





文摘
支气管哮喘是由于持续的慢性气道炎症和各种各样的结构性变化,或改造,包括平滑肌和杯状细胞增生。
探讨气道重塑的机制,目前的作者使用了一个建立过敏原(卵白蛋白;卵子)简况啮齿动物模型(布朗挪威鼠)。
布朗挪威老鼠敏感卵子和挑战三次每隔5天唤起气道重塑。表皮生长因子(EGF)的影响受体抑制剂,AG1478,和半胱氨酰leukotriene-1受体拮抗剂,montelukast、上皮和气道平滑肌细胞增殖(ASM)在活的有机体内针对重复卵子挑战测试。三个挑战与白三烯(LT) D4,对改造和检查他们的影响没有AG1478预处理。
卵子挑战ASM增生引起的,与质量的增加,上皮细胞增生,杯状细胞增殖。AG1478阻止变化,montelukast也是如此。多个卵子挑战增加heparin-binding EGF-like气道上皮生长因子但不是EGF的表达。有限公司4再现了改造改变诱导卵子,这被AG1478。
Allergen-induced气道上皮细胞和气管平滑肌改造是由半胱氨酰白三烯等通过与下游的半胱氨酰leukotriene-1受体对表皮生长因子受体轴的影响。
哮喘气道通常显示广泛而复杂的改造1- - - - - -4。平滑肌有潜力增长的最重要的病理生理后果通过过度气道狭窄和气道高反应性5,6。增加气道平滑肌(ASM)航空公司已经与哮喘的严重程度有关3,7目前,当超过大型航空公司与死亡率相关8。众所周知,平滑肌增生的动物模型9- - - - - -11增生和肥大在人体呼吸道标本12,13为ASM质量的增加作出贡献。还提出,牙龈myofibroblasts可能增加组织迁移质量7。
肌肉的增长反应的机制很不确定,虽然在人类生长因子表达的一些描述性研究气道组织已报告14- - - - - -16和许多生长因子已被证明对ASM在文化促有丝分裂的影响17- - - - - -20.。半胱氨酰(半胱氨酸)白细胞三烯(LTs)已知参与allergen-induced ASM细胞增殖在活的有机体内21,22但在体外这些物质是ASM弱的有丝分裂原23,24。敏感的鼠标,cys-LT1受体(cys-LT1R)对抗阻止ASM增厚的增加经过反复过敏原的挑战25,26。有可能是cys-LTs的影响在活的有机体内是间接的和由改变的表达或其余酪氨酸kinase-linked生长因子的影响,如表皮生长因子(EGF)27,或者通过上游影响免疫反应的过敏原的挑战28。
表皮生长因子已被证明是调节人类哮喘气道16和刺激ASM增长在体外23。Heparin-binding EGF-like生长因子(HB-EGF)、表皮生长因子受体(EGFR)的另一个配体也被报道在哮喘气道上皮的科目29日对血管平滑肌,促有丝分裂的30.,31日。因此,这些因素都是似是而非的候选人在哮喘ASM增生。EGFR被卷入杯状细胞分化在过敏性哮喘小鼠模型26。表皮生长因子受体的刺激引起的杯状细胞分化和upregulation粘蛋白基因32。当前作者提出,过敏原挑战增加了ASM的质量,通过表皮生长因子受体激活气道杯状细胞的数量在活的有机体内。为了验证这个假说,使用allergen-induced气道重构的大鼠模型9- - - - - -11,只需要三个过敏原暴露增加ASM大规模发生33。目前作者想阐明的可能作用表皮生长因子受体在ASM cys-LTs的明显的促有丝分裂的影响21,22在气道上皮改造26,报告说,抑制表皮生长因子受体或cys-LT1R完全废除allergen-induced ASM增长和上皮杯状细胞分化。外生有限公司4模仿过敏原的挑战导致ASM和杯状细胞增生的机制包括表皮生长因子受体,表明cys-LT1R是上游的表皮生长因子受体在改造级联。
材料和方法
动物准备和治疗
布朗挪威(BN)注射的老鼠敏感1毫克的卵清蛋白(OVA)和100毫克的明矾皮下注射2×109heat-killed百日咳博德特氏菌杆菌i.p。犀牛老鼠挑战aerosolised卵子或盐水气管内交付14天,19 - 24。的影响AG1478(表皮生长因子受体酪氨酸激酶抑制剂)或montelukast (cys-LT1R拮抗剂),或适当的车辆,因为i.p。或强饲法分别1 h每个挑战之前在独立样本进行测试。在第三个示例中,unsensitised老鼠管理有限公司4每隔5天(100μg)气管内的三倍,有或没有与AG1478预处理。这些组织与vehicle-treated控制。动物被杀后2天最后的挑战,组织收集。
研究协议是麦吉尔大学动物保健委员会批准(加拿大蒙特利尔,QC)。
疣状HB-EGF, EGF和EGFR
左肺formalin-fixed在25而言不啻2O压力和随后石蜡包埋。Mid-sagittal和5µm厚para-hilar部分被剪掉了。本土化和量化HB-EGF EGF蛋白表达在气道,疣状与兔多克隆抗体对人类HB-EGF(研发系统,Inc .,明尼阿波利斯,美国)和大鼠表皮生长因子(生物医学技术,Inc .)、斯托顿,妈,美国)。表皮生长因子受体是应用使用兔多克隆抗体免疫球蛋白(Ig) G(圣克鲁斯生物技术有限公司、圣克鲁斯、钙、美国)。消极的控制运行使用相同的同形像的浓度控制免疫球蛋白(研发系统,Inc .)每个主要的抗体。信号与向量®开发红色(向量实验室,伯林盖姆、钙、美国),紧随其后的是甲基绿(Sigma-Aldrich有限公司,圣路易斯,密苏里州,美国)对比染色。为了执行气道细胞的数量的形态学分析阳性HB-EGF EGF,剂量反应曲线的应用组织执行初级抗体和抗体的浓度产生明确的选择但弱阳性染色控制组织,为了允许检测免疫染色的增加,如果存在。免疫反应性的上皮细胞的数量统计的一个观察者(m . Hassan)失明组状态,和结果表示每毫米的周边基底膜(PBM)。
ASM的测量质量
ASM的质量估计的面积测量的光滑阳性α-actin(α-SMA)免疫反应性(如前所述)33。短暂,部分沾α-SMA小鼠单克隆抗体(克隆1 a4;Sigma-Aldrich有限公司)和生物素化的马anti-mouse免疫球蛋白,老鼠吸附(向量实验室)。信号检测与向量®红色。ASM的面积是追踪使用相机lucida侧臂附件显微镜和数字化。ASM的质量标准化了ASM的面积除以气道大小的平方PBM。
细胞增殖ASM的量化在活的有机体内
ASM细胞扩散研究使用双疣状增殖细胞核抗原(PCNA)和α-SMA使用PCNA单克隆抗体(美国Calbiochem,圣地亚哥,CA)如前所述33。染色之前是高温抗原决定部位暴露在抗原检索解决方案(向量实验室)和permeabilisation 0.2% Triton x - 100 (Sigma-Aldrich有限公司)。部分被堵塞20%马血清(向量实验室)普遍屏蔽解决方案(Dako Cytomation,斯特鲁普,丹麦),和一个单克隆抗体与生物素化的马anti-mouse PCNA检测免疫球蛋白,亲和素/ biotin-alkaline磷酸酶复杂,5-bromo-4-chloro-3-indolyl磷/氮蓝四唑发色体衬底(向量实验室)。双重免疫染色anti-α-SMA单克隆抗体与向量®开发红色和紧随其后的是甲基绿(Sigma-Aldrich有限公司)复染色。PCNA-positive ASM细胞的数量清点,纠正了气道大小的平方除以PBM。
量化的上皮改造
上皮细胞的营业额是决定使用PCNA染色。列举了杯状细胞周期acid-Schiff (PAS)染色组织和表示为每毫米P PAS-positive细胞的数量BM。
实时定量PCR对肺癌和呼吸道上皮细胞生长因子
调查的mRNA表达HB-EGF EGF在整个肺和呼吸道上皮细胞治疗后,实时定量PCR进行。正确的肺在液氮快速冻结,保持在-80°C。总RNA提取使用试剂盒从冷冻肺试剂(美国表达载体,卡尔斯巴德,CA)根据制造商的指示。气管被切割出来,沉浸在RNAlater一夜之间(试剂盒、希尔登,德国)和上皮细胞通过刮的腔气管细胞刮刀。从上皮细胞总RNA提取使用RNeasy迷你包(试剂盒)根据制造商的指示。逆转录执行使用上标二世逆转录酶(表达载体)。实时定量PCR与LightCycler执行®瑞士巴塞尔(罗氏)使用DNA主人SYBR绿色我由于“快速上手”项目(罗氏)。HB-EGF引物的序列是如前所述34。引物对大鼠表皮生长因子、表皮生长因子受体和核糖体蛋白S9使用基于网络的软件设计,PrimerQuest(集成DNA技术,鲁汶,比利时)。每一对引物的序列如下。HB-EGF: 5′-ACTTGGAAGGGACCGATCTGGA-3′, 5′-TAGGGTCAGCCCATGACACCTC-3′;EGF: 5′-TGCCTTGCCCTGACTCTAC-3′, 5′-AGCCAATGACACAGTTGCAC-3′;表皮生长因子受体:5′-TCCCTTTGGAGAACCTGCAGATCA-3′, 5′-GTTGCTAAATCGCACAGCACCGAT-3′;S9: 5′-AGGATTTCTTGGAGAGAAGGCTGC-3′, 5′-CTTCTGAGAGTCCAGGCGAACAAT-3′。建立了标准曲线为每个生长因子和看家基因。简而言之,PCR产品从琼脂糖凝胶中提取和纯化GFX PCR和DNA凝胶乐队净化设备(Amersham生物科学(通用电气医疗集团生命科学),皮斯卡塔韦,新泽西,美国)。使用微PCR产物的数量计算。 By step dilution, 101-10年10标准的副本。
有限公司的影响4、EGF和HB-EGF鼠ASM的主要文化细胞的扩散
评估ASM的可能性增长后重复卵子挑战涉及EGFR和互动有限公司4的平滑肌本身,EGF和HB-EGF的影响,结合有限公司4是培养ASM上测试过。大鼠气管被切割和多余的结缔组织35。组织在弹性蛋白酶消化在37°C为30分钟,然后放在冰停止反应。组织被允许站和上层的收集。组织碎片留在管洗了汉克斯平衡盐溶液(哈佛商学院)和上层的收集。这个过程重复了三次。最后,收集的细胞在1400转离心5分钟,resuspended 25厘米2烧瓶1:1杜尔贝科的修改鹰介质(DMEM):火腿的F12培养基补充10%胎牛血清的边后卫,NaHCO 0.224%35%和1%的青霉素、链霉素的有限公司2。细胞培养试剂购买从GIBCO(表达载体)。表型确认如前所述35。
在章节2 - 4细胞用于实验。细胞被镀上6-well板块(40000细胞2毫升介质/)和饥饿DMEM 0.2%牛血清白蛋白(BSA)当他们到达confluency∼70%。与0.2% BSA,细胞在DMEM 48 h后刺激了人类HB-EGF (1 - 10 ng·毫升−1;研发系统,Inc .)有限公司4(1)μM;Biomol国际普利茅斯会议上,美国宾夕法尼亚州)或HB-EGF和有限公司4。在一个单独的一系列实验,细胞使用montelukast(500海里;Merck-Frosst加拿大,加拿大蒙特利尔)管理有限公司前30分钟4。细胞与哈佛商学院含有0.25%胰蛋白酶和0.02% EDTA trypsinised分离板。胰蛋白酶是通过添加DMEM中和:火腿的F12 10%的边后卫,离心法收集的细胞在1400 rpm的5分钟和resuspended 1毫升的PBS。这些细胞被使用血球计数器计数。溴脱氧尿苷(BrdU)合并分析,细胞被饿死,如上所述,BrdU被添加到每个刺激刺激后6 h。细胞收获后的18 h BrdU(刺激后24 h)和处理流仪检测事务局BrdU使用荧光素isothiocyanate-BrdU流工具包(BD生物科学,米西索加、加拿大)根据制造商的指示。
有限公司的影响4在细胞内钙培养ASM细胞的反应
演示的功能cys-LT ASM受体,细胞内钙的变化以应对投资有限公司4(1μM)测量。胞质钙进行测量(如前所述)36ASM细胞孵化为30分钟37°C·汉克斯的缓冲区(NaHCO氯化钠137毫米34.2毫米,葡萄糖10毫米,Na2HPO43毫米,氯化钾5.4毫米,KH2阿宝40.4毫米,CaCl21.3毫米,MgCl20.5毫米,MgSO40.8毫米,羟乙基哌嗪乙磺酸5毫米)的5μM Fura-2-acetoxymethylester。加载的细胞被洗了,封面都放置在一个莱顿室(美国医疗系统集团,格林维尔,纽约)包含450μL汉克斯缓冲区在舞台上倒置显微镜的细胞成像装备40×油浸物镜(尼康,东京,日本)。细胞刺激前后的图像有限公司4获得使用一个加剧相机(视频示波器IC 200;光子技术International Inc .,伦敦,加拿大)和PTI软件(光子技术International Inc .)在一个发射波长(510海里)双兴奋波长(345和380海里)。荧光比例(345/380)测定单个细胞和自由钙使用Grynkiewicz公式计算37。
统计分析
数据意味着±扫描电镜。单向方差分析之后,未配对的双尾t检验进行了分析。当数据不是正态分布,日志转换应用。Bonferroni调整用于正确的多重比较。p < 0.05的值被认为是重要的。
结果
过敏原的挑战对EGF和HB-EGF mRNA表达的影响在整个肺和呼吸道上皮细胞
卵巢敏化作用的影响和挑战的mRNA的表达两个表皮生长因子受体配体,EGF和HB-EGF,以及表皮生长因子受体,接受了检查,他们的表情是使用管家基因S9标准化。这是证实S9学习小组之间没有显著变化,此外,是高度相关的其他两个管家基因的表达水平,还有β-actin和(数据没有显示)。这些生长因子量化的表达在整个肺组织利用mRNA的肺匀浆和上皮细胞收获从气管刮。HB-EGF mRNA表达多个卵子后气管上皮细胞是调节双重挑战挑战细胞(0.0014±0.00029与0.00073±0.000073控制;p = 0.035;图1一个⇓mRNA),而EGF和EGFR没有显示变化(表皮生长因子:0.013±0.0011与对挑战细胞0.014±0.0016与分别控制,p = 0.51;表皮生长因子受体:0.013±0.0012与0.015±0.0016,p = 0.43;图1 b⇓和c)。没有可检测变更任何三个基因的表达在整个肺组织(图1 d-f⇓)。
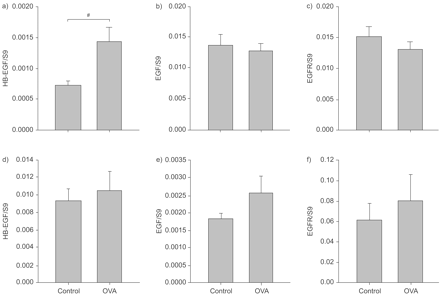
Heparin-binding表皮生长因子(EGF)——如生长因子(HB-EGF;a和d), EGF (b和e)和表皮生长因子受体(EGFR);c、f) mRNA表达在气道上皮细胞(a - c)和全肺组织(d-f)经过反复卵白蛋白(OVA)的挑战。mRNA的表达HB-EGF、EGF和EGFR被实时定量PCR和测量标准化的看家基因S9气道上皮细胞获得从气管腔的表面刮后连续三个卵子挑战每隔5天。卵子挑战的气溶胶交付5%卵子解决气管内插管犀牛老鼠。数据意味着±扫描电镜;n = 8。#:p = 0.035。
Immunolocalisation和地貌形态示量评估HB-EGF EGF表达后卵子的挑战
疣状进行本土化网站HB-EGF和EGF的表达的航空公司。图2⇓显示了免疫反应性的显微照片和数据HB-EGF和EGF。HB-EGF表达主要在气道上皮细胞和形态学评估HB-EGF的免疫反应性表现出显著增加OVA-challenged动物细胞(110.7±4.15毫米−1PBM细胞)与控制(28.2±4.91毫米−1PBM;p < 0.0001;图2一个⇓,c和e)。在气道上皮细胞EGF表达强烈和更少的强烈ASM细胞和肺泡细胞。有证据的组成型表达EGF但没有区别的免疫反应性的上皮细胞之间的细胞治疗组(36.32±3.23毫米−1在OVA-challenged动物与35.88±2.86控制;p = 0.92;图2 b⇓,d和f)。这些观测结果符合定量实时PCR信使rna从气管上皮细胞(图1 a - c⇑)。
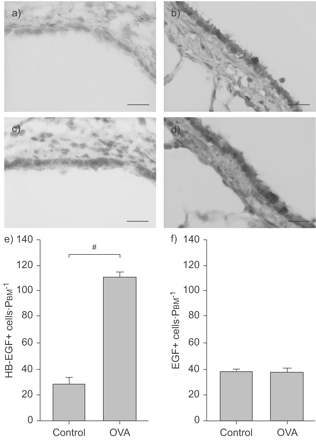
多个卵清蛋白(OVA)挑战增加heparin-binding表皮生长因子(EGF)——如生长因子(HB-EGF)蛋白表达在气道上皮细胞。本地化和量化HB-EGF (a和c)和表皮生长因子(b和d)在蛋白质水平进行免疫染色和分析morphometrically生理盐水对照组(a和b)和OVA-challenged老鼠(c和d)。免疫反应性的HB-EGF检测主要是在气道上皮细胞和增加OVA-challenged动物(a和c)。在气道上皮细胞EGF表达强烈和更少的强烈在气道平滑肌细胞和肺泡细胞(b和d)。HB-EGF-positive (e)和EGF-positive (f)气道上皮细胞数和纠正使用周边的基底膜(PBM)。HB-EGF-positive上皮细胞增加OVA-challenged组的3.5倍。没有卵子后EGF蛋白表达变化的挑战。数据意味着±扫描电镜;n = 6。酒吧= 50μm规模。#:p < 0.0001。
表皮生长因子受体的影响封锁allergen-induced ASM改造:ASM的质量
EGFR抑制的影响在ASM研究OVA-induced增加。收缩组织被免疫组织化学鉴定α-SMA,如图3所示⇓(a和b)。ASM的质量决定morphometrically和表达为一个区域组织纠正气道大小(ASM·PBM−2;图3 f⇓)。ASM的size-corrected区域卵子吸入后会增加2.8倍的挑战(0.0020±0.0004)相比saline-challenged控件(0.0007±0.0002;p = 0.0007)。ASM的增加质量显著抑制AG1478治疗,一个EGFR-specific酪氨酸激酶抑制剂(0.0009±0.0002;p = 0.0011)。
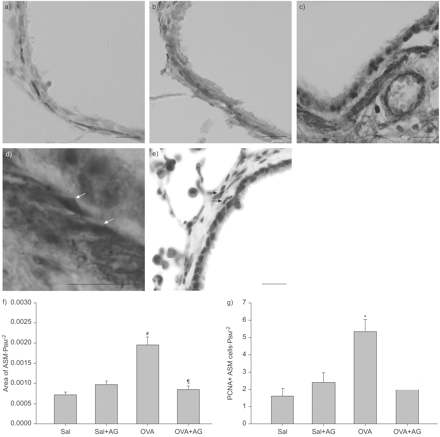
表皮生长因子受体(EGFR)酪氨酸激酶抑制剂AG1478 (AG)阻塞气管平滑肌(ASM)扩散引起的多个卵清蛋白(OVA)的挑战。代表显微照片光滑阳性α-actin(α-SMA)盐水的气道组织化学染色控制(a)和多个OVA-challenged组(b),显示ASM束红色。c和d)双疣状增殖细胞核抗原(PCNA)和α-SMA。核染色深紫色代表PCNA-positive ASM细胞(箭头)和上皮细胞。e)说明性的显微照片的上皮细胞表皮生长因子受体免疫反应性,在牙龈细胞纺锤状核可能是ASM细胞和肺泡巨噬细胞(箭头)。ASM的f)的形态学测量质量。ASM是标准化的平方的面积周长的基底膜(PBM),是更大的在OVA-challenged组(n = 9)与生理盐水相比控制(Sal;n = 6)和减少AG)治疗(n = 6)。g) PCNA-positive ASM细胞增加3.4倍卵子组(n = 9)和AG)显著增加。AG(5毫克公斤−1在二甲亚砜)或车辆是由i.p。注射1 h之前每个挑战。数据意味着±扫描电镜。酒吧= 50μm规模。#:p = 0.004与萨尔控制;¶:p = 0.016与卵巢组织;+:p = 0.001与萨尔控制;§:p = 0.01与卵巢组织。
表皮生长因子受体的影响封锁allergen-induced ASM细胞的扩散
为了确认ASM细胞发生增生性的增长,PCNA-positive核immunolocalisedα-SMA-positive细胞;一个说明性的例子如图3 c⇑和d。增殖ASM细胞的数量被PCNA染色高出3.4倍OVA-challenged动物细胞(5.4±0.69毫米−2)与生理盐水对照相比(1.6±0.46;p = 0.001;图3 g⇑),证实了ASM扩散对过敏原的挑战。表皮生长因子受体表达在上皮细胞和ASM,以及肺泡巨噬细胞。在牙龈上皮的表达EGFR和表达细胞位于平滑肌层如图3所示⇑。AG1478阻塞后细胞增殖ASM的增加多个卵子挑战(p = 0.01)。
表皮生长因子受体的影响封锁allergen-induced气道上皮改造
过敏性气道重构包括气道上皮细胞表型的变化,最明显的是杯状细胞的增加。杯状细胞的增加由过敏原诱发挑战如图4所示⇓(a和b)。量化PAS-positive细胞显示增加8倍后卵子挑战(p = 0.004;图4摄氏度⇓)。增加被AG1478显著降低(p = 0.006)。增殖气道上皮细胞的数量,确定PCNA染色,也增加了多种过敏原后挑战·mm(17.4±1.87细胞−1;p = 0.001;图4 d⇓)。这一增长也被AG1478 (9.1±2.1;p = 0.017)。
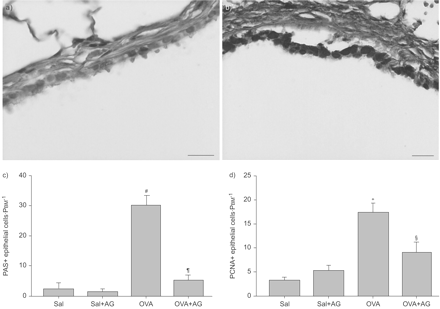
表皮生长因子受体酪氨酸激酶抑制剂AG1478 (AG)阻塞气道上皮改造引起的多个卵清蛋白(OVA)的挑战。代表显微照片的周期性acid-Schiff (PAS)染色从盐水(Sal)控制动物(a)和繁殖OVA-challenged动物(b),显示增加OVA-challenged动物的杯状细胞。c) PAS-positive上皮细胞数和纠正使用周边的基底膜(PBM)。他们在多个卵子挑战增加了8倍。AG)减少PAS-positive细胞诱导由多个卵子的挑战。d)多个卵子挑战引起气道上皮细胞增殖检测增殖细胞核抗原(PCNA)。这是被AG)。数据意味着±扫描电镜;n = 6。酒吧= 50μm规模。#:p = 0.0042与萨尔控制;¶:p = 0.0056与卵巢组织;+:p = 0.0013与萨尔控制;§:p = 0.017与卵巢组织。
影响cys-LT1R拮抗剂montelukast ASM多个卵子质量和增生的挑战
为了证实cys-LT的角色1R在气道重塑,反复过敏原的挑战是在一个单独的执行一系列的动物,其中一些使用montelukast,特定的cys-LT1R拮抗剂。的形态学分析OVA-induced ASM执行质量的变化,有或没有montelukast预处理。和之前一样,ASM质量增加OVA-challenged老鼠(0.0015±0.00014;p = 0.04),而与saline-challenged控制动物(0.0011±0.00016),和ASM OVA-induced增加质量预防montelukast预处理(0.0010±0.0001;p = 0.01),这表明它是依赖cys-LT1r . PCNA-positive ASM OVA-challenged老鼠细胞的数量,确定morphometrically,卵子后也增加了挑战和显著减少montelukast(图5所示⇓)。
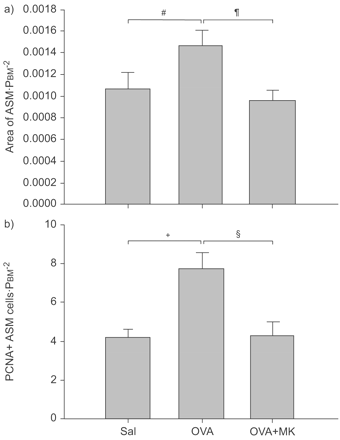
气管平滑肌(ASM)质量的增加经过多次卵白蛋白(OVA)挑战被cysteinyl-leukotriene-1受体拮抗剂抑制montelukast(可)。ASM的质量和增殖细胞核抗原(PCNA)阳性ASM细胞被标准化的广场周边的基底膜(PBM)。OVA-challenged组(n = 7)有显著提高ASM质量与生理盐水相比控制(Sal;n = 9)。ASM是减少OVA-challenged MK-treated组(n = 9)与OVA-challenged组。b) PCNA-positive ASM细胞增加1.9倍后卵子的挑战。这个增加是由可预防。可注射一剂3毫克公斤−1在甲基纤维素填喂法1 h之前挑战。数据意味着±扫描电镜。#:p = 0.041;¶:p = 0.036;+:p = 0.0059;§:p = 0.016。
重复的影响有限4挑战ASM增长和杯状细胞增殖
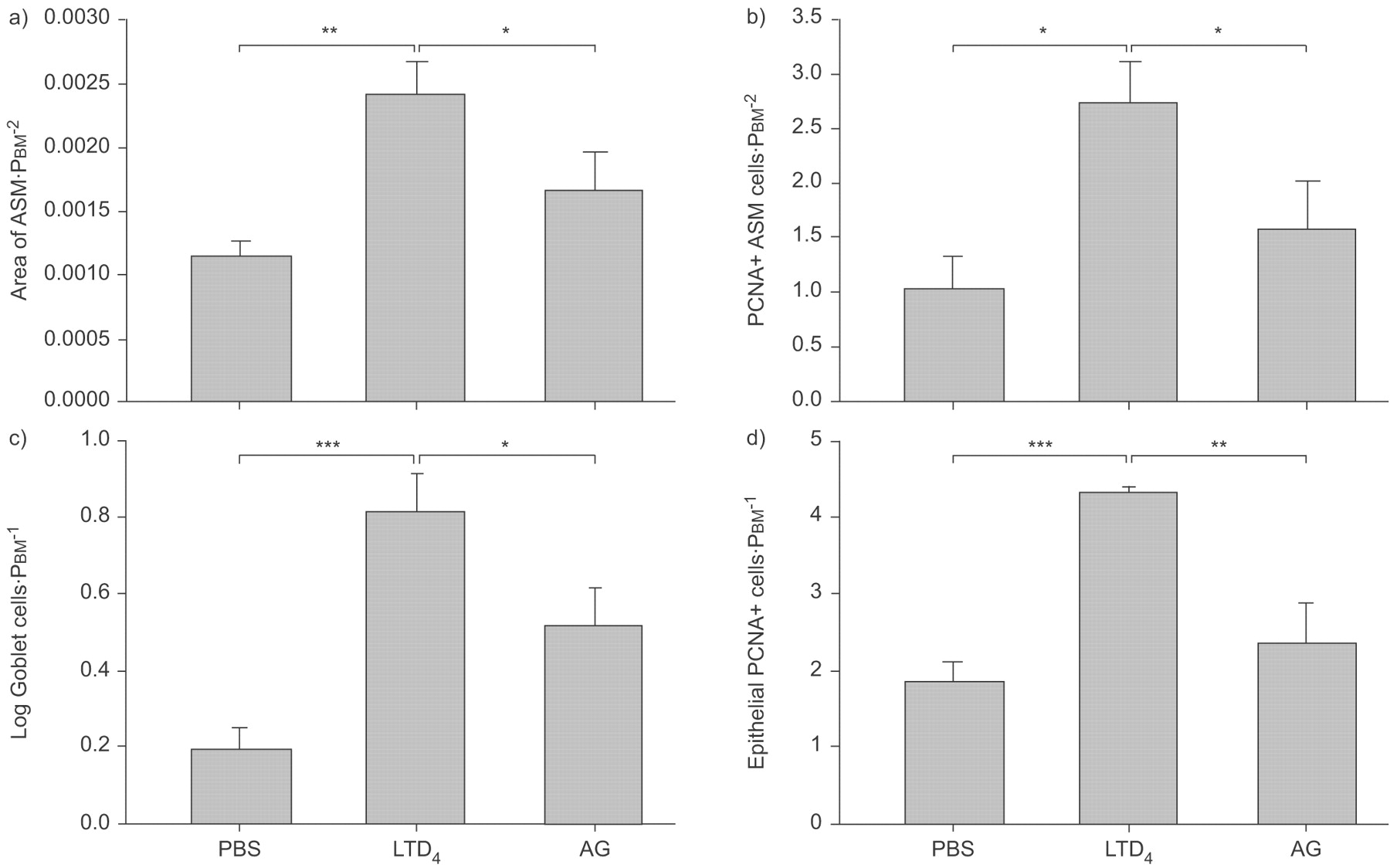
检查潜在的有限公司4单独诱导气道重构在天真的老鼠,动物暴露在有限公司4三次每隔5天,模仿重复卵子的挑战。有明显的改造ASM和杯状细胞在这些动物;ASM增加超过两倍和杯状细胞数量增加了大约四倍(图6所示⇓)。
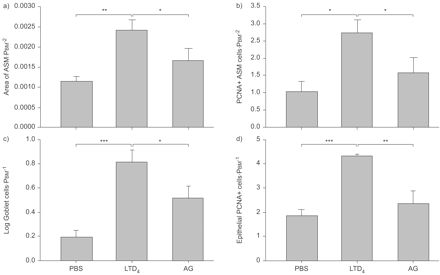
重复白三烯(LT) D4灌注物引起气道平滑肌(ASM)增长和杯状细胞增殖。ASM地区由形态测量学),增加了三个气管内的灌注物1μg有限公司4100年μL PBS。增加被AG1478 (AG)。b)增殖细胞核抗原(PCNA)免疫反应性的细胞局部双免疫染色ASM的光滑阳性α-actin。有限公司后细胞的数量增加4这增加被AG)。c)有限公司4杯状细胞数量增加评估的周期性acid-Schiff染色。杯状细胞被AG)的增加。d)有增加上皮细胞PCNA-immunoreactive有限公司后4。AG阻塞PCNA-positive上皮细胞的增加。PBM:基底膜的周边。*:p < 0.05;* *:p < 0.01;* * *:p < 0.001。
抑制表皮生长因子受体在有限公司的影响4全身的气道重塑
评估公司的可能性4是通过表皮生长因子受体大鼠进行重复有限公司吗4挑战是用AG1478预处理,在同一剂量,有效预防allergen-induced ASM和上皮改造。表皮生长因子受体的抑制与AG1478显著降低ASM和杯状细胞增生,增殖细胞核抗原免疫反应性评估(图。6⇑)。杯状细胞数量和ASM面积比例减少。
有限公司后的形态学评估上皮HB-EGF表达式4挑战
重复挑战公司的影响4在上皮表达HB-EGF morphometrically评估。有限公司的表达显著增加4挑战动物与控制(图7所示⇓)。效果是完全由montelukast抑制,表明它是由cys-LT1R。
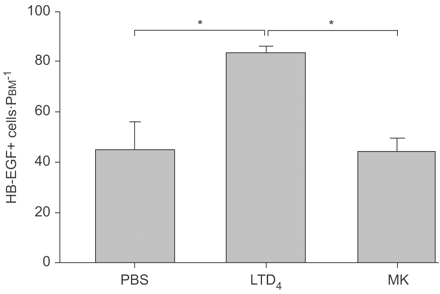
影响白三烯(LT) D4挑战上皮heparin-binding表皮生长因子生长因子(HB-EGF)免疫反应性。有限公司4每隔5天是由气管内的三次滴注法和48 h后组织的第三个挑战是收获进行分析。HB-EGF-expressing细胞的数量增加了近两倍与PBS控制相比,这增加显著抑制montelukast(可)。PBM:基底膜的周边。*:p < 0.05。
有限公司的影响4在ASM扩散在体外单独和伴随的表皮生长因子受体刺激
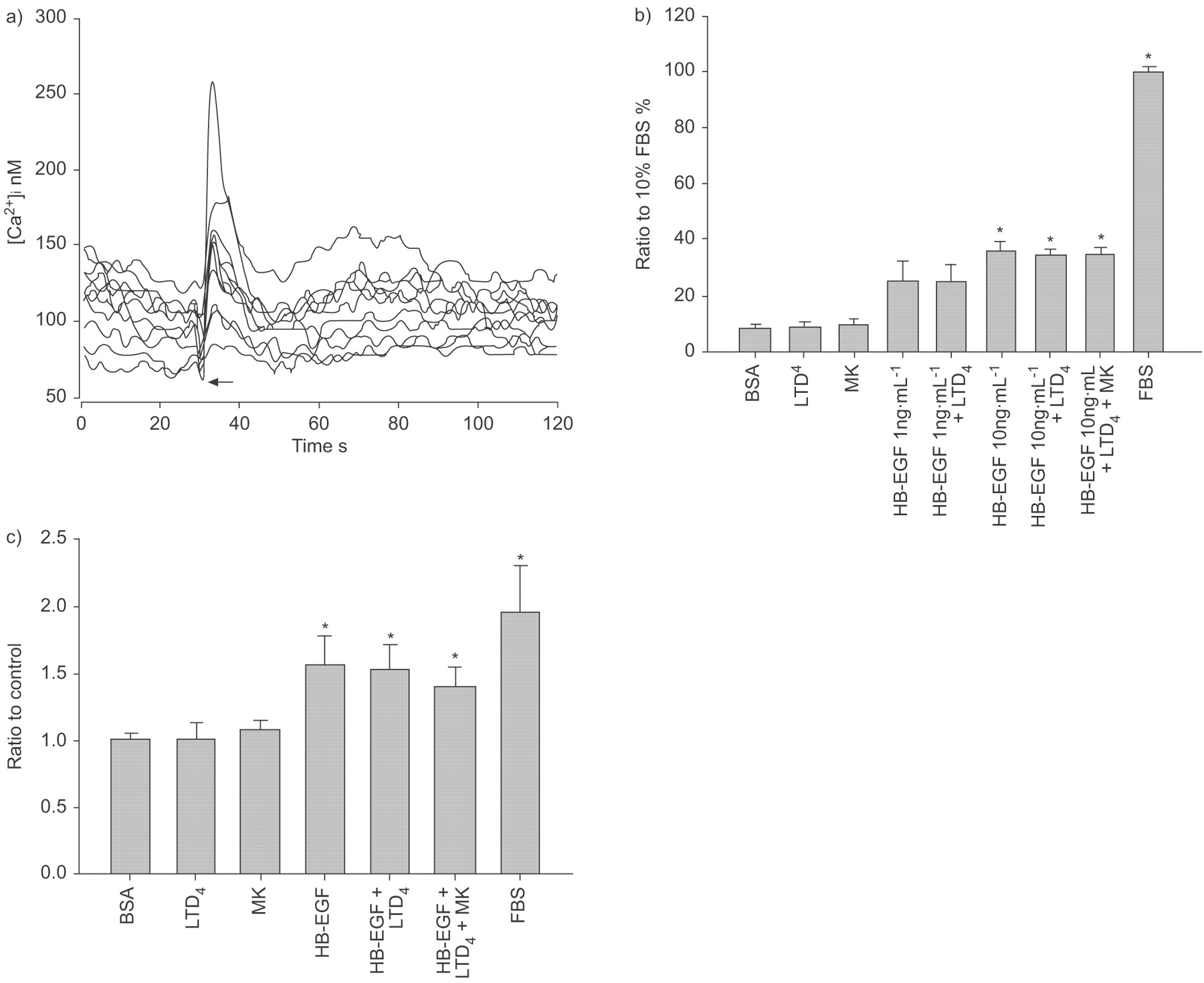
ASM本身是一个潜在的公司网站的交互4表皮生长因子受体轴,通过协同作用或transactivation EGFR。这是确认功能cys-LT受体存在于ASM细胞通过展示上升细胞胞内钙的反应接触有限公司4(1)μM;图8⇓)。测试cys-LTs协同行动的可能性与表皮生长因子受体在促进平滑肌细胞生长,仅HB-EGF和EGF的促有丝分裂的影响,结合有限公司4在ASM BN大鼠在初级文化进行检测时,发现使用BrdU合并和细胞计数。HB-EGF 10 ng·毫升−1刺激ASM显著增殖,通过衡量BrdU合并(图8 b⇓)和细胞计数(图8 c⇓)。外生有限公司4没有增加BrdU合并或细胞数量超出独自HB-EGF后观察到的值。表皮生长因子和表皮生长因子+有限公司也获得了类似的调查结果4(数据没有显示)。AG1478应对EGF和HB-EGF抑制了增长,然而它并没有减少对血小板源生长因子或multi-mitogenic刺激的边后卫(10%;数据未显示)。
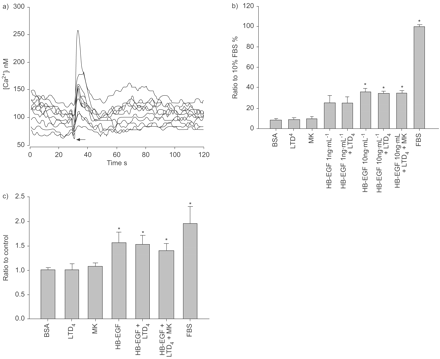
Heparin-binding表皮生长因子生长因子(HB-EGF)促有丝分裂的布朗挪威大鼠气道平滑肌细胞(ASM)在体外。)一个代表性的例子ASM细胞钙反应1µM白三烯(LT) D4。每一行代表一个细胞的钙瞬变。受体激动剂应用程序用一个箭头表示。b)溴脱氧尿苷(BrdU)结合流式细胞仪测量,与BrdU-incorporating细胞的数量表达的细胞治疗的比例在10%胎牛血清的边后卫。细胞治疗在HB-EGF (10 ng·毫升−1)单独和HB-EGF (10 ng·毫升−1)+4(1μM)显示增加到了原来的4倍BrdU公司相比之下,0.2%的牛血清白蛋白(BSA)的控制。有限公司4并没有改变BrdU合并或影响HB-EGF的促有丝分裂的影响。Montelukast(可;500海里)并不影响BrdU合并。c)细胞计数和细胞的数量的比率,表示为控制(0.2% BSA)。在协议BrdU整合研究,HB-EGF (10 ng·毫升−1)单独细胞数量增加1.5倍,而无论是有限公司4(1μM)也可(500海里)细胞数量的影响。数据意味着±扫描电镜;n = 3。*:p < 0.05与BSA控制。
讨论
反复过敏原挑战积极敏感BN大鼠增生引起的ASM导致大幅提高ASM质量。它也引起气道上皮细胞和杯状细胞的增殖分化。生长因子的增加表达HB-EGF气道上皮细胞,但不是EGF,多次发生卵子的挑战。ASM增生,杯状细胞分化是由预处理预防选择性表皮生长因子受体酪氨酸激酶抑制剂,表明EGFR在这些组件的主要角色allergen-induced气道重塑。Montelukast也封锁allergen-induced ASM增生,确认cys-LT的角色1在这个过程中R。气管内的滴剂的有限公司4改造引起的类似于卵子挑战单独服用时天真的老鼠,而这些改造变化被AG1478抑制,表明有限公司4通过表皮生长因子受体介导改造通过间接手段。有限公司4没有检测到直接对老鼠ASM促有丝分裂的影响文化,不与表皮生长因子受体诱导ASM增长。
尽管许多生长因子有可能增加ASM增长,基于活动在体外作者,当前选择集中在表皮生长因子受体配体,EGF和HB-EGF,因为最近的证据表明,这些配体可能是调节人类哮喘组织29日。增加EGF的免疫反应性已被证明在哮喘的黏膜下层16在支气管上皮,腺体和平滑肌在人类哮喘气道29日。表皮生长因子受体及其配体的功能意义的航空哮喘受试者还不清楚。目前的数据阐明了表皮生长因子受体的作用和建议一个可能的候选人受体的配体allergen-induced改造的上皮和ASM在活的有机体内。ASM的抑制增长在活的有机体内由特定的表皮生长因子受体抑制剂AG1478几乎是完整和杯状细胞分化也减少到基底的水平。表皮生长因子受体已经被证明参与杯状细胞化生过敏原后挑战在ASM改造使用小鼠模型,但其作用没有任何详细地检查在活的有机体内26。目前的研究没有发现EGF和EGFR upregulation在mRNA水平在整个肺标本或孤立的上皮细胞,这表明监管扩散并不源于改变表皮生长因子受体或EGF水平本身。的upregulation HB-EGF提供另一种机制来解释观察到的表皮生长因子受体在改造的重要性虽然数据不排除选择表皮生长因子受体配体的作用。
表皮生长因子受体之间的关系和cys-LTs调停气道重构还不清楚。改造的抑制ASM的完整性和上皮细胞特定的表皮生长因子受体的拮抗剂和cys-LT1R提出一个可能的交互途径调解他们的影响。Cys-LTs过敏性气道狭窄的主要介质36- - - - - -41。目前的数据是一致的在改造他们的角色,在几个以前的研究证明21,22,25,26。当前的作者预期,抑制表皮生长因子受体可能降低公司的改造效果4本身,因为报道机械压力放在上皮的作用,比如将引起的支气管收缩释放HB-EGF从气道上皮细胞42。符合这些报道发现,抑制表皮生长因子受体的AG1478阻止有限公司4全身的改造。另外,重复曝光的航空有限公司4引起的upregulation HB-EGF,暗示的可能性,这种配体可能负责观察改造介导通过表皮生长因子受体。释放的HB-EGF有限公司的直接证据4需要证实这一假说。
的互动有限公司4和表皮生长因子受体在ASM本身是寻求。有限公司4已被证明导致培养的人类表皮生长因子受体transactivation ASM细胞43。人类成纤维细胞4还把与表皮生长因子受体,但不涉及受体transactivation机制27。相比人类ASM的数据23,43和成纤维细胞27,没有证据表明公司的促有丝分裂的影响4有限公司之间的交互也不是显而易见的4HB-EGF或EGF在老鼠ASM,尽管明显存在功能性cys-LT受体在ASM文化。老鼠和人类之间的差异的原因ASM细胞并不清楚。
可能,除了重装对表皮生长因子受体轴由下游的影响,可能存在影响cys-LTs发布后卵子在上游网站涉及炎症级联的挑战。的cys-LT1R已经涉及到辅助细胞的起源2型炎症通过对树突状细胞功能的影响28。符合cys-LTs的免疫调节作用,montelukast减少气道嗜酸性粒细胞和interleukin-5 allergen-challenged BN大鼠,表明涉及t细胞的抗炎作用44。
总之,目前的研究表明,表皮生长因子受体和cysteinyl-leukotriene-1受体是必要的一些特点,过敏原后气道重构的挑战。然而,cysteinyl-leukotriene-1受体是上游的表皮生长因子受体在改造的过程。表皮生长因子受体的配体可能是heparin-binding表皮生长因子生长因子,因为它是由过敏原和白三烯D调节4挑战。Cysteinyl-leukotrienes过敏原挑战可能发布的版本,反过来,heparin-binding表皮生长因子生长因子,从而介导上皮和气道平滑肌细胞改造。然而,是否发生这种现象的确是一个问题,需要进一步研究。
支持声明
m . Tamaoka拨款支持中心哮喘在工作场所由加拿大卫生研究院资助的研究。d . Ramos-Barbon是加拿大的接受者肺脏协会/ BI /辉瑞奖学金,,目前支持的国家西班牙卫生系统(洋底de Investigaciones疗养地,基金没有。CP04/00313)。
感兴趣的语句
声明对j。马丁和d Ramos-Barbon可以找到www.www.qdcxjkg.com/misc/statments.shtml
- 收到了2007年12月11日。
- 接受2008年7月14日。
- ©人期刊有限公司











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}