





文摘gydF4y2Ba
的机制gydF4y2Ba铜绿假单胞菌gydF4y2Ba殖民支气管扩张患者的气道和囊性纤维化是知之甚少。gydF4y2Ba铜绿假单胞菌gydF4y2Ba可以通过坚持逃避黏膜纤毛的间隙区域剥蚀的基底膜完整呼吸道上皮细胞。gydF4y2Ba
作者开发了一个gydF4y2Ba在体外gydF4y2Ba模型来研究gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持它基底膜使用扫描电镜iv型胶原蛋白。gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持密度确定的数量gydF4y2Ba铜绿假单胞菌gydF4y2Ba每20个显微镜领域(2000×)日志inocular大小在37°C孵化后45分钟。gydF4y2Ba
phytohaemagglutinin (PHA) - e,特别结合d-galactose-β1-4-d -gydF4y2BaNgydF4y2Ba乙酰氨基葡萄糖,显著降低gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持密度与控制。肝素钠和钙的存在也显著降低gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持密度。gydF4y2Ba铜绿假单胞菌gydF4y2Ba依从性不受脯氨酸的存在,trans-hydroxyproline,甘氨酸,半乳糖,gydF4y2BaNgydF4y2Ba-acetylneuraminic酸,gydF4y2BaNgydF4y2Ba乙酰氨基葡萄糖或gydF4y2Ba落花生hypogeagydF4y2Ba。gydF4y2Ba
铜绿假单胞菌gydF4y2Ba可能依从性行为gydF4y2Ba通过gydF4y2Ba承认d-galactose-β1-4-d -gydF4y2BaNgydF4y2Ba乙酰氨基葡萄糖iv型胶原蛋白序列,这一过程可能会抑制肝素和钙。是持久的gydF4y2Ba铜绿假单胞菌gydF4y2Ba殖民囊肿性纤维化和支气管扩张患者是有害的,目前还没有有效的治疗根除,这些结果可能会导致新的治疗持久的gydF4y2Ba铜绿假单胞菌gydF4y2Ba感染。gydF4y2Ba
本研究支持了从香港研究资助局研究资助,中国香港特别行政区。gydF4y2Ba
支气管扩张,定义为病态的永久性扩张的支气管树,东亚人之间是一种常见的呼吸道疾病。没有有效的治疗支气管扩张和许多严重影响患者长期感染gydF4y2Ba铜绿假单胞菌gydF4y2Ba占重要的发病率和死亡率gydF4y2Ba1gydF4y2Ba。目前,唯一的治疗gydF4y2Ba铜绿假单胞菌gydF4y2Ba肺部感染是政府的抗生素,这是消除无效的gydF4y2Ba铜绿假单胞菌gydF4y2Ba。更好的理解的机制gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持这些患者的肺部有潜在的新发展的关键和新的治疗方法抗感染。gydF4y2Ba
人们普遍认为目标黏膜表面细菌坚持在疾病的发病机制中扮演了重要的角色,因为坚持为进一步建立锚固与宿主的相互作用gydF4y2Ba2gydF4y2Ba。细菌可以通过表达丰富的表面,实现这一过程,以特定的方式与上皮细胞表面受体结合。gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持大量的哺乳动物细胞类型包括口腔上皮细胞gydF4y2Ba3gydF4y2Ba、呼吸道上皮细胞gydF4y2Ba4gydF4y2Ba、呼吸道黏液gydF4y2Ba5gydF4y2Ba和暴露的胶原蛋白gydF4y2Ba6gydF4y2Ba。一个gydF4y2Ba在活的有机体内gydF4y2Ba最近的研究表明,gydF4y2BaP.aeruginosagydF4y2Ba坚持公开支气管结缔组织和管腔内的分泌物,而不是完整的囊性纤维化患者的呼吸道粘膜(CF)gydF4y2Ba7gydF4y2Ba。Collagen-binding蛋白质已确定gydF4y2Ba链球菌引起的肺炎gydF4y2Ba葡萄球菌,调解他们坚持哺乳动物细胞外基质材料gydF4y2Ba8gydF4y2Ba。利用透射电子显微镜,目前集团最近显示gydF4y2Ba铜绿假单胞菌gydF4y2Ba具有高度的亲和力人类基底膜胶原原纤维gydF4y2Ba在体外gydF4y2Ba9gydF4y2Ba。因此,坚持基底膜是一个重要的问题没有研究。因此,作者最近建立了一个模型来研究细菌粘附基底膜胶原蛋白和应用这一评估各种化学品的依从性的影响gydF4y2Ba铜绿假单胞菌gydF4y2Ba对胶原蛋白gydF4y2Ba在体外gydF4y2Ba10gydF4y2Ba。gydF4y2Ba
材料和方法gydF4y2Ba
接种的gydF4y2Ba铜绿假单胞菌gydF4y2Ba
临床孤立nonmucoid和piliated应变的gydF4y2Ba铜绿假单胞菌gydF4y2Ba(PACS001)存储在大脑/心浸液中含有20%甘油在液态氮。gydF4y2Ba铜绿假单胞菌gydF4y2Ba对大脑检索/心浸液琼脂(英国贝辛斯托克Oxoid)板块和孵化一夜之间在37°C。通过有限的前三次实验。在孵化后,一群gydF4y2Ba铜绿假单胞菌gydF4y2Ba激动于4毫升的大脑/心脏灌注6毫升透明塑胶管安装在辊阶段为24小时37°C。结果细菌悬液然后离心机在2000×10分钟gydF4y2BaggydF4y2Ba。上清液被抛弃,取而代之的是4毫升的磷酸盐(PBS;Oxoid)。这是重复三次洗细菌,最后resuspended在PBS。最后一个gydF4y2Ba铜绿假单胞菌gydF4y2Ba悬架是用于孵化与埃普多夫盖子(见下面的描述)。gydF4y2Ba
胶原蛋白涂层gydF4y2Ba
无菌人类iv型胶原蛋白(σ,圣路易斯,密苏里州,美国)解决方案(2 mg·毫升gydF4y2Ba−1gydF4y2Ba在1%乙酸)是每个实验前准备的。根据制造商,胶原蛋白后三大乐队dodecylsulphate-polyacrylamide钠凝胶电泳与基底膜胶原蛋白减少的条件下一致的gydF4y2Ba11gydF4y2Ba。盖子的塑料埃普多夫(微型离心机)管(美国索伦森,盐湖城,UT)被精心修剪过的,从身体中取出的管子通过高压灭菌和消毒。胶原蛋白溶液(50µL)添加到内部是一个倒埃普多夫盖子,允许在一个孵化器风干24小时保持在37°C。Collagen-coated盖子被沉浸在无菌PBS清洗三次,风干了30分钟的unhumidified孵化器在37°C。该协议提供一致和统一的涂层的iv型胶原在埃普多夫盖子(图1所示gydF4y2Ba⇓gydF4y2Ba)。gydF4y2Ba
{kind=link}

{kind=link}
扫描电子显微图显示测试表面涂有一层光滑的人类iv型胶原蛋白处理后成为微皱的边缘。酒吧= 500µm规模。gydF4y2Ba
孵化的gydF4y2Ba铜绿假单胞菌gydF4y2Ba与collagen-coated盖子gydF4y2Ba
铜绿假单胞菌gydF4y2Ba悬挂在PBS(50µL),其中含有没有或不同浓度的测试代理,被温柔小心地添加到collagen-coated盖子移液。活菌计数的接种gydF4y2Ba铜绿假单胞菌gydF4y2Ba悬架也执行确定细菌浓度和纯度。盖子被孵化gydF4y2Ba铜绿假单胞菌gydF4y2Ba悬挂在45分钟37°C unhumidified氛围。作者先前确定45分钟是最大最优的依从性没有显著改变gydF4y2Ba铜绿假单胞菌gydF4y2Ba活菌计数。孵化后,gydF4y2Ba铜绿假单胞菌gydF4y2Ba悬架是仔细倾析collagen-coated盖子。盖子是在无菌PBS溶液清洗三次(5毫升)删除不依从细菌。后,盖子固定在4%戊二醛和储存在4°C到处理电子显微镜。gydF4y2Ba
扫描电子显微镜的处理gydF4y2Ba
Collagen-coated盖子与孵化gydF4y2Ba铜绿假单胞菌gydF4y2Ba在4%的戊二醛固定24小时在钠甲次砷酸盐缓冲液冲洗之前,和1%的四氧化锇1 h后缀。分级标准脱水乙醇随后(三次三次50%,70%,在90%,三次,三次每5分钟)为100%到100%丙酮。标本被极度干在二氧化碳和安装在铝存根sputter-coated之前用金子包裹。这些标本随机编码并存储在单独的干燥管扫描电子显微镜检查之前的观察者不知道治疗。gydF4y2Ba
扫描电子显微镜的评估gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持collagen-coated埃普多夫盖子gydF4y2Ba
每个盖子放置在舞台上的扫描电子显微镜(SEM)和低倍镜下观察(200×)证实胶原涂层均匀,如图1所示gydF4y2Ba⇑gydF4y2Ba;否则,样品将被拒绝。对于每个标本,20个随机扫描电镜字段检查2000×放大中心的盖子。杆菌的数量计算手动为每个SEM字段。的总数gydF4y2Ba铜绿假单胞菌gydF4y2Ba杆菌被计算gydF4y2Ba铜绿假单胞菌gydF4y2Ba胶原蛋白表面密度,反映了gydF4y2Ba铜绿假单胞菌gydF4y2Ba对胶原蛋白在特定实验条件下亲和力。坚持密度计算的总数gydF4y2Ba铜绿假单胞菌gydF4y2Ba杆菌检测20 SEM领域除以inocular规模的对数gydF4y2Ba铜绿假单胞菌gydF4y2Ba,确定可行的菌落计数。gydF4y2Ba
凝集素的影响,阳离子、糖和其他试剂gydF4y2Ba铜绿假单胞菌gydF4y2Ba依从性gydF4y2Ba
试剂,从σ,购买,除非另有说明,以前所影响gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持或适当的生化反应在形式的胶原蛋白成分,喜忧参半的gydF4y2Ba铜绿假单胞菌gydF4y2Ba暂停来评估他们的影响gydF4y2Ba铜绿假单胞菌gydF4y2Ba依从性。CagydF4y2Ba2 +gydF4y2Ba提出了氯化钙(CaCl吗gydF4y2Ba2gydF4y2Ba·HgydF4y2Ba2gydF4y2BaO)解决方案(默克,柏林,德国)。中使用这些试剂的浓度gydF4y2Ba铜绿假单胞菌gydF4y2Ba悬架表1 - 4所示gydF4y2Ba⇓gydF4y2Ba⇓gydF4y2Ba⇓gydF4y2Ba⇓gydF4y2Ba。gydF4y2Ba
凝集素的依从性密度的影响gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持人类胶原iv型gydF4y2Ba
电荷的依从性密度的影响gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持胶原iv型gydF4y2Ba
胶原蛋白成分的影响氨基酸的依从性gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持胶原iv型gydF4y2Ba
糖的依从性密度的影响gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持胶原iv型gydF4y2Ba
统计分析gydF4y2Ba
数据表示为均值±sem,除非另有说明。魏克森讯号等级测试是用来比较配对数据相同的实验。的假定值< 0.05为两组数据之间的一个统计上的显著差异。gydF4y2Ba
结果gydF4y2Ba
一般的观察gydF4y2Ba
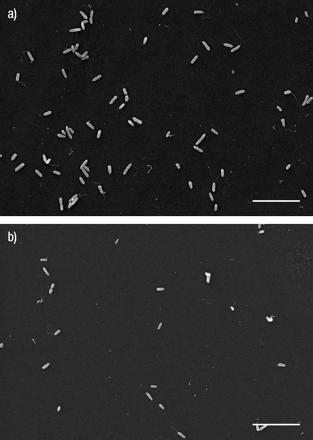
有一个一致的模式的扫描电镜检查gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持胶原表面。绝大多数的扫描电镜检查显示字段奇异相同的细菌附着的胶原蛋白,通常的长轴杆菌直接接触后者(图。2gydF4y2Ba⇓gydF4y2Ba)。gydF4y2Ba
{kind=link}
{kind=link}
扫描电子显微图显示collagen-coated表面附着gydF4y2Ba铜绿假单胞菌gydF4y2Ba杆菌在磷酸盐包含一个45分钟的孵化后)不phytohaemagglutinin (PHA) - e和b) 0.1 mg·毫升gydF4y2Ba−1gydF4y2BaPHA-E,明显减少了gydF4y2Ba铜绿假单胞菌gydF4y2Ba依从性。酒吧= 10µm规模。gydF4y2Ba
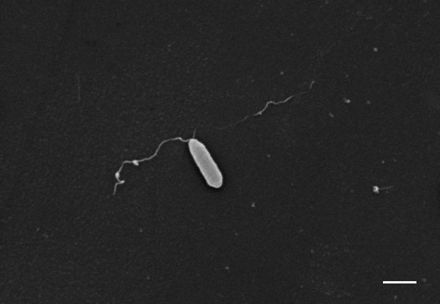
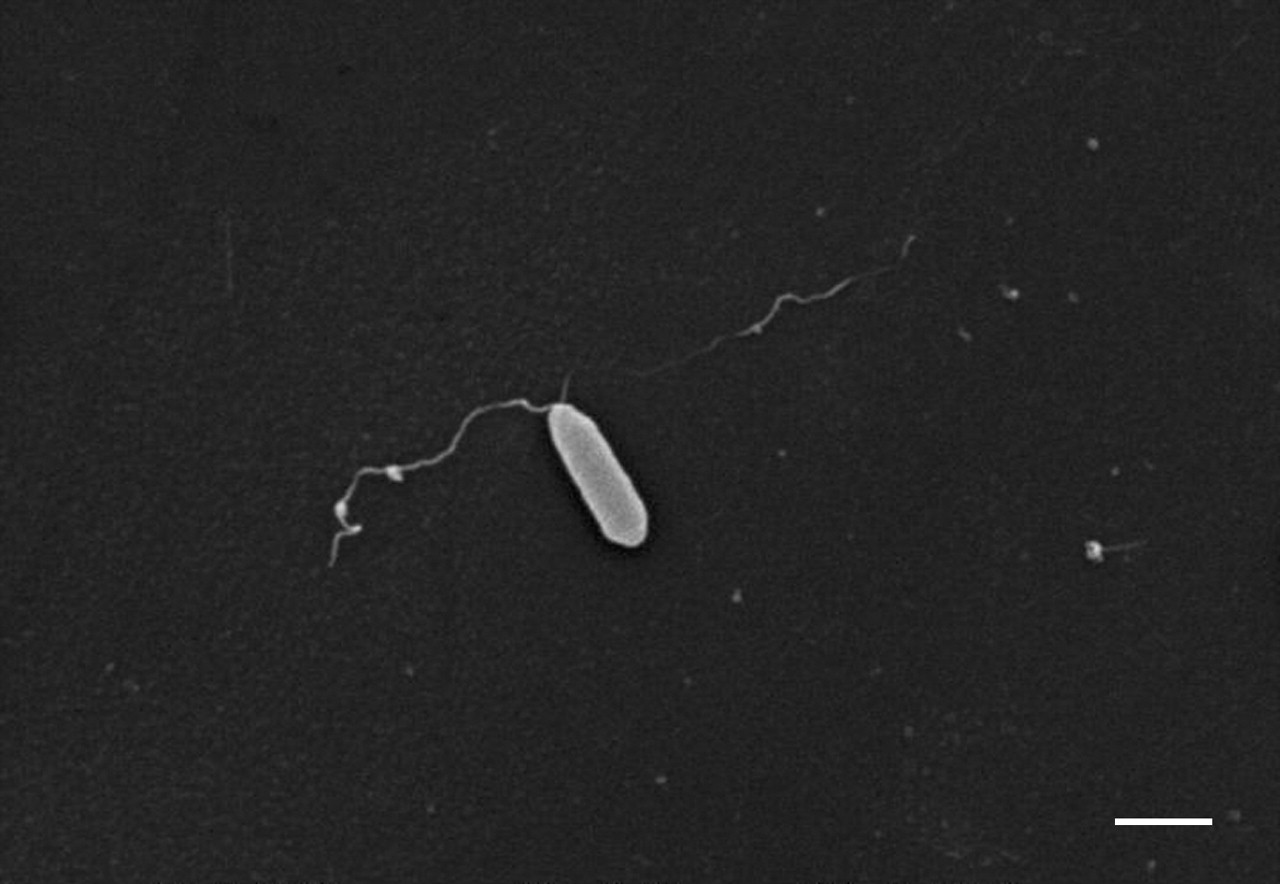
在< 1%的检查,SEM字段gydF4y2Ba铜绿假单胞菌gydF4y2Ba杆菌出现在一个集群中,就像一串葡萄。几乎没有证据表明超然的最初附着细菌,asthere没有细菌的“足迹”或其他可识别的胶原蛋白表面变形。细菌极地pili也发现胶原蛋白表面(图3所示gydF4y2Ba⇓gydF4y2Ba)。gydF4y2Ba
{kind=link}
{kind=link}
一个大功率的扫描电子显微照片显示的依从性gydF4y2Ba铜绿假单胞菌gydF4y2Ba杆菌胶原蛋白表面,通常在细菌长轴。有极地杆菌菌毛,这似乎与胶原蛋白表面。酒吧= 1µm规模。gydF4y2Ba
凝集素对gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持密度gydF4y2Ba
表1gydF4y2Ba⇑gydF4y2Ba表明phytohaemagglutinin (PHA) - e有抑制性影响gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持密度。PHA-E 0.1和1毫克·毫升的浓度gydF4y2Ba−1gydF4y2Ba显著降低gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持密度与缺乏PHA-E相比(p < 0.05)。的存在gydF4y2Ba落花生hypogeagydF4y2Ba似乎减少gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持密度,尽管没有统计学意义(p > 0.05)。gydF4y2Ba
费用的影响gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持密度gydF4y2Ba
表2gydF4y2Ba⇑gydF4y2Ba显示Ca的存在gydF4y2Ba2 +gydF4y2Ba在浓度为1、5、10毫米,但不是0.1毫米,明显减少了gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持密度相比,Ca的缺失gydF4y2Ba2 +gydF4y2Ba(p < 0.05)。似乎没有剂量依赖性抑制gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持胶原蛋白在CagydF4y2Ba2 +gydF4y2Ba测试。肝素也减少gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持密度显著的浓度10和100国际单位(IU)·毫升gydF4y2Ba−1gydF4y2Ba,但不是1000 IU·毫升gydF4y2Ba−1gydF4y2Ba相比,无肝素(p < 0.05)。gydF4y2Ba
胶原蛋白成分的影响gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持密度gydF4y2Ba
表3gydF4y2Ba⇑gydF4y2Ba表明,脯氨酸,trans-hydroxyproline和甘氨酸浓度的0.1,1,10 mg·毫升gydF4y2Ba−1gydF4y2Ba没有任何重大影响gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持胶原蛋白相比,缺乏测试试剂(p > 0.05)。gydF4y2Ba
糖的影响gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持密度gydF4y2Ba
表4gydF4y2Ba⇑gydF4y2Ba表明,半乳糖,gydF4y2BaNgydF4y2Ba-acetylneuraminic酸,gydF4y2BaNgydF4y2Ba乙酰氨基葡萄糖浓度的0.01,0.1,把mg·毫升gydF4y2Ba−1gydF4y2Ba没有任何重大影响依从性的gydF4y2Ba铜绿假单胞菌gydF4y2Ba胶原蛋白相比,缺乏测试代理(p > 0.05)。gydF4y2Ba
讨论gydF4y2Ba
作者描述了一种新的模式直接研究细菌粘附基底膜使用扫描电子显微镜gydF4y2Ba10gydF4y2Ba。通过使用表面附着的直接人工计数gydF4y2Ba铜绿假单胞菌gydF4y2Ba杆菌与扫描电子显微镜,他们已经确定胶原蛋白表面的附着细菌的确切数字。这可能是一个更直接的和具体的,尽管更费力,比先前的间接检测方法来确定细菌粘附来坚持,比如radiolabelling技术。Byrecent使用该模型,作者表明,gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持基底膜胶原蛋白减少在低剂量红霉素的存在,可能部分由于细菌morophology变更gydF4y2Ba10gydF4y2Ba。这些结果表明,外源凝集素PHA-E,但不是gydF4y2Ba答:hypogeagydF4y2Ba,显著抑制gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持胶原蛋白。PHA-E似乎抑制gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持为0.01,0.1和1毫克·毫升gydF4y2Ba−1gydF4y2Ba,尽管只有后两个浓度显著抑制依从性。CagydF4y2Ba2 +gydF4y2Ba抑制gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持的浓度> 0.1毫米,尽管也并未明显的剂量依赖性的影响。肝素只抑制gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持在10和100 IU·毫升gydF4y2Ba−1gydF4y2Ba但不是在1000 IU·毫升gydF4y2Ba−1gydF4y2Ba。胶原蛋白的主要氨基酸成分,即脯氨酸,trans-hydroxylproline和甘氨酸,并没有影响gydF4y2Ba铜绿假单胞菌gydF4y2Ba依从性显著。同样,糖,包括半乳糖,gydF4y2BaNgydF4y2Ba-acetylneuraminic酸和gydF4y2BaNgydF4y2Ba乙酰氨基葡萄糖,并没有改变gydF4y2Ba铜绿假单胞菌gydF4y2Ba依从性显著。gydF4y2Ba
脱氧核糖核酸指纹技术表明,大多数CF患者港口基因相关gydF4y2Ba铜绿假单胞菌gydF4y2Ba病毒在呼吸道长期的时间gydF4y2Ba12gydF4y2Ba。然而,鲜为人知的机制(s)gydF4y2Ba铜绿假单胞菌gydF4y2Ba持久性bronchiectatic气道。优惠的依从性gydF4y2Ba铜绿假单胞菌gydF4y2Ba受损组织也在很大程度上是无法解释的,尽管损伤气道上皮细胞表达asialo-GgydF4y2BaM1gydF4y2Ba寡糖,这可能是一个gydF4y2Ba铜绿假单胞菌gydF4y2Ba受体gydF4y2Ba13gydF4y2Ba。它是可能的gydF4y2Ba铜绿假单胞菌gydF4y2Ba杆菌逃避黏膜纤毛的清除,坚持基底膜完整的纤毛上皮粘膜网站剥蚀。gydF4y2Ba铜绿假单胞菌gydF4y2Ba外毒素,如绿脓菌素,1-hydroxylphenazine和鼠李糖脂,也可以暴露的基底膜gydF4y2Ba铜绿假单胞菌gydF4y2Ba细菌通过纤毛跳动减缓,分离上皮紧密连接,和呼吸道粘膜受损脱落gydF4y2Ba7gydF4y2Ba,gydF4y2Ba9gydF4y2Ba。尽可能多的管腔内的细菌附着呼吸道粘液,许多工人认为,这可能是一个持续的呼吸道病原体,水库等gydF4y2Ba铜绿假单胞菌gydF4y2Ba和gydF4y2Ba流感嗜血杆菌gydF4y2Ba7gydF4y2Ba,gydF4y2Ba14gydF4y2Ba。然而,呼吸道粘液最终咯血,随后不能保留这些病原体在航空公司。因此,上述假说可以更好地解释持续气道殖民化的呼吸道病原体,如gydF4y2Ba铜绿假单胞菌gydF4y2Ba和nontypablegydF4y2Ba流感嗜血杆菌gydF4y2Ba。然而,(s)的机制gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持基底膜还没有系统的研究。gydF4y2Ba
的附着力gydF4y2Ba铜绿假单胞菌gydF4y2Ba呼吸道粘膜是复杂和多gydF4y2Ba铜绿假单胞菌gydF4y2Ba丰富和上皮受体似乎参与。gydF4y2Ba铜绿假单胞菌gydF4y2Bapili高度毒株特异性蛋白质的附属物,丰富的调停坚持人类气管黏膜gydF4y2Ba15gydF4y2Ba。菌毛的表面上gydF4y2Ba铜绿假单胞菌gydF4y2Ba认识到d -gydF4y2BaNgydF4y2Baasialo-G -acetylgalactosamine-β1-4-d-galactose (GalNAcβ1-4Gal)二糖gydF4y2BaM1gydF4y2Ba和ggydF4y2Ba平方米gydF4y2Ba受体gydF4y2Ba16gydF4y2Ba。黏液状的菌株gydF4y2Ba铜绿假单胞菌gydF4y2Ba胞外多糖产生,形成一个松散的组织的线性链多糖胶囊,从细胞表面向外辐射。这已经被证明可以调解对人体呼吸道上皮细胞gydF4y2Ba9gydF4y2Ba,gydF4y2Ba17gydF4y2Ba。作者还观察到的直接并列gydF4y2Ba铜绿假单胞菌gydF4y2Ba极地pili胶原蛋白表面的许多SEM字段,尽管在很多情况下gydF4y2Ba铜绿假单胞菌gydF4y2Ba杆菌也直接附加到胶原蛋白表面。gydF4y2Ba
铜绿假单胞菌gydF4y2Ba和其他常见呼吸道病原体,如nontypablegydF4y2Ba流感嗜血杆菌gydF4y2Ba和gydF4y2Ba肺炎链球菌,gydF4y2Ba绑定到glycoconjugates糖脂和黏蛋白。具体来说,GalNAcβ1-4Gal二糖鞘糖脂中发现人类肺上皮细胞表面的受体外植体是一个候选人gydF4y2Ba18gydF4y2Ba。细胞表面的唾液酸已被确认为上皮受体的一个至关重要的组成部分gydF4y2Ba铜绿假单胞菌gydF4y2Baadhesin (s)gydF4y2Ba19gydF4y2Ba。一些其他呼吸道病原体,如gydF4y2Ba分枝杆菌肺炎gydF4y2Ba利用唾液acid-containing glycoconjugates受体gydF4y2Ba20.gydF4y2Ba。表面束缚神经氨酸酶可能参与最初的识别系统,除了唾液酸残留的去除,允许增加adhesin之间的亲和力和asialo-terminal残留的细胞表面受体gydF4y2Ba21gydF4y2Ba。数据还表明,唾液酸和可用gydF4y2BaNgydF4y2Ba乙酰氨基葡萄糖arecomponents粘蛋白受体(s)和两个1型d-galactose-β1-3-d -gydF4y2BaNgydF4y2Ba乙酰氨基葡萄糖(Galβ1-3GlcNAc)和2型d-galactose-β1-4-d -gydF4y2BaNgydF4y2Ba乙酰氨基葡萄糖(Galβ1-4GlcNAc)二糖单位参与绑定gydF4y2Ba铜绿假单胞菌gydF4y2Ba22gydF4y2Ba。最近,gydF4y2Ba铜绿假单胞菌gydF4y2Ba也被证明具有高亲和性结合位点sialyl-Lewis X共轭,一个gydF4y2BaNgydF4y2Ba-acetylneuraminic酸α2-3-d-galactose-β1-4 (d-fucoseα1-3)- d -gydF4y2BaNgydF4y2Ba乙酰氨基葡萄糖低聚糖的黏蛋白序列中常见的CF患者gydF4y2Ba23gydF4y2Ba。这表明,除了中性碳水化合物的识别因素,gydF4y2Ba铜绿假单胞菌gydF4y2Ba丰富的特定于酸性glycoconjugates产生应对当地的气道粘膜的炎症。gydF4y2Ba
基底膜主要是由iv型胶原、层粘连蛋白、纤连蛋白和硫酸乙酰肝素蛋白聚糖。他们构成上皮和内皮细胞环绕周围神经和肌肉细胞gydF4y2Ba24gydF4y2Ba。iv型胶原蛋白是最丰富的nonfibril-forming胶原蛋白在肺癌和提供了其他基底膜组件的支架附着。gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持i型胶原蛋白矩阵gydF4y2Ba25gydF4y2Ba,纤连蛋白gydF4y2Ba26gydF4y2Ba,层粘连蛋白gydF4y2Ba通过gydF4y2Banonpilus-mediated机制gydF4y2Ba27gydF4y2Ba。gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持i型,——胶原蛋白由d-galactose抑制,d-mannose和gydF4y2BaNgydF4y2Ba-acetylneuraminic酸gydF4y2Ba28gydF4y2Ba,这表明,糖类可以扮演一个角色gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持i型,——胶原蛋白。然而,的依从性gydF4y2Ba铜绿假单胞菌gydF4y2Baiv型胶原蛋白,基底膜的最丰富的框架,没有被研究过。gydF4y2Ba
凝集素PHA-E和gydF4y2Ba答:hypogeagydF4y2Ba被用来抵销的坚持gydF4y2Ba铜绿假单胞菌gydF4y2Baiv型胶原在目前的模型。PHA-E具体结合Galβ1-4GlcNAc Manα1-6有关的手臂gydF4y2Ba复杂类型的gydF4y2BaNgydF4y2Ba聚糖gydF4y2Ba11gydF4y2Ba,并有可能与竞争gydF4y2Ba铜绿假单胞菌gydF4y2Baadhesin识别受体轴承这二糖单位gydF4y2Ba22gydF4y2Ba。iv型胶原蛋白的7 s域熊位点asparagine-linked聚糖的bi -与终端β1-4-d-galactose-d triantennary类型gydF4y2BaNgydF4y2Ba-acetylgalactosamine (Galβ1-4GalNAc)gydF4y2Ba29日gydF4y2Ba和当前结果表明PHA-E减少gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持胶原蛋白,gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持iv型胶原蛋白可能涉及Galβ1-4GalNAc序列。gydF4y2Ba答:hypogeagydF4y2Ba凝集素结合专门β1-3-d-galactose-d -gydF4y2BaNgydF4y2Ba-acetylgalactosamine (Galβ1-3GalNAc)终端gangliotetrosylceramide序列。绑定的gydF4y2Ba铜绿假单胞菌gydF4y2Ba后者表明这个醣脂类可能是一个上皮受体gydF4y2Ba铜绿假单胞菌gydF4y2Ba18gydF4y2Ba。然而,缺乏的影响gydF4y2Ba答:hypogeagydF4y2Ba在gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持在这个研究表明终端Galβ1-3GalNAc序列没有参与gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持iv型胶原蛋白。缺乏影响依从性的糖,包括半乳糖,gydF4y2BaNgydF4y2Ba-acetylneuraminic酸,gydF4y2BaNgydF4y2Ba乙酰氨基葡萄糖(表4gydF4y2Ba⇑gydF4y2Ba),表明三维结构的这些更重要的行列式二糖或集群adhesin识别比个人糖,和与凝集素研究的结果是一致的。gydF4y2Ba
肝素是一种粘多糖类似硫酸乙酰肝素的二糖重复d-glucuronic acid-d -gydF4y2BaNgydF4y2Ba乙酰氨基葡萄糖,但不同于二糖的广泛领域重复与N -取代和O-sulphates。肝素可能的行为gydF4y2Ba通过gydF4y2Ba竞争与硫酸乙酰肝素蛋白多糖的半个出现在组织,抑制坚持ofurinary病原体膀胱粘膜gydF4y2Ba30.gydF4y2Ba。现在结果表明,肝素显著抑制gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持10 - 100 IU·毫升gydF4y2Ba−1gydF4y2Ba。硫酸乙酰肝素可能出现的蛋白聚糖的大型分子聚合组件iv型胶原蛋白制备,因此参与gydF4y2Ba铜绿假单胞菌gydF4y2Ba依从性。这个有趣的现象应该进一步评估,因为这应该实现低浓度的肝素nebulisation的航空公司没有系统性的低剂量肝素溶液的效果。更高层次的肝素,即1000 IU·毫升gydF4y2Ba−1gydF4y2Ba,也降低gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持密度与控制相比,尽管这种差异无统计学意义(表2gydF4y2Ba⇑gydF4y2Ba)。缺乏响应存在剂量依赖的相关性是令人费解的,无法解释当前的理解antiadherence肝素的影响。进一步研究areclearly保证这些可以提供见解gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持设计实验小说的治疗机制和线索gydF4y2Ba铜绿假单胞菌gydF4y2Ba感染使用肝素。gydF4y2Ba
马库斯gydF4y2Ba等gydF4y2Ba。gydF4y2Ba31日gydF4y2Ba表明supraphysiological浓度的CagydF4y2Ba2 +gydF4y2Ba(15毫米)增强gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持仓鼠气管上皮细胞,表明金属离子参与adhesin-oligosaccharide绑定。相比之下,目前的作者发现,生理浓度的CagydF4y2Ba2 +gydF4y2Ba抑制gydF4y2Ba铜绿假单胞菌gydF4y2Ba依从性(表2gydF4y2Ba⇑gydF4y2Ba)。针对硫酸乙酰肝素蛋白聚糖的可能性与iv型胶原,CagydF4y2Ba2 +gydF4y2Ba结合硫酸乙酰肝素可以改变了半个可用硫酸乙酰肝素gydF4y2Ba铜绿假单胞菌gydF4y2Ba依从性。胶原蛋白的主要成分氨基酸成分,即脯氨酸,trans-hydroxyproline和甘氨酸,没有影响gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持iv型胶原蛋白(表3所示gydF4y2Ba⇑gydF4y2Ba)。这表明,这些氨基酸并不直接参与之间的粘附过程gydF4y2Ba铜绿假单胞菌gydF4y2Ba和iv型胶原蛋白。这与上面的结果一致,坚持过程更有可能涉及Galβ1-4GlcNAc但不是氨基酸。他们也很有可能,这些组件骨骼中的胶原蛋白氨基酸都是锁着的,而不是直接暴露的依从性的过程。gydF4y2Ba
这项研究的结果表明gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持iv型胶原蛋白可能的行为gydF4y2Ba通过gydF4y2Ba具体机制(s)涉及adhesin d-galactose-β1-4-d的认可gydF4y2BaNgydF4y2Ba乙酰氨基葡萄糖序列。此外,肝素和CagydF4y2Ba2 +gydF4y2Ba似乎也抑制gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持与基底膜相关蛋白多糖成分iv型胶原蛋白。是持久的gydF4y2Ba铜绿假单胞菌gydF4y2Ba殖民囊肿性纤维化和支气管扩张患者是有害的,目前还没有有效的治疗根除,这些结果可能导致toa持续治疗的新方法gydF4y2Ba铜绿假单胞菌gydF4y2Ba感染。进一步的研究应该追求使用这个模型gydF4y2Ba铜绿假单胞菌gydF4y2Ba坚持其他基底膜组件。gydF4y2Ba
- 收到了gydF4y2Ba2002年10月23日。gydF4y2Ba
- 接受gydF4y2Ba2003年1月23日。gydF4y2Ba
- ©人期刊有限公司gydF4y2Ba
引用gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba









