





文摘gydF4y2Ba
同时治疗人类气管平滑肌(对比)细胞lysophosphatidic酸(LPA)和表皮生长因子(EGF)有丝分裂发生的导致惊人的协同刺激。本研究的目的是探索潜在的信号集成中介合作网站,专注于细胞外signal-regulated激酶(ERK)和转录因子参与增殖和炎症可能的候选人。gydF4y2Ba
激活ERK被免疫印迹分析。转录因子激活使用对比评估细胞转导与荧光素酶报告基因结构。gydF4y2Ba
LPA和表皮生长因子激活ERK但没有协同效应的总和。LPA和表皮生长因子激活激活蛋白(美联社)高1环磷酸腺苷反应元件结合蛋白,核转录因子激活T细胞和血清的反应元素;然而,只有美联社1激活合作展出。激活ERK信号通路的抑制鸟嘌呤nucleotide-binding蛋白质和被要求对于大多数转录因子对LPA的反应。相比之下,核因子(NF)κB激活LPA但不能EGF和NFκB激活只有当ρ是抑制完全封锁。快速激活ρ在应对LPA但不要EGF。重要的是,抑制ρ选择性阻塞合作在美联社1激活和有丝分裂发生。gydF4y2Ba
总之,细胞外signal-regulated激酶激活需要许多转录因子反应lysophosphatidic酸和表皮生长因子,然而它不协同。激活的激活蛋白1是应承担的协同,ρ合作需要lysophosphatidic酸激活的激活蛋白1应承担的激活和有丝分裂发生。gydF4y2Ba
这项研究得到了内布拉斯加州大学医学中心的种子给马丁加拿大,内布拉斯加州大学的Emley奖学金T.L. Ediger科研经费HL52810和HL56107从国家心脏,肺和血液研究所T.J.墨菲。gydF4y2Ba
调节人类气管平滑肌(对比)细胞增殖是哮喘的病理临床相关,因为增加气道平滑肌质量是哮喘和有助于增强气道狭窄gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2gydF4y2Ba。增加气道平滑肌质量是气道重塑的一个组成部分,一个重要的病理过程不仅哮喘,而且纤维化疾病,如慢性支气管炎。高度增殖的研究对比细胞文化可以揭示潜在的监管机制gydF4y2Ba在活的有机体内gydF4y2Ba1gydF4y2Ba。gydF4y2Ba
对比细胞生长gydF4y2Ba在体外gydF4y2Ba在应对各种各样的刺激增殖,包括G protein-coupled受体(GPCR)受体激动剂以及多肽生长因子作用gydF4y2Ba通过gydF4y2Ba受体酪氨酸激酶(rtk)gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2gydF4y2Ba。从现在和其他作者最近的研究提供了证据表明lysophosphatidic酸(LPA)、血清磷脂促分裂原存在,可能是一个重要的监管机构气道细胞的功能,也建议LPA在气道疾病的可能角色gydF4y2Ba3gydF4y2Ba。对比显示细胞有丝分裂发生增加LPA, GPCR促分裂原,和表皮生长因子(EGF)、RTK生长因子。有趣的是,对比细胞治疗LPA EGF一起显示惊人的协同刺激有丝分裂发生,远远大于反应的总和两个代理gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba。理解信号通路参与协同有丝分裂发生,当细胞接受多重刺激,是了解细胞反应的一个重要方面gydF4y2Ba在活的有机体内gydF4y2Ba,细胞必须集成多个刺激的信号通路激活。gydF4y2Ba
一个关键增殖信号中介是细胞外signal-regulated激酶(ERK)的成员mitogen-activation蛋白激酶(MAPK)的家庭,LPA和表皮生长因子信号通过ERK在许多细胞类型gydF4y2Ba6gydF4y2Ba,gydF4y2Ba7gydF4y2Ba。然而,ERK活化和转录因子受ERK不是唯一参与扩散机制。众所周知,许多转录因子被激活下游GPCRs和rtk的。激活蛋白(美联社)高1环磷酸腺苷反应元件结合蛋白(营)(分子),核转录因子激活T细胞(NFAT)应承担的核转录因子(NF)高κB血清反应元素(行为)复杂、信号传感器和转录激活(STAT)是所有已知参与增殖和/或炎症,涉及相关的炎性状态导致哮喘和呼吸道疾病gydF4y2Ba8gydF4y2Ba。gydF4y2Ba
的协同激活gydF4y2Ba3gydF4y2BaH胸腺嘧啶核苷掺入LPA + EGF前面描述的gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba意味着通路激活这两个有丝分裂原收敛在细胞核中或之前。收敛点的两条途径调解合作代表了有丝分裂发生的主要监管一步感应。本研究的目的是确定网站的信号集成参与合作和利用药理抑制剂调查机制。gydF4y2Ba
材料和方法gydF4y2Ba
研究设计gydF4y2Ba
因为对比细胞表现出协同刺激的有丝分裂发生在回应LPA + EGF,既然兵是一个关键的中介的促有丝分裂的信号,ERK激活的程度和持续时间是决定利用免疫印迹分析细胞治疗LPA, EGF和LPA + EGF。分析转录因子激活作为合作的另一个网站,对比细胞转导与含有一系列的荧光素酶报告基因的逆转录病毒载体由特定转录因子可能参与增殖信号gydF4y2Ba9gydF4y2Ba。在这些研究过程中,作者得到的证据参与的小三磷酸鸟苷5′检测(三磷酸鸟苷)绑定蛋白质ρ,和ρ活化进行了进一步的研究使用Rhotekin Rho-binding域肽选择性识别ρ的三磷酸鸟苷结合应承担的激活状态。gydF4y2Ba
细胞的荧光素酶活性,并对美联社记者基因结构高1分子,NFAT, NF高κB行为复杂,统计,与LPA在治疗后化验,EGF, LPA + EGF。药理抑制剂的信号通路被用来评估特定信号通路的相对贡献的每个转录因子和ERK的活化。接触之前,一项研究调查的有丝分裂原刺激所需的脱氧核糖核酸(DNA)合成并没有发现合作一个8 h后曝光时间LPA + EGF,但强劲的合作后一个12 h曝光时间gydF4y2Ba10gydF4y2Ba。因此,目前的研究都集中在事件发生期间或之前这段时间窗口。gydF4y2Ba
材料gydF4y2Ba
杜尔贝科修改鹰的介质(DMEM)和胎牛血清(的边后卫)从生活购买技术(美国纽约大岛)。18:1(油酰)必经LPA购买从两代情(美国艾尔雪花石膏)和重组在10毫米0.25%脂肪无酸的牛血清白蛋白从σ(圣路易斯,密苏里州,美国)。百日咳毒素(PTX)从列表购买生物制剂(美国坎贝尔,CA)。AG1478, U0126和雷帕霉素是从Calbiochem购买(La Jolla、钙、美国)。Calphostin C是来自Biomol(美国普利茅斯会议上,PA), EGF是Biosource国际(贝南、钙、美国),和gydF4y2Ba3gydF4y2BaH胸苷是从欧宁(美国波士顿)。其他化学物质来自σ。gydF4y2Ba
C3毒素制备和细胞治疗gydF4y2Ba
互补DNA的一个融合蛋白谷胱甘肽S转移酶(GST)和gydF4y2Ba肉毒梭状芽胞杆菌gydF4y2BaC3毒素(包裹量9 e10.c3)是一个慷慨的礼物d·爱尔(Van Andel研究所,大急流城,MI,美国)。向量转化为BL21gydF4y2Ba大肠杆菌gydF4y2Ba(美国Stratagene拉霍亚,Ca);C3毒素净化从对数增长gydF4y2Ba大肠杆菌gydF4y2Ba与0.3毫米isopropylthiogalactoside诱导。细胞细胞溶解在缓冲包含25毫米三pH值7.4,100毫米氯化钠,MgCl 10毫米gydF4y2Ba2gydF4y2Ba,10µg·毫升gydF4y2Ba−1gydF4y2Ba抑肽酶,1毫米phenylmethylsulphonyl氟化物和1毫米二硫苏糖醇(德勤)。澄清提取物与glutathione-Sepharose孵化(分子动力学/ Amersham法玛西亚生物技术,皮斯卡塔韦,新泽西,美国)。洗后,消费税标签在一夜之间被孵化与凝血酶裂解。凝血酶被孵化para-aminobenzamidine-Sepharose和C3毒素被Coomassie量化染色蛋白质通过钠来解决dodecysulphate聚丙烯酰胺凝胶electropheresis (SDS检测页面)。细胞治疗通过添加培养基纯化毒素,交货方法,已被证明是有效的在对比细胞gydF4y2Ba11gydF4y2Ba。gydF4y2Ba
对比细胞培养gydF4y2Ba
对比之前细胞从人类分离气管被酶分解被好心的提供的m·考利考夫(纽约伊萨卡康奈尔大学兽医学院,美国)gydF4y2Ba12gydF4y2Ba。对比细胞培养在high-glucose (4.5 g·LgydF4y2Ba−1gydF4y2Ba每周)DMEM和10%的边后卫和通道。人类胚胎肾(HEK)量293 t细胞对逆转录病毒生产也生长在high-glucose DMEM 5%的边后卫。细胞常规培养37°C湿润5%二氧化碳培养箱。gydF4y2Ba
细胞外signal-regulated激酶激活gydF4y2Ba
对比细胞被镀上12量好或者6孔板,增加融合和饥饿血清DMEM的24小时之前被视为表示。治疗后细胞细胞溶解,溶解产物加载10% SDS聚丙烯酰胺凝胶分离蛋白质。蛋白质被转移到膜和涂抹使用一个antiphospho ERK (Thr202 / Tyr204)抗体,紧随其后的是一个antirabbit辣根过氧化物酶(合)共轭二次抗体(细胞信号技术,贝弗利,妈,美国)。使用化学发光信号检测LumiGLO试剂(细胞信号技术)和暴露于电影。带密度与ImageQuant量化使用扫描激光测密度术和分析软件(分子动力学/ Amersham法玛西亚生物技术)。gydF4y2Ba
生产对比逆转录病毒和转导的细胞gydF4y2Ba
基于小鼠逆转录病毒载体Moloney白血病病毒和荧光素酶报告基因已经生成的先前研究转录因子调节血管平滑肌细胞gydF4y2Ba9gydF4y2Ba。每个向量包含下列转录因子之一的增强剂,克隆的上游最小白介素(IL)量2启动子驱动的规范表达荧光素酶:美联社高1分子,NFAT, NF高κB行为复杂(包括麋鹿、三元复杂的因素,血清反应因子),并统计。pKA9控制向量包含IL 2启动子和荧光素酶基因,但缺乏任何增强子序列。amphotropic包装矢量pSVΨ应承担(-)还是一个高MLV来自o·威特(加州大学洛杉矶分校、钙、美国),获得了从t Smithgall(美国宾夕法尼亚州匹兹堡大学医学院)。逆转录病毒是由cotransfecting记者gene-containing矢量和amphotropic包装成HEK使用磷酸钙转染293 t细胞应承担的协议。逆转录病毒上清液收集在2、3和4天转染后,立即0.45µm无菌过滤器,用于过滤转导细胞对比。对比细胞通过四个或五个镀在40000个细胞·gydF4y2Ba−1gydF4y2Ba在6孔板和转导confluency∼50%。转导,4毫升的逆转录病毒上清液和0.8 mg·毫升gydF4y2Ba−1gydF4y2Ba聚凝胺之前被添加到每个细胞离心机在1200×30分钟gydF4y2BaggydF4y2Ba450年Jouan离心机(美国弗吉尼亚州Jouan实验室设备,温彻斯特),在∼32°C,如前所述gydF4y2Ba13gydF4y2Ba。细胞被离心机共有四次每次都用新鲜的逆转录病毒上清液。对比并行细胞也转导galactosidase-containing向量和沾Xgal确定转导效率,> 95%。转导后,转导细胞通道,用于实验三到六个段落。对比细胞继续生长和分裂后与逆转录病毒转导记者构造用于作者的研究,与细胞周期阻滞之前报道使用adenovirus-mediated转导的荧光素酶报告结构gydF4y2Ba14gydF4y2Ba。gydF4y2Ba
荧光素酶检测gydF4y2Ba
对比细胞被镀在40000细胞·gydF4y2Ba−1gydF4y2Ba在6孔板和发展到融合。采用无血清细胞然后饥饿24小时治疗前无血清培养基的促分裂原。细胞被洗两次在CagydF4y2Ba2 +gydF4y2Ba- - - MggydF4y2Ba2 +gydF4y2Ba200年无磷酸盐和细胞溶解µL裂解缓冲包含三25毫米,4毫米ethyleneglycol-bis -(βaminoethylether应承担)所致gydF4y2BaNgydF4y2Ba,gydF4y2BaNgydF4y2Ba,gydF4y2BaNgydF4y2Ba′,gydF4y2BaNgydF4y2Ba′必经四乙酸(EGTA)、10%甘油、1% Triton X量100,2毫米德勤。10分钟后潜伏期在裂解缓冲,细胞刮和溶菌产物然后由离心澄清在20000×5分钟gydF4y2BaggydF4y2Ba贝克曼微型离心机(美国贝克曼库尔特公司,富勒顿,CA)。荧光素酶活性化验使用Pharmingen Monolight 3010光度计(Pharmingen、圣地亚哥、钙、美国)。50µL细胞溶解产物添加到试管包含350µL荧光素酶检测缓冲区(25毫米三,20毫米MgSOgydF4y2Ba4gydF4y2Ba,EGTA 4毫米,2毫米三磷酸腺苷和1毫米德勤)。注射后100µL 1毫米荧光素在10毫米德勤,相对光单位测量10 s后5 s应承担延迟。gydF4y2Ba
3gydF4y2BaH胸腺嘧啶核苷掺入化验gydF4y2Ba
的gydF4y2Ba3gydF4y2BaH与汇合的胸苷分析进行对比采用无血清细胞饥饿24小时,然后接受促分裂原24 H,正如先前的研究gydF4y2Ba4gydF4y2Ba。gydF4y2Ba
ρ激活化验gydF4y2Ba
选择性分离后的活性形式ρ,西方墨点法是用来评估ρ激活;这种方法是基于的亲和力Rhotekin Rho-binding域。对比细胞被镀在250000细胞/ 100毫米盘和融合发展。采用无血清细胞饥饿24 h,然后对有丝分裂原3分钟。细胞与冰冷的Tris-buffered盐水洗两次,然后细胞溶解在0.5毫升ρ分析裂解缓冲(50毫米三羟甲基氨基甲烷、液pH值7.2,500毫米氯化钠,MgCl 10毫米gydF4y2Ba2gydF4y2Ba,1% Triton X量100,0.5%钠脱氧胆酸盐,0.1% SDS, 1毫米phenylmethylsulphonyl氟化物)。事先批准了溶解产物离心14000×2分钟gydF4y2BaggydF4y2Ba。激活ρ当时孤立使用琼脂糖珠共轭一个销售税融合蛋白包含Rho-binding Rhotekin域(美国纽约北部的生物技术,普莱西德湖),这只三磷酸鸟苷结合量约束ρ的活动形式。一个30µL整除的珠子被加入到溶解产物从每道菜,孵化与温柔旋转45分钟在4°C,孤立的离心14000×5 sgydF4y2BaggydF4y2Ba,洗了三次ρ测定洗缓冲区(50毫米三羟甲基氨基甲烷、液pH值7.2,1% Triton X量100,150毫米氯化钠,MgCl 10毫米gydF4y2Ba2gydF4y2Ba和0.1毫米phenylmethylsulphonyl氟化物)。珠子被resuspended 20µL 2 x Laemmli减少样品的缓冲和煮5分钟。珠子被离心分离颗粒状,上层清液受到地理SDS页10 - 20%梯度凝胶。蛋白质被转移到聚乙二烯二氟化物膜使用半干转移装置。这些膜被涂抹用鼠标anti-RhoA抗体(圣克鲁斯生物技术有限公司、圣克鲁斯、钙、美国),其次是孵化与合量共轭antimouse二级抗体其次是可视化和量化如上所述的ERK激活化验。gydF4y2Ba
分析gydF4y2Ba
数据使用计算机软件进行分析。刺激值ERK和ρ活化利用光密度分析和计算了实验被转换为正常折叠刺激基底然后褶皱刺激值多次实验取平均值。结果的意义是由使用单向方差分析(方差分析)其次是Bonferroni测试后。gydF4y2Ba
转录因子的响应,在每个实验数据被转换为褶皱刺激基底值和数据从多个实验(使用两个或三个独立的转导细胞)然后取平均值。1样本t检测测试被用来确定每个荧光素酶的激活的统计学意义记者构造gydF4y2Ba与gydF4y2Ba控制值为1.0。褶皱的大小反应与褶皱的大小反应enhancerless pKA9控制向量。刺激不明显不同的价值比pKA9控制表明,刺激是由于非特异性转录反应。LPA + EGF治疗的效果与之和与LPA独自孤单,EGF治疗。合作被定义为刺激的≥150%的两个人有丝分裂原刺激的的总和所作者的先前的研究gydF4y2Ba4gydF4y2Ba。双向方差分析被用来统计比较刺激通过LPA + EGF刺激之和仅靠LPA和EGF。在实验测试中抑制剂,未配对的双尾t检测没有测试是用来比较刺激gydF4y2Ba与gydF4y2Ba抑制剂的存在。gydF4y2Ba
结果gydF4y2Ba
在对比细胞ERK活化处理LPA, EGF和LPA + EGFgydF4y2Ba
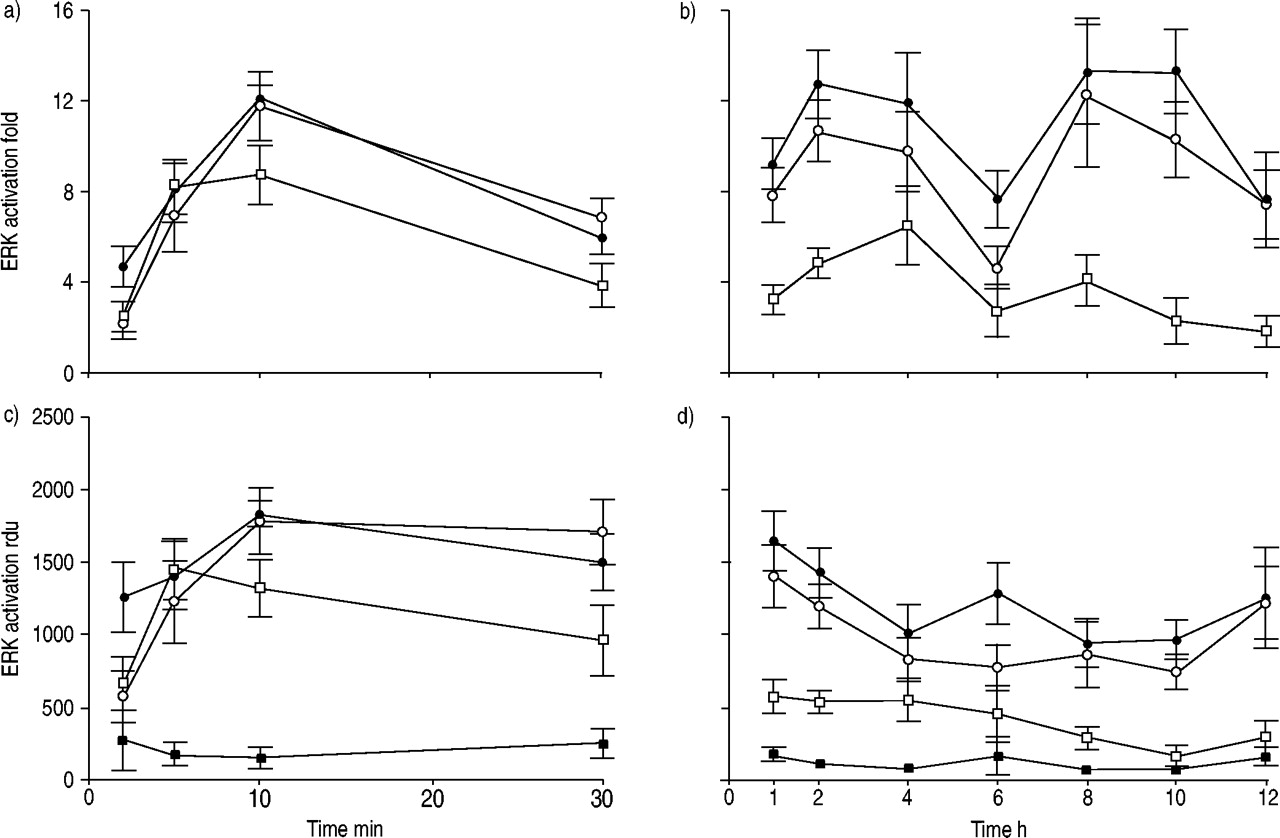
由LPA ERK激活的程度和持续时间,EGF和LPA + EGF测定一个12 h治疗期间(图1所示gydF4y2Ba⇓gydF4y2Ba),合作出现的时间进程gydF4y2Ba10gydF4y2Ba。LPA的治疗产生了强烈对比细胞ERK激活,这是明显的在2分钟,10分钟后到达顶峰。ERK活化,LPA瞬态,重大刺激的和包容的6 h应承担的时间点;6 h后没有观察到显著ERK激活。治疗对比细胞EGF在更早的时间点产生了类似的ERK激活;然而,强大的激活是保留在整个时间进程,其中包括12 h后处理。当对比细胞治疗与LPA结合EGF、ERK激活平行,仅与EGF,显示出强大的激活整个时间进程(图1所示gydF4y2Ba⇓gydF4y2Ba)。一些实验表明添加剂ERK活化反应在指定时间点;最常见的是在6 h。然而,这种添加剂效果并不是始终存在协同效应对ERK激活并没有观察到。gydF4y2Ba
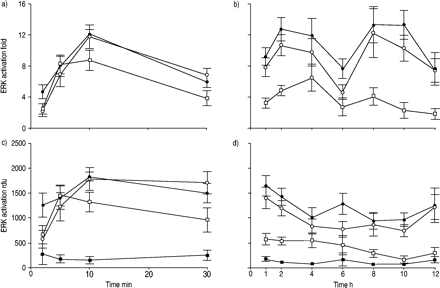
激活细胞外signal-regulated激酶(ERK)在人类气管平滑肌细胞治疗后评估与牛血清白蛋白(BSA;▪),10μm lysophosphatidic酸(LPA;□),60 ng·毫升gydF4y2Ba−1gydF4y2Ba表皮生长因子(EGF);○)或LPA + EGF (•)。显示的值代表均值±sem四到七个独立的实验。单向方差分析显示ERK显著激活LPA (p < 0.05),其中包括6 h时间点,但不是在稍后时间。ERK激活通过EGF和LPA + EGF是重要的时间点。b)和d)中所示的数据表明,低水平的刺激在6 h主要反映了BSA控制价值的增加。rdu:相对密度单位。gydF4y2Ba
信号通路参与ERK激活LPA和EGFgydF4y2Ba
LPA的ERK信号途径的激活,EGF和LPA + EGF评估使用药理抑制剂在10分钟或2 h化验。ERK激活LPA被PTX预处理,暗示抑制鸟嘌呤nucleotide-binding蛋白(GgydF4y2Ba我gydF4y2Ba)在ERK激活LPA(图2所示gydF4y2Ba⇓gydF4y2Ba)。正如所料,EGF不是由PTX抑制ERK激活。ERK激活通过EGF被EGF RTK抑制剂AG1478地理。在一些系统中GPCR有丝分裂原,例如LPA,导致EGF的独立transactivation应承担的表皮生长因子受体(EGFR),因此,表皮生长因子受体也被认为在促有丝分裂的信号下游许多GPCR的有丝分裂原gydF4y2Ba7gydF4y2Ba,gydF4y2Ba15gydF4y2Ba。然而,AG1478没有阻断ERK激活LPA,表明EGF检测不需要RTK LPA刺激ERK激活。U0126是地理MAPK ERK激酶的抑制剂(MEK),直接上游激酶ERK和负责ERK激活。U0126阻塞ERK激活LPA和表皮生长因子,主要由MEK表明ERK激活,尽管不完全抑制活化的LPA + EGF可能允许激活ERK的一个额外的小通道。光密度分析的多个实验显示,U0126抑制ERK激活LPA + EGF只有76±4%,而U0126阻塞LPA的刺激ERK激活92±4%和EGF刺激ERK激活99±1% (n = 4)。gydF4y2Ba
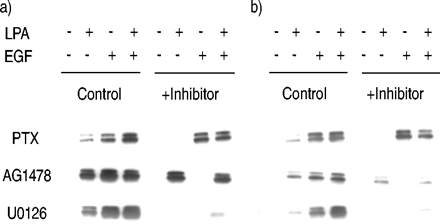
抑制剂对细胞外signal-regulated激酶(ERK)激活lysophosphatidic酸(LPA)和表皮生长因子(EGF)后)10分钟,b) 2 h治疗。人类气管平滑肌细胞治疗百日咳毒素(PTX;100 ng·毫升gydF4y2Ba−1gydF4y2Ba),AG1478(2.5µM)或U0126(10µM) 30分钟,然后用牛血清白蛋白治疗,10µM LPA, 60 ng·毫升gydF4y2Ba−1gydF4y2BaEGF、LPA + EGF 10分钟或2 h,和磷量ERK然后分析。一个代表性的污点从至少三个实验。gydF4y2Ba
其他抑制剂检测其对ERK活化的影响包括Src抑制剂PP1磷脂酰肌醇3′检测激酶(PI3K)抑制剂LY294002核糖体蛋白s6激酶(p70gydF4y2Bas6kgydF4y2Ba)抑制剂雷帕霉素,磷脂酶C抑制剂U73122,ρ抑制剂C3毒素。这些抑制剂被发现改变ERK激活LPA, EGF,或者LPA + EGF在10分钟或2 h化验(数据未显示)。gydF4y2Ba
激活的转录因子在12 h治疗与LPA, EGF, LPA + EGFgydF4y2Ba
对比细胞转导面板的荧光素酶记者构造处理LPA, EGF,或LPA + EGF 12 h(表1所示gydF4y2Ba⇓gydF4y2Ba)。细胞转导pKA9矢量,不包含增强剂,包括作为一个消极的控制。此外,反应为每个转录因子是典型的代理作为积极的控制。gydF4y2Ba
转录因子的激活报告基因结构在人类气管平滑肌细胞(对比)gydF4y2Ba
LPA单独治疗诱导荧光素酶活性在细胞基因结构轴承美联社高1分子,NFAT, NFκB和行为复杂。LPA没有显著激活统计记者构造。EGF诱导转录由美联社高1分子,NFAT和行为复杂。尽管NFκB和属性的值> 1.0,他们没有显著高于控制pKA9向量,表明EGF并不特别激活这些转录因子。当细胞被LPA + EGF处理,只有美联社1记者构造表现出协同激活。分子的报告基因,NFAT行为复杂显示激活的生长因子,大约是这对个人成长因素的总和。相比之下,NFκB显示激活的LPA强劲,但不能通过EGF和NFκB激活通过LPA没有明显改变表皮生长因子的存在。gydF4y2Ba
荧光素酶活性还测量了4 h后治疗。有趣的是,据美联社1激活应承担LPA, EGF, LPA + EGF只是添加剂4 h,与协同响应在12 h。4 h褶皱的刺激值分别为2.4±0.4,2.0±0.1,4.8±0.8,分别为(n = 3)。其他记者的刺激值构造类似4和12 h;NFκB应承担的激活,LPA 4 h为4.7±1.2折(n = 4)又无显著激活NFκB应承担的观察EGF(1.3±0.1折;n = 4)。gydF4y2Ba
信号通路参与AP-1激活LPA和EGFgydF4y2Ba
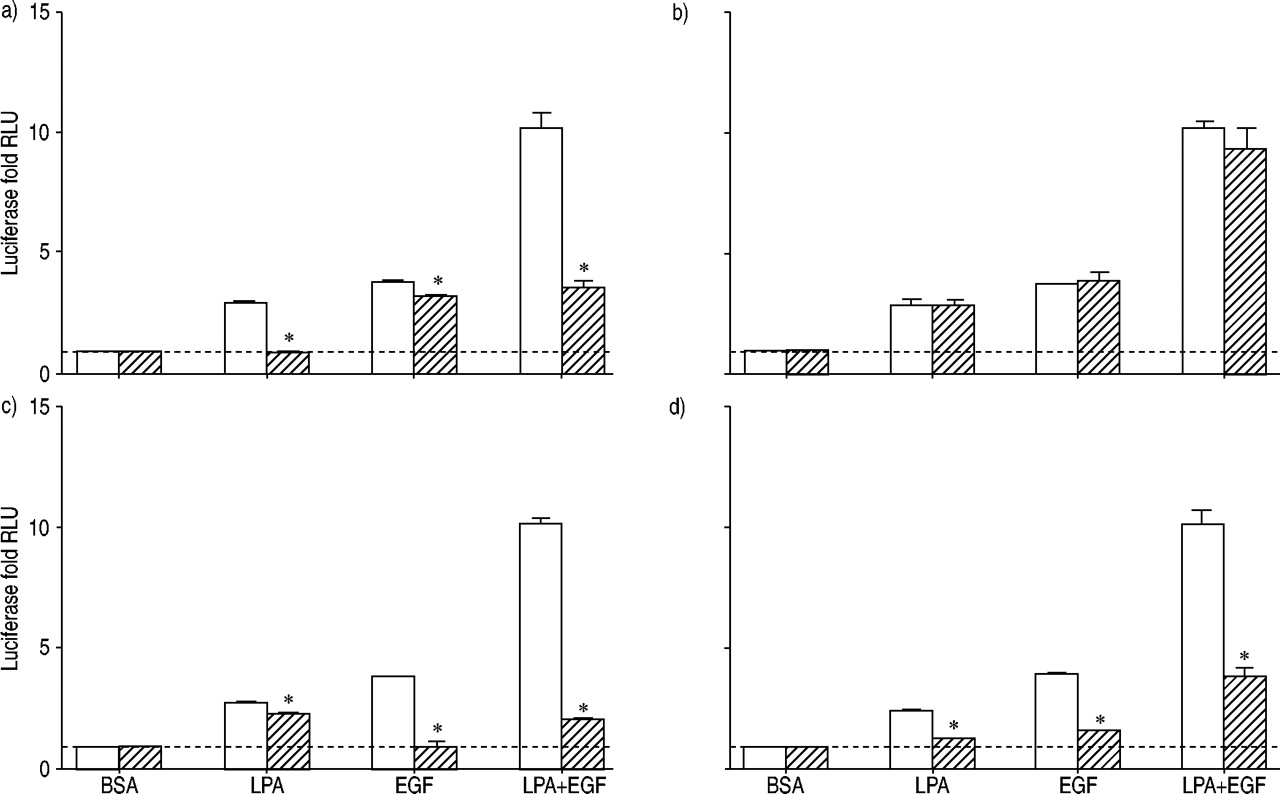
因为转录因子美联社检测1建立了扩散的重要中介gydF4y2Ba16gydF4y2Ba并协同激活LPA + EGF,信号通路抑制剂的影响在美联社1激活LPA和表皮生长因子进行调查(图3所示gydF4y2Ba⇓gydF4y2Ba)。预处理细胞PTX完全阻塞LPA必经介导美联社高1激活在暗示GgydF4y2Ba我gydF4y2Ba进入LPA刺激美联社1激活。PTX造成只有很小(但统计学意义)抑制EGF的AP 1应承担的激活。在PTX面前,激活AP所致LPA + EGF等于1,独自与EGF。因为LPA在对比也可以激活《Gq》和磷脂酶C细胞gydF4y2Ba17gydF4y2Ba,可能作用的蛋白激酶C (PKC)据美联社1激活了。PKC抑制剂calphostin C没有阻止美联社高1激活LPA在EGF、LPA + EGF(图3所示gydF4y2Ba⇓gydF4y2Ba不需要),这表明PKC激活AP 1激活这些刺激。gydF4y2Ba
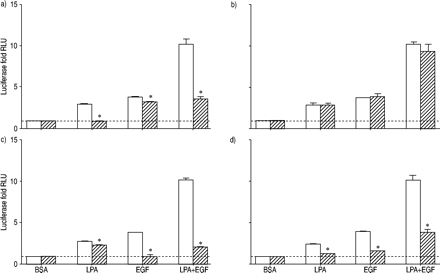
荧光素酶活性评估人类气管平滑肌细胞包含激活蛋白(美联社)高1报告基因治疗12 h和10µM lysophosphatidic酸(LPA), 60 ng·毫升gydF4y2Ba−1gydF4y2Ba表皮生长因子(EGF), LPA + EGF,或适当的牛血清白蛋白(BSA)控制在没有(□)或存在(└)的抑制剂:百日咳毒素100 ng·毫升gydF4y2Ba−1gydF4y2Ba,b) calphostin C 1µM, C) AG1478 2.5µM, d) U0126µg·10毫升gydF4y2Ba−1gydF4y2Ba。值所代表的意思是±sem三至六实验利用两个转导的细胞。- - -:基线正常化BSA控制;RLU:相对光单位。*:p < 0.05。gydF4y2Ba
细胞治疗与LPA + EGF RTK抑制剂AG1478不应承担的块LPA量刺激美联社1激活(图。3gydF4y2Ba⇑gydF4y2Ba),虽然小但发现统计上显著的抑制。这些结果表明,表皮生长因子受体酪氨酸激酶活性的扮演一个小角色,如果有的话,进入调解LPA刺激美联社1激活。AG1478做块表皮生长因子介导应承担的美联社高1激活。与LPA + EGF刺激AG1478的存在是一样的,看到LPA孤单。PTX和AG1478孵化的细胞,阻止GgydF4y2Ba我gydF4y2Ba介导LPA效应和表皮生长因子受体介导的EGF应承担的效果,完全封锁了美联社高1激活LPA + EGF在按预期(数据没有显示)。gydF4y2Ba
MEK抑制剂U0126阻塞AP 1激活LPA和EGF,暗示ERK的活化据美联社1由代理(图。3gydF4y2Ba⇑gydF4y2Ba)。有趣的是,然而,U0126没有完全阻止AP所致1激活LPA + EGF的结合。没有U0126,美联社量1激活LPA + EGF增加了10.2±1.1必经褶皱基底,而在U0126的存在,美联社量1激活LPA + EGF还增加了3.5±0.1折。U0126无法完全阻止美联社1激活LPA + EGF是一致的与观察,U0126并不完全阻断ERK激活LPA + EGF(图。2gydF4y2Ba⇑gydF4y2Ba)。这表明U0126未能完全阻止AP所致1激活LPA + EGF可能结果的不完全抑制ERK激活这个组合的有丝分裂原。或者,可能有一个兵独立分量的AP 1激活LPA + EGF。gydF4y2Ba
最近的一项研究表明p70的角色gydF4y2Bas6kgydF4y2Ba在对比细胞促有丝分裂的信号和合作gydF4y2Ba14gydF4y2Ba。然而,在目前的实验中,p70gydF4y2Bas6kgydF4y2Ba通路抑制剂雷帕霉素不据美联社1激活和抑制可能增强AP 1激活。褶皱的刺激值LPA、EGF和LPA + EGF分别为2.5±0.2,3.3±0.3,9.3±0.1没有雷帕霉素和3.2±0.2,4.8±0.3,16.3±0.6的1µM雷帕霉素(n = 3)。因此,p70gydF4y2Bas6kgydF4y2Ba似乎并不需要协同美联社1对比细胞激活和p70的贡献吗gydF4y2Bas6kgydF4y2Ba有丝分裂发生或合作在这些细胞可能通过其他机制比美联社1激活。gydF4y2Ba
ERK和GgydF4y2Ba我gydF4y2Ba激活LPA和EGF监管其他转录因子gydF4y2Ba
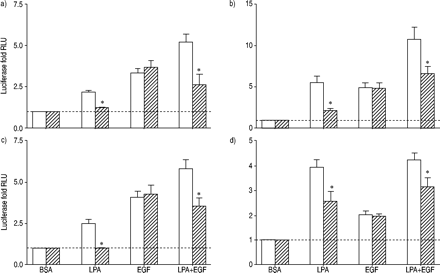
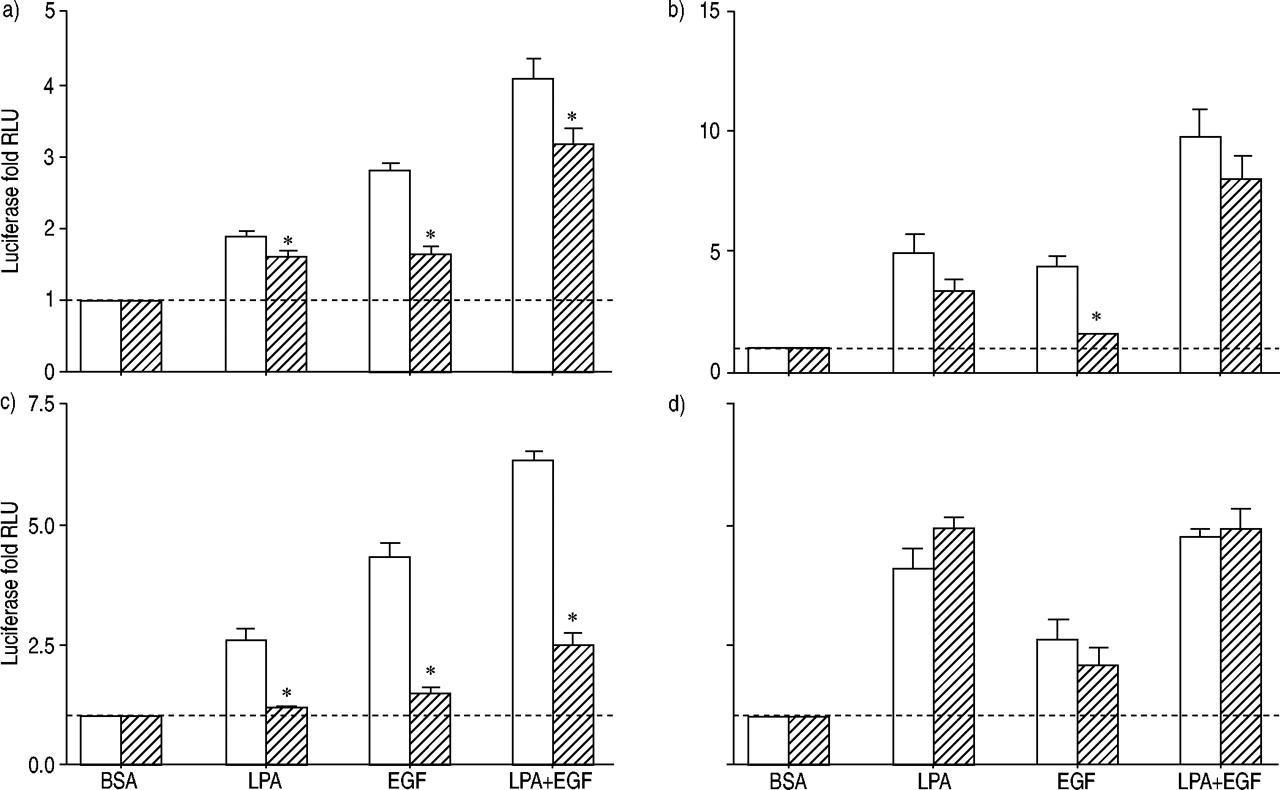
为LPA列车激活转录因子介导的信号途径除了美联社1了。分子的U0126对活化的影响,行为复杂,NFAT和NFκB LPA, EGF和LPA + EGF评估(图4所示gydF4y2Ba⇓gydF4y2Ba)。U0126没有显著抑制LPA的刺激激活分子;然而,通过EGF刺激明显抑制。活化分子的LPA + EGF没有明显抑制。结果是复杂的分子是相似的;gydF4y2Ba即。gydF4y2BaLPA弱抑制,EGF明显抑制,LPA + EGF略微下降。U0126几乎完全阻塞NFAT激活LPA, EGF和LPA + EGF的结合。相比之下,U0126对NF量没有影响κB激活LPA或LPA + EGF的结合。gydF4y2Ba
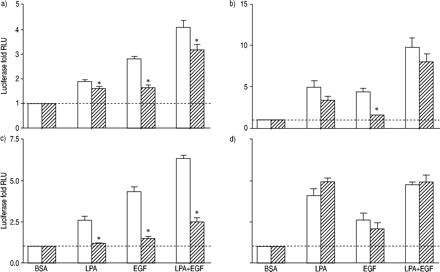
荧光素酶活性评估人类气管平滑肌细胞包含)的环磷酸腺苷反应元件结合蛋白,b)血清反应元素,c)核转录因子激活T细胞,应承担和d)核因子高κB报告基因治疗12 h和10µM lysophosphatidic酸(LPA), 60 ng·毫升gydF4y2Ba−1gydF4y2Ba表皮生长因子(EGF), LPA + EGF,或适当的牛血清白蛋白(BSA)控制在没有(□)或存在(└)10µM U0126。值所代表的意思是±sem三到四个实验有两个细胞转导。- - -:基线正常化BSA控制;RLU:相对光单位。*:p < 0.05。gydF4y2Ba
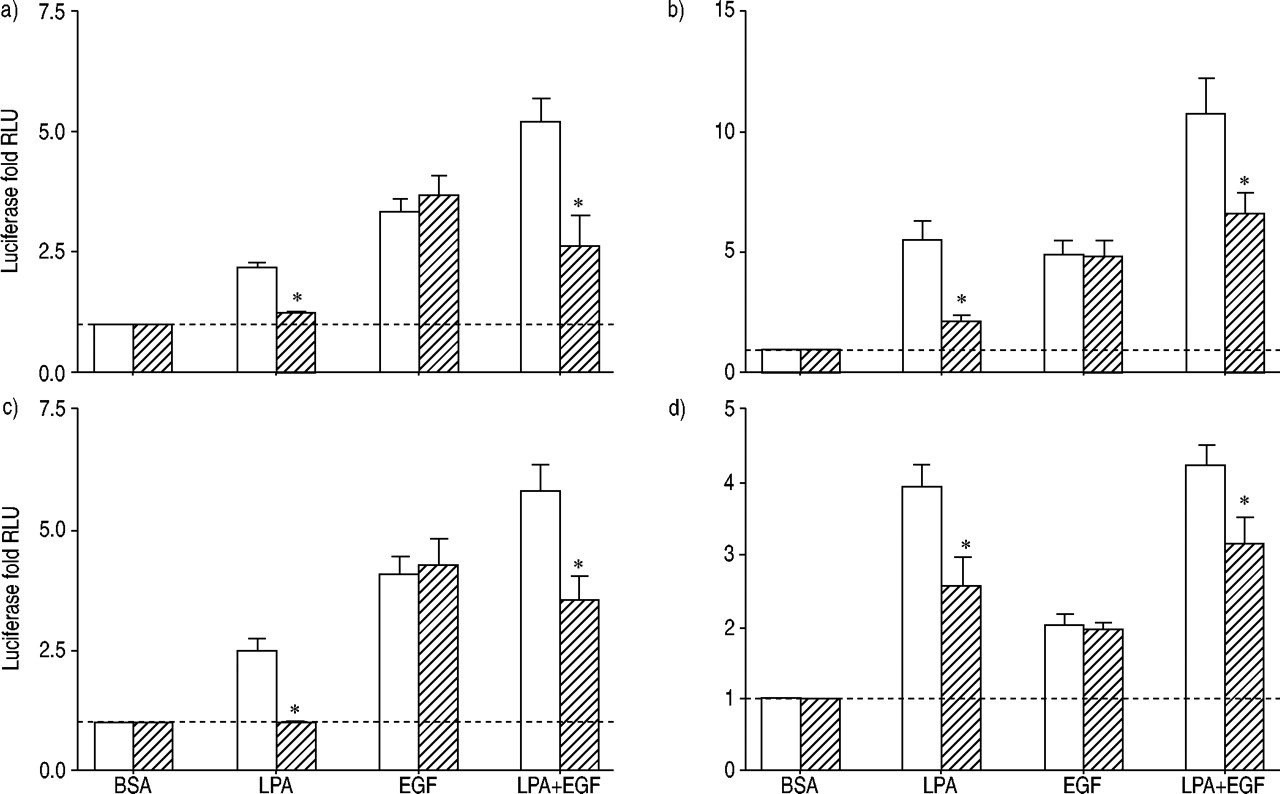
PTX在LPA的影响量调节分子的活化,行为复杂,NFAT, NFκB应承担的评估(图5所示gydF4y2Ba⇓gydF4y2Ba)。激活的分子和NFAT几乎完全被PTX。激活的行为复杂的主要是PTX高敏感而NFκB激活应承担的由PTX LPA仅略有降低。EGF激活所有的这些反应是PTX高不敏感。的行为复杂,分子和NFAT PTX治疗应承担的细胞表现出反应LPA + EGF类似与EGF。gydF4y2Ba
荧光素酶活性评估人类气管平滑肌细胞包含)的环磷酸腺苷反应元件结合蛋白,b)血清反应元素,c)核转录因子激活T细胞,应承担和d)核因子高κB报告基因治疗12 h和10µM lysophosphatidic酸(LPA), 60 ng·毫升gydF4y2Ba−1gydF4y2Ba表皮生长因子(EGF), LPA + EGF,或适当的牛血清白蛋白(BSA)控制在没有(□)或存在(└)100 ng·毫升gydF4y2Ba−1gydF4y2Ba百日咳毒素。值所代表的意思是±sem三到四个实验有两个细胞转导。- - -:基线正常化BSA控制;RLU:相对光单位。*:p<0.05 using a two-tailed t‐test.
ρ的影响还是灭活C3毒素LPA激活的NFκB和美联社1和协同刺激的有丝分裂发生gydF4y2Ba
LPA的相对不敏感所致介导NFκB应承担的激活PTX或U0126建议另一种途径NFκB应承担的激活可能参与其中。最近的报告提出了ρ的中介NF其他GPCRκB激活受体激动剂gydF4y2Ba18gydF4y2Ba,gydF4y2Ba19gydF4y2Ba。因此,ρ抑制剂C3毒素的影响在NFκB激活测试。C3毒素排除NF必经κB激活LPA,暗示ρNFκB激活(图。6gydF4y2Ba⇓gydF4y2Ba)。因为C3毒素阻塞NFκB应承担的激活,因为美联社的一个组成部分1激活LPA + EGF U0126麻木不仁,C3毒素的影响在美联社1激活还测试了。治疗与C3的细胞毒素并没有阻止刺激美联社1 LPA或EGF。然而,C3毒素排除了合作在美联社1激活LPA + EGF。这些结果表明,ρ可能是唯一参与协同激活AP所致1×LPA + EGF,即使它似乎并不需要单独或生长因子的响应。gydF4y2Ba
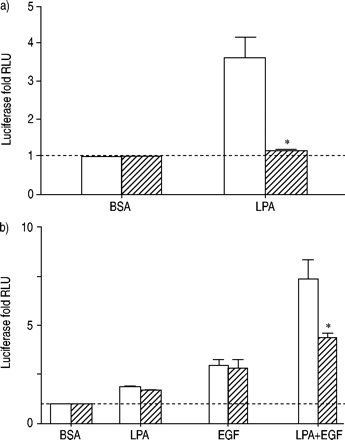
荧光素酶活性评估人类气管平滑肌细胞包含一个)核转录因子(NF)κB或者b)激活蛋白(美联社)1,治疗12 h和10µM lysophosphatidic酸(LPA), 60 ng·毫升gydF4y2Ba−1gydF4y2Ba表皮生长因子(EGF), LPA + EGF、牛血清白蛋白(BSA)控制在没有(□)或存在(└)10 ng·毫升gydF4y2Ba−1gydF4y2BaC3毒素。价值观代表了均值±sem的两到四个实验中使用两个细胞转导。C3毒素增加基底值2.9±0.4折等控制AP所致1细胞,7.8±1.3折等控制NFκB应承担的细胞,和1.7±0.1折为血清反应元素控制复杂的细胞。显示的值是治疗的褶皱增加细胞相对于这些升高值的控制。- - -:基线正常化BSA控制;RLU:相对光单位。*:p < 0.05使用一个双尾t测试。gydF4y2Ba
C3毒素在有丝分裂发生评估的影响gydF4y2Ba3gydF4y2BaH胸苷并入DNA也被调查。发现部分C3毒素抑制有丝分裂发生仅靠LPA刺激;然而,它没有影响仅靠EGF刺激(图7所示gydF4y2Ba⇓gydF4y2Ba)。重要的是,合作是消除C3毒素,刺激LPA + EGF水平降低,只有添加剂,而非协同。这表明,ρ激活参与之间的合作LPA和EGF水平的DNA合成,类似于上述结果与美联社1激活化验(图。6gydF4y2Ba⇑gydF4y2Ba)。gydF4y2Ba
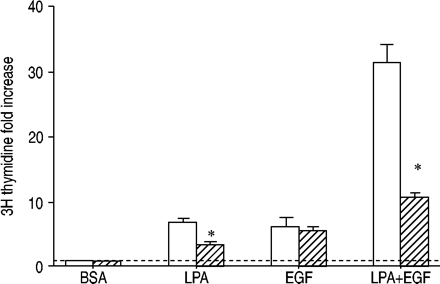
3gydF4y2BaH胸腺嘧啶核苷掺入被评估人气道平滑肌细胞治疗与牛血清白蛋白(BSA), 10µM lysophosphatidic酸(LPA), 60 ng·毫升gydF4y2Ba−1gydF4y2Ba表皮生长因子(EGF),或LPA + EGF 24 h没有(□)或存在(└)µg·10毫升gydF4y2Ba−1gydF4y2BaC3毒素。值所代表的意思是±sem对三个实验。- - -:基线正常化BSA控制;*:p < 0.05使用一个双尾t测试。gydF4y2Ba
LPA和EGFρ激活的影响gydF4y2Ba
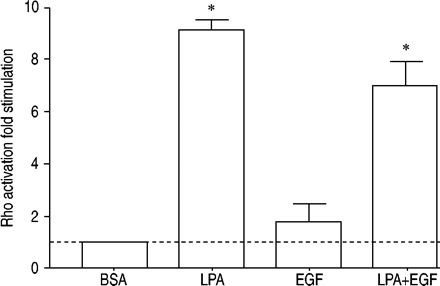
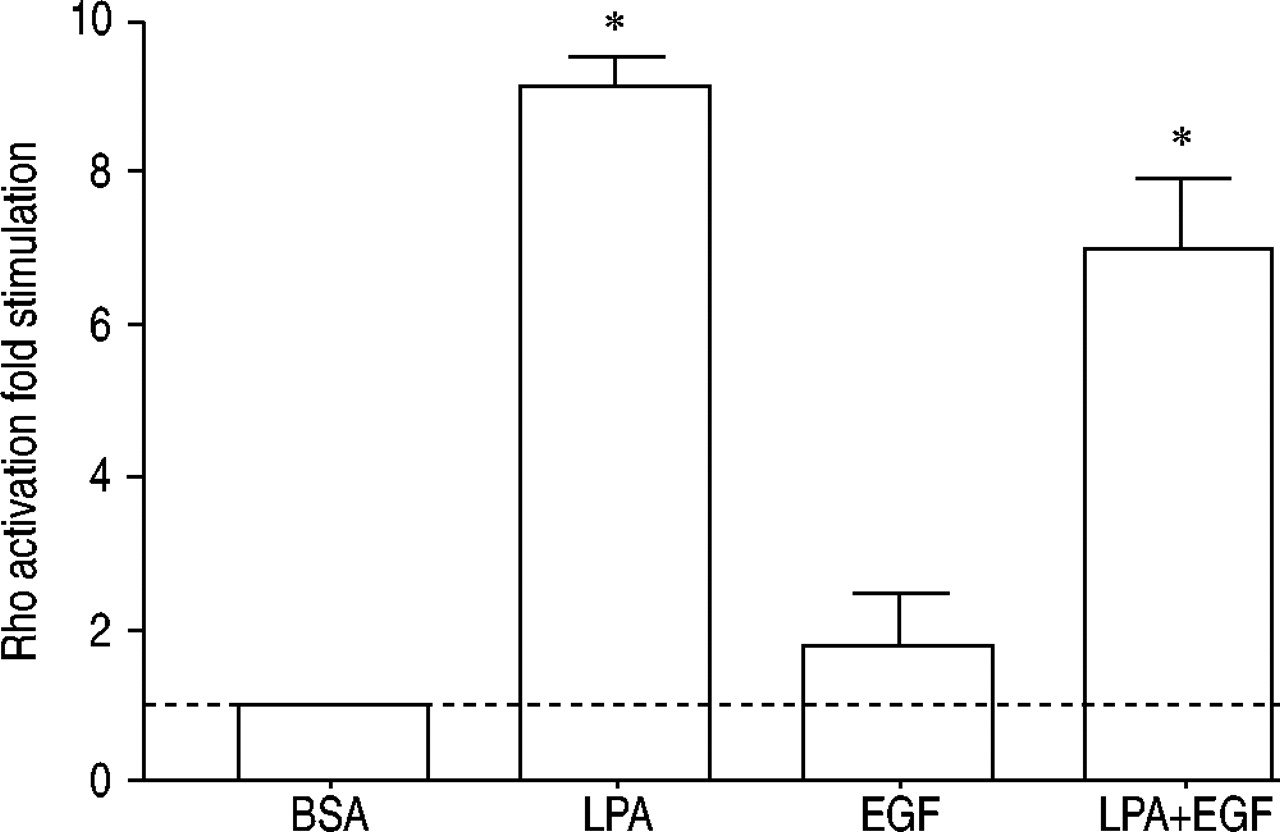
因为C3毒素抑制作用的具体对比细胞有丝分裂发生协同组成部分,LPA的影响,EGF和LPA + EGFρ激活进行评估。这些研究被进行确认ρ是激活,识别哪些促分裂原调和ρ激活和确定合作是否观察到ρ的激活水平。Rho-binding域Rhotekin粘附在琼脂糖珠的GST融合蛋白用于应承担的地理隔离三磷酸鸟苷结合活性形式的ρ,当时量化通过免疫印迹anti-Rho抗体(图8所示gydF4y2Ba⇓gydF4y2Ba)。LPA诱导显著ρ的刺激激活,但没有显著激活的ρEGF。LPA + EGF的结合并没有表现出合作ρ激活,而是效果低于LPA单独出现。gydF4y2Ba
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
ρ激活被评估人气道平滑肌细胞刺激与牛血清白蛋白(BSA), 10µM lysophosphatidic酸(LPA), 60 ng·毫升gydF4y2Ba−1gydF4y2Ba表皮生长因子(EGF),或LPA + EGF 3分钟。值表示为褶皱的刺激与BSA控制和从三个均值±sem实验。- - -:基线正常化BSA控制;*:p < 0.05单向方差分析其次是Bonferroni测试后。gydF4y2Ba
讨论gydF4y2Ba
这些研究的目的是探讨LPA的潜在的集成点,EGF-signalling通路下游对比的受体激活细胞,识别这些通路的点收敛于调解DNA合成的协同刺激。LPA和EGF-activated途径导致ERK激活;然而,合作没有观察到的ERK激活水平。LPA和EGF导致多个转录因子的激活,包括分子NFAT,美联社量1,行为复杂。此外,ERK信号通路似乎参与这些转录因子激活基于U0126的抑制效应。与LPA + EGF治疗对比细胞产生协同激活AP 1。抑制ρ激活与C3毒素封锁了合作在美联社高1激活和协同有丝分裂发生在暗示ρ在协同信号。LPA而不是EGF似乎是介导该ρ激活的主要因素。因此,ERK活化和ρ激活所需的促有丝分裂的信号;然而,无论是合作的轨迹。 AP‐1 activation does exhibit synergism and as such represents a likely point of integration for signals from LPA and EGF.
ERK激活的信号通路进行调查,因为可能ERK活化的重要性导致下游合作一些轨迹。符合已知的ERK活化途径其他细胞,激活通过EGF表皮生长因子受体酪氨酸激酶活性的要求。激活LPA是由GgydF4y2Ba我gydF4y2Ba或者另一个PTX敏感的G蛋白但似乎没有要求应承担的EGF RTK应承担的活动。这一发现是一致的与另一个来自作者的实验室的最新研究表明更直接,LPA不刺激表皮生长因子受体酪氨酸的磷酸化gydF4y2Ba10gydF4y2Ba。MEK激活ERK激活所需的LPA和EGF。gydF4y2Ba
了多方面的证据表明ERK激活合作是不够的。首先,ERK激活通过LPA + EGF没有大于单靠EGF。第二,尽管持续ERK激活似乎的关键信号在几个细胞类型包括对比细胞增殖gydF4y2Ba20.gydF4y2Ba- - - - - -gydF4y2Ba22gydF4y2Ba,激活ERK的LPA瞬态无关紧要的ERK激活LPA在8 - 12 h治疗期,这是合作的重要时间窗口gydF4y2Ba10gydF4y2Ba。然而,ERK激活通过EGF持续12小时里的曝光与对比细胞以前的研究显示出类似的结果gydF4y2Ba21gydF4y2Ba。这种持续ERK活化可能代表的重大贡献EGF在对比细胞协同促有丝分裂的信号。第三,PTX敏感性LPA反对的ERK激活ERK作为合作的关键,因为LPA和EGF之间的合作不是PTX十分敏感gydF4y2Ba5gydF4y2Ba。尽管ERK激活本身不是协同,激活ERK还是促有丝分裂的信号和合作的关键,基于抑制U0126对转录因子的影响下游ERK活化的反应。gydF4y2Ba
在最近的一项研究中,作者表明,治疗的对比与LPA诱导细胞EGFR检测绑定的一个近似双重upregulation活动是由增加举的表达式gydF4y2Ba10gydF4y2Ba。Upregulation LPA举的是一个有吸引力的解释合作的机制,因为它可以解释的要求LPA和EGF刺激最大,因为表皮生长因子受体Upregulation课程的时间与课程的时间开始合作。一个明显的从这个模型预测的upregulation EGFRs将导致EGF刺激ERK激活应承担的相应增加。然而,在当前研究ERK激活不是LPA + EGF比单独使用EGF在12 h,时间点的最大EGFR upregulation LPA发生gydF4y2Ba10gydF4y2Ba。这些结果表明,任何贡献的EGFR upregulation合作发生通过ERK激活以外的信号通路。gydF4y2Ba
目前研究还文档的多个转录因子激活LPA和EGF对比细胞。最好的作者的知识,这是第一次报告的LPA介导活化分子。尽管LPA对阵营形成抑制性影响,分子也可以通过ERK激活,PI3K和calcium-signalling通路gydF4y2Ba23gydF4y2Ba,都是激活LPA在这些或其他细胞gydF4y2Ba6gydF4y2Ba。同样,LPA介导NFAT反应之前没有记录。NFAT监管主要是通过激活钙信号和钙调磷酸酶,LPA并增加细胞内钙对比细胞(未发表的数据)。NF必经κB LPA已经证明活化的成纤维细胞和内皮细胞gydF4y2Ba24gydF4y2Ba,gydF4y2Ba25gydF4y2Ba。LPA行为复杂的激活增加的影响建立了纤维母细胞细胞系gydF4y2Ba26gydF4y2Ba。因为c量gydF4y2Ba”丛书gydF4y2Ba是一个组件的美联社1和c量gydF4y2Ba”丛书gydF4y2Ba基因包含特征明显行为和CRE序列gydF4y2Ba27gydF4y2Ba,转录增加c量gydF4y2Ba”丛书gydF4y2Ba表达这些因素的激活是一个潜在的解释LPA激活AP 1应承担的能力,尽管这还有待建立。表皮生长因子已被证实能激活c量gydF4y2Ba”丛书gydF4y2Ba行为gydF4y2Ba26gydF4y2Ba和分子gydF4y2Ba28gydF4y2Ba在其他类型的细胞,这可能也是EGF机制激活AP 1。尽管EGF不是古典NFAT的活化剂,最近的一份报告表明EGF可能激活NFAT Vav1通过激活的T细胞gydF4y2Ba29日gydF4y2Ba。gydF4y2Ba
所有被AG1478抑制表皮生长因子介导应承担的反应,表明中介的古典EGF RTK通路。所有EGF反应也被U0126,暗示MEK和大概ERK downstream-signalling介质的这些转录因子对EGF的反应。美联社量1分子行为和NFAT响应U0126 LPA也被抑制。然而,NFκB应承担的激活报告基因是U0126完全不敏感。同样,LPA激活AP高1分子,行为和NFAT PTX高敏感而NFκB激活不应承担PTX十分敏感。gydF4y2Ba
ρ是最好的一个小G蛋白作为细胞骨架调制器的响应特征,但最近的证据显然涉及ρ在调节基因转录和细胞增殖gydF4y2Ba30.gydF4y2Ba。重要的是,ρ抑制剂C3毒素阻塞协同美联社1激活LPA + EGF但不能美联社1激活仅靠LPA或EGF。因此,ρ激活似乎所需的额外美联社1激活观察到当LPA和EGF同时存在。这些数据表明ρ作为一个信号通路导致合作的重要组成部分。在对比结果与C3毒素细胞有丝分裂发生化验表明,ρ激活可能是合作的关键观察有丝分裂发生的评估gydF4y2Ba3gydF4y2BaH胸腺嘧啶核苷掺入也类似于结果的激活AP 1。这些研究都指向一个角色ρ作为协同信号在扩散的关键信号。进一步的研究表明,LPA的主要中介而不是EGFρ激活,符合最近的证据ρLPA激活的细胞对比gydF4y2Ba31日gydF4y2Ba也在其他细胞类型gydF4y2Ba32gydF4y2Ba。ρ的机制参与LPA激活这些细胞及相关ρ效应器参与合作是令人兴奋的未来研究方向。gydF4y2Ba
NF PTXκB激活是最低限度敏感和不受U0126,表明NFκB应承担的激活发生,另一种LPA的信号通路。因为合作不是PTX高敏感的,而不是由ERK在这选择NFκB应承担的途径也可能促成合作。有趣的是,两种NFκB和ρ被EGF LPA但不激活。此外,NF必经κB LPA被激活的C3毒素,表明ρLPA需要激活激活NFκB。这个结果是一致的与其他最近的报告,NFκB应承担的激活可能是受ρ。Rho-mediated NF必经κB激活已经证明了缓激肽治疗A549肺癌细胞gydF4y2Ba18gydF4y2Ba和鞘氨醇1磷酸盐处理的HEK293细胞gydF4y2Ba19gydF4y2Ba。因此,Rho-mediated NFκB应承担的激活是一个有趣的可能性的贡献LPA的合作中观察到两个AP 1激活和有丝分裂发生化验。gydF4y2Ba
美联社1应承担的核心作用是由最近的一项研究调查其他协同对比细胞分裂素组合,特别是EGF加凝血酶,组胺、氯化氨甲酰胆碱gydF4y2Ba14gydF4y2Ba。虽然ERK激活这些组合不协同,增强激活AP所致的1,麋鹿1观察,即使使用adenovirus-mediated转导的细胞周期阻滞引起的荧光素酶报告结构gydF4y2Ba14gydF4y2Ba。本研究的时间进程数据也符合一个角色美联社1在合作,如美联社量1激活后没有协同4 h与LPA + EGF治疗,但经过12 h的治疗。这些结果平行之前报道的时间进程协同活化的有丝分裂发生在12 h与LPA + EGF治疗被要求遵守合作gydF4y2Ba10gydF4y2Ba。进一步研究AP所致1激活地理描述具体美联社1因素参与合作和他们是如何受到信号通路激活LPA和EGF可能进一步洞察AP所致1合作的机制。此外,激活AP 1可能会进一步把与其他转录因子如NF必经κB直接基因启动子区域的交互调节扩散中扮演关键的角色。自美联社1和NFκB都被卷入的规定应承担的气道重塑的炎症信号发生疾病,如哮喘、进一步研究这两个转录因子的调控及其相关激活应该产生结果,对呼吸道疾病的发病机理和治疗的认识。gydF4y2Ba
最近发表的一项研究表明,同时激活Ras-mediated和Rho-mediated通路可以在血管平滑肌细胞协调合作。持续活跃的Ras和ρ构造转染成大鼠血管平滑肌细胞协同刺激扩散引起的gydF4y2Ba33gydF4y2Ba。基于研究结果在最近的研究中,一个类似的模型提出了气道平滑肌细胞,与Ras-activated Rho-activated通路相互作用导致合作gydF4y2Ba3gydF4y2Ba。表皮生长因子,提出了通过ERK激活持续合作作出贡献,可能是下游的Ras。这种持续的ERK激活细胞内未见LPA单独对待。LPA有助于合作通过Rho-mediated通路的激活,一个候选人ρ的下游效应器NFκB应承担的这个系统。因此协同激活AP 1和细胞有丝分裂发生在对比的协同刺激可能造成的影响一起LPA激活ρ的强烈影响表皮生长因子激活Ras和调解持续激活ERK。信号通路激活的收敛Ras和ρ调节转录因子包括NFκB和美联社1提出了一个简单而优雅的模型对协同信号有丝分裂发生,LPA和表皮生长因子激活互动小G蛋白通路。gydF4y2Ba
这些研究发现的许多重要转录因子刺激lysophosphatidic酸和表皮生长因子和关键信号通路调节气道平滑肌细胞的活动。更详细的调查所涉及的分子机制之间的相互作用这些mitogenic-signalling通路可能提供新的目标调制气管平滑肌的增加gydF4y2Ba在活的有机体内gydF4y2Ba在病理条件下的哮喘气道重塑和其他疾病。gydF4y2Ba
确认gydF4y2Ba
作者要感谢j·林奇与统计分析寻求帮助,t . Smithgall o·威特,a . Alberts和m·考利考夫试剂和r·刘易斯有益的讨论。gydF4y2Ba
- 收到了gydF4y2Ba2002年8月15日。gydF4y2Ba
- 接受gydF4y2Ba2002年12月13日。gydF4y2Ba
- ©人期刊有限公司gydF4y2Ba
引用gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba









