





摘要gydF4y2Ba
已知来自嗜酸性粒细胞的活性氧(ROS)会引起过敏性炎症中的组织损伤。CC趋化因子,特别是eotaxin,并受激活、正常T细胞表达和分泌(RANTES)调节,不仅参与趋化,也参与嗜酸性粒细胞激活,如ROS的产生。已经证明,过敏患者的嗜酸性粒细胞在功能上与正常人的不一样。在本研究中,比较了过敏患者和正常对照组嗜酸性粒细胞中化学因子启动的ROS产生的特点。gydF4y2Ba
用趋化因子预处理后,用钙离子载体A23187刺激嗜酸性粒细胞。用发光依赖性化学发光法测定嗜酸性粒细胞产生的ROS。gydF4y2Ba
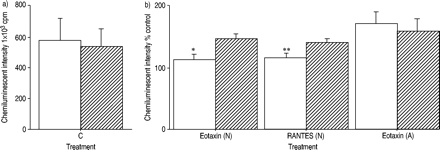
RANTES和eotaxin对钙离子诱导的嗜酸性粒细胞产生ROS具有启动效应。尽管CC趋化因子受体3的表达没有差异,但RANTES和eotaxin在患者嗜酸性粒细胞中的启动作用显著增强。白介素- 5进一步增强了正常受试者嗜酸性粒细胞中趋化因子的启动效应,而非过敏受试者。gydF4y2Ba
目前的结果表明,过敏患者的嗜酸性粒细胞对趋化因子的反应上调,并且白介素‐5可以诱导类似的表型gydF4y2Ba体内gydF4y2Ba在过敏的病人。gydF4y2Ba
- CC趋化因子受体3gydF4y2Ba
- 嗜酸性粒细胞gydF4y2Ba
- eotaxingydF4y2Ba
- 白介素5应承担的gydF4y2Ba
- 活性氧gydF4y2Ba
- 在激活规范gydF4y2Ba
- 正常T细胞表达和分泌gydF4y2Ba
这项研究得到了教育科学省和卫生福利省(日本东京)科学研究资助计划的支持。gydF4y2Ba
过敏性疾病的一个特征是组织炎症,涉及T淋巴细胞和嗜酸性粒细胞的激活gydF4y2Ba1gydF4y2Ba.变态反应性疾病的严重程度受嗜酸性粒细胞活化程度的影响。在过敏性炎症过程中,嗜酸性粒细胞进入组织,释放有毒颗粒蛋白和活性氧(ROS),导致组织损伤gydF4y2Ba2gydF4y2Ba.gydF4y2Ba
ROS的产生是由多种刺激引起的,如免疫球蛋白(Igs)和细胞因子gydF4y2Ba3.gydF4y2Ba.此前已有报道,粘附分子的信号在嗜酸性粒细胞产生ROS中起关键作用gydF4y2Ba4gydF4y2Ba.ccc趋化因子,特别是eotaxin和激活调控,正常T细胞表达和分泌(RANTES),对嗜酸性粒细胞具有选择性趋化活性。除了趋化作用外,这些趋化因子还参与嗜酸性粒细胞的活化。事实上,最近的研究表明,趋化因子启动了嗜酸性粒细胞产生ROSgydF4y2Ba5gydF4y2Ba,gydF4y2Ba6gydF4y2Ba.gydF4y2Ba
研究表明,过敏患者的嗜酸性粒细胞在效应功能上与正常受试者不相同gydF4y2Ba7gydF4y2Ba- - - - - -gydF4y2Ba9gydF4y2Ba.然而,嗜酸性粒细胞对趋化因子的不同响应尚未完全阐明。因此,在本文中,对比较研究含有含有患者的患者和正常主题,了解嗜酸性粒细胞的趋化因子对ROS产生的初探。gydF4y2Ba
材料和方法gydF4y2Ba
主题gydF4y2Ba
抽取12例健康非过敏成人静脉血(年龄18-40岁,平均27.3岁;15例呼吸道变应性疾病患者(23-32岁,平均25.1岁;五个女性)。年龄和性别分布在正常受试者和患者组之间没有显著差异。所有受试者均给予知情同意,研究根据《赫尔辛基宣言》的原则进行。采血前均未接受药物治疗≥24小时或类固醇治疗≥2周。正常受试者的定义是基于缺乏过敏或其他类似疾病的临床史。所有患者均有过敏性哮喘和/或过敏性鼻炎,IgE浓度为>400国际单位(IU)·mLgydF4y2Ba−1gydF4y2Ba以及针对至少一种常见空气传播过敏原(如室内尘螨、花粉或真菌)的IgE放射变应原测试结果高于3级。哮喘和变应性鼻炎患者分别为7人和11人(3人同时患有哮喘和变应性鼻炎)。参与本研究的哮喘患者符合美国胸科协会对哮喘的定义。所有的过敏性鼻炎患者在采血时都有症状(鼻塞、打喷嚏、流鼻涕、眼睛发痒、gydF4y2Ba等。gydF4y2Ba).患者外周血嗜酸性粒细胞计数明显高于正常对照组(683.3±374.0)gydF4y2Ba与gydF4y2Ba126.0±80.4细胞·毫米gydF4y2Ba−3gydF4y2Bap<0.01)。gydF4y2Ba
嗜酸性粒细胞隔离gydF4y2Ba
如前所述,使用改性的CD16负选择方法从肝素化静脉血中分离出嗜酸性粒细胞。gydF4y2Ba10gydF4y2Ba.简而言之,从泛白涂层中获得的细胞与anti - d16、anti - d3、anti - d20和anti - d14单克隆抗体(小鼠IgG;随后与抗鼠IgG磁珠反应(Dynal, Oslo, Norway)。CD16、CD3、CD20和cd14阴性嗜酸性粒细胞使用磁性细胞分选系统(Miltenyi Biotec, Bergisch Gladbach, Germany)获得。嗜酸性粒细胞纯度为>97%。gydF4y2Ba
Luminoldependent化学发光gydF4y2Ba
通过发光依赖性化学发光的方法检测了嗜酸性粒细胞产生的ROSgydF4y2Ba5gydF4y2Ba.此前,eotaxin和RANTES在培养15min后对嗜酸性粒细胞氧化代谢有明显影响gydF4y2Ba5gydF4y2Ba,gydF4y2Ba6gydF4y2Ba.因此纯化的嗜酸性粒细胞(1×10gydF4y2Ba6gydF4y2Ba细胞·ml.gydF4y2Ba−1gydF4y2Ba)悬浮在Roswell Park Memorial Institute (RPMI) 1640培养基中,与1 - 100 nM eotaxin (R & D Systems, Minneapolis, MN, USA)或RANTES (Sigma, St Louis, MO, USA)在96孔平底板中孵育,存在或不存在1 ng·mLgydF4y2Ba−1gydF4y2Ba白细胞介素(IL)‐5或粒细胞巨噬细胞集落刺激因子(GMCSF) (R & D系统)在37℃下处理15分钟。在一些实验中,嗜酸性粒细胞用抗il‐5受体α (IL5Rα)抗体(小鼠IgG1κ;pharmacingen, San Diego, CA, USA)或同型匹配对照(Pharmingen),均为0.2µg·mLgydF4y2Ba−1gydF4y2Ba或CC趋化因子受体(CCR) 3拮抗剂(化合物X;这是日本筑波坂友制药有限公司的礼物),在37°C下加热30分钟。添加50µL钙离子载体A23187 (Sigma;最终浓度1×10gydF4y2Ba−5gydF4y2BaM)至100µL嗜酸性粒细胞悬液(5×10gydF4y2Ba4gydF4y2Ba电池),含0.25 mM鲁米诺(双叶医学,东京,日本)。使用ARGUS50/2D光度计(Hamamatsu Photonics, Hamamatsu, Japan)测定60分钟的最大强度和积分强度化学发光。gydF4y2Ba
嗜酸性粒细胞表面CC趋化因子受体3的流式细胞术分析gydF4y2Ba
纯化的嗜酸性粒细胞(<1×10gydF4y2Ba6gydF4y2Ba用荧光素异硫氰酸酯(FITC)标记的抗人CCR3单克隆抗体(小鼠IgG2;DAKO、斯特鲁普、丹麦;0.5µg·毫升gydF4y2Ba−1gydF4y2Ba)30年 37°C下的最低温度。一种FITConjugated IgG2同型匹配对照单克隆抗体(美国加利福尼亚州圣何塞市贝克顿迪金森;0.5 微克·毫升gydF4y2Ba−1gydF4y2Ba)来评估非特异性程度。洗涤细胞后,使用FACScan流式细胞仪(BecktonDickinson)分析染色细胞。gydF4y2Ba
测定细胞内钙浓度gydF4y2Ba
从正常受试者纯化的嗜酸性粒细胞悬浮在含钙的汉克平衡盐溶液(HBSS)中gydF4y2Ba2+gydF4y2Ba(0.14 g·毫升gydF4y2Ba−1gydF4y2BaCaClgydF4y2Ba2gydF4y2Ba)、镁gydF4y2Ba2+gydF4y2Ba(0.1g·mlgydF4y2Ba−1gydF4y2BaMgClgydF4y2Ba2gydF4y2Bah·6gydF4y2Ba2gydF4y2BaO;0.1 g·毫升gydF4y2Ba−1gydF4y2BaMgSOgydF4y2Ba4gydF4y2Bah·7gydF4y2Ba2gydF4y2BaO) 和2%胎牛血清(Sigma),细胞密度为2×10gydF4y2Ba6gydF4y2Ba细胞·ml.gydF4y2Ba−1gydF4y2Ba.Fura‐2‐乙酰氧基甲酯(DOJINDO,熊本,日本)以2 μ M的最终浓度加入。孵育40 min后,270×离心5 min去除多余染料gydF4y2BaggydF4y2Ba在4°C下,将细胞重新悬浮在含有20 mM-羟乙基哌嗪乙烷磺酸(HEPES)(pH 7.4)在2×10的浓度下gydF4y2Ba6gydF4y2Ba细胞·ml.gydF4y2Ba−1gydF4y2Ba.在340和380 nm的荧光光谱仪(ARGUS;滨松光子学)。gydF4y2Ba
统计分析gydF4y2Ba
使用配对和未配对的T检验分析数据,方差分析(ANOVA)或Mannwhitney U-Test。认为p值为≤0.05表示意义。gydF4y2Ba
结果gydF4y2Ba
正常和过敏受试者嗜酸性粒细胞的发光依赖性化学发光gydF4y2Ba
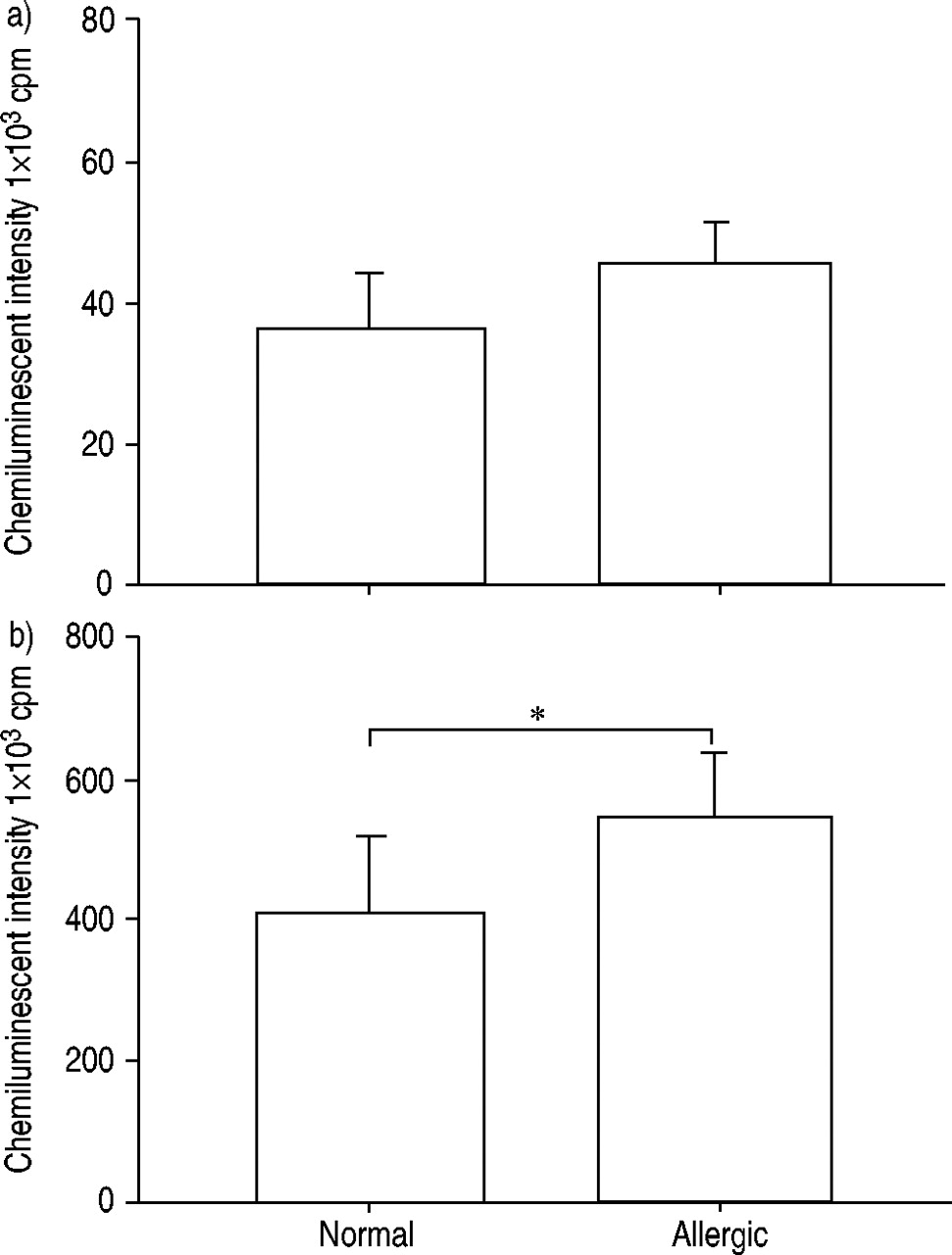
通过钙离子载体A23187诱发的发光依赖性化学发光检测嗜酸性粒细胞产生ROS,并比较正常受试者和过敏患者。测定嗜酸性粒细胞产生ROS的情况gydF4y2Ba通过gydF4y2Ba过敏性患者的整体强度比正常受试者在显着更大(图1gydF4y2Ba⇓gydF4y2Ba)(P <0.05)。gydF4y2Ba
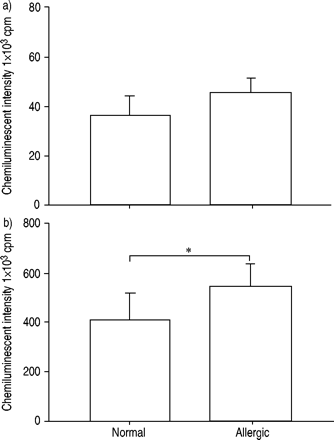
正常受试者(n=12)和过敏受试者(n=15)嗜酸性粒细胞产生的Reactive oxygen species通过a)最大和b)积分强度发光依赖性化学发光60分钟来确定。添加50μL钙离子载体A23187(终浓度1×10)刺激嗜酸性粒细胞gydF4y2Ba−5gydF4y2BaM)至100 μL嗜酸性粒细胞悬液(5×10gydF4y2Ba4gydF4y2Ba电池)含有0.25 mM鲁米诺。数据以平均值±表示gydF4y2Ba扫描电镜gydF4y2Ba.Cpm:每分钟计数。*:P <0.05gydF4y2Ba与gydF4y2Ba正常受试者(未配对t‐测试)。gydF4y2Ba
eotaxin和regulated对正常和过敏受试者嗜酸性粒细胞产生活性氧的活化、正常T细胞表达和分泌的影响gydF4y2Ba
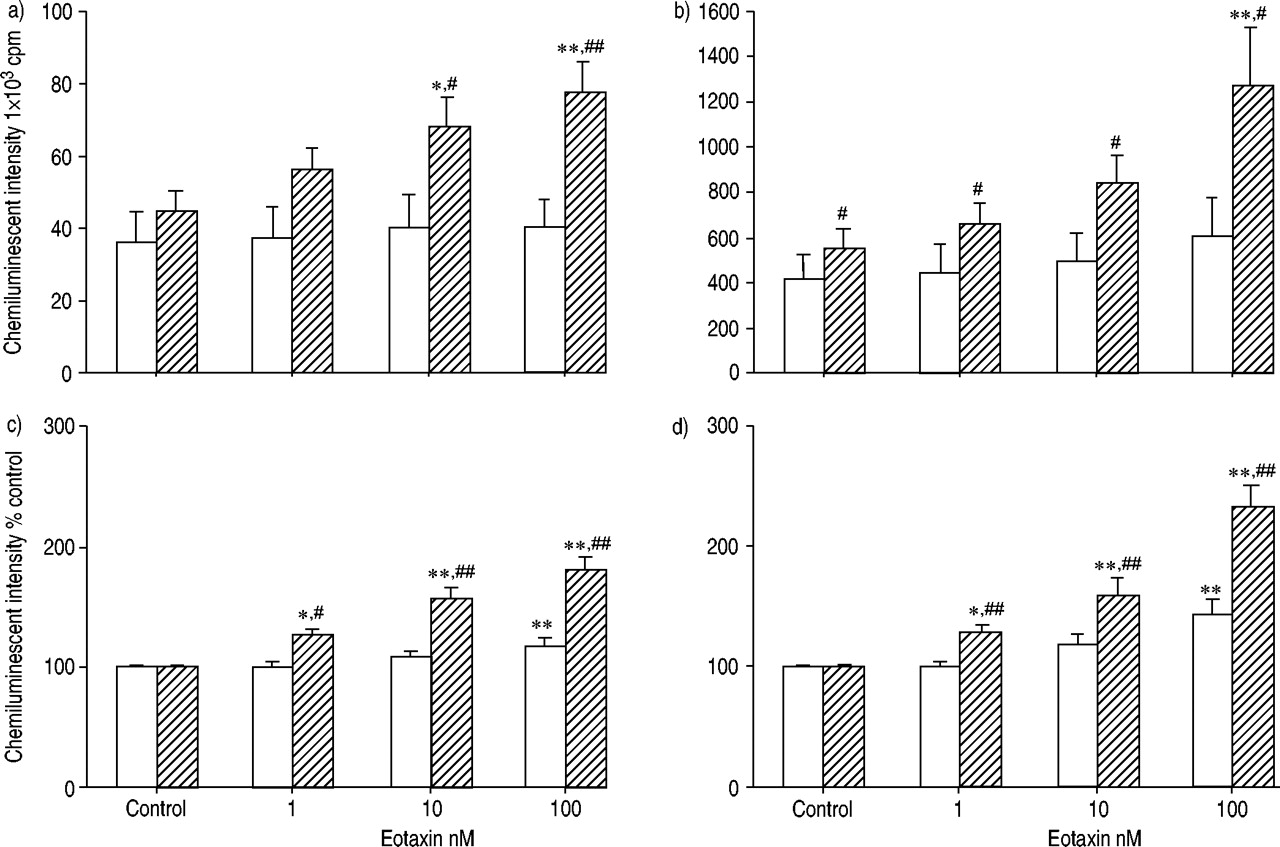
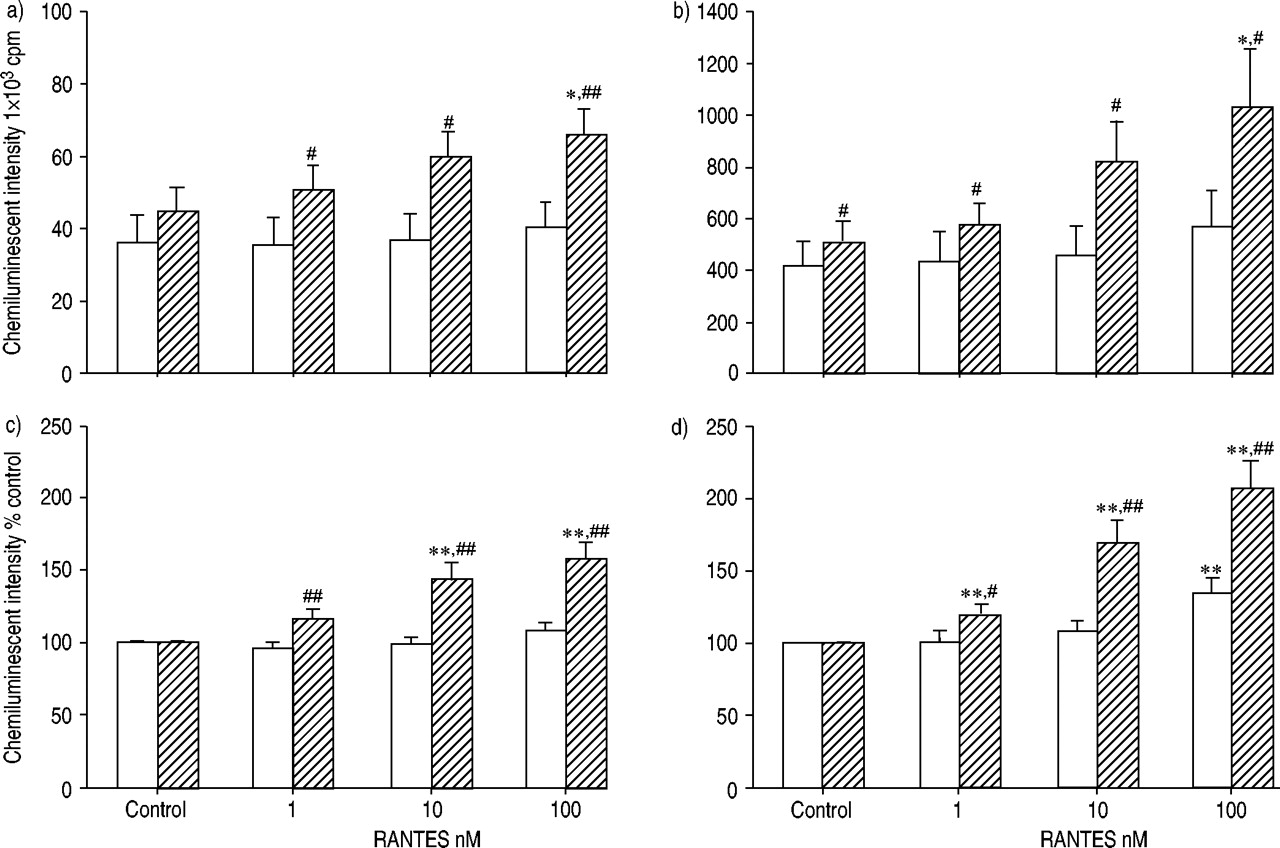
比较了eotaxin和RANTES对正常受试者和过敏患者ROS产生的启动效应。图2a及bgydF4y2Ba⇓gydF4y2Ba显示最大强度和积分强度。在过敏患者中,嗜酸性粒细胞与eotaxin的预孵育明显增强了ROS的生成,而在正常受试者中则没有。在eotaxin启动条件下,过敏和正常受试者之间ROS产生的差异要大得多。为了研究eotaxin的增强作用,结果也与那些没有趋化因子的结果相关(图2c和d)gydF4y2Ba⇓gydF4y2Ba).eotaxin对过敏患者的嗜酸性粒细胞的增强作用强于正常人。也观察到RANTES的类似效果(图3)gydF4y2Ba⇓gydF4y2Ba).gydF4y2Ba
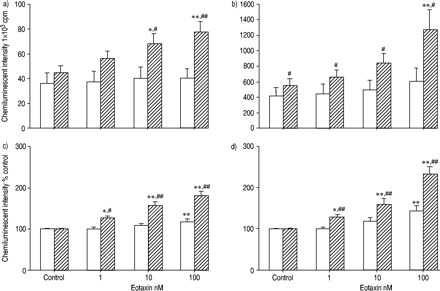
由嗜酸性粒细胞(1-100nm)从正常(□; n = 12)和过敏(└; n = 15)受试者通过最大(a,c)和积分(b)(b)来产生反应性氧气种类(ros)生产。,d)强度乳液依赖性化学发光60分钟。通过添加50µL钙离子载体A23187(最终浓度1×10)来刺激嗜酸性粒细胞gydF4y2Ba−5gydF4y2BaM)至100µL嗜酸性粒细胞悬液(5×10gydF4y2Ba4gydF4y2Ba电池)含有0.25 mM鲁米诺。数据以平均值±表示gydF4y2Ba扫描电镜gydF4y2Ba.嗜酸性粒细胞与eotaxin的预孵育可增强过敏患者和正常受试者的ROS生成。eotaxin的启动效应在过敏受试者的嗜酸性粒细胞中比在正常受试者中的更强。Cpm:每分钟计数。*,gydF4y2Ba* *: p < 0.05, p < 0.01gydF4y2Ba与gydF4y2Ba控制(方差分析);gydF4y2Ba#gydF4y2Ba,gydF4y2Ba##gydF4y2Ba:p<0.05,p<0.01gydF4y2Ba与gydF4y2Ba正常受试者(未配对t‐测试)。gydF4y2Ba
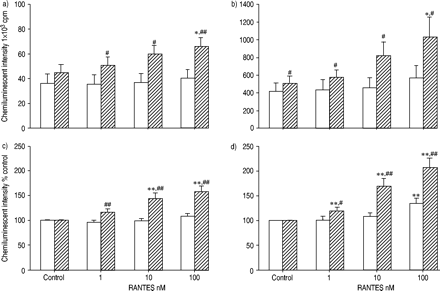
经活化调节、正常T细胞表达和分泌(RANTES;1-100)处理的嗜酸性粒细胞产生活性氧(ROS) 纳米)从正常(□; n=12)和过敏性(└; n=15)受试者,根据60天的最大(a,c)和积分(b,d)强度发光依赖性化学发光测定 通过添加50%嗜酸性粒细胞进行最小嗜酸性粒细胞刺激 µL钙离子载体A23187(最终浓度1×10gydF4y2Ba−5gydF4y2BaM)至100µL嗜酸性粒细胞悬液(5×10gydF4y2Ba4gydF4y2Ba电池)含有0.25 mM鲁米诺。数据以平均值±表示gydF4y2Ba扫描电镜gydF4y2Ba.RANTES对嗜酸性粒细胞的预培养可增强过敏患者和正常受试者的ROS生成。RANTES的启动效应在过敏受试者的嗜酸性粒细胞中比在正常受试者中更强。Cpm:每分钟计数。*,gydF4y2Ba* *: p < 0.05, p < 0.01gydF4y2Ba与gydF4y2Ba控制(方差分析);gydF4y2Ba#gydF4y2Ba,gydF4y2Ba##gydF4y2Ba:p<0.05,p<0.01gydF4y2Ba与gydF4y2Ba正常受试者(未配对t‐测试)。gydF4y2Ba
CC趋化因子受体3拮抗剂对嗜酸性粒细胞趋化因子启动的活性氧产生的影响gydF4y2Ba
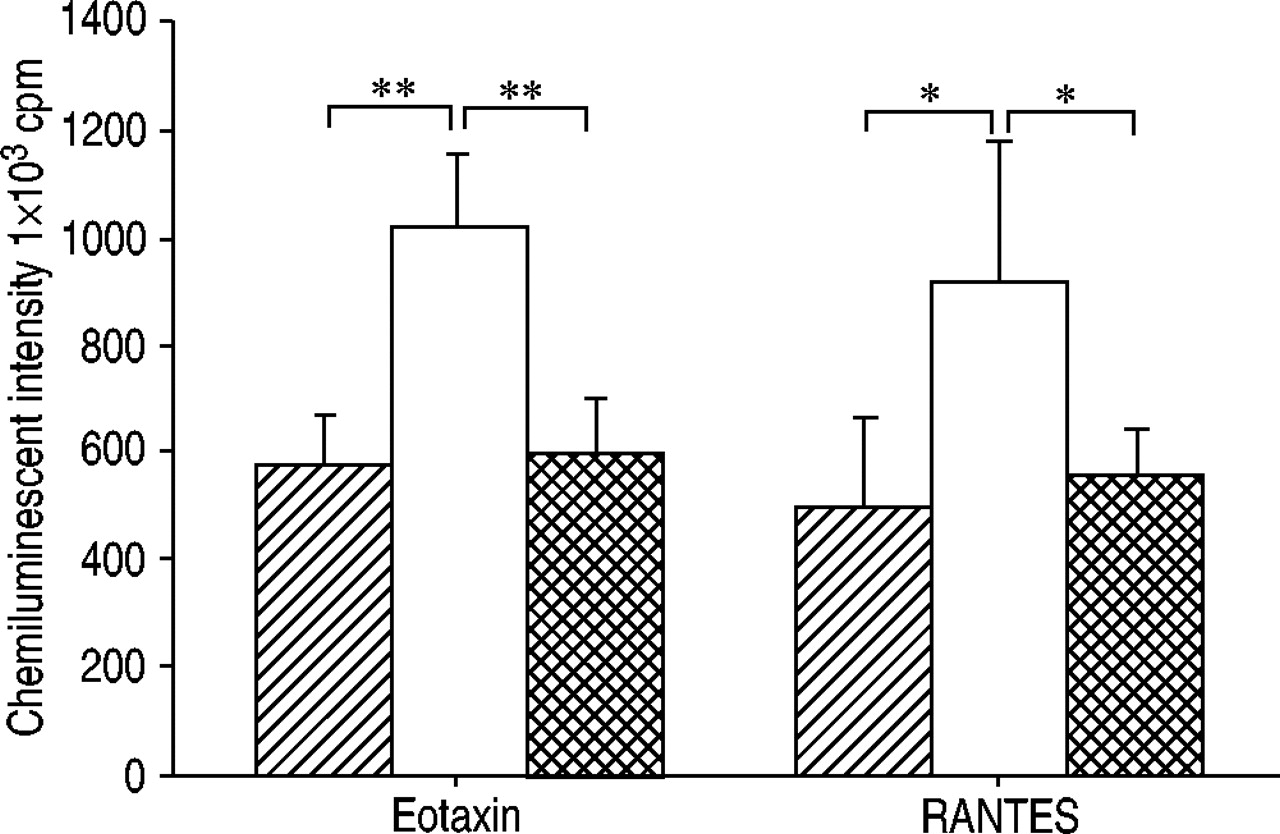
为了证实CCR3参与趋化因子启动的ROS生成,研究了一种CCR3拮抗剂抑制eotaxin与人类嗜酸性粒细胞结合的作用gydF4y2Ba11gydF4y2Ba是被投虫病的。CCR3拮抗剂完全抑制ETAXIN和RANTESPRIMED ROS生产(图4gydF4y2Ba⇓gydF4y2Ba).gydF4y2Ba
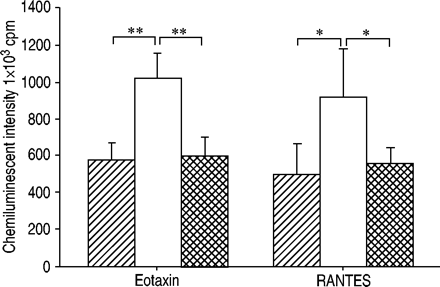
CC趋化因子受体(CCR)3拮抗剂对嗜酸性粒细胞产生的趋化因子启动的活性氧(ROS)的影响,通过积分强度发光依赖化学发光检测60分钟。□:趋化因子;┼:趋化因子+拮抗剂)。纯化嗜酸性粒细胞(1×10gydF4y2Ba6gydF4y2Ba细胞·ml.gydF4y2Ba−1gydF4y2Ba)用CCR3拮抗剂(化合物X;1×10)孵育从过敏患者(n=5)获得的药物gydF4y2Ba−6gydF4y2BaM) 30分钟。然后用eotaxin处理嗜酸性粒细胞或调节激活、正常T细胞表达和分泌(RANTES)(均为100 nM) 15分钟。通过添加50µL钙离子载体A23187(最终浓度1×10)来刺激嗜酸性粒细胞gydF4y2Ba−5gydF4y2BaM)至100µL嗜酸性粒细胞悬液(5×10gydF4y2Ba4gydF4y2Ba电池)含有0.25 mM鲁米诺。数据以平均值±表示gydF4y2Ba扫描电镜gydF4y2Ba.CCR3拮抗剂完全抑制eotaxin和RANTES的启动效应。Cpm:每分钟计数。*、**:p<0.05, p<0.01(配对t‐检验)。gydF4y2Ba
正常人和过敏者嗜酸性粒细胞中CC趋化因子受体3的表达gydF4y2Ba
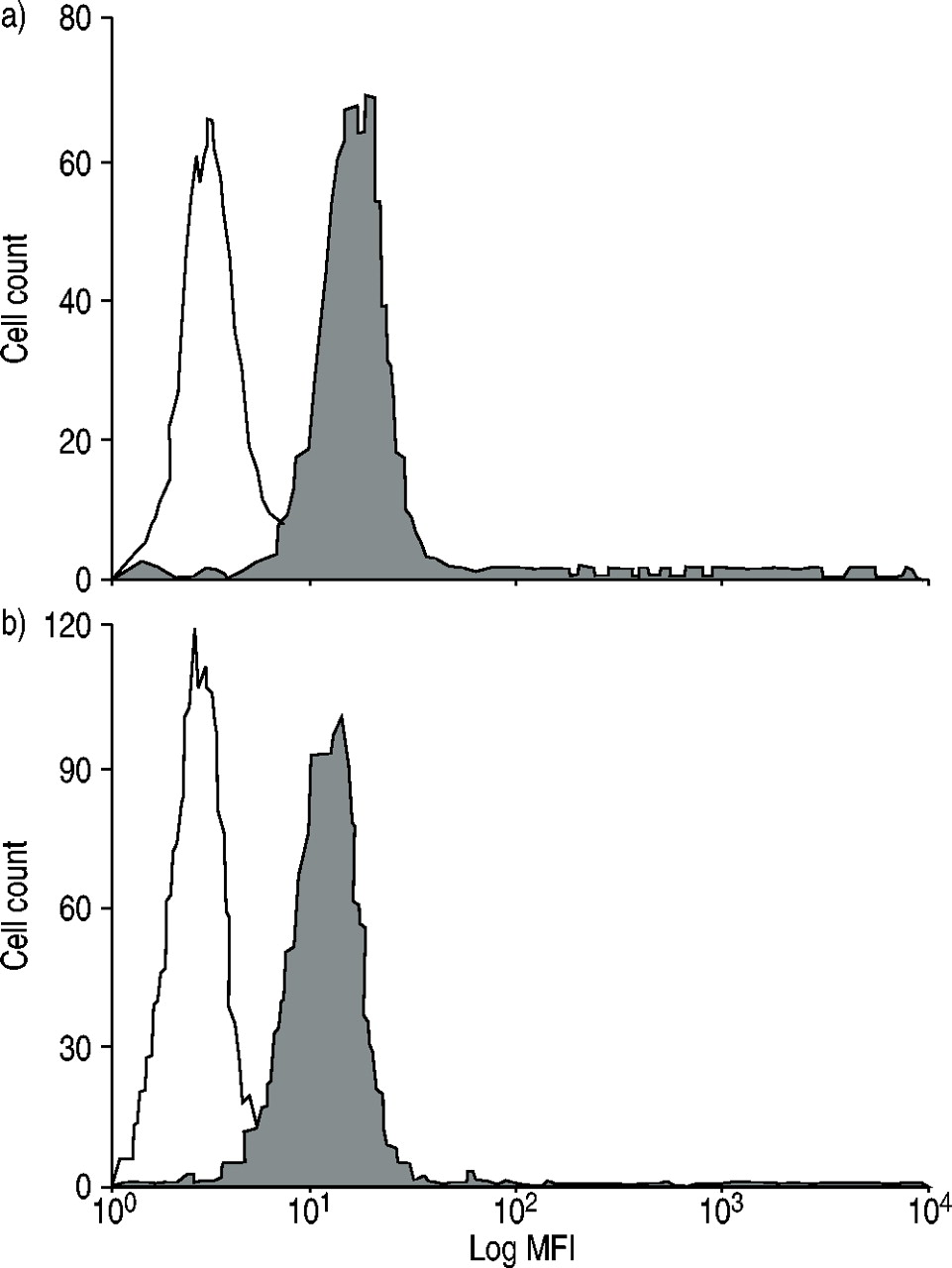
为了研究过敏患者嗜酸性粒细胞的不同反应,我们测定了RANTES和eotaxin的共同受体CCR3的表达。以ccr3阳性细胞百分比和与对照组相比的平均荧光强度作为受体表达的参数。在过敏者和正常人的嗜酸性粒细胞之间没有观察到显著差异(图5)gydF4y2Ba⇓gydF4y2Ba).gydF4y2Ba
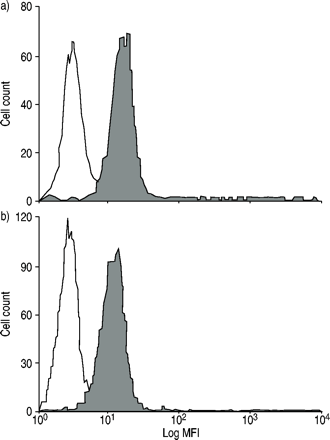
a)正常人(n=7)和b)过敏患者(n=8)(□:免疫球蛋白G2a(阴性对照);▓:CCR3)。有代表性的直方图显示。过敏组和正常组嗜酸性粒细胞表面表达无显著差异(14.5±2.2)gydF4y2Ba与gydF4y2Ba平均荧光强度(MFI)与阴性对照差异13.9±1.8;82.2±6.7gydF4y2Ba与gydF4y2Ba83.0±7.8% CCR3positive细胞)。gydF4y2Ba
白介素‐5对嗜酸性粒细胞产生的趋化因子启动的活性氧的影响gydF4y2Ba
IL‐5已被证明可以增强嗜酸性粒细胞的效应功能gydF4y2Ba12gydF4y2Ba,gydF4y2Ba13gydF4y2Ba.IL‐5增强嗜酸性粒细胞对血小板激活因子、甲酰基甲基亮氨酸苯丙氨酸、血小板因子‐4和补体因子5a的反应gydF4y2Ba13gydF4y2Ba,gydF4y2Ba14gydF4y2Ba.因此,我们检测了IL - 5在正常受试者和患者对趋化因子的不同嗜酸性粒细胞反应中的可能参与。在一项初步研究中,1‐ng·mLgydF4y2Ba−1gydF4y2BaIL-5剂量,如前一体化患者血清浓度的报告中所示gydF4y2Ba15gydF4y2Ba在没有趋化因子的情况下,对嗜酸性粒细胞产生ros没有影响(图6a)gydF4y2Ba⇓gydF4y2Ba).在正常受试者中,IL - 5的作用进一步增强了趋化因子的启动效应(图6b和c)gydF4y2Ba⇓gydF4y2Ba).有趣的是,在过敏患者的嗜酸性粒细胞中没有观察到这种效应(图6d)gydF4y2Ba⇓gydF4y2Ba).gydF4y2Ba
白介素(IL)‐5对活性氧(ROS)产生的影响正常受试者)和b)化学启动因子(正常(N)或过敏(A)受试者)通过积分强度发光依赖性化学发光60分钟测定嗜酸性粒细胞。纯化嗜酸性粒细胞(1×10gydF4y2Ba6gydF4y2Ba细胞·ml.gydF4y2Ba−1gydF4y2Ba)从正常(n=7)和过敏(n=6)受试者中获得的,在有IL‐5 (1 ng·mL)或没有IL‐5 (1 ng·mL)的情况下,用eotaxin预孵育或调节激活、正常T‐细胞表达和分泌(RANTES)(均为10 nM)gydF4y2Ba−1gydF4y2Ba).通过添加50µL钙离子载体A23187(最终浓度1×10)来刺激嗜酸性粒细胞gydF4y2Ba−5gydF4y2BaM)至100µL嗜酸性粒细胞悬液(5×10gydF4y2Ba4gydF4y2Ba电池)含有0.25 mM鲁米诺。数据以平均值±表示gydF4y2Ba扫描电镜gydF4y2Ba.在一项初步研究中,1‐ng·mLgydF4y2Ba−1gydF4y2Ba单独剂量的IL‐5并不影响嗜酸性粒细胞产生ROS (a)。在正常受试者中,IL‐5的作用进一步增强趋化因子的启动效应。在过敏患者的嗜酸性粒细胞中未观察到这种作用。Cpm:每分钟计数。*、**:p<0.05, p<0.01gydF4y2Ba与gydF4y2Ba趋化因子加IL-5(配对t检验)。gydF4y2Ba
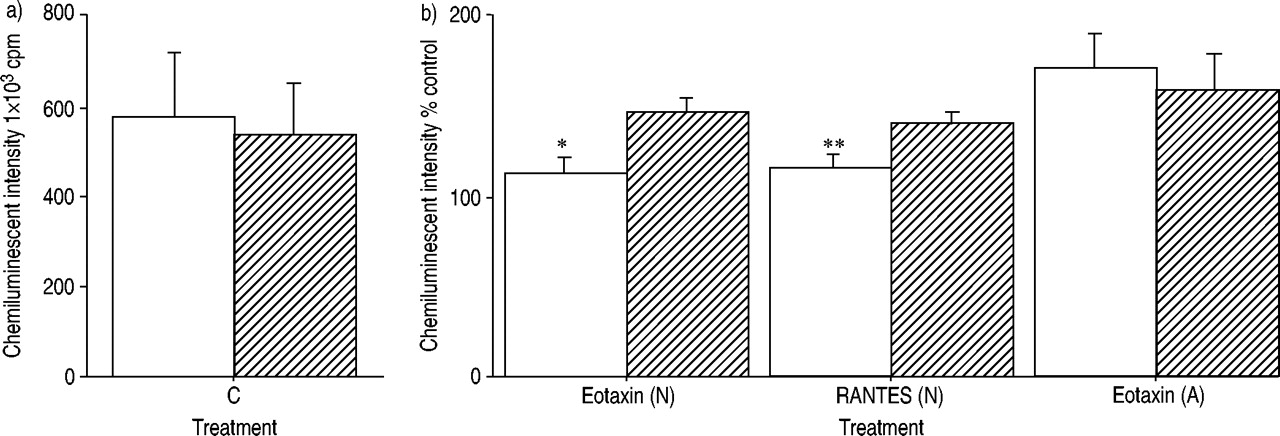
此外,GMCSF(1 ng·mlgydF4y2Ba−1gydF4y2Ba)不影响eotaxin和RANTES的启动效应(125.5±17.6)gydF4y2Ba与gydF4y2Ba119.7±9.4%控制整体化学发光强度,单独使用eotaxingydF4y2Ba与gydF4y2Baeotaxin +刺激因子)。此外,在IL - 5处理后,嗜酸性粒细胞的CCR3表达没有变化(图7)gydF4y2Ba⇓gydF4y2Ba).gydF4y2Ba
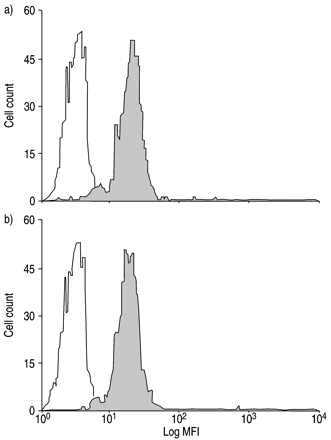
在与a)磷酸盐缓冲盐水(PBS)和b)白细胞介素(IL)-5(1)孵育后,使用针对CCR3的荧光素异硫氰酸连接单克隆抗体治疗的正常受试者(n=5)嗜酸性粒细胞上CC趋化因子受体(CCR)3表达的荧光活化细胞分选分析 ng·mLgydF4y2Ba−1gydF4y2Ba)作用30min(□:免疫球蛋白G2a(阴性对照);▓:CCR3)。有代表性的直方图显示。与PBS相比,IL - 5处理30min后嗜酸性粒细胞CCR3表达没有变化(14.8±2.5)gydF4y2Ba与gydF4y2Ba来自阴性对照的平均荧光强度(MFI)的15.4±1.7差;85.5±5.5gydF4y2Ba与gydF4y2Ba89.7±3.5% CCR3positive细胞)。gydF4y2Ba
为了研究对过敏嗜酸性粒细胞阻断IL5Rα是否能够逆转趋化因子启动的增强作用,在eotaxin刺激前,将过敏嗜酸性粒细胞与抗IL5Rα抗体预孵育。阻断IL - 5受体并不影响eotaxin在过敏性嗜酸性粒细胞中的启动效应(156.6±10.2)gydF4y2Ba与gydF4y2Ba控制整体化学发光强度159.6±18.8%,单独ETAXINgydF4y2Ba与gydF4y2BaEotaxin加抗菌5Rα;n = 4)。gydF4y2Ba
白细胞介素-5对趋化因子诱导的嗜酸性粒细胞钙内流的影响gydF4y2Ba
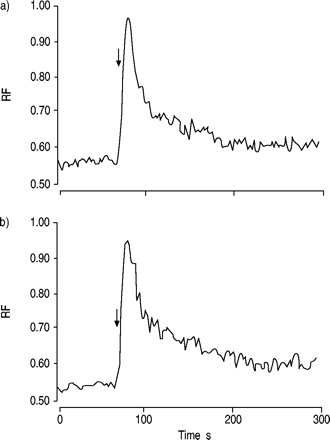
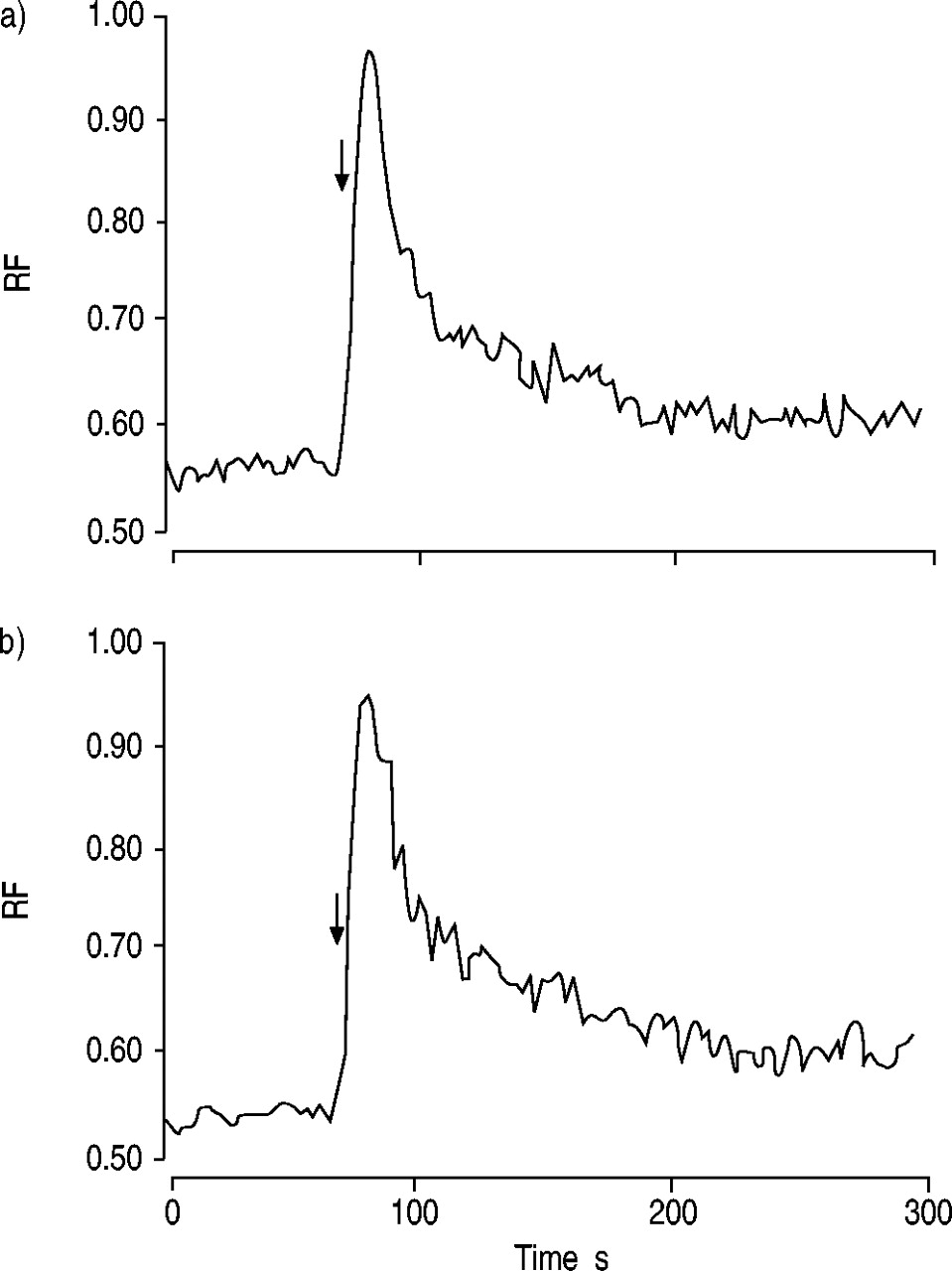
为了研究IL-5是否调节CCR3的下游信号以增强对eotaxin的反应,研究了IL-5对趋化因子诱导的钙内流的影响。然而,IL-5并不影响eotaxin诱导的钙内流(图。 8.gydF4y2Ba⇓gydF4y2Ba).gydF4y2Ba
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
白介素(IL)‐5对eotaxin诱导的钙流入嗜酸性粒细胞的影响从正常受试者获得的嗜酸性粒细胞在a)无IL‐5和b)有IL‐5 (1ng·mL)的情况下进行预培养gydF4y2Ba−1gydF4y2Ba).然后在最终浓度为100 nM(箭头)时用eotaxin刺激嗜酸性粒细胞,并按照测量细胞内钙浓度所描述的方法测量钙内流gydF4y2Ba部分。所显示的数据代表了来自不同捐赠者的三个独立分析,每个分析都显示了相似的结果。IL - 5预孵育不影响eotaxin诱导的钙内流。射频:相对荧光。gydF4y2Ba
讨论gydF4y2Ba
一些研究报道,嗜酸性粒细胞功能高度依赖于变态反应性疾病的病理生理条件gydF4y2Ba7gydF4y2Ba,gydF4y2Ba16gydF4y2Ba- - - - - -gydF4y2Ba19gydF4y2Ba.目前的研究表明,与正常受试者相比,过敏患者获得的嗜酸性粒细胞中氧化代谢上调。在过敏患者中也观察到嗜酸性粒细胞产生ROS的类似增加gydF4y2Ba16gydF4y2Ba,gydF4y2Ba17gydF4y2Ba。研究还表明,接受过敏原激发的受试者或有此类症状的患者的嗜酸性粒细胞表现出活性氧生成增强gydF4y2Ba18gydF4y2Ba,gydF4y2Ba19gydF4y2Ba.将这些研究的结果和目前的观察结果结合起来,过敏患者的嗜酸性粒细胞可能在渗入组织之前就已经在外周血中被激活了。gydF4y2Ba
此外,在本研究中,观察到过敏患者嗜酸性粒细胞对ROS产生中趋化因子反应的功能性上调。RANTES和eotaxin对ROS产生的启动效应均显著大于正常受试者嗜酸性粒细胞。即使在嗜酸性粒细胞的次优剂量下在正常受试者中,过敏患者的嗜酸性粒细胞在用趋化因子治疗后表现出增强的ROS生成。这些结果表明过敏患者的嗜酸性粒细胞对趋化因子更敏感和更敏感。gydF4y2Ba
据报道,eotaxin和IL-5在过敏性炎症期间协同调节嗜酸性粒细胞的转运gydF4y2Ba20gydF4y2Ba,gydF4y2Ba21gydF4y2Ba.年代gydF4y2BachweizergydF4y2Baet al。gydF4y2Ba22gydF4y2Ba据报道,趋化因子诱导的反应对灌注细胞因子(如IL-5)非常敏感。因此,为了延长对这些上调的敏感性的理解及其对趋化因子的反应,研究了细胞因子,例如IL-5和GMCSF的可能涉及趋化因子的引发效果。表明低浓度(1ng·mlgydF4y2Ba−1gydF4y2Ba)增强嗜酸性粒细胞介导的趋化因子启动的ROS生成,提示IL - 5可能增强对趋化因子的响应。尽管在嗜酸性粒细胞的其他功能,如脱颗粒和迁移中也观察到类似的趋势gydF4y2Ba14gydF4y2Ba,gydF4y2Ba22gydF4y2Ba,gydF4y2Ba23gydF4y2Ba,这是第一个关于IL - 5对嗜酸性粒细胞中趋化因子启动的ROS产生的启动效应的报道。有趣的是,尽管β亚基(βc)在IL - 5和GMCSF受体中都是共同的,但GMCSF并没有显示出增强作用。虽然βc在IL‐5信号通路中起主要作用gydF4y2Ba24gydF4y2Ba最近的证据表明,特异性的IL‐5受体IL‐5Rα也参与信号转导。GgydF4y2BaeijsengydF4y2Baet al。gydF4y2Ba25gydF4y2Ba克隆了一个il5r α相关分子syntenin,该分子是激活转录因子Sox4所必需的。IL5Rα还与一种新的信号分子IL‐5受体相互作用蛋白相关,它可以激活嗜酸性粒细胞中的Lyn和HckgydF4y2Ba26gydF4y2Ba.因此,这些il5r α特异性分子可能是IL - 5特异性反应的原因。gydF4y2Ba
据报道,IL‐5是由嗜酸性粒细胞本身产生的,特别是在过敏条件下gydF4y2Ba27gydF4y2Ba.一种可能是,过敏嗜酸性粒细胞可以通过自身产生的IL - 5进行启动。然而,研究表明,在过敏嗜酸性粒细胞上阻断IL - 5受体并不能逆转趋化因子的启动效应。这一结果表明,在过敏嗜酸性粒细胞中观察到的趋化因子反应上调不是由嗜酸性粒细胞分离后产生的IL - 5引起的。此外,IL - 5的增强作用仅在正常受试者的嗜酸性粒细胞中观察到,而在过敏患者的嗜酸性粒细胞中没有观察到。这一独特的表型与数据显示一致gydF4y2Ba体内gydF4y2Ba过敏性疾病患者外周血嗜酸性粒细胞粘附相关反应的启动gydF4y2Ba13gydF4y2Ba,gydF4y2Ba14gydF4y2Ba.因此,过敏患者的嗜酸性粒细胞可能会在血液中暴露IL - 5,导致对趋化因子的反应性大大增强,正如本研究所证明的那样。gydF4y2Ba
检查CCR3表达作为解释对趋化因子的不同反应的方法的可能性,但在患者与正常受试者之间的CCR3表达中没有发现显着差异。此外,用IL-5处理后,嗜酸性粒细胞的CCR3表达并未改变。这些观察结果表明,通过CCR3在过敏患者中的响应功能上调并不依赖于CCR3表达的增加。关于嗜酸性粒细胞的信号,检查IL-5是否调节趋化因子诱导的钙动员。然而,IL-5不影响eotaxin诱导的细胞内钙流量。最近据报道,与正常受试者相比,磷脂酰肌醇3-激酶的基线活性在过敏患者中升高,与IL-5在磷脂酰肌醇3-激酶活化中的累积gydF4y2Ba28gydF4y2Ba,gydF4y2Ba29gydF4y2Ba.因此,可以假设IL - 5调节CCR3的下游信号通路以增强对eotaxin的应答。在过敏患者嗜酸性粒细胞对趋化因子的上调反应中,除了细胞因子(如IL‐5)的参与外,可以推测还有其他机制,如CCR3多态性gydF4y2Ba30gydF4y2Ba最近,CCR3已成为哮喘、特应性皮炎和过敏性鼻炎等过敏性疾病的治疗靶点。事实上,已发现CCR3拮抗剂对趋化因子介导的嗜酸性粒细胞功能的抑制作用(正在编写的手稿)。gydF4y2Ba
总之,本研究表明过敏患者嗜酸性粒细胞活性氧产生中对趋化因子的反应增强,白细胞介素-5可能参与了这种增强,CC趋化因子受体3的表达没有变化。需要进一步研究来阐明其机制CC趋化因子受体3的不同反应的ms。gydF4y2Ba
致谢gydF4y2Ba
作者要感谢Y. Kamada, T. Takahashi和H. Oyamada的帮助和支持。gydF4y2Ba
- 收到了gydF4y2Ba2002年4月4日。gydF4y2Ba
- 接受gydF4y2Ba2003年1月14日。gydF4y2Ba
- ©ERS期刊有限公司gydF4y2Ba
参考文献gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba









