





摘要
假设膈肌损伤激活了肌肉修复中基于时间的基因表达程序。
不同物质的基因表达,如蛋白酶(calpain 94 (p94)),转录因子(myogenin和cFos),生长因子(碱性成纤维细胞生长因子(bFGF)和胰岛素样生长因子(IGF)-II),结构蛋白(肌凝蛋白重链(MHC)和肌联蛋白),通过RT-PCR对暴露于咖啡因损伤的大鼠横膈膜进行定量。分别于不同时间点(1 ~ 240 h)切除损伤和非损伤(对照)大鼠半膈肌。
在受伤的半膈肌中,与对照肌肉相比,p94的表达水平在1 损伤后h(PI),cFos mRNA水平在最初下降后开始升高,并在96小时达到峰值 而肌生成素mRNA水平早在12岁时就开始升高 h-PI、IGF-II mRNA水平最初下降至48 h-PI随后增加,在72时达到峰值 h-PI、bfgfmrna水平在96℃时达到最大值 h-PI、MHC和titin的mRNA水平在72岁时显著升高 h PI。
咖啡因诱导的膈肌损伤之后是一个基于时间的不同基因的表达程序,以满足肌肉修复的需要。
横膈膜纤维损伤与肌肉力量减少有关1.,2.这可能会导致呼吸肌肉功能障碍。膜片损伤均可观察到体外3.和在活的有机体内吸气后重载4.–6.脓毒症,7.,延长机械通气时间8.,9. 在人类中,慢性阻塞性肺疾病(COPD)患者的膈肌肌节断裂比健康人更为普遍10. 肌肉损伤的程度与气道阻塞的程度有关,COPD患者的高阈值吸气负荷增加了损伤的程度10,11. 基于这些发现,可以假设横膈膜纤维在受伤后能够有效再生,就像四肢肌肉在一生中反复修复时一样12,13.骨骼肌再生需要激活静止的肌原性前体细胞(mpc), mpc经历多次细胞分裂,分化成成肌细胞,融合到受损的纤维上,最后成熟成肌纤维。尽管不同分子机制的作用尚未明确,但一些研究已经提示了基于时间的基因表达程序在肢体肌肉损伤后调控mpc进展13–15.
在一个在活的有机体内大鼠模型2.,之前已有研究表明咖啡因诱导的膈肌损伤导致33%的肌纤维肌膜破裂,与1小时时70%的肌力减少相关 h损伤后(PI)。4个月时膜损伤完全愈合 天圆周率。然而,完全恢复力只发生在10点 days PI,表明肌节和其他细胞结构的组装,需要正常的力量生产,需要比膜愈合更长的时间。这支持了膈肌修复过程中基于时间的计划概念,包括不同分子因子的有组织表达,如蛋白酶、肌源性调节因子、生长因子和结构蛋白。
目前作者假设隔膜损伤激活了肌肉修复中的基于时间基因表达的程序。因此,主要目标是研究肌肉再生过程中涉及不同阶段的基因的mRNA表达的时间过程,即蛋白酶(Calpain 94(P94)),转录因子(CFOS和Myogenin),生长因子(碱性成纤维细胞生长因子(BFGF)和胰岛素样生长因子(IGF)-II)和结构蛋白(肌球蛋白重链(MHC)和三肽)。
材料和方法
方法
所有实验均在体重250-300克的Sprague-Dawley大鼠(Charles River, St. Constant, QC, Canada)上进行,按照加拿大动物保护委员会(Ottawa, on, Canada)的指导方针,经动物保护委员会(Centre hospitalier del 'Université de Montréal (CHUM), Hôpital Notre-Dame,加拿大蒙特利尔,QC)。目前的作者研究了咖啡因诱导的损伤模型,该模型已经显示33%的横膈膜纤维存在肌细胞膜损伤2..
实验步骤
腹腔注射戊巴比妥钠(50 mg·kg)麻醉大鼠−1.).在腹壁中间进行无菌切口后,将右侧膈肌肋部暴露于100 mm的咖啡因溶液(Sigma Chemicals, St. Louis, MO, USA)中,该溶液溶解在hepes缓冲的Krebs溶液中。咖啡因包含在一个直径2厘米的塑料吸盘中,用负压(-20厘米h)保持10分钟2.O)。用充满盐水的吸盘对所有大鼠作为对照肌肉的左膈肌进行同样的操作。然后关闭腹壁,让动物在有食物和水的笼子里恢复自由.
为了检查局部使用咖啡因是否会导致缺氧,我们进行了一系列补充实验。在33只成年雄性大鼠中,测量了整个横膈膜注射咖啡因期间(10分钟)血红蛋白的经皮氧饱和度。这33种动物都没有饱和。
研究方案和隔膜样品制备
与之前一样,建立9个时间点组(n = 7)2.,分别于1、4、6、12、24、48、72、96和240 h处死,每只作为单独对照,因为左侧膈肌未处理。隔膜被移除整体,并浸入平衡的正规克雷布斯溶液(95% O2., 5%的公司2.,博士 7.38)在4°C下冷却以进行进一步解剖。咖啡因和生理盐水暴露区域均被定位并分为两部分。解剖一个矩形块,并在-80°C下冷冻,用于随后的分子生物学分析。第二块在异戊烷中快速冷冻,异戊烷已用液氮预冷,并在-80°C下保存以进行组织学检查。
组织学分析
肌条(10 μm横截面),在低温切片机(Leica crycut 1800;用苏木素和伊红(H&E)染色,用尼康Eclipse TE600显微镜(尼康,梅尔维尔,纽约,美国)观察,该显微镜连接到Photometrics®CoolSNAPTM值相机(Roper科学公司,图森,AZ,美国)。标准改编自里德和贝尔卡斯特罗16用于识别异常肌肉形态。当肌肉区域包含形态异常的活肌肉、坏死肌肉或被炎性细胞侵入(没有明显的肌肉细胞轮廓)时,视为异常.正常和异常区域使用Metarm®成像系统4.6软件(美国宾夕法尼亚州唐宁敦环球成像公司)的面积计算器工具进行测量首先,计算总横截面积,包括正常和异常肌肉,并以校准单位显示。然后,使用手动轮廓工具分隔所有异常区域,并以类似方式计算。异常组织的部分面积由异常肌肉相对于总cr的百分比表示oss截面积17.
总RNA提取
冷冻膈肌组织均质化(9855370型组织撕裂器;Biospec Products, Bartlesville, OK, USA)在1 mL的TRIzol®试剂(Life Technologies Inc., Rockville, MD, USA)根据制造商的说明。提取的总RNA经乙醇沉淀、干燥、重悬于RNAse-free水中,-80℃保存。
RT-PCR分析
表1列出了用于每个研究因子的寡核苷酸引物对⇓.纯化总RNA(1 μg/反应)是Titan®单管RT-PCR系统(加拿大拉瓦尔罗氏诊断公司)中基因片段RT-PCR扩增的底物。每个基因特异性片段与对应于mRNA甘油醛-3-磷酸脱氢酶(GAPDH)的237碱基对片段共扩增.GAPDH作为参考基因,因为其表达被广泛认可用于标准化不同组织中不同基因的表达水平,以及在许多生理条件下,包括肌肉损伤和修复18,19.在循环前,逆转录在50℃下进行30分钟。根据所研究的基因进行28-39个周期的线性扩增(表1)⇓).每个循环在94℃下由1分钟组成,然后在55℃下在68℃下进行1分钟,3分钟。通过将GAPDH引物加入反应混合物中通过剩余28个循环来实现共同扩增。然后在55℃下循环停止5分钟以允许GAPDH特异性序列的逆转录,并且剩余的28个扩增循环被恢复。最后一个循环的伸长率为10分钟而不是3分钟。在当前作者的经验中,该试剂盒中的逆转录酶非常温度稳定。所有RT-PCR产物在1.5%标准琼脂糖/ 1×三硼酸乙酸乙酸乙酰乙酸乙酸乙酰乙酸乙酰乙酸盐(TBE)凝胶中分离,并用溴化乙锭染色,除了MHC / GAPDH共同扩增,它们以3%高分离- 溶解(3:1)琼脂糖/ 1×Tbe凝胶。
光密度分析
扩增产物在Alpha Imager 2000文件和分析系统(美国加利福尼亚州圣莱安德罗Alpha Inotech公司)中进行分析。相对于GAPDH mRNA,损伤纤维中不同分子标记物的mRNA表达正常化。因此,代表每个样本中目标基因的条带强度除以样本对应GAPDH条带的强度。然后将处理后的横膈膜计算的比率与未损伤对照组织的比率进行比较,并根据相对表达绘制时间曲线。
统计分析
值表示为相对表达式的平均值±扫描电镜. 首先通过Wilcoxon检验对相关样本比较受损半膈肌和对侧半膈肌在不同时间点各因子的相对mRNA表达和异常肌肉部分面积。在确认正态性和方差相等性后,通过单因素方差分析比较各因素在不同时间点的表达。邦费罗尼酒店事后通过检验确定不同时间点之间的显著差异。此外,使用Kruskal-Wallis单因素方差分析对排名进行分析。显著性水平p<0.05。
后果
异常肌肉区域的组织学定量
通过H&E染色监测损伤和对照半膈肌的坏死和炎症进展(图1)⇓).对照肌肉未显示任何单核细胞/巨噬细胞浸润(图。 1a⇓).中度炎症细胞浸润早在PI 6 h就出现,但在24 h时更为明显(图1b)⇓).通过单核细胞肿胀的形态和肿胀的坏死细胞浸润的肌坏死达到48的最大值(图1C⇓), 72 h PI。这些异常随后在96 h下降,在240 h PI完全消失(图1d)⇓).统计比较如图2所示⇓.
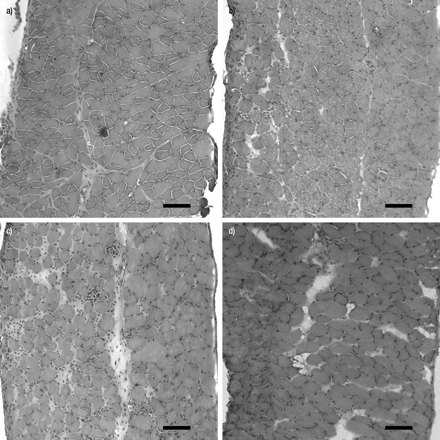
控制非生成隔膜(A)。单核细胞的间质空间和肌细胞侵袭在损伤后24小时(pi; b)中明显明显,并且在48小时(c)中显着上升。单核细胞浸润完全消失在240小时(D)。秤条=50μm。
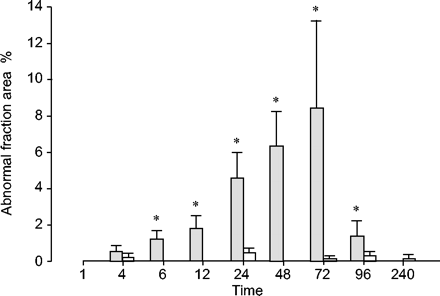
咖啡因治疗后苏木精和伊红组织异常的演变( )和未治疗的对照半膈肌(□)。结果用百分比±表示扫描电镜每个受伤后时间点的总面积。未观察到渗透时,未表示钢筋。*:p<0.05,与对照组有显著性差异。
)和未治疗的对照半膈肌(□)。结果用百分比±表示扫描电镜每个受伤后时间点的总面积。未观察到渗透时,未表示钢筋。*:p<0.05,与对照组有显著性差异。
标记的时间-过程表达
蛋白酵素
P94 mRNA在隔膜损伤后1小时达到峰值,两倍于基线值(P <0.05),并在4小时下降至基线。在240小时PI观察到晚期显着增加(图3A⇓).
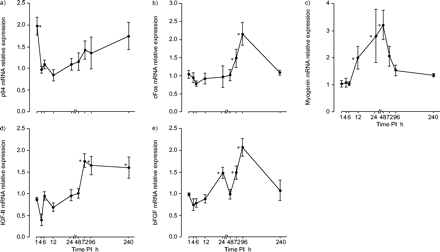
损伤后(PI)CALPAIN 94(p94; a)的表达模式,肌原素转录因子CFOS(b)和肌菌素(c),以及生长因子胰岛素样生长因子(IgF)-ii(d)和碱性成纤维细胞生长因子(BFGF; e)。相对表达±扫描电镜每个时间点显示与对照组比较的结果(对照组横膈膜中mRNA表达的均值设为1,PI mRNA表达值归一化至对照组)。*: p<0.05(治疗组半膈肌mRNA表达与对照组差异显著)。
转录因子
与对照组相比,cFos mRNA表达在6 h PI时降低(p<0.05),并在12 h PI时恢复到基线水平(图3b)⇑).此后,在PI 48 h时开始逐渐升高,在PI 96 h时急剧升高(p<0.05)。有趣的是,cFos mRNA的开始升高与肌原蛋白mRNA的下降直接相关(72 h PI)。肌原蛋白mRNA表达在12 h时开始升高(p<0.05),在48 h时达到峰值,是对照组的3倍(p<0.05)。在72 h PI之前,它的表达一直保持高水平(图3c)⇑).
生长因子
IGF-II mRNA表达下降1 h PI,并在6时达到最低水平 h(p<0.05),持续低至48小时 h PI。其最高水平为72 术后恢复h,几乎是对侧膈肌恢复h的两倍(p<0.05),在240分钟前显著升高 h PI(图。 三维⇑).bFGF mRNA表达在PI 6 h时开始下降(p<0.05),在PI 24 h时显著升高(p<0.05)。第二个表达高峰是对照膈肌的2倍,在96 h PI时出现(p<0.05),在240 h PI时达到基线(图3e)⇑).
结构蛋白
MHC mRNA表达在1 h PI时出现最初的抑制(p<0.05),在72 h PI时显著升高(图4a)⇓).在96 h PI时表达量达到最大值,此后仍保持较高水平(p<0.05)。最初的下降并不显著,但72 h PI后,titin mRNA表达显著上升,此后保持升高(p<0.05;图4 b⇓).
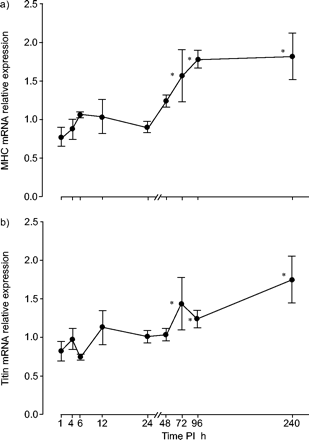
肌肉结构蛋白肌球蛋白重链(MHC;a)和titin(b)的损伤后(PI)mRNA表达模式。相对表达式±扫描电镜每个时间点显示与对照组比较的结果(对照组横膈膜中mRNA表达的均值设为1,PI mRNA表达值归一化至对照组)。*: p<0.05(治疗组半膈肌mRNA表达与对照组差异显著)。
讨论
本研究的主要发现如下:1)p94表达水平在1 h PI达到峰值;2)转录因子cFos mRNA水平在最初下降后开始升高,在96h PI时达到峰值,而肌原蛋白mRNA水平早在12h PI时就升高;3) IGF-II水平在48 h PI前呈先下降后上升趋势,在72 h PI时达到峰值;4)最初下降后,bFGF mRNA水平在96 h PI时达到最大值;5) MHC和titin mRNA水平在72 h PI显著升高。表2⇓总结这些数据。
组织学研究
在以前的一项研究中,有报道称,局部应用咖啡因可诱导大鼠膈肌肌膜损伤在1小时PI时达到可重复的峰值,伴随最大的力损失2.. 事实上,尽管H&E对炎症过程没有真正的特异性,也不允许目前的作者区分炎性细胞群,但研究表明PI炎性浸润出现的时间晚于心肌细胞膜损伤,并且仅在48–72岁时才出现 h PI。炎症浸润的作用仍有待确定,不排除炎症可能导致参与肌肉修复过程的分子因子活化20.–22.Lapointe等.23提示非甾体抗炎药抑制肌肉损伤后的修复过程。
基因激活模式
蛋白酵素
P94,一种不可缺少的、肌肉特异性的、钙依赖的、非溶酶体半胱氨酸蛋白酶24,调节消化蛋白酶的功能25.calpain在败血症引起的肌肉损伤时被激活26以及电阻性负载引起的膜片损伤6.,16.与其他因素相比,p94 mRNA没有降低,但在损伤后立即达到峰值,同时肌膜损伤和力丧失最大2.. 这一快速增加表明,咖啡因诱导的细胞内钙立即升高可能触发p94 mRNA上调。尽管由于组织数量较少,无法对酶的活化水平进行量化,但有理由认为p94 mRNA峰促进了酶的合成,并导致其活化形式的更大可用性。此外,钙蛋白酶与细胞迁移和信号传递有关27、成肌细胞融合和肌原蛋白表达28.
转录因子
原癌基因cFos可促进肌细胞修复和肥大,对两种损伤均有即时反应29和机械应力30.,31.而肌生成素是肌肉发生和再生过程中终末肌肉分化的有效调节因子32.–34..出乎意料的是,肌生成素早在PI 12小时就升高了,而cFos在PI 72小时升高的比预期的晚。这与肌生成素转录本的减少相一致,可能是因为肌生成素抑制了cFos启动子35..此外,最高水平的炎症反应与cFos的升高同时发生,因此,这表明一些促炎因子在诱导cFos表达和mpc分裂方面具有潜在作用36.,37..作为终末分化标记,肌原蛋白基因的晚期表达是预期的。然而,一些外周肌肉挤压损伤或中毒性损伤的研究已经在卫星细胞的肌核/核中检测到早期肌原蛋白mRNA和蛋白的表达29,38.,39..这意味着有一个从属细胞亚群的存在,该亚群在没有先前有丝分裂的情况下立即开始最终分化,而其他mpc只在之后增殖和分化40. 这些高度抗辐射的卫星细胞,在短期内保持形成肌肉的能力,并直接进行终末分化40,可能存在于横膈膜中。
生长因子转录本
bFGF和IGF-II分别在PI 24和72 h时升高。bFGF是一种强的有丝分裂因子41.,增强肌肉再生42.与肌源性细胞迁移43..横膈膜bFGF的表达尚不清楚。缺氧诱导的过度通气增强44.在MDX小鼠中降低45..目前的作者发现bFGF mRNA的升高持续了96小时,证实了bFGF参与了膈肌修复程序。IGF-II在肌肉修复过程中起信号因子的作用46.,激活mpc增殖和终末分化47..关于IGF-II在横膈膜中的表达和作用知之甚少。虽然igf似乎参与了隔膜重塑,但大多数现有数据与IGF-I有关。IGF-II增加膈肌比力48.,促进纤维生长,保持其大小49.,50.,并预防肺气肿性仓鼠皮质激素诱导的膈肌萎缩51..相反,大量皮质类固醇治疗后,大鼠膈肌中IGF-I和-II的表达降低52..根据目前的数据,IGF-II转录本在隔膜修复过程中上调。此外,IGF-II mRNA只有在肌生成蛋白基因激活后才会增加,这在肌生成模型中得到了证实47..IGF-II mRNA升高在结构蛋白转录物表达之前,在MHC和三肽转录物表达和力回收期间保持高水平2..这表明IGF-II参与了结构蛋白的构建,最近的一项研究表明,IGF-II促进骨骼肌力量和肌纤维异质性,这一点很有趣53..
最后,MHC和titin mRNA的晚期增加,伴随着炎症浸润消退和功能恢复2.,证实膈肌修复过程的效率。MHC mRNA较早出现表明收缩蛋白基因先于肌联蛋白等非收缩蛋白基因活化。肌凝蛋白转录本的最大表达量,这是一种弹性蛋白,被认为是一种帮助肌凝蛋白丝在肌节中定位的分子54.,仅在240时观察到 h PI。
研究的局限性
Caffeine-induced损伤模型
咖啡因引起的损伤并不是经典意义上的生理损伤模型。尽管如此,在目前的研究中,目前的作者对咖啡因肌肉损伤模型的偏好有两个方面:1)已经证明,局部应用咖啡因溶液会引起可重复的膈肌损伤,并导致膈肌力量的大幅减少2.;2)膈肌暴露在咖啡因中产生损伤,而不受其他容易造成肌纤维损伤的因素的干扰,如缺氧和酸中毒,如在阻力负荷模型中所见16.因此,咖啡因诱导的伤害的口语是它消除了几个混杂因素。此外,已显示骨骼肌咖啡因暴露刺激CA2+通过激活瑞尼诺受体从肌淋式网释放,从而增加细胞内Ca2+浓度55.. 细胞内钙的大量升高是肌肉损伤的一个已知因素。一些研究表明,咖啡因诱导的细胞内钙释放可引起哺乳动物肌膜损伤56.肌丝降解57..
H&E染色允许评估组织学损伤。如前所述,该技术不允许特异性分化炎性细胞类型,但清楚地显示细胞浸润(图1⇑).
rt - pcr
本研究旨在筛选和监测膈肌损伤后参与心肌细胞再生的几个因素的基因表达时间过程。RT-PCR是检测所选因子的少量mRNA的最合适技术,因为从大鼠膈肌获得的损伤组织数量非常有限。尽管RT-PCR不能提供有关产生的蛋白质的存在、数量或位置的信息,但mRNA水平的显著改变对于相关基因的PI激活是可重复的和有用的。GAPDH作为管家基因的适用性通过当前结果的内部一致性得到了证明。由于GAPDH表达水平在研究所涵盖的整个PI期间保持不变,因此从当前数据中出现的不同时间基因表达模式是可能的。
结论
总而言之,目前的作者得出结论,咖啡因诱导的损伤触发了一种基于时间的多种基因表达程序,至少在大鼠横膈膜中,满足肌肉修复的需要。这些基因的表达模式涵盖了涉及肌肉完全修复的各种不同现象,包括从受损肌肉蛋白的蛋白质水解到新的肌肉纤维的合成。未来的研究需要阐明在其他慢性膈肌损伤情况下,如败血症和慢性阻塞性肺疾病,肌肉修复过程中基因表达的确切模式。
致谢
作者要感谢O.Da Silva(加拿大QC蒙特勒艾尔Hôtel Dieu医院CHUM研究中心研究支持办公室)的编辑协助。
- 收到了2004年4月23日。
- 认可的2004年11月12日。
- ©ERS期刊有限公司










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}