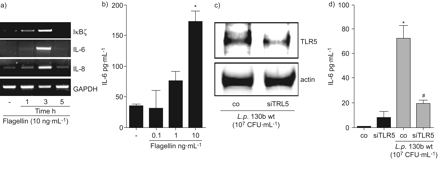
嗜肺性军团菌诱导表达白介素(IL) 6和IκBζ激活toll样受体(TLR) 5。A549细胞被刺激与鞭毛蛋白(10 ng·毫升−1))表示时间和mRNA表达决心,或b)表示浓度24 h,并释放il - 6在上层的决定。A549细胞与未指明的nucleofected(有限公司)或TLR5-specific (siTLR5)核和48 h后感染退伍军人野生型菌株130 b (107CFU·毫升−1)24 h . c) TLR5表达式是由免疫印迹和d)释放il - 6通过ELISA(▓)和没有(▪)退伍军人野生型菌株130 b (107CFU·毫升−1)。如果*:p < 0.05与刺激细胞;#:p < 0.05不具体的与TLR5核。滴在c),代表三个独立的实验。
Type-IVB点/ Icm分泌的胞质识别系统相关的活动被认为是重要的军团菌鞭毛蛋白和随后的细胞活化38。在A549细胞的感染退伍军人JR32突变不足为队伍的一个组成部分军团菌type-IVB系统8,我们观察到一个类似的感应IκBζ,il - 6,并引发mRNA在早期的时间点与感染细胞JR32野生型,但明显减少5 h后(图3解放和b)以及降低il - 6 (图3 c)。总的来说,细胞内,type-IV-dependent识别军团菌鞭毛蛋白似乎是重要的持续感应IκBζ和il - 6的A549肺癌细胞。
NF-κB-related转录因子和IκBζ退伍军人来华的上皮细胞
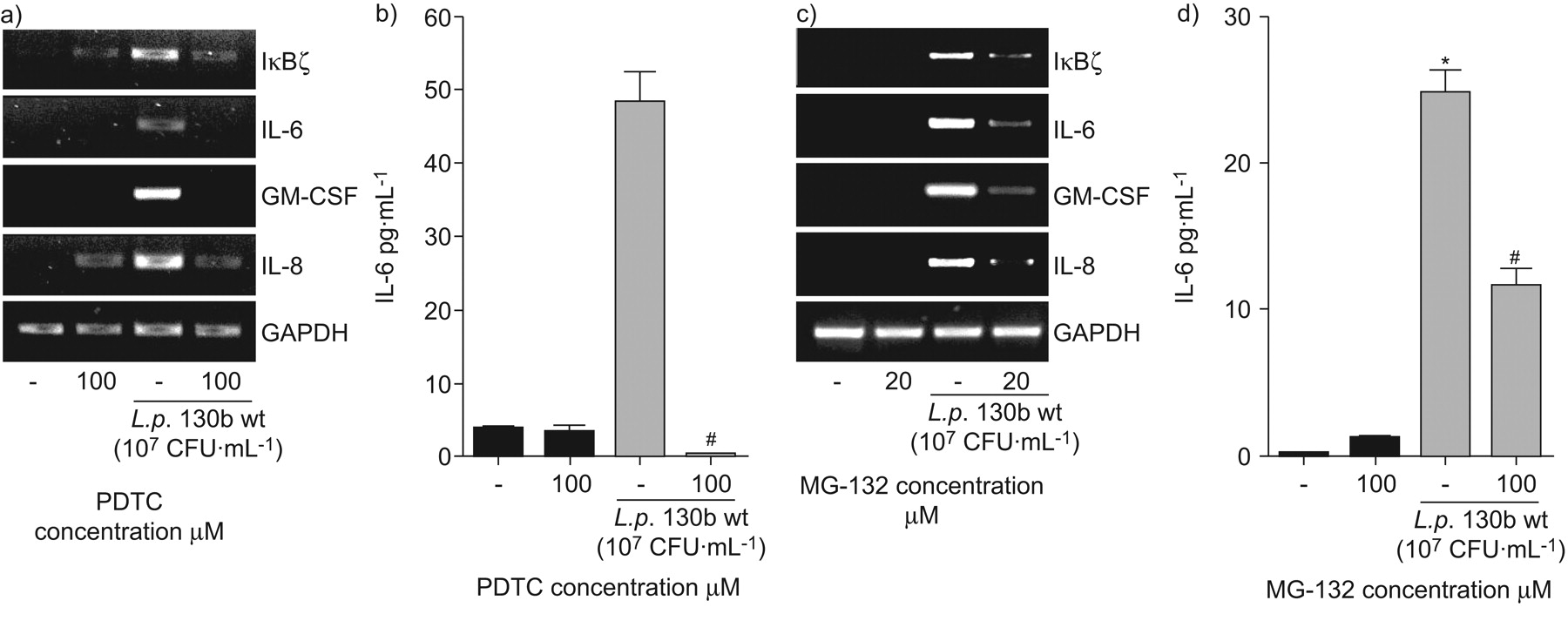
因为NF-κB似乎中央IκBζ和重要的促炎细胞因子的表达,包括il - 6,两个无关的NF-κB通路的抑制剂,PDTC和mg - 13239,40关于测试军团菌相关的细胞活化。在mRNA水平,两种抑制剂减少IκBζ的表达,il - 6, gm - csf和引发(图5和c)。此外,pre-incubation细胞抑制剂显著降低退伍军人130 b-induced il - 6 (图5 b和d)和引发(数据未显示)蛋白表达,证明了ELISA表明NF-κB途径的核心作用军团菌有关的上皮细胞激活。
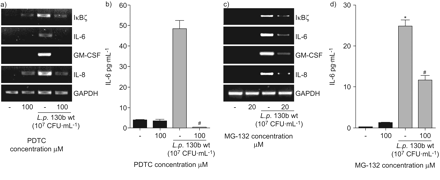
核因子(NF)的细胞因子和IκBζ-κB-related转录嗜肺性军团菌来华的上皮细胞。肺泡上皮细胞(A549) preincubated 30分钟的抑制剂,b) PDTC(100μM)或c, d) mg - 132(浓度),然后感染退伍军人野生型菌株130 b (107CFU·毫升−1)(▓)。7小时后,信使rna表达决心通过rt - pcr (a和c)。a, c)代表三个独立实验的凝胶。24小时后,白介素6 (IL)释放在上层的决定(b和d)。*: p < 0.05如果与刺激细胞;#细胞:p < 0.05与没有抑制剂。
的影响军团菌相关的信号的iκbζ和白细胞介素6基因启动子
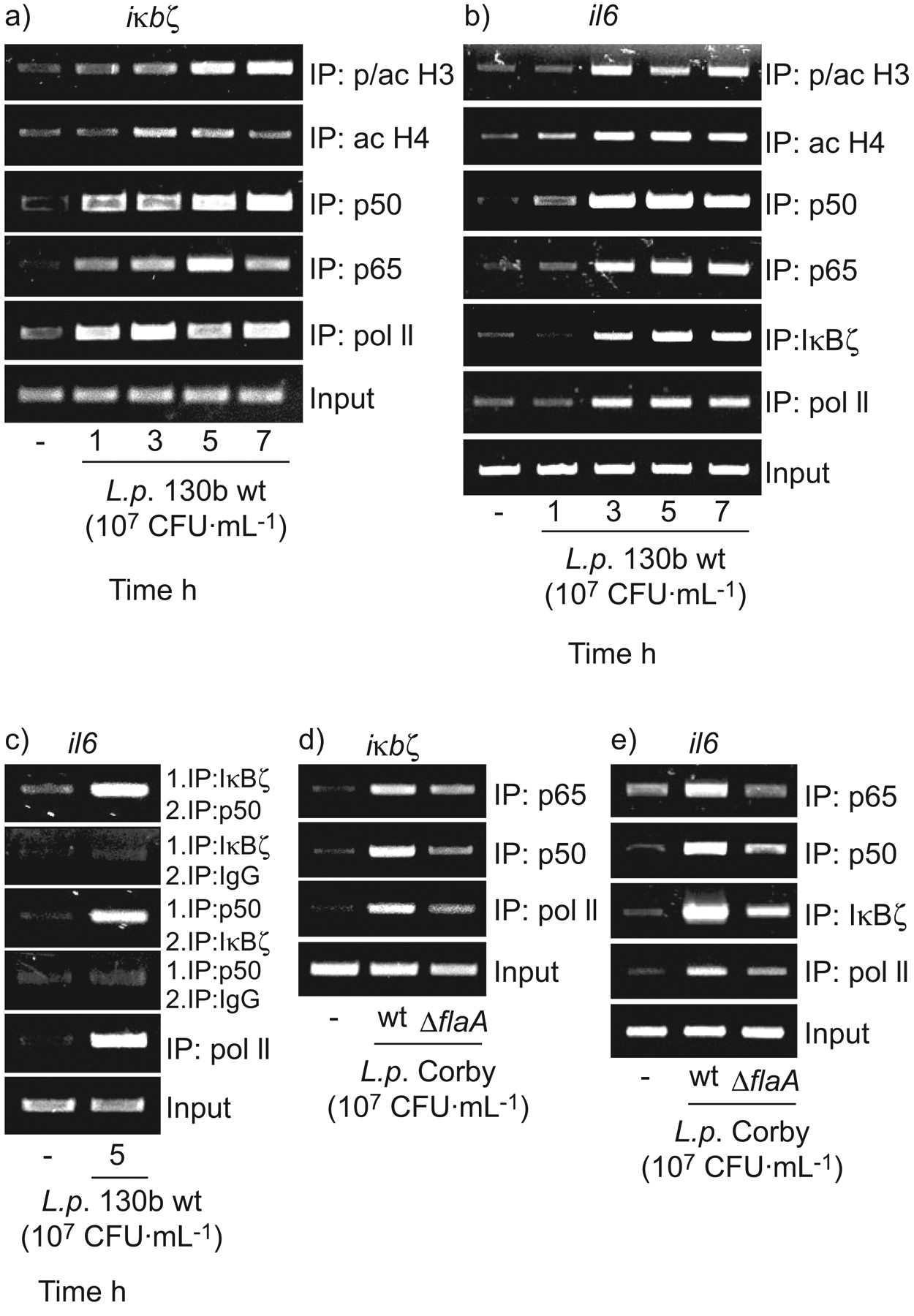
由于我们的数据显示IκBζ和NF-κB il - 6表达的一个重要的角色,我们利用芯片评估内生iκbζ和白细胞介素6基因启动子的更多细节退伍军人来华的细胞(图6)。在iκbζ基因催化剂,军团菌感染导致phospho-acetylation组蛋白H3 (PSer10/交流Lys14)和H4 pan-acetylation表明基因转录促进基因的改造(图6)。招聘NF-κB子单元p50和观察p65伴随着绑定的Pol II表明基因转录的开始(图6)。在白细胞介素6基因启动子,我们注意到相同的组蛋白修饰和招聘模式的p50 p65-NF-κB单元(图6 b)。此外,招聘IκBζ蛋白质以及波尔二世的白细胞介素6基因启动子记录退伍军人来华的上皮细胞。
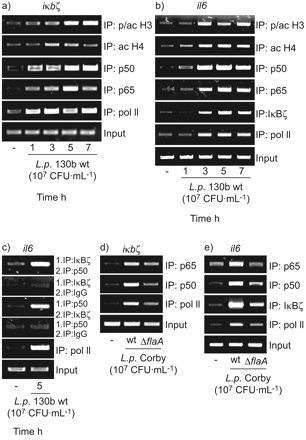
的影响军团菌相关的信号的iκbζ和白细胞介素6基因启动子。A549细胞感染107CFU·毫升−1嗜肺性军团菌菌株130 b (a - c)或寇比野生型(wt) /ΔflaA把缺失突变体(ΔflaA)(d和e)表示时间(a和b)或5 h(汉英)。染色质免疫沉淀反应(芯片)对指定的目标执行内生的倡导者白细胞介素6或(b, c和e)iκbζ凝胶(a和d)。代表三个独立的实验。
因为山崎et al。41证明强相互作用与p50 IκBζ,但p65弱绑定,我们测试了两个蛋白质之间的相互作用(IκBζ和p50)白细胞介素6基因启动子军团菌暴露的细胞。ChIP-IP,实验中我们发现内生p50协会和IκBζ蛋白质表明两分子的合作行动白细胞介素6基因启动子(图6 c)。
最后,我们想知道如果一个缺陷军团菌鞭毛蛋白表达会导致减少刺激转录因子基因启动子和波尔II招聘。因为它所示图6 d,flagellin-deficient军团菌诱导弱p65 p50和波尔II招聘iκbζ基因启动子。沿着这些线路,减少p65、p50 IκBζ绑定和波尔II招聘当时还演示了白细胞介素6基因启动子(图6 e)。量化的凝胶图6 d和e显示显著降低为波尔二世以及转录因子(数据没有显示)。
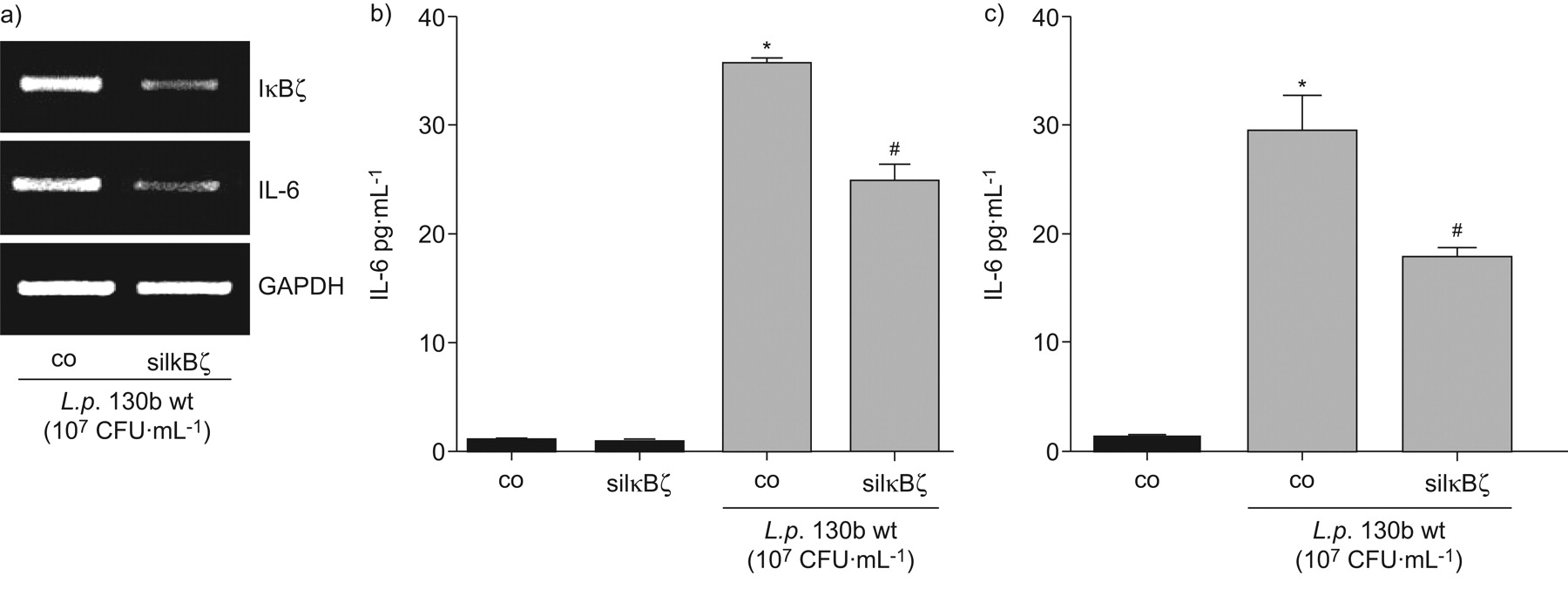
IκBζ降低il - 6表达的损耗退伍军人来华的上皮细胞
进一步分析IκBζ的作用军团菌有关的il - 6表达,我们利用IκBζ核。IκBζ-specific,但不是控制核,减少IκBζ以及il - 6 mRNA表达退伍军人感染A549细胞(图7)。此外,我们观察到显著降低il - 6蛋白的表达(图7 b)。对蛋白质含量相对较低的影响可能是由于有限的转染效率,从而由siRNA-unaffected细胞被克服。排除的影响不完全nucleofection,我们创建了A549细胞稳定表达非特定的或IκBζ-specific成分。细胞被感染退伍军人野生型菌株130 b (107CFU·毫升−1)24 h和il - 6发布由ELISA检测(图7 c)。在IκBζ-depleted细胞中,退伍军人全身的细胞il - 6版本相比,降低了41%表达不具体的成分。
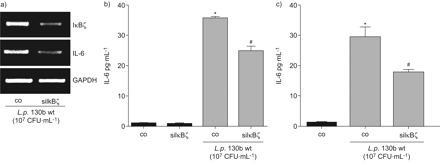
损耗降低IκBζ白介素6 (IL)表达式嗜肺性军团菌来华的上皮细胞。(a, b) A549细胞与未指明的nucleofected(有限公司)或IκBζ-specific (siIκBζ)核和后36 h感染退伍军人野生型菌株130 b (107CFU·毫升−1)24 h。IκBζ表达式是由一个rt - pcr和ELISA b)释放il - 6。c) A549稳定表达不具体的(公司)或IκBζ-specific成分(shIκBζ)被感染退伍军人130 b (10 wt压力7CFU·毫升−1)24 h和il - 6释放被ELISA检测。如果*:p < 0.05与刺激细胞;#:p < 0.05不具体的与IκBζsi /成分。凝胶),代表三个独立的实验。
MEK1和p38 MAP激酶抑制il - 6的表达而不是IκBζ通路阻塞
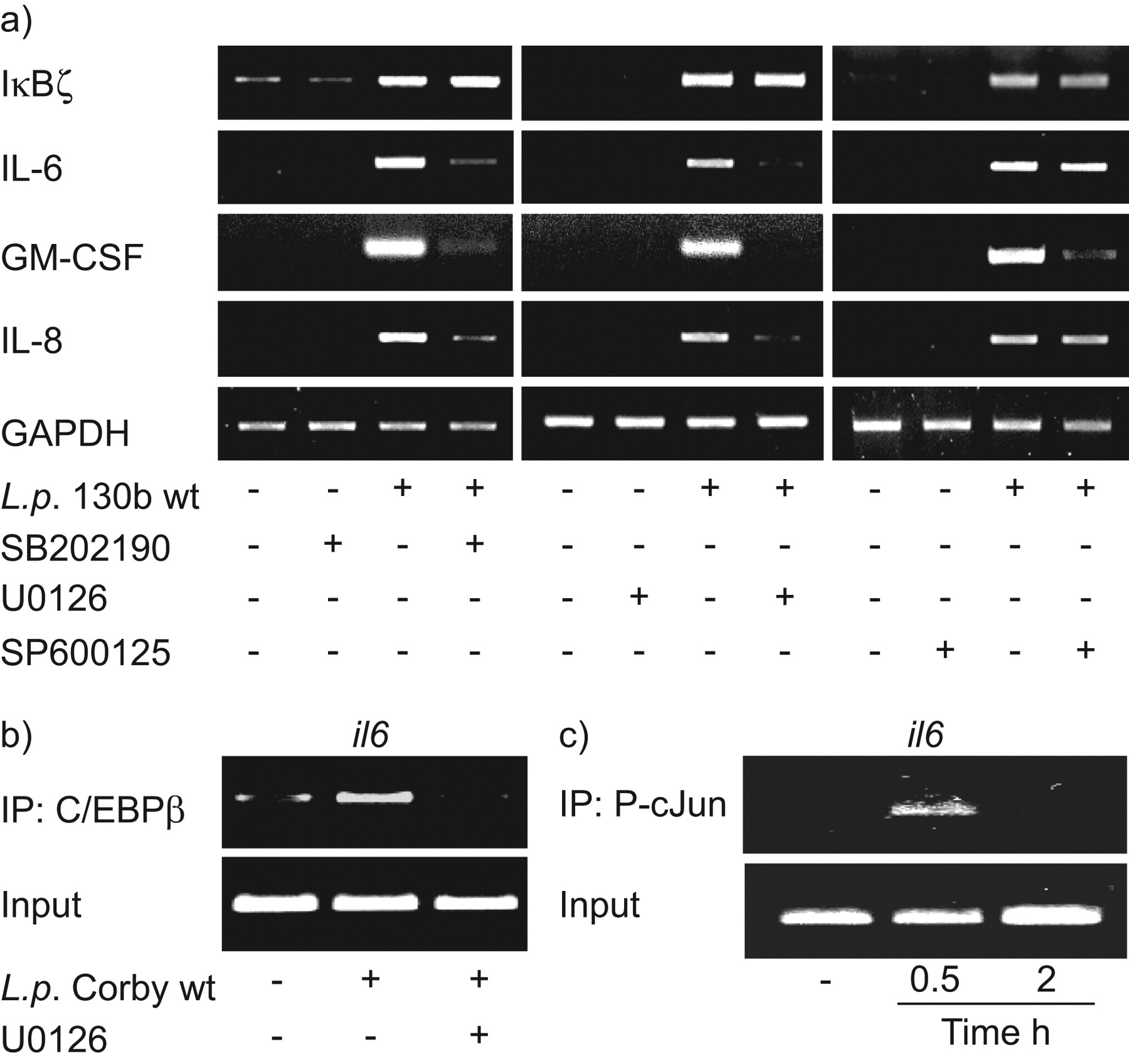
除了IκBζ和规范化NF-κB通路,几个激酶通路被怀疑是真核细胞的重要bacteria-related激活10,14,42。尤其是,他们的作用军团菌相关IκBζ和il - 6表达是未知的。因此,我们利用一些化学为重要的激酶抑制剂在IκBζ——测试他们的作用,和cytokine-expression军团菌受感染的细胞(图8和图S2补充材料)。抑制MEK1激酶(U0126 10μM)和p38 MAP激酶(SB202190 10μM)都封锁了退伍军人有关的il - 6、gm - csf和引发的表情。阻塞PI3激酶(Ly294002 10μM)和木菠萝亚型1 - 3(激酶抑制剂,10μM)阻止il - 6的表达和gm - csf,但不会引发的军团菌来华的细胞。然而,PKC抑制剂(Go6976 10μM)和物(SP600125 10μM)只有降低gm - csf表达。值得注意的是,无论是抑制剂影响退伍军人全身IκBζ表达式在肺上皮细胞虽然指出了有效的激酶抑制作用的抑制剂细胞因子来解决。

表达白介素6 (IL),但不是IκBζmRNA,取决于p38激酶和MEK1地图。一)肺泡上皮细胞(A549) preincubated与抑制剂SB202190 120分钟(10μM), U0126(10μM)或SP600125(10μM)然后感染退伍军人(l . p。(10)野生型菌株130 b7CFU·毫升−1)。7小时后,信使rna表达决心通过rt - pcr。代表三个独立实验的凝胶。A549细胞感染107CFU·毫升−1退伍军人寇比预培养后30分钟b)与抑制剂U0126 120分钟(10μM)或c)表示时报没有预孵化。对显示芯片进行目标的内源性启动子白细胞介素6。代表三个独立实验的凝胶。
转录因子C / EBPβ和AP-1涉及il - 6表达和下游目标MEK1和p38激酶地图,分别43。感染后退伍军人早期招聘C / EBPβ和的磷酸化形式AP-1亚基cJun发生在白细胞介素6启动子(图8 b和c)。抑制MEK1屏蔽退伍军人全身的C / EBPβ招聘的白细胞介素6启动子(图8 b)。
讨论
在这里,我们表明,退伍军人诱导flagellin-dependent IκBζ表达式和随后的il - 6表达在人类肺上皮细胞。军团菌诱导组蛋白修饰和招募p50 p65 / RelAiκbζ和随后的白细胞介素6基因启动子启动基因转录。同时,IκBζ和p50交互白细胞介素6基因催化剂军团菌来华的细胞和基因沉默的IκBζ减少军团菌有关的il - 6表达。
对肺的宿主-病原体相互作用的分子机制。这限制了创新的干预策略的发展尽管抗生素耐药细菌的出现使抗生素治疗肺炎。因此,我们旨在阐明il - 6表达的机制军团菌感染。
表达式的多效性的细胞因子il - 6在肺炎被怀疑导致感染和炎症的局部控制20.- - - - - -22例如通过调节中性粒细胞招募和功能23,24。间质性肺炎的发展转基因老鼠overexpressing人类白介素突出了il - 6的力量来促进肺部炎症过程44。感染的肺上皮细胞退伍军人菌株130 b, JR32和寇比(这项研究中,15,45)以及小鼠巨噬细胞46导致强烈的释放il - 6。
检测军团菌鞭毛蛋白在宿主细胞模式识别受体(PRR)似乎是一个关键的步骤,军团病。在早期感染时间点,鞭毛蛋白被transmembraneous TLR547和共同主导TLR5终止密码子多态性与人类对军团病的易感性5。鞭毛蛋白诱导重组il - 6表达,而flagellin-deficient退伍军人显示降低il - 6感应和TLR5-depletion真核细胞减少军团菌有关的il - 6表达指示flagellin-TLR5-related诱导il - 6的分泌。此外,胞质识别Naip5鞭毛蛋白单体的巨噬细胞导致的限制退伍军人感染小鼠38。值得注意,信使rna IκBζ和il - 6水平的差异,由野生型或flaA-deletion突变体消失后7 h。il - 6蛋白的差异释放持续24小时,额外的机制,如。在翻译或释放il - 6可能参与其中。此外,TLR2的作用可以从这些数据不能排除48。
鞭毛蛋白被怀疑到宿主细胞细胞质的iv型分泌system-induced穿孔时间38,激活,例如NAIP5或要诀37。在协议中,我们观察到在之后的时间点降低il - 6 mRNA表达在细胞感染军团菌应变JR32缺乏队伍,这些细菌的type-IVB系统不可分割的一部分8。总的来说,细胞外,可能是细胞内的认可退伍军人似乎是重要的肺上皮细胞il - 6表达。有趣的是,军团菌longbeachae(西澳大利亚的军团病的常见原因)包含鞭毛蛋白但没有检测到造孔活动,不激活Naip5-related活动,因此暗示可能重要的毒株特异性效应军团菌相关细胞的激活49。
il - 6的表达是受一些转录因子,包括NF-κB AP-1 C / EBP25,26。NF-κB-related基因表达中观察到退伍军人来华的肺细胞7,11,15。此外,转录因子AP-1 / cJun和C / EBPβ招募到白细胞介素6启动子后感染退伍军人。然而,更知道的作用诱导核蛋白质IκBζ肺部感染,这对il - 6表达似乎是至关重要的27。我们观察IκBζ的感应退伍军人130 b、寇比,JR32-infected A549和初级人类肺上皮细胞。它的表达持续24小时。符合山本的研究et al。27实验用鞭毛蛋白重组,flagellin-deficient退伍军人表示flagellin-related IκBζ诱导的肺上皮细胞。宿主细胞信号通路分析证明NF-κB IκBζ表达式的依赖。p38,然而,尽管抑制MEK1木菠萝或PI3激酶抑制il - 6表达,我们观察到不影响IκBζ表达式。此外,退伍军人全身的C / EBPβ招聘白细胞介素6子被MEK1抑制剂U0126。这些数据表明,kinase-related, IκBζ-independent信号也导致了il - 6表达退伍军人来华的上皮细胞。此外,PKC和物途径导致il - 6和IκBζ表达式,但减少了军团菌全身的gm - csf表达。两个激酶扮演重要角色已被证明上皮细胞和巨噬细胞的激活,分别11,50。
在感染的细胞退伍军人,组蛋白H3 phosphoacetylated和H4乙酰化iκbζ基因启动子,表明染色质重塑与转录相关的活动14,33,34,51。这是伴随着招募p50和p65 NF-κB子单元,这是已知重要IκBζ表达式52。除了这些修改和招聘同时记录白细胞介素6基因启动子,我们观察到IκBζ绑定的白细胞介素6基因启动子退伍军人来华的上皮细胞。
最近,Kayamaet al。52证明LPS-induced组蛋白H3 trimethylation在启动子的一个子集依赖IκBζ老鼠巨噬细胞。与我们的结果,似乎可能IκBζ参与TLR -或者NLR-induced核小体重塑调节二次基因的表达。这也可能解释IκBζ-dependence的物种,和细胞与类型相关的学位27。
IκBζ似乎优先与NF-κB单元p50交互41。事实上,交互的p50 IκBζ被观察到白细胞介素6基因启动子军团菌Chromatin-IP-IP暴露的细胞。沉默IκBζ表达的特定核或成分导致下降,但不完全废除,il - 6表达退伍军人来华的细胞,这表明IκBζ中扮演一个重要的但不是不可或缺的作用,il - 6表达系统中进行测试。
最后,flagellin-deficient退伍军人弱诱导转录因子在招聘和波尔II绑定,iκbζ和白细胞介素6基因启动子,进一步强调在激活基因转录的鞭毛蛋白的作用。总的来说,这些数据显示一系列flagellin-dependent染色质重塑,p50, p65,和波尔II招聘、IκBζ表情,和随后的il - 6表达退伍军人来华的肺上皮细胞。此外,其他途径,如MEK-1-C / EBPβ和p38地图kinase-AP-1以及规范化NF-κB途径似乎参与il - 6表达。
从观察的复杂特性军团菌有关的il - 6表达出现几个问题:例如,胞质受体有助于IκBζ和il - 6表达?此外,除了NF-κB分子的贡献,导致IκBζ表达广泛未知的信号机制和不同的激酶途径可能在本研究被排除在外。除了退伍军人其他重要的病原体,不表达鞭毛(如。肺炎双球菌)引起肺炎和il - 6表达42。此外,虽然如。il - 6对肺炎双球菌传播的重要作用21,22是已知的,潜在的分子机制pneumococci-related il - 6(和潜在IκBζ)表达式是未知的。最后,在活的有机体内研究IκBζ缺陷小鼠在肺炎将有助于分析其作用。
总之,我们的数据突出显示复杂的il - 6感应途径在人类肺上皮,由早期flagellin-TLR5-dependent组蛋白修饰,IκBζ表达式,以及随后IκBζ-p50-related il - 6表达。
确认
这部分工作将包括在j . Lorenz和i Pollok的博士论文。f·塞弗特的技术援助,m . Laumer j . Hellwig f·施赖伯和d·斯托尔(所有Universitatmedizin柏林,柏林,德国)我将非常感谢。
脚注
可以从本文的补充材料www.www.qdcxjkg.com
支持声明
这项工作是Bundesministerium毛皮的赠款支持的陶冶和大幅减退和德意志Forschungsgemeinschaft b Schmeck (BMBF-FORSYS-Partner“Forsys肺”F-Kz 0315256, BMBF网络的进步,DFG-SFB / TR84), j . Zahlten (BMBF网络的进步,DFG-SFB / TR84),美国Hippenstiel (BMBF网络的进步,DFG HI-789/6-1, DFG-SFB / TR84), n Suttorp (BMBF网络的进步,DFG-SFB / TR84),和柏林夏洛——夏洛蒂Schmeck。
感兴趣的语句
没有宣布。
- 收到了2009年12月19日。
- 接受2010年6月26日。
- ©2011人队











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}