





摘要
肺缺血-再灌注损伤在心脏手术,特别是胸部器官移植后肺功能障碍中的中心作用已经得到公认。
长期缺血引起的肺组织坏死是肺功能恶化的主要原因。最近的研究表明,缺血-再灌注后的肺凋亡在术后肺功能障碍的发展中同样重要。
本文就目前有关肺功能障碍的机制和途径,特别是肺缺血再灌注后细胞凋亡的相关文献作一简要综述。
更好地了解肺凋亡及其上游途径,可能有助于制定治疗策略,使心肺移植患者受益。
缺血再灌注引起的肺功能障碍是心脏外科特别是肺移植中一个重要的临床问题1.体外循环术后肺功能障碍是一种常见的与肺缺血-再灌注损伤相关的现象2,3..高达20%的患者在肺移植术后早期出现危及生命的移植物功能障碍1.然而,了解血糖再灌注诱导的肺损伤的复杂病理生理学仍然不完整。与人体中的任何其他器官不同,肺部拥有两个血液供应网络,具有广泛的吻合点,并且共有三种潜在的肺组织氧气来源,从而使肺部血液再灌注损伤更加有趣的研究2.自20世纪80年代中期以来,广泛研究了中性粒细胞,自由基和其他炎症介质的作用,已被广泛调查4然而,这些介质似乎仅部分参与了肺缺血-再灌注损伤。肺缺血和再灌注后,导致肺上皮细胞损伤、坏死或凋亡的最终途径仍有待完全阐明。最近,肺缺血后肺细胞凋亡的相关机制尚不清楚aemia–再灌注已经开始被理解5.目前的评论突出了急性肺患者再灌注损伤后细胞凋亡的最新研究结果。
细胞凋亡是否参与肺缺血-再灌注损伤?
肺缺血-再灌注可导致肺上皮组织细胞破裂和死亡,这可能导致体外循环和肺移植后肺功能障碍的程度和持续时间2,4.自20世纪90年代末以来,科学家们已经认识到,缺血-再灌注后的不同过程虽然密切相关,但通过激活不同的炎症途径导致肺损伤2,4,6.因此,这些变化可能导致肺细胞坏死或凋亡。肺缺血-再灌注损伤后的组织坏死已被确认与肺功能显著恶化相关,与高度炎症相关。然而,缺血-再灌注与肺凋亡之间的复杂关系才刚刚开始显现。
细胞凋亡,也称为程序性细胞死亡,由Kerr重新引入现代临床实践et al。7,尽管这个词最初在医学上的使用可以追溯到希波克拉底时代(c。460-370 BC)和Galen(AD 129-201)8.细胞凋亡在希腊语中的意思是“脱落或脱落”,这个过程是正常器官发育的基础。形态学上,细胞凋亡的特征是细胞核收缩,染色质浓缩和DNA片段到寡核小体大小的片段,而质膜和细胞内细胞器保持完整9.最近的证据表明,细胞凋亡也可能在肺癌,间质肺纤维化和成人呼吸窘迫综合征(ARDS)如肺癌等肺部疾病过程中起重要作用10.,11..有许多因素包括细胞死亡的正常和异常调节,导致肺上皮细胞中的细胞凋亡,这可能在每个独特的情况下具有不同5.
在啮齿类动物单肺移植的实验研究中,短时间的缺血(20分钟)和再灌注与移植肺相比,移植肺的细胞坏死和凋亡均未增加12..在长时间(长达18小时)冷缺血肺保存无再灌注后,可观察到高百分比的细胞坏死(而非凋亡)12.,13..众所周知,当移植肺遭受长时间冷缺血时,肺上皮细胞再灌注会增加损伤程度,因为细胞坏死和凋亡的数量显著受缺血时间的影响。例如,在中度冷缺血保存(6或12小时)后,再灌注时移植肺的肺上皮细胞死亡模式以凋亡为主(<2%坏死,30%凋亡)。相比之下,在持续18或24小时的冷缺血后,这种细胞死亡方式以坏死为主(21-29%坏死,<1%凋亡)。12..同样,在临床肺移植中,在冷或甚至温暖的缺血后,几乎没有发现细胞凋亡持续时间最多5小时14..肺泡上皮细胞的凋亡在再灌注后才变得明显,而且凋亡细胞的数量随着再灌注时间的延长而增加14..在再灌注后,发现缺血性肺肺气胸凋亡在2小时内达到峰值,随后随后延长再灌注13..此外,细胞凋亡与总缺血时间、保存液或肺切除前的通气时间无关14..因此,在冷暖条件下单独的肺缺血,或仅在短时间的冷缺血后再灌注肺,似乎对肺凋亡的影响很小。而长时间缺血后肺再灌注会导致再灌注后早期出现明显的细胞凋亡,缺血时间进一步延长则会导致坏死。公认的再灌注肺损伤理论主要是基于再灌注后活性氧(ROS)的产生,导致细胞损伤和凋亡。另一个有趣的概念是细胞凋亡是一个能量依赖的细胞死亡程序。因此,长时间缺血后细胞凋亡的缺失可能仅仅是能量储备衰竭的反映。此外,延长的肺缺血可能通过增加抗炎细胞因子,如白细胞介素(IL)-10的水平,使平衡向肺上皮细胞坏死而不是凋亡转变15..
肺缺血再灌注后肺凋亡程度与肺功能障碍之间的直接关系尚存争议。有趣的是,缺血-再灌注后肺功能的恶化,包括动脉氧压下降(PA,O.2),只与细胞坏死百分比相关,而不是细胞凋亡的百分比12.,15..较差的肺功能与更多的细胞坏死可能部分与周围坏死细胞的高度炎症有关。这种现象在细胞凋亡中是不存在的,因为凋亡细胞很容易被巨噬细胞吞噬,从而限制了炎症细胞内酶以细胞膜碎片的形式从凋亡细胞中释放出来16..人肺移植再灌注后凋亡细胞的百分比与某些临床参数无相关性,如PA,O.2, 30天死亡率或术后机械通气时间14..在啮齿类动物肺移植模型中也发现了类似的结果,肺功能(PA,O.2,潮湿:干重比和峰值气道压力)和细胞凋亡程度17..实际上,改善的肺功能可能与减少的促炎症有关17.但增强抗炎15.细胞因子的反应。然而,在一项实验研究中,肺缺血再灌注后细胞凋亡的数量与肺分流率之间以及细胞凋亡与细胞氧化损伤之间均存在显著的相关性18..因此,人类研究似乎证实了动物模型的数据,即凋亡主要是一种再灌注后事件,缺血-再灌注后的细胞坏死程度似乎与肺功能紊乱的相关性强于与凋亡的相关性4,12..虽然已有大量实验研究表明细胞凋亡与肺功能障碍之间存在关系,但主要是在ARDS等急性肺损伤的情况下进行的,与缺血再灌注引起的肺损伤特异性相关的较少10..
肺缺血再灌注损伤细胞凋亡的机制
细胞凋亡可以通过机械损伤和暴露于某些环境条件引发,这导致活化内在(线粒体)和外在(死亡受体)途径(图1⇓)。通过特异性受体 - 配体相互作用激活外部途径,例如Fas / Fas-LigAnd(FAS-L),血管紧张素(A)II和肿瘤坏死因子(TNF)/受体,导致随后激活涉及半胱天冬酶的细胞内凋亡级联(半胱氨酸阿斯巴氨氨酸蛋白酶)。具有细胞组形成,称为线粒体途径,称为线粒体途径,称为线粒体途径,涉及BH3相互作用的域死亡激动剂(BID)切割,线粒体膜渗透性,线粒体细胞色素C和凋亡蛋白酶激活因子(APAF)释放。此外,在周围细胞环境中,血管内皮生长因子,包括血管内皮生长因子,血小板衍生的生长因子,表皮刺激因子,包括血管内皮生长因子,血小板衍生的生长因子和巨噬细胞菌落刺激因子的水平可以通过细胞内蛋白酪氨酸激酶和蛋白质酪氨酸磷酸酶影响肺上皮细胞凋亡.这些途径可以由某些丝裂原激活的蛋白激酶(MAPK)和来自B细胞白血病/淋巴瘤-2基因产物(BCL-2)家族的分子下调。磷脂酰肌醇-3'-激酶和蛋白激酶B途径也已显示在肺细胞凋亡中是重要的。然而,没有研究专门探讨了该途径在肺患者再灌注诱导的细胞凋亡中的作用19..这些细胞内促凋亡因子和抗凋亡因子之间的平衡可能最终决定一个细胞是否存活或下行通路经历凋亡。目前,各凋亡因子及凋亡通路在缺血-再灌注肺损伤中的相对重要性尚不明确。此外,临床环境中其他外部因素与上皮细胞凋亡的相互作用(即。呼吸机诱导的肺上皮细胞凋亡或坏死)、中性粒细胞激活和凋亡,以及来自全身炎症反应的介质,也可能显著影响最终结果10..目前主要研究缺血再灌注诱导肺实质细胞凋亡,而非炎症细胞凋亡(即。中性粒细胞和巨噬细胞)。在这两种细胞群中,凋亡的调节对缺血-再灌注后肺损伤的程度可能有不同的结果。
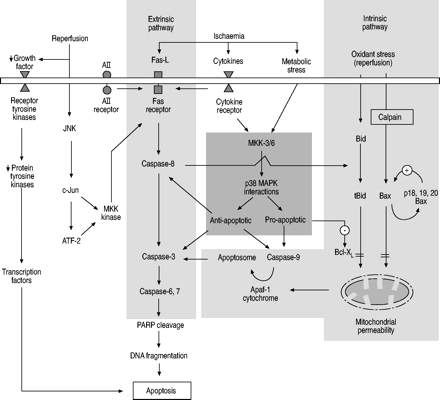
主要的内在(线粒体)和外在(死亡受体)肺缺血-再灌注损伤后调节凋亡的途径。还显示了影响抗凋亡和促凋亡介质之间平衡的重要途径。AII:血管紧张素II;Fas-L:Fas配体;JNK:c-Jun N-末端激酶;MAPK:丝裂原激活蛋白激酶;MKK:MAPK激酶;ATF-2:转录因子2激活;PARP:聚ADP核糖聚合酶;Apaf:凋亡蛋白酶激活因子;Bid:BH3相互作用域死亡激动剂;tBid:截短Bid;Bax:Bcl-2相关X蛋白;Bcl-Xl:Bcl-2-like 1。↓: 减少;+:激活;-:抑制作用; = : 抑制(阻断)。
外在途径
Fas和caspase级联,也被称为外源性途径,可能是细胞凋亡起始最有力的途径。Fas (CD95)是一种细胞膜受体,可在多种细胞类型中发现,包括肺泡上皮细胞。其相应的配体Fas- l (CD95L)是TNF家族的成员细胞因子,其与Fas受体的相互作用可以触发凋亡途径的启动,特别是通过激活caspase-8。研究表明,在肺组织缺血后,启动caspase (caspase-8和caspase-9)以这种方式被激活20..随后,通过蛋白裂解刽子手caspase (caspase-3, -6和-7)的激活可以在凋亡级联中攻击它们的细胞底物20.,导致更多的下游caspases和多聚腺苷二磷酸核糖聚合酶的激活和随后的DNA片段。此外,在缺血肺中,另一个重要的凋亡途径涉及通过Bid切割和线粒体细胞色素c释放激活本征通路的caspase-820.,21.,尽管在缺血条件下,通过TNF和Fas死亡信号传导的替代直接外源途径也是可能的21..在肺缺血-再灌注过程中抑制上述任何一种半胱天冬酶均可减轻肺凋亡20..
众所周知,在支气管肺泡液和肺泡细胞中,急性肺损伤或急性肺损伤或ARDS,FAS和FAS-L水平显着增加,所述肺泡液和上皮细胞10.,22..这种增加可以通过可溶性FAS-L的肺部局部释放部分占核对22.,23.来自活化的肺中性粒细胞和中性粒细胞细胞膜裂解过程中膜结合Fas-L的释放23..与肺损伤相关的促炎细胞因子水平的增加可以通过抑制中性粒细胞凋亡和延迟抑制炎症反应的延迟延长中性粒细胞寿命,并且可以延长FAS-L释放23..有趣的是,肺损伤患者的肺部损伤和多器官失败以及肺部患者的血液表现和多器官衰竭以及多器官衰竭以及临床结果不良,有趣22..在实验环境中,肺缺血再灌注也被证明会导致Fas和Fas- l表达增加,并导致肺组织和肺动脉内皮细胞的相关凋亡20..用Fas- l抗体消除或拮抗Fas可显著降低肺缺血再灌注后细胞凋亡20..
丝裂原活化蛋白激酶的作用
已知MAPK信号通路在控制细胞增殖、分化和凋亡中起重要作用。各种MAPK模块(包括p38和c-Jun N-末端激酶(JNK))的直接和间接激活,等等。)通过细胞外刺激最近被整齐总结了24.,25..p38 MAPK是MAPK超家族的成员,其有三种亚型α、β和γ,参与细胞周期调控和存活的细胞膜到细胞核信号转导。通过MAPK激酶-3、-4和-6激活p38 MAPK可在代谢应激、细胞因子和缺血反应中发生。p38 MAPK活性水平与肺缺血-再灌注损伤后的促凋亡(通过产生促炎症细胞因子和Bid切割)和抗凋亡活性有关20..
在啮齿动物肺缺血-再灌注模型中,抑制p38 MAPK激活导致促炎细胞因子IL-1β水平降低,这与更少的肺泡损伤和肺水肿有关,效果更好PA,O.2再灌注后26..此外,在犬肺移植模型中,在冷藏期间加入Euro-Collins溶液中的p38 MAPK抑制剂可显著改善其抑制作用PA,O.2,肺泡-动脉氧张力差,肺血管阻力和心输出量,以及减少肺水肿和中性粒细胞浸润27..心肺功能的改善与肺再灌注30分钟后p38 MAPK的磷酸化抑制相对应27.,28..有趣的是,在鼠异体和同种异体肺移植模型中,p38 MAPK的激活在再灌注2小时后显著降低,表明p38 MAPK的激活可能也具有一些抗凋亡的特性28..这种促凋亡和抗凋亡特性的不一致可能是由于不同的MAPK激酶和p38 MAPK亚型(α, β和γ)在不同刺激的影响下被激活或组合而引起的20..例如,MAPK激酶3优先激活p38-α和-γ,而MAPK激酶6激活p38-α、-β和-γ。活化的p38-α MAPK亚型通过抑制caspase-3、-8和-9在肺缺血-再灌注损伤动物模型中发挥抗凋亡作用20.,可能通过诱导细胞保护性血氧合酶-1基因29.. p38 MAPK和MAPK激酶亚型活性的不同组合对肺缺血再灌注损伤后细胞凋亡的影响仍有待阐明。
JNK是MAPK家族的另一成员,已知参与应激诱导的细胞凋亡。特别是,氧化应激,如再灌注后发现的氧化应激,可能诱导中性鞘磷脂酶活性,导致信号分子神经酰胺的积累30.随后,神经酰胺可激活JNK,导致c-Jun磷酸化和转录因子(ATF)-2的激活11..ATF-2和c-Jun可以结合并打开MAPK激酶激酶启动子,导致Fas-L的转录增加。这也可能是氧化诱导肺上皮细胞凋亡的重要信号机制。
酪氨酸磷酸化的作用
已知酪氨酸激酶的细胞酪氨酸磷酸化可防止多种细胞类型的凋亡并促进细胞存活。促凋亡酪氨酸磷酸酶和抗凋亡酪氨酸激酶之间的平衡对决定细胞存活非常重要。人肺冷缺血保存2-5年后 h、 蛋白酪氨酸磷酸化活性增加,从而有利于肺移植后和再灌注前的抗凋亡行为31.然而,在肺再灌注2 h后,酪氨酸激酶的酪氨酸磷酸化活性显著下降,然后恢复到基础水平,这可能与实验和人肺移植中再灌注后早期凋亡细胞死亡的发展有关28.,31.有趣的是,在实验移植研究中再灌注缺血性肺部后,促凋亡蛋白酪氨酸磷酸酶的活性也降低了2小时28..因此,促凋亡酪氨酸磷酸酶和抗凋亡酪氨酸激酶活性之间的平衡可能比它们各自的水平更为重要。
血管紧张素II的作用
经典上,肾素 - 血管紧张素系统(RAS)已被视为血压调节中的内分泌系统。然而,最近的证据表明,RAS可以通过帕拉卡碱和自分泌机制参与细胞凋亡的调节32.局部内在RAS在远端肺实质中表达,通过AII在肺泡上皮细胞中的凋亡中发挥着核心作用,通过与上皮AII受体相互作用,对于FAS介导的细胞凋亡10.,11.,33.在患者在各种形式的急性肺损后,在支气管肺泡中发现了转化AI至AII的血管紧张素转换酶(ACE)水平,其在支气管肺泡中发现10.,11..显示对啮齿动物肺部血糖再灌注模型中ACE的抑制导致再灌注后显着更好PA,O.2和峰值气道压力,较少的肺水肿比对照34.ACE抑制对肺功能的保护似乎与中性粒细胞隔离或脂质过氧化无关,提示AII可能对缺血-再灌注后的肺功能产生不利影响通过其他途径,如Fas/Fas- l凋亡34.
bcl - 2家族(投标/伯灵顿/ Bcl-Xl)
氧化应激可诱导肺泡上皮细胞内促凋亡蛋白Bid和bcl -2相关x蛋白(Bax)的表达35,通过内在途径引起细胞凋亡。蛋白裂解Bid,使截短的Bid和Bax与通透性过渡孔复合物结合,增加线粒体通透性,导致线粒体内跨膜电位耗散,细胞色素c通过线粒体外膜释放35,36.此外,BAX可以通过CALPAIN切入P18和P19 BAX,这可以为BAX和线粒体之间的相互作用提供阳性反馈37.线粒体释放的细胞色素c和Apaf-1,与caspase-9一起形成凋亡小体,激活caspase-3,随后caspase级联导致细胞凋亡。有趣的是,肺组织缺血和再灌注对促凋亡Bax的表达没有影响20..
bcl-2状1(bcl-xl)属于Bcl-2家族,可防止多种刺激诱导的细胞凋亡通过半胱天冬酶抑制。此外,Bcl-Xl已被证明可以防止缺氧诱导的肺上皮细胞凋亡38. 它作用于Bid下游,其表达可抑制激活的Bid诱导的所有凋亡变化,从而改善线粒体损伤21.,减少线粒体细胞色素c的释放,限制caspase-9的激活38.此外,Bcl-x可以防止Bax对线粒体膜的影响l35.缺血-再灌注损伤可降低抗凋亡蛋白Bcl-2和Bcl-X的水平l,可能通过MAPK激酶3/p38 MAPK途径20..凋亡抑制蛋白和线粒体衍生的caspases通路的第二激活因子在肺癌凋亡中也被证明是重要的39,40. 然而,迄今为止还没有研究专门探讨这些通路在肺缺血再灌注诱导的细胞凋亡中的作用。
缺血-再灌注诱导的肺凋亡的其他刺激
反应性氧气
ROS在ARDS和肺缺血再灌注损伤中的重要性早已被认识41.活性氧可由活化的中性粒细胞通过还原的烟酰胺腺嘌呤二核苷酸磷酸(NADPH)氧化酶途径产生4,或从肺部血液再灌注后的肺实质细胞(缺氧重新氧化)。iscaemia可以赋予肺组织,从而在再灌注时重新引入分子氧导致有毒氧代谢物的产生通过线粒体细胞色素P450.系统42.缺氧与ATP降解相关,已发现可激活细胞p38 MAPK,通过磷酸化激活黄嘌呤氧化酶,从而在肺组织再灌注(再引入氧气)时产生ROS43,44.此外,一氧化氮合酶(NOS)和最近发现的非吞噬细胞膜/胞浆NADPH氧化酶也可能参与ROS的生成,作为凋亡的信号分子44,45.ROS可通过以下途径诱导肺细胞损伤:1)直接提高内皮细胞内钙水平,激活钙依赖性蛋白酶46(2)通过激活MAPK作为凋亡信号的生化中间产物,导致细胞色素c释放和随后的聚ADP核糖聚合酶切割44,45.
一氧化氮
活性氧(ROS),如NO,通过生成其他氧化介质直接或间接介导细胞缺氧-复氧损伤。高浓度NO可使许多线粒体呼吸酶失活,包括线粒体乌头酸酶、烟酰胺腺嘌呤二核苷酸-泛素氧化还原酶(复合物1)和琥珀酸-泛素还原酶(复合物2),以及细胞色素氧化酶47.此外,缺氧条件会增加诱导性NOS的产生,这可能会增加氧化自由基的副产物,如过氧亚硝酸盐阴离子,导致ros型细胞损伤44.然而,NO也可能发挥细胞保护的抗氧化作用,并减轻复氧损伤,特别是当来自组成性NOS时。例如,缺氧复氧后组成性NOS的表达降低,通过增加Fas和降低Bcl-2的表达,促进细胞凋亡48.
细胞因子
在肺缺血和再灌注后,肺组织和体循环中一些促炎细胞因子(如TNF和IL-8)的水平显著升高,这些因子被认为可以刺激肺中性粒细胞迁移、激活和脱颗粒4,49.在临床肺移植中,再灌注后肺组织中的IL-8水平显着与低电平相关PA,O.2,高气道压力和急性生理和慢性健康评估评分,这可能导致更长的重症监护室住院时间和更高的术后死亡率49.缺血-再灌注肺损伤后TNF局部释放增加可能通过JNK活化诱导细胞凋亡,而JNK活化促进caspase-8和Bid活性,最终导致肺功能障碍50.
已经发现抗炎细胞因子IL-10通过抑制炎症和T细胞介导的免疫反应来保护肺部免受isChemia再灌注损伤。内源性IL-10可以通过平衡血浆纤溶酶原激活剂抑制剂和组织纤溶酶原激活剂的活性,降低肺血管纤维蛋白沉积和微血管血栓形成,并且在这样做的情况下,也衰减了IL-1的释放51.在啮齿动物肺缺血-再灌注模型中,IL-10可以通过降低肺血管通透性来限制肺再灌注损伤,可能是通过抑制肺泡巨噬细胞TNF-α的产生52.
在缺血-再灌注肺损伤后,促炎细胞因子和抗炎细胞因子与肺功能的相互作用已经得到了很好的研究。然而,细胞因子网络对缺血-再灌注肺损伤后细胞凋亡的影响尚不清楚。在一个IL-10基因转染模型中,IL-10通过改变细胞死亡模式,使细胞凋亡而不是坏死,从而改善肺移植后的肺损伤15..然而,在另一个肺移植模型中,高水平的IL-10和低水平的TNF-α与肺组织中凋亡细胞的数量或Bcl-2的表达无关17..
细胞凋亡和缺血-再灌注肺损伤的预处理
在主要缺血性侮辱之前短暂的缺血性和再灌注造成的缺血预处理明显限制了心脏,肝脏和肾脏等器官的缺血再灌注损伤程度。然而,对预处理对肺部的类似保护作用很少。由于其双血供水和通风,在肺中创造缺血预处理比其他器官更复杂;因此,研究更难以研究。例如,据报道,在通风期间阻断肺血液供应不会导致ATP水平降低53.然而,肺萎陷时停止通气可能不同于缺氧通气,因为萎陷和再充气的机械作用比缺血挑战本身更重要53.在肺缺血预处理实验模型中,肺缺血再灌注损伤标志物(即。缺血再灌注损伤后,预处理组的丙二醛(丙二醛和谷胱甘肽)和肺动脉压均低于未预处理组54.此外,肺中的预处理诱导的热休克蛋白合成还有助于保持离子输送和肺泡流体清除,从而保持气体交换功能并降低肺部缺血性侮辱后的肺血管渗透性55.有趣的是,实验研究表明,缺血预处理的持续时间和模式在实现针对血糖再灌注肺损伤的充分保护作用方面也很重要。重复而不是单一的缺血预处理可以赋予肺功能保存更好的肺功能保存,如肺结血再灌注肺损伤53.此外,全身缺血或热预处理的概念最近获得了普及55.
在肺部缺血再灌注之前,各种化学物质也已用于肺预处理,包括吸入不含和3-硝基丙酸盐56,57.所提出的机制,其中不提供预处理,包括肺中性粒细胞凋亡的诱导,抑制来自肺泡巨噬细胞的中性粒细胞和炎性细胞因子的超氧化物释放56.线粒体复合物II的抑制剂,3-硝基丙酸,通过抑制再灌注后细胞的氧化磷酸化和氧化应激,也能先决条件肺和减轻随后的缺血-再灌注损伤57.
临床意义
细胞凋亡及其上游途径明显促进了肺缺血-再灌注损伤的发生和一些相关的临床表现。实验和临床研究证实了两个主要途径,即外部Fas-L-caspase和内在线粒体途径,通过这两个途径,肺上皮发生程序性细胞死亡。在肺缺血和再灌注后的凋亡反应的微调中,有许多调节介质和补充通路。了解这些调控缺血-再灌注后肺凋亡的途径可以为减轻急性肺损伤的严重程度开辟新的前景。例如,制药(一氧化碳)20.和生物分子干预(FAS-L)58,以及腺病毒和质粒的基因操作15.,58,最近提出了。这些潜在的治疗策略可以有助于预防凋亡途径的繁殖,随后,可以降低接受心肺移植患者的围手术性发病率和死亡率。
总结
自20世纪90年代初以来,凋亡及其介质在肺部缺血再灌注损伤中的显着作用已经开始理解。新兴的证据表明,肺部缺血再灌注后细胞凋亡的调节可能比以前设想更复杂。令人惊讶的是,肺细胞凋亡与肺部血症再灌注引起的肺功能障碍之间相关的证据并不完全一致。在血液再灌注后肺部的预期和高度选择的细胞凋亡可能不会像以前认为一样损坏。相反,细胞坏死与其相关的炎症,对肺可能会更伤。与再灌注相比,单独血症似乎只对肺细胞凋亡产生了微小的影响。外在和内在凋亡途径以及它们相关的调节因素的全部程度仍应阐明肺部缺血再灌注凋亡的凋亡。在分子水平需要进一步探索其他胱天蛋白酶在肺部缺血再灌注过程中的作用。对肺细胞凋亡引发和繁殖负责的上游途径的未来研究将是在肺缺血再灌注和肺移植后最小化肺功能障碍的可能治疗策略的基础。
- 已收到2004年3月10日。
- 接受2004年8月21日。
- ©ERS期刊有限公司










{kind=link}
{kind=link}