





摘要GydF4y2Ba
哮喘患者慢性肥大细胞活化的机制尚不清楚。在没有过敏原的情况下,单体免疫球蛋白(Ig)E诱导啮齿动物肥大细胞释放介质,表明IgE可能在哮喘支气管粘膜肥大细胞的持续激活中发挥作用。本研究探讨了单体IgE是否诱导CaGydF4y2Ba2+GydF4y2Ba人肺肥大细胞(HLMC)的内流和介质释放。GydF4y2Ba
将纯化的HLMC培养4周,然后暴露于单体人骨髓瘤IgE。比率约定GydF4y2Ba2+GydF4y2Ba对单呋毛2负载细胞进行成像。组胺释放通过射精测定测量;白三烯C.GydF4y2Ba4.GydF4y2Ba(LTC.GydF4y2Ba4.GydF4y2Ba)通过ELISA测量白细胞介素(IL)-8。GydF4y2Ba
在经历的浓度下GydF4y2Ba体内GydF4y2Ba,单体IgE诱导剂量依赖性组胺释放,LTCGydF4y2Ba4.GydF4y2Ba生产和IL-8合成。这与细胞源性免费CA的增加有关GydF4y2Ba2+GydF4y2Ba.在初始接触IgE后,增强的组胺释放仍然是明显的1周,表明继续暴露保持增强的分泌。GydF4y2Ba
单独的单体免疫球蛋白E单独激活培养的人肺肥大细胞启动CA.GydF4y2Ba2+GydF4y2Ba内流、脱颗粒、花生四烯酸代谢和细胞因子合成。这些发现支持哮喘气道肥大细胞免疫球蛋白E负荷与哮喘气道生理紊乱有关的假设。GydF4y2Ba
肥大细胞在哮喘免疫病理学中发挥着核心作用,通过持续分泌血清炎介质,细胞因子和蛋白酶(审查)GydF4y2Ba1GydF4y2Ba).它们渗入哮喘气道的关键结构,如气道平滑肌(ASM)。GydF4y2Ba2GydF4y2Ba和气道粘液腺GydF4y2Ba3.GydF4y2Ba,这些事件对于将调解人定位到其预定的行动地点可能至关重要。GydF4y2Ba
通常假设哮喘中的慢性肥大细胞活化通过呼吸气道暴露于吸入气泡剂的哮喘驱动,导致过敏原特异性免疫球蛋白(Ig)E与肥大细胞高亲和力IgE受体FcεRi的交联。然而,过敏原避免通常只对已建立的哮喘的状态影响,这似乎变得“自我延续”GydF4y2Ba4.GydF4y2Ba.因此,持续哮喘中的肥大细胞分泌过多可能是由替代刺激引起的,例如蛋白酶GydF4y2Ba5.GydF4y2Ba,腺苷GydF4y2Ba6.GydF4y2Ba,细胞因子,如干细胞因子(SCF)GydF4y2Ba7.GydF4y2Ba.令人感兴趣的是,在没有IgE通过抗原交联的情况下,单独的IgE单体可以诱导大鼠肥大细胞系RBL-2H3和小鼠骨髓源性肥大细胞(BMMC)释放介质GydF4y2Ba8.GydF4y2Ba那GydF4y2Ba9.GydF4y2Ba.此外,IgE延长啮齿动物肥大细胞存活,部分通过存活增强细胞因子的自分泌释放,例如白细胞介素(IL)-6GydF4y2Ba8.GydF4y2Ba.出于未知的原因,这些响应的程度取决于IgE的来源。因此,某些类型的IgE诱导组胺和细胞因子合成,而其他类型的IgE仅诱导细胞因子的释放GydF4y2Ba8.GydF4y2Ba.这些观察结果特别有趣,因为在人类中,血清总IgE浓度、支气管高反应性和哮喘之间存在可重复的相关性GydF4y2Ba10.GydF4y2Ba-GydF4y2Ba13.GydF4y2Ba.因此,假设在哮喘中具有吸引力,在哮喘中,气道内的高度桅杆细胞活化至少部分地从IgE与FcεRi的增加的增加来产生。GydF4y2Ba
单体IgE对人肺肥大细胞(HLMC)功能的影响是未知的。啮齿动物和人肥大细胞之间存在重要的功能差异,这些功能差异是由离子通道活动的显着差异,目前作者最近证明了GydF4y2Ba14.GydF4y2Ba.因此,最终,必须对人细胞和感兴趣的组织进行研究。在这项研究中,本作者已经迈出了测试其假设的第一步。他们证明IgE在没有过敏原的情况下,激活培养的HLMC导致CaGydF4y2Ba2+GydF4y2Ba流入和释放组胺,白三烯C.GydF4y2Ba4.GydF4y2Ba(GydF4y2BaLTC.GydF4y2Ba4.GydF4y2Ba)和IL-8。这对哮喘发病机制具有重要意义。GydF4y2Ba
方法GydF4y2Ba
HLMC纯化和培养GydF4y2Ba
所有受试者都给予书面知情同意,该研究得到了英国莱斯特郡研究伦理委员会的批准。利用免疫亲和磁选择,将肺癌切除后1小时内获得的宏观正常肺中的HLMC分散并纯化GydF4y2Ba15.GydF4y2Ba.最终HLMC纯度> 98%,细胞活力> 97%(通过排除台盼蓝监测)。在分离后,HLMC在Dulbecco的改性基本培养基(DMEM)/谷氨酸/ HEPE中培养,含有10%FBS,1%MEMORICIES氨基酸(来自Life Technologies,Paisley,UK),1%抗生素/抗催化溶液(Sigma,Poole,英国),100 ng·mlGydF4y2Ba-1GydF4y2Ba重组人(RH)SCF,50ng·mLGydF4y2Ba-1GydF4y2Barhil-6和10 ng·mlGydF4y2Ba-1GydF4y2Barh -10 (R&D, Abington, UK),在37°C的湿化培养箱中用5% CO冲洗GydF4y2Ba2GydF4y2Ba在实验前至少4周GydF4y2Ba16.GydF4y2Ba.每7天更换媒体的一半。GydF4y2Ba
加利福尼亚州GydF4y2Ba2+GydF4y2Ba成像GydF4y2Ba
无细胞溶胶CA的变化GydF4y2Ba2+GydF4y2Ba([CaGydF4y2Ba2+GydF4y2Ba]GydF4y2Ba一世GydF4y2Ba通过使用CA荧光测量荧光GydF4y2Ba2+GydF4y2Ba- 如前所述的敏感探针Fura-2GydF4y2Ba16.GydF4y2Ba.数据采集以一个双波长图像的速率发生,每6秒作为340/380nm比率。这是转换为[加利福尼亚州GydF4y2Ba2+GydF4y2Ba]GydF4y2Ba一世GydF4y2Ba使用市售的校准试剂盒(分子探针)。根据需要将IgE加入到盘子中。GydF4y2Ba
调节释放的HLMC激活GydF4y2Ba
肥大细胞(1×10GydF4y2Ba4.GydF4y2Ba)加热至37℃,共3个重复,分别为50µL DMEM和50µL人骨髓瘤IgE (Calbiochem-Novabiochem, Nottingham, UK)或人IgG (Sigma, Poole, Dorset),添加2×最终浓度。37℃孵育30 min或24 h后,250×离心GydF4y2BaGGydF4y2Ba4分钟后,倒上清液。对照细胞微丸在无菌去离子水中裂解以测定总组织胺含量。GydF4y2Ba
为了研究IgE制剂中是否存在IgE的聚集物,将新鲜组成的IgE以14000×离心GydF4y2BaGGydF4y2Ba20分钟,除去上清液和重悬PBS中的任何聚集体。然后将离心IgE,任何聚集的IgE和正常IgE的剂量响应。GydF4y2Ba
介质分析GydF4y2Ba
如前所述,通过射精测定法测量组胺GydF4y2Ba15.GydF4y2Ba.LTC.GydF4y2Ba4.GydF4y2Ba(Cayman Chemical Co.,Ann Arbor,MI,USA)和IL-8(BD Biosciences,San Diego,CA,USA)根据制造商的说明通过ELISA测量。GydF4y2Ba
数据展示和统计分析GydF4y2Ba
数据表示为平均值±GydF4y2BaSEM.GydF4y2Ba除非另有说明。组胺,LTCGydF4y2Ba4.GydF4y2Ba和IL-8在NG·10中表达GydF4y2Ba-6GydF4y2Ba细胞释放到上清液中。使用配对或未配对(两个尾型)T-TEST评估数据组之间的差异。P值为<0.05被认为是统计学上显着的。GydF4y2Ba
结果GydF4y2Ba
单体IgE诱导组胺,LTC的释放GydF4y2Ba4.GydF4y2BaIL-8来自HLMCGydF4y2Ba
由于SCF是人肥大细胞的基本生长因子,因此诱导细胞凋亡的戒断,并且即使在正常组织中也存在,但在哮喘的气道中升高GydF4y2Ba17.GydF4y2Ba,首先检查单体IgE在其正常的SCF补充生长培养基中孵育HLMC的效果。预形成的颗粒衍生的组胺的早期释放(30分钟)和花生酸代谢LTC的新合成产物GydF4y2Ba4.GydF4y2Ba,并研究了趋化因子IL-8的晚(24 h)释放。有趣的是,目前的作者发现,来自几个不同捐赠者的HLMC释放组胺和LTCGydF4y2Ba4.GydF4y2Ba在SCF存在下与IgE孵育30分钟时以剂量依赖性方式(图1GydF4y2Ba⇓GydF4y2Ba因此,对照组组胺释放量为258±26 ng·10GydF4y2Ba-6GydF4y2Ba583±99 ng·10GydF4y2Ba-6GydF4y2Ba3µg·mLGydF4y2Ba-1GydF4y2BaIgE,对IgE依赖的组胺释放量为325±88 ng·10GydF4y2Ba-6GydF4y2Ba细胞(总量的12.8%)(p = 0.008,n = 8;图1GydF4y2Ba⇓GydF4y2Ba一种)。LTC的释放有一个平行增加GydF4y2Ba4.GydF4y2Ba净单体IgE依赖释放4.3±1.2 ng·10GydF4y2Ba-6GydF4y2Ba3µg·mLGydF4y2Ba-1GydF4y2BaIgE(p = 0.038,n = 4;图1GydF4y2Ba⇓GydF4y2Bab).用3µg·mL活化细胞24 h后检测IL-8浓度GydF4y2Ba-1GydF4y2Ba与对照相比,IgE和显着升高(对照2.77±0.51 ng·10GydF4y2Ba-6GydF4y2Ba细胞,IgE 7.33±0.19 ng·10GydF4y2Ba-6GydF4y2BaCells, p = 0.003, n = 4;图1GydF4y2Ba⇓GydF4y2BaC)。为了看该效果是IgE特异性的,使用IgG运行并行测试,发现这些细胞中没有释放组胺(299±27 Ng·10GydF4y2Ba-6GydF4y2Ba对照细胞中的细胞,与273±42 ng·10相比GydF4y2Ba-6GydF4y2Ba3µg·mLGydF4y2Ba-1GydF4y2BaIgG,p = 0.37,n = 4)。GydF4y2Ba
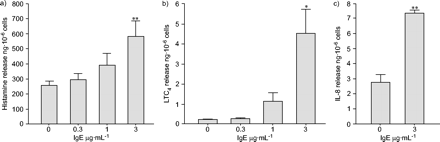
在干细胞因子100ng·mL存在下通过单体免疫球蛋白(Ig)e活化人肺肥豆细胞GydF4y2Ba-1GydF4y2Ba.单独的IgE诱导剂量依赖性释放a)预形成的颗粒衍生的介体组胺(n = 8),b)GydF4y2Ba德诺维GydF4y2Ba合成的花生酸代谢物白酮CGydF4y2Ba4.GydF4y2Ba(LTC.GydF4y2Ba4.GydF4y2Ba;n = 4)和c)新合成的白细胞介素(IL)-8 (n = 4)。所有值均为平均值±GydF4y2BaSEM.GydF4y2Ba.*:P <0.05;**:P <0.01。GydF4y2Ba
体内GydF4y2Ba,血清IgE浓度保持相当稳定,因此评估单体IgE的作用是否持续很重要。在另外三个供体中,分别用或不用3µg·mL培养HLMCGydF4y2Ba-1GydF4y2Ba在洗涤前在其正常的SCF补充生长培养基中持续7天,以除去累积的组胺并重新悬浮在中±IgE。组胺释放为125±27 ng·10GydF4y2Ba-6GydF4y2Ba没有IgE的细胞与1033±480 ng·10GydF4y2Ba-6GydF4y2Ba在3μg·ml存在下的细胞GydF4y2Ba-1GydF4y2BaIgE,净组胺释放909±288 ng·10GydF4y2Ba-6GydF4y2Ba细胞(24.6%的细胞总量)(n = 3,p = 0.05)。这表明当FcεRi被占用时,保持IgE信令。GydF4y2Ba
尽管缺乏生理学,但大多数研究都是在没有SCF的情况下研究人肥大细胞组胺释放。当在没有SCF的情况下,用单体IgE激活HLMC时,14个供体中有12个出现急性组胺释放,尽管这明显低于有SCF的实验。因此,取3µg·mLGydF4y2Ba-1GydF4y2Ba单体IgE,组胺释放从162±51 ng·10增加GydF4y2Ba-6GydF4y2Ba细胞浓度降至213±59 ng·10GydF4y2Ba-6GydF4y2Ba细胞,给出净组胺释放51±10 ng·10GydF4y2Ba-6GydF4y2Ba细胞(细胞总数的1.7%)(所有14个供体,p = 0.0002)。即使在scf缺失的细胞中,这种组胺释放也伴随着花生四烯酸代谢,以LTC的释放为例GydF4y2Ba4.GydF4y2Ba在IgE处理的细胞中,从175±28 pg·10开始GydF4y2Ba-6GydF4y2Ba对照细胞中的细胞到348±76 pg·10GydF4y2Ba-6GydF4y2Ba在3μg·ml中GydF4y2Ba-1GydF4y2BaIgE处理的细胞(p = 0.046,n = 10)(净释放为172±75 pg·10GydF4y2Ba-6GydF4y2Ba细胞)。GydF4y2Ba
单独的IgE激活肥大细胞通常会提出IgE是真正的单体的问题。可以想到,报告的效果是由在制剂中的IgE的低水平聚集体引起的,当添加到细胞交叉链路FcεRi中时,因此模拟过敏原暴露。为了解决这个问题,新建的IgE被离心为14000×GydF4y2BaGGydF4y2Ba20分钟,除去上清液和重生于PBS中的任何潜在聚集体。然后用任何聚集的IgE和正常的IgE和正常的IgE和正常的IgE和正常IgE之间的含量响应,并且在离心和正常的IgE之间没有显着差异(净组胺释放为51±7 ng·10GydF4y2Ba-6GydF4y2Ba细胞和57±8 ng·10GydF4y2Ba-6GydF4y2Ba细胞,分别具有3μg·mlGydF4y2Ba-1GydF4y2BaIgE)。此外,如果任何聚集体在沉淀物中,它们并未引发细胞中的任何反应(净组胺释放5±2ng·10GydF4y2Ba-6GydF4y2Ba细胞),这与Kalesnikoff的先前观察一致GydF4y2Ba等GydF4y2Ba.GydF4y2Ba18.GydF4y2Ba.GydF4y2Ba
单体IgE增加了无核糖的CAGydF4y2Ba2+GydF4y2Ba
在接下来的情况下是从HLMC中的单体IgE依赖性组胺释放是否与[CA的升高相关GydF4y2Ba2+GydF4y2Ba]GydF4y2Ba一世GydF4y2Ba.研究发现,在缺乏SCF的情况下,单体IgE诱导[CaGydF4y2Ba2+GydF4y2Ba]GydF4y2Ba一世GydF4y2Ba由添加IgE前的277.4±4.7 nM 3 min增至添加3µg·mL后的376.2±8.39 nM 2 minGydF4y2Ba-1GydF4y2BaIgE (p = 0.0001, n = 162个来自4个捐献者的细胞;图2GydF4y2Ba⇓GydF4y2Ba).目前的作者最近表明,在HLMC中[CaGydF4y2Ba2+GydF4y2Ba]GydF4y2Ba一世GydF4y2Ba和抗IgE依赖性组胺释放的幅度GydF4y2Ba19.GydF4y2Ba.从这种关系中,[CA的变化GydF4y2Ba2+GydF4y2Ba]GydF4y2Ba一世GydF4y2Ba在本研究中,预测净组胺释放的〜3%的细胞总量,其非常接近实际测量的细胞(2.0%不包括非反应者)。加利福尼亚州GydF4y2Ba2+GydF4y2Ba高原相对光滑,在抗IGE依赖性激活后保持HLMC中报告的变化,尽管偶尔细胞表现出与嗜碱性粒细胞中所述的相似的振荡GydF4y2Ba20.GydF4y2Ba.这表明,单体IgE和抗IgE激活的信号通路可能是相似的,这与最近从啮齿动物研究中得出的结论一致GydF4y2Ba21.GydF4y2Ba.GydF4y2Ba

{kind=link}
{kind=link}
![Heterogeneity of changes in [Ca2+]i in human lung mast cells during incubation with 3 µg·mL−1 monomeric immunoglobulin (Ig)E. Each panel represents a different donor and each trace an individual cell.](http://www.qdcxjkg.com/content/erj/25/5/858/F2.large.jpg?width=800&height=600&carousel=1){kind=link}
{kind=link}
[CA的变化的异质性GydF4y2Ba2+GydF4y2Ba]GydF4y2Ba一世GydF4y2Ba用3µg·mL孵育人肺肥大细胞GydF4y2Ba-1GydF4y2Ba单体的免疫球蛋白(Ig) E。每个面板代表一个不同的供体,每个追踪一个细胞。GydF4y2Ba
讨论GydF4y2Ba
该研究使新的观察结果培养HLMC释放组胺,LTCGydF4y2Ba4.GydF4y2Ba和IL-8关于急性暴露于单体IgE,而IgE仍然存在,则保持这种效果。与抗IgE依赖性激活相比,效果相对较小,特别是在没有SCF的情况下,甚至这种介质释放的幅度甚至可能具有较大的生理意义,其中肥大细胞渗透关键组织结构。GydF4y2Ba
许多GydF4y2Ba在体外GydF4y2Ba对肥大细胞的研究一直依赖于FcϵRI交联最大限度地激活细胞,导致过敏脱颗粒和大量组胺释放。然而,这一水平的最大细胞激活是值得怀疑的生理相关性,因为它可能从来没有实现在人类GydF4y2Ba体内GydF4y2Ba,也许在过敏反应和实验室过敏原刺激期间除外。当然,在慢性持续哮喘中,肥大细胞活化的侵袭性要小得多,在电子显微镜下可见碎片性脱颗粒,而不是最大细胞活化时出现的过敏性脱颗粒GydF4y2Ba22.GydF4y2Ba.与过敏原攻击后稳定的哮喘,在支气管肺泡灌洗液中也存在显着较低的介质浓度。GydF4y2Ba23.GydF4y2Ba.因此,当存在靶细胞的近似近似时,用单体IgE证明的介质的相对细微释放可能是重要的。这在哮喘中特别相关,其中在特定组织结构中存在肥大细胞的微生物,例如ASMGydF4y2Ba2GydF4y2Ba.观察结果,HLMC不仅脱粒,而且还响应IgE响应IgE而合成的胰岛素和合成细胞因子表明,随后会出现几种不同的组织效应。例如,组胺是支气管电池,胰蛋白酶是ASM丝裂原GydF4y2Ba24.GydF4y2Ba,Cysteinyl白三烯是有效的支气管扩张剂和暴露于IL-13后的增强ASM增殖GydF4y2Ba25.GydF4y2Ba,而IL-8也是一种有效的平滑肌丝分裂原GydF4y2Ba26.GydF4y2Ba.此外,由于SCF的浓度在哮喘呼吸道中升高GydF4y2Ba17.GydF4y2Ba,而SCF是ASM的产物GydF4y2Ba27.GydF4y2Ba,很容易设想正反馈环,其中单体Ige与ASM衍生的SCF一起激活肥大细胞,其又激活ASM。哮喘中的这种肥大细胞活化的途径将有助于解释总体而不是特异性血清IgE和支气管高反应性程度之间的强烈相关性GydF4y2Ba10.GydF4y2Ba-GydF4y2Ba13.GydF4y2Ba.GydF4y2Ba
随着时间的推移,IgE的血清浓度仍然相对恒定,因此当前作者已经表明,暴露于IgE后1周,保持增强的HLMC分泌反应。这与来自RBL-2H3细胞的数据一致,显示电池信令只要存在未结合的IgE即可。删除“自由”IgE快速将细胞返回到静态。迅速推荐信号传导的IgE对敏化单元的重新引入,建议绑定的IgE不会阻止“自由”IgE信号传导GydF4y2Ba21.GydF4y2Ba.因此,这些观察结果为从哮喘的肥大细胞中提高了增强的本组成介质介质释放的进一步机制GydF4y2Ba与GydF4y2Ba正常的课程GydF4y2Ba28.GydF4y2Ba.还要注意,IgE的顶部浓度研究,3μg·mlGydF4y2Ba-1GydF4y2Ba,相当于~ 1250 iU·mLGydF4y2Ba-1GydF4y2Ba良好的范围内GydF4y2Ba体内GydF4y2Ba.GydF4y2Ba
在本研究中,已经使用过至少4周的培养物中的肺部肥大细胞,如目前的作者所述GydF4y2Ba16.GydF4y2Ba.这是有几个原因。首先,与来自哮喘受试者的那些,来自正常供体的新鲜分离的肥大细胞相对静态,并且大部分细胞不表达FcεRiGydF4y2Ba28.GydF4y2Ba那GydF4y2Ba29.GydF4y2Ba.那些表达FcϵRI的细胞很可能被内源性IgE占据,这需要在低pH下剥离,从而导致细胞损伤。相比之下,培养的细胞都表达FcϵRI,当IgE致敏时,比新鲜分离的细胞释放更多的抗IgE介质GydF4y2Ba16.GydF4y2Ba那GydF4y2Ba19.GydF4y2Ba.更重要的是,大约一半的细胞在培养的第一周内死亡,但随后将增殖,因此在4周内,起始数量有多达五倍GydF4y2Ba16.GydF4y2Ba.因此,≥90%的细胞在4周将暴露于IgE,使实验可行。此外,由于与正常气道相比,SCF的表达明显增加了SCF的表达GydF4y2Ba17.GydF4y2Ba,已经在SCF中培养的这些细胞也可以比来自正常受试者的细胞更代表哮喘表型。GydF4y2Ba
当检查IgE对HLMC激活的影响时,重要的是确保所用的IgE是单体的。因此离心制剂,并平行地将离心IgE,未凝胶的IgE和任何聚集的IgE的剂量反应进行离心,并且发现离心和未经干燥的制剂之间没有显着差异。这与Kalesnikoff的观察一致GydF4y2Ba等GydF4y2Ba.GydF4y2Ba18.GydF4y2Ba尽管他们实验中使用的IgE (SPE-7抗二硝基酚)含有聚集体,但聚集体本身在引发反应方面的效果远不如单体IgEGydF4y2Ba18.GydF4y2Ba.GydF4y2Ba
来自小鼠和RBL-2H3细胞的先前工作表明,单体IgE导致肥大细胞活化的机制是通过增强FcεRI的二聚化GydF4y2Ba18.GydF4y2Ba那GydF4y2Ba21.GydF4y2Ba.这表明激活的信令通路应该类似于由抗IgE激活的。在加利福尼亚州GydF4y2Ba2+GydF4y2Ba据报道,动员,在以前的鼠研究中,据据报道,IgE引起CAGydF4y2Ba2+GydF4y2Ba动员导致[加利福尼亚州GydF4y2Ba2+GydF4y2Ba]GydF4y2Ba我,GydF4y2Ba但是,这未能达到要发生黎明所需的阈值。Huber研究GydF4y2Ba等GydF4y2Ba.GydF4y2Ba30.GydF4y2Ba和KalesnikoffGydF4y2Ba等GydF4y2Ba.GydF4y2Ba18.GydF4y2Ba发现野生型BMMC对IgE单独的去颗粒反应并不显著,尽管[CaGydF4y2Ba2+GydF4y2Ba]GydF4y2Ba一世GydF4y2Ba并且证据表明FcεRi的β和γ亚基以及许多ERK激酶和含SH2的肌醇5'-磷酸酶,是酪氨酸磷酸化GydF4y2Ba18.GydF4y2Ba.奥卡的另一个研究GydF4y2Ba等GydF4y2Ba.GydF4y2Ba9.GydF4y2Ba报道,向RBL-2H3细胞添加单体IgE诱导[CA的显着增加GydF4y2Ba2+GydF4y2Ba]GydF4y2Ba一世GydF4y2Ba和调解员释放。然而,在同一研究中,鼠BMMC表现出比RBL-2H3细胞显着较少(但仍然显着的)脱粒,尽管如此GydF4y2Ba2+GydF4y2Ba]GydF4y2Ba一世GydF4y2Ba增加是可比较的GydF4y2Ba9.GydF4y2Ba.已经表明了CAGydF4y2Ba2+GydF4y2Ba在抗ige依赖的细胞激活后,HLMC中观察到的信号与当前作者和其他人之前描述的类似GydF4y2Ba19.GydF4y2Ba那GydF4y2Ba20.GydF4y2Ba.这表明,抗IgE和单体IgE激活的信号通路具有多种成分,但是否相同还需要进一步研究。GydF4y2Ba
有趣的是,与目前作者的结果相反的是,之前唯一的单体IgE对人肥大细胞激活的研究使用了脐血来源的肥大细胞,并没有报道对组胺或IL-8释放有显著影响,尽管这是在没有SCF的情况下GydF4y2Ba31.GydF4y2Ba.众所周知,物种和来自同一物种内不同组织之间的肥大细胞反应中有明显的异质性,因此数据再次突出了从感兴趣的组织学习细胞的重要性。GydF4y2Ba
总之,结果表明单体免疫球蛋白E单独诱导组胺,白酮胺C的释放GydF4y2Ba4.GydF4y2Ba来自人肺肥大细胞的白细胞介素-8。释放的幅度小于抗免疫球蛋白E或过敏原可以获得的大规模释放,但这实际上可能更为生理相关。由于肥大细胞渗透到哮喘中的呼吸道平滑肌,例如气道平滑肌,响应于单体免疫球蛋白E分泌的各种介质阵列可能对这些细胞的功能具有关键后果。这些发现将有助于解释总血清免疫球蛋白E和哮喘和支气管高反应性之间观察到的强烈相关性,以及抗免疫球蛋白E治疗与过敏原的避免相比,抗免疫球蛋白E治疗的能力GydF4y2Ba32.GydF4y2Ba.因此,单体免疫球蛋白E激活人肺肥大细胞的能力可以是患有哮喘的慢性症状的核心。GydF4y2Ba
致谢GydF4y2Ba
作者要感谢S. Harper(AstraZeneca R&D Charnwood,英国Loughborough,Loughborough,Loughborough,Loughne组胺甲基转移酶的慷慨礼物,D. Parker和L. Woodman为他们的技术援助。GydF4y2Ba
- 已收到GydF4y2Ba2004年8月3日。GydF4y2Ba
- 公认GydF4y2Ba2005年1月20日。GydF4y2Ba
- ©ERS期刊有限公司GydF4y2Ba
参考GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba
- ↵GydF4y2Ba









