条文本gydF4y2Ba

摘要gydF4y2Ba
背景gydF4y2Ba囊性纤维化(CF)肺病的定义是气道中存在大量中性粒细胞和相关的损伤产物。CF中描述了延迟性中性粒细胞凋亡,尽管尚不清楚这是原发性中性粒细胞缺陷还是对慢性炎症的反应。在CF中检测到中性粒细胞细胞外陷阱(NETs)水平的增加,我们的目的是研究这些现象与其作为炎症驱动因素的潜力之间的因果关系。我们假设CF中细胞凋亡的延迟是一个主要缺陷,并优先允许CF中性粒细胞形成NETs,从而导致炎症。gydF4y2Ba
方法gydF4y2Ba从CF患者、CF猪和适当对照组中分离出血液中性粒细胞。在开始ivacaftor之前和之后,还从CF患者中获得了中性粒细胞。形态学和流式细胞仪检测细胞凋亡。通过荧光显微镜和DNA释放测定确定NET的形成。ELISA和qRT-PCR检测细胞因子的生成,检测NET与巨噬细胞的相互作用。gydF4y2Ba
结果gydF4y2Ba由于细胞凋亡减少,CF中性粒细胞寿命延长。囊性纤维化跨膜电导调节因子(CFTR)缺失仔猪和CF患者均观察到这种情况,并且在门控(G551D)突变患者中,ivacaftor (CFTR增强剂)可以逆转这种情况。CF中性粒细胞形成了更多的net,而暴露于周期蛋白依赖性激酶抑制剂可以逆转这一情况。NETs对巨噬细胞提供促炎刺激,在CF中增强。gydF4y2Ba
结论gydF4y2BaCF中性粒细胞具有促生存表型,这与CFTR功能的缺失有关,并允许增加NET的产生,这反过来又可以诱导炎症。增强CF中的中性粒细胞凋亡可能允许更适当的中性粒细胞处理,减少NET的形成,从而减少炎症。gydF4y2Ba
- 囊性纤维化gydF4y2Ba
- 中性粒细胞生物学gydF4y2Ba
这是一篇根据创作共用署188滚球软件名(CC BY 4.0)许可条款发布的开放获取文章,该许可允许其他人出于商业用途分发、混音、改编和构建此作品,前提是正确引用原始作品。看到的:gydF4y2Bahttp://creativecommons.org/licenses/by/4.0/gydF4y2Ba
数据来自Altmetric.comgydF4y2Ba
关键信息gydF4y2Ba
关键问题是什么?gydF4y2Ba
囊性纤维化(CF)中性粒细胞是否活得更长,并允许中性粒细胞细胞外陷阱(NET)优先形成?gydF4y2Ba
底线是什么?gydF4y2Ba
CF中性粒细胞具有原发性缺陷,导致自发凋亡减少,并允许NET形成水平增加,从而促进炎症。gydF4y2Ba
为什么要读下去呢?gydF4y2Ba
这项工作将中性粒细胞凋亡的基本囊性纤维化跨膜电导调节相关缺陷与NETs促进CF中的炎症联系起来,并证明增强中性粒细胞凋亡可能是减少CF中NET形成和炎症的治疗策略。gydF4y2Ba
简介gydF4y2Ba
囊性纤维化(CF)是高加索人群中最常见的致命单基因疾病,患病率为1 / 2000,是一种影响肺、胰腺、汗腺、肠道、肝脏和肾脏的多器官疾病。CF中下呼吸道病原体感染的炎症反应被夸大,大多数患者死于肺部疾病。gydF4y2Ba1gydF4y2BaCF肺病的病理生理学尚不清楚,但重要因素包括细菌杀灭受损、粘液清除减少和过度炎症。gydF4y2Ba2gydF4y2Ba中性粒细胞是CF肺中的关键炎症细胞,其作用与专业吞噬细胞一样,但它们与肺中其他免疫细胞的相互作用可能同样重要,需要进一步研究。gydF4y2Ba
中性粒细胞自发凋亡的能力对宿主具有保护作用,对解决感染性或炎性损伤至关重要。gydF4y2Ba3.gydF4y2Ba一些研究表明,细胞凋亡缺陷导致CF中中性粒细胞存活增加,gydF4y2Ba4个5gydF4y2Ba但其后果尚不清楚。在没有自发凋亡的情况下,中性粒细胞处理的其他机制,如中性粒细胞细胞外陷阱(NET)形成(NETosis)可能变得越来越重要。NETosis被认为是中性粒细胞杀死细菌的另一种机制,除吞噬体杀死外还起作用。gydF4y2Ba6 7gydF4y2BaNETosis发生在受刺激的中性粒细胞(例如,由白介素(IL)-8、脂多糖(LPS)、肉豆蔻酸乙酸磷酯(PMA)或细菌引起)中,这些中性粒细胞经历氧化爆发并释放出由DNA、组蛋白、抗菌蛋白和潜在的促炎蛋白组成的网络。net在CF气道中过量,gydF4y2Ba8 9gydF4y2Ba这可能是由于CF肺的产量增加或清除率减少。NETs还与关节炎、系统性红斑狼疮(SLE)和痛风等炎症疾病有关,gydF4y2Ba10 - 12gydF4y2Ba最近,NETs已被证明与巨噬细胞相互作用并引发血管疾病中的炎症。gydF4y2Ba13gydF4y2Ba因此,NETs与巨噬细胞的相互作用作为炎症的驱动因素可能在CF中特别相关,其中巨噬细胞对炎症刺激有过度夸张的反应。gydF4y2Ba14gydF4y2Ba
我们假设,由于细胞凋亡的减少,CF中的中性粒细胞存活率构成性增加,允许更多的中性粒细胞形成NETs,与巨噬细胞相互作用并促进炎症。gydF4y2Ba
方法gydF4y2Ba
CF患者样本的收集gydF4y2Ba
从参加苏格兰国家CF服务的稳定CF患者中采集外周血。如果患者在过去2周内不需要静脉注射抗生素,则认为病情稳定。肺移植患者被排除在外。患者给予了书面同意,研究得到了区域伦理委员会的批准(苏格兰东部研究伦理委员会,15/ES/0094,苏格兰西部研究伦理委员会,11/WS/0074)。在当地招募了匿名匹配的健康对照组(洛锡安研究伦理委员会,08/S1103/38)。都柏林圣文森特大学医院研究和伦理委员会批准了ivacaftor治疗前后患者的样本收集。gydF4y2Ba
人中性粒细胞和外周血单个核细胞的分离gydF4y2Ba
取人外周血加入3.8%柠檬酸钠。全血350倍离心后抽吸血浆gydF4y2BaggydF4y2Ba20分钟。多形核细胞和外周血单个核细胞(PBMC)经6%右旋糖酐沉淀分离,经不连续(72.9、63.0%和49.5%)Percoll (GE Healthcare, Buckinghamshire, UK)梯度分离。gydF4y2Ba15gydF4y2Ba分离的细胞在无阳离子的Dulbecco’s磷酸盐缓冲盐水(DPBS)中洗涤两次gydF4y2Ba-/-gydF4y2Ba),然后在适当的培养基中重新悬挂。在一些实验中(参见“结果”部分),使用Ficoll-Paque (GE Healthcare)右旋糖酐沉淀和低渗裂解残余红细胞分离中性粒细胞。gydF4y2Ba
细胞活力和凋亡测定gydF4y2Ba
分离的中性粒细胞(5×10gydF4y2Ba6gydF4y2Ba/mL)在24孔或96孔板中培养,其中Iscove 's Modified Dulbecco 's Medium (IMDM)(在一些实验中取代了Roswell Park Memorial Medium (RPMI)),补充5%的自体血清(在一些实验中10%胎牛血清(FCS)或不替换血清),1%的青霉素和链霉素,1%的l -谷氨酰胺单独或存在AT7519 (Astex Pharmaceuticals, Cambridge, UK),粒细胞-巨噬细胞集落刺激因子(GM-CSF;R&D Systems, Abingdon, UK)或大肠杆菌LPS (Sigma, Dorset, UK),在37°C, 5% CO下保存24小时gydF4y2Ba2gydF4y2Ba.在规定的时间点,中性粒细胞在DPBS中重悬(1:5)gydF4y2Ba-/-gydF4y2Ba添加25 mM氯化钙,用Annexin V-FLUOS (Sigma)以1:500和1µg/mL碘化丙酯(PI)标记,然后按所述在BD FACS Scan, FACS Calibur或BD Accuri细胞分析仪上分析。gydF4y2Ba16gydF4y2Ba用Diff-Quick (Gamidor, Didcot, UK)对细胞离心制剂进行染色,以评估细胞凋亡的形态学变化。gydF4y2Ba
西方墨点法gydF4y2Ba
如前所述,进行了Western blotting,gydF4y2Ba17gydF4y2BaMcl-1 (1:1000;圣克鲁兹,达拉斯,德克萨斯州,美国),BAX (1:1000;β-肌动蛋白(1:50 000;Sigma)和辣根过氧化物酶偶联二抗(1:2500;Dako,剑桥郡,英国)。gydF4y2Ba
net的显微检测gydF4y2Ba
中性粒细胞被播种(5×10gydF4y2Ba4gydF4y2Ba/well)在含5% FCS的RPMI中放入24孔板,让其粘附30分钟,然后用10 nM PMA刺激,在37°C, 5% CO下孵育4小时gydF4y2Ba2gydF4y2Ba.在一些实验中,在刺激前,允许它们在单独的培养基或含有1 μ M AT7519和/或2.5 ng/mL GM-CSF的培养基中粘附6小时,并进一步孵育4小时。培养时间结束后,加入0.15µM SYTOX绿色(Invitrogen, Thermo Scientific, UK),然后在EVOS FL细胞成像系统上捕获亮场和荧光(470/22 nm发光二极管激发)图像。NET形成被量化为SYTOX阳性NET每10×场的百分比(NET占亮场总细胞计数的百分比),如所述。gydF4y2Ba15日18gydF4y2Ba所有样品都被重复电镀,每孔计数多个区域。gydF4y2Ba
NET DNA释放动力学试验gydF4y2Ba
这是基于一项已发表的分析。gydF4y2Ba15日19gydF4y2Ba分离的中性粒细胞被接种(5×10gydF4y2Ba4gydF4y2Ba在RPMI 1640介质中添加5% FCS,放入平底96孔板中,在37°C, 5% CO下粘附30分钟gydF4y2Ba2gydF4y2Ba.通过添加10 nM PMA诱导NETs,并通过添加0.15µM SYTOX green(一种细胞渗透性核酸染料,激发/发射光谱为504/523 nM)在Synergy HT BioTek平板阅读器中每隔30分钟检测NETs。在一些实验中,在PMA刺激前将1µM AT7519和/或25 ng/mL GM-CSF添加到培养基中6小时。gydF4y2Ba
人巨噬细胞培养gydF4y2Ba
分离的pmcs在IMDM中接种到6孔板中60分钟(Nunc Upcell, ThermoFisher, Waltham, MA)。然后去除培养基,细胞在IMDM中洗涤两次,剩余的粘附单核细胞在添加10%自体血清、1%青霉素和链霉素、1% l -谷氨酰胺的IMDM中培养,在37°C、5% CO下培养5天gydF4y2Ba2gydF4y2Ba.第5天,细胞在DPBS中洗涤两次gydF4y2Ba-/-gydF4y2Ba在分离之前。然后将单核细胞来源的巨噬细胞(MDM)种在2.5×10的48孔组织培养板中gydF4y2Ba5gydF4y2Ba在37°C, 5% CO条件下,注入补充10%自体血清,1%青霉素和链霉素,1% l -谷氨酰胺的IMDM,持续1-2天gydF4y2Ba2gydF4y2Ba.gydF4y2Ba
人net与巨噬细胞共培养gydF4y2Ba
在汉克斯平衡盐溶液(HBSS)中,用100 nM PMA或二甲基亚砜对照刺激分离的中性粒细胞gydF4y2Ba-/-gydF4y2Ba)在室温下滚动悬浮15分钟(以防止结块)。中性粒细胞在HBSS中洗涤三次gydF4y2Ba-/-gydF4y2Ba在播种5×10之前去除任何残留的PMAgydF4y2Ba5gydF4y2Ba在500 μ L IMDM中添加10%巨噬细胞供体血清,1%青霉素和链霉素,1% L-谷氨酰胺,在37°C, 5% CO下24小时(中性粒细胞与MDM比例为2:1)gydF4y2Ba2gydF4y2Ba.pma处理的中性粒细胞被认为是NETing中性粒细胞(或NETs),非pma处理的中性粒细胞作为对照中性粒细胞。24小时收集上清液,300倍离心gydF4y2BaggydF4y2Ba5分钟以去除细胞碎片,并在−80°C冷冻。根据制造商的说明,使用Direct-Zol RNA提取试剂盒(Zymo Research, Irvine, CA)从剩余的粘附MDMs中提取RNA。gydF4y2Ba
共培养上清ELISAgydF4y2Ba
按照制造商的说明,使用市售的elisa(研发系统)在培养上清中测量IL-8和肿瘤坏死因子(TNF)。gydF4y2Ba
巨噬细胞基因qRT-PCR使用市售的TaqMan基因表达测定法(ThermoFisher),根据制造商说明对IL8、TNF、CXCL9、CCL17和18s RNA进行qRT-PCR。数据以相对于对照的折数变化表示。gydF4y2Ba
支气管组织扫描电子显微镜gydF4y2Ba
支气管组织取自移植的CF肺(经机构委员会批准,爱荷华大学),用2.5%戊二醛固定在0.1 M碳酸钙缓冲液中,并进行电子显微镜(EM)处理。要了解完整的方法,请参见在线gydF4y2Ba补充数据gydF4y2Ba.gydF4y2Ba
补充材料gydF4y2Ba
统计分析gydF4y2Ba
所有数据均以均数±标准差表示。流式细胞术数据使用FlowJo软件(TreeStar, Ashland, Oregon)或BD Accuri平台专用软件进行分析。使用GraphPad Prism (La Jolla, CA)对数据进行t检验、Mann-Whitey U检验或方差分析,并进行适当的后测,如图所示。gydF4y2Ba
结果gydF4y2Ba
由于细胞凋亡减少,CF中性粒细胞存活率增加gydF4y2Ba
通过细胞离心制剂的形态学评估,CF中性粒细胞显示生存率增加(gydF4y2Ba图1 a - cgydF4y2Ba, p < 0.001)。我们用流式细胞仪Annexin V和PI染色证实了这一点。对照组中性粒细胞在24小时内比CF中性粒细胞显示出更多的完全凋亡,(gydF4y2Ba图1 d, EgydF4y2Ba, p<0.01),在24小时时,CF样本中有更多的中性粒细胞存活,这表明细胞凋亡减少后出现了促生存表型(gydF4y2Ba图1 fgydF4y2Ba, p = 0.0004)。Pan-caspase抑制可有效抑制CF中性粒细胞的所有细胞死亡(gydF4y2Ba图1 ggydF4y2Ba, p<0.001),证实细胞凋亡是CF中性粒细胞死亡的主要机制,但与健康对照组相比,细胞凋亡延迟。gydF4y2Ba

由于细胞凋亡减少,CF中性粒细胞存活率增加。(A)通过形态学评估,培养的CF中性粒细胞在24小时时比健康对照更有活力(n=6 CF, 5对照)。(B)体外培养中新鲜分离的活中性粒细胞显示多裂核(×100目标)。(C)长时间体外培养后凋亡中性粒细胞显示特征性核凝结(×100目的)。(D)对照中性粒细胞体外培养24小时的流动图,显示少量活细胞(V=膜联蛋白V−ve/PI−ve细胞),大多数细胞已进入凋亡(A=膜联蛋白V+ve/PI−ve)或已完成凋亡(CA=膜联蛋白V+ve/PI+ve)。(E)对照组中性粒细胞完全凋亡率高于CF (n=19 CF和20对照组,患者详细信息见在线)gydF4y2Ba补充表E1gydF4y2Ba).(F) 24小时后CF培养中存在更多存活的中性粒细胞(n=19 CF和20对照组)。(G)将泛caspase抑制剂Q-VD-OPh水合物添加到CF中性粒细胞培养物中,通过抑制构成性凋亡来增加存活率。有关患者详情,请参阅网上gydF4y2Ba补充表E1gydF4y2Ba.数据以平均值±SEM表示。采用双向方差分析和Bonferroni (A, E, G)分析,未配对t检验(F). **p<0.01;* * * p < 0.001。CF,囊性纤维化;PI,碘化丙啶;QVD, Q-VD-OPh水合物。gydF4y2Ba
CF中延迟的中性粒细胞凋亡与囊性纤维化跨膜电导调节功能的丧失有关gydF4y2Ba
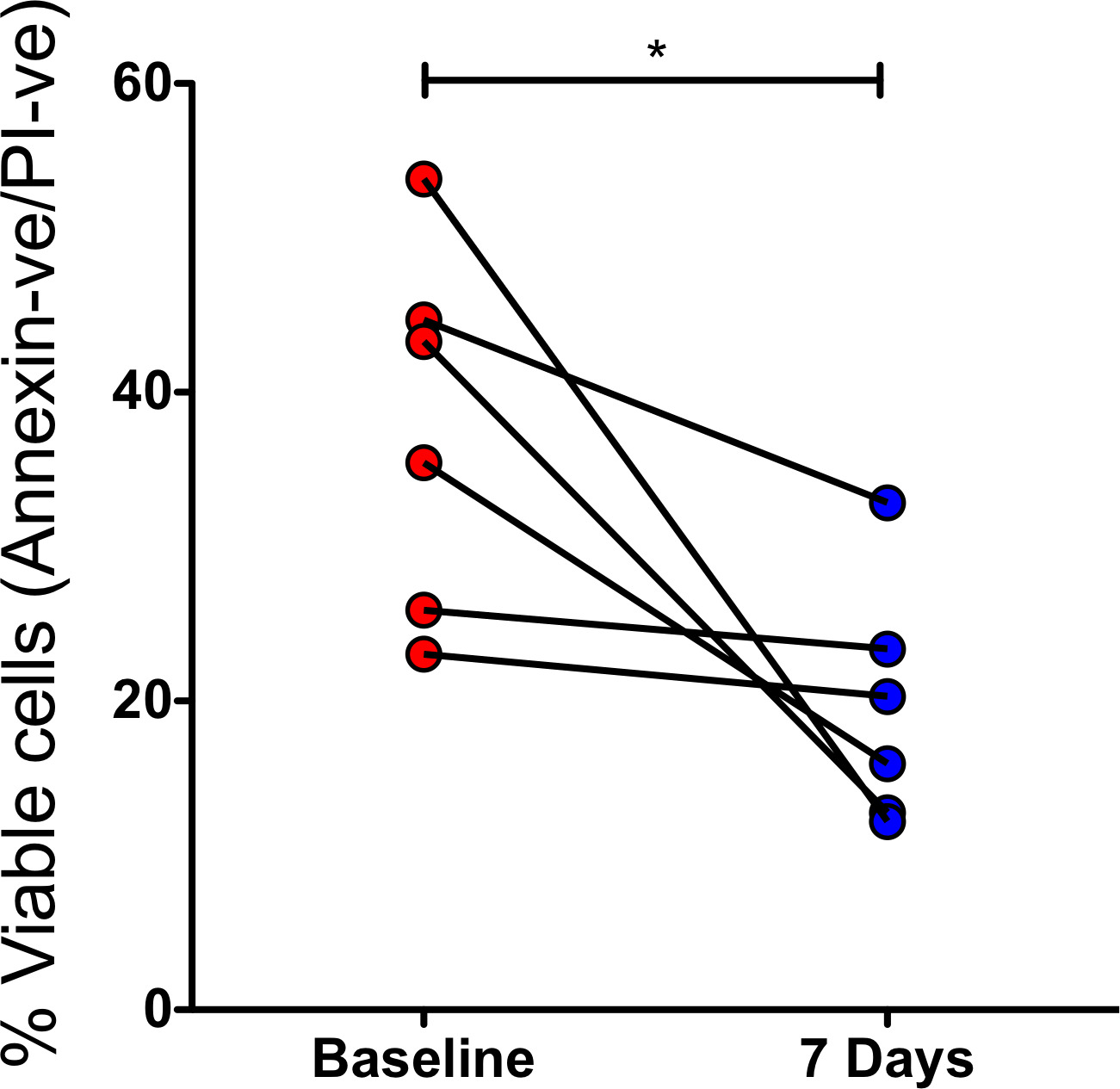
接下来,我们评估了CF中延迟的凋亡是否与中性粒细胞中囊性纤维化跨膜电导调节(CFTR)功能下降有关。我们从12例至少有一种G551D突变的患者中获得中性粒细胞(使用Ficoll-Paque方法和红细胞溶解法),这些患者在开始ivacaftor治疗(150mg,每天两次)前和2天内发生突变,ivacaftor是一种CFTR增强剂药物,可增加异常CFTR的电导,并具有已证实的临床效果。gydF4y2Ba20.gydF4y2Ba这些样本是之前发表的一项观察研究的一部分,gydF4y2Ba21日22gydF4y2Ba但其他工作是分开准备和分析的。ivacaftor治疗后,中性粒细胞24小时存活率显著降低,这表明CFTR增强逆转了CF中性粒细胞的原存活表型(gydF4y2Ba图2 a - cgydF4y2Ba, p = 0.014)。当患者接受了7天的治疗时,也可以看到这种效果(可在线获得6名患者的数据)gydF4y2Ba补充图S1gydF4y2Ba, p = 0.0313)。为了确保ivacaftor的凋亡诱导作用不是药物的非特异性作用,我们在培养的健康对照中性粒细胞中测试了ivacaftor诱导中性粒细胞凋亡的能力。在一定剂量的体外培养20小时后,没有显示出任何影响gydF4y2Ba补充图S2gydF4y2Ba).gydF4y2Ba
补充图s1gydF4y2Ba
补充图s2gydF4y2Ba

CF中延迟的中性粒细胞凋亡与囊性纤维化跨膜电导调节(CFTR)功能的丧失有关。在启动CFTR增强剂ivacaftor之前和之后,从至少有一个G551D突变的12名患者中收集中性粒细胞(患者详细信息,请参阅在线gydF4y2Ba补充表E2gydF4y2Ba),体外培养24小时。(A)和(B)展示了单个患者在(A)开始ivacaftor前和(B)开始ivacaftor后2天的示例流图(24小时体外培养后),显示中性粒细胞存活率随着治疗而下降。(C) 12例接受ivacaftor治疗的患者的联合数据显示,中性粒细胞活力随治疗显著下降。(D)中性粒细胞取自2周龄的CF仔猪和野生型(WT)对照组。体外培养24小时后的流式细胞仪显示CF存活率增加(5头CF猪和5头WT对照组)。(E)具有代表性的CF和WT中性粒细胞在0和24小时的细胞离心制剂(×100目标)显示,在24小时,WT中凋亡中性粒细胞的数量增加。数据以平均值±SEM表示。采用配对t检验(C)、Mann-Whitney U检验(D)进行分析,*p<0.05。CF,囊性纤维化;PI,碘化丙啶。gydF4y2Ba
接下来,我们利用CFTR中CFTR缺失的中性粒细胞来研究中性粒细胞存活表型是否与中性粒细胞中CFTR缺失有关gydF4y2Ba-/-gydF4y2Ba仔猪(CF仔猪)。从2周龄CF仔猪和野生型对照组中收获中性粒细胞(采用Ficoll-Paque法和红细胞裂解法),并在10% FCS存在下体外培养。CF仔猪中性粒细胞表现出较长的生存时间,这表明增加的中性粒细胞生存(由于凋亡减少)是与中性粒细胞缺乏CFTR相关的主要缺陷(gydF4y2Ba图2 d, EgydF4y2Ba, p = 0.027)。gydF4y2Ba
CF中性粒细胞寿命延长不是由炎症引起的gydF4y2Ba
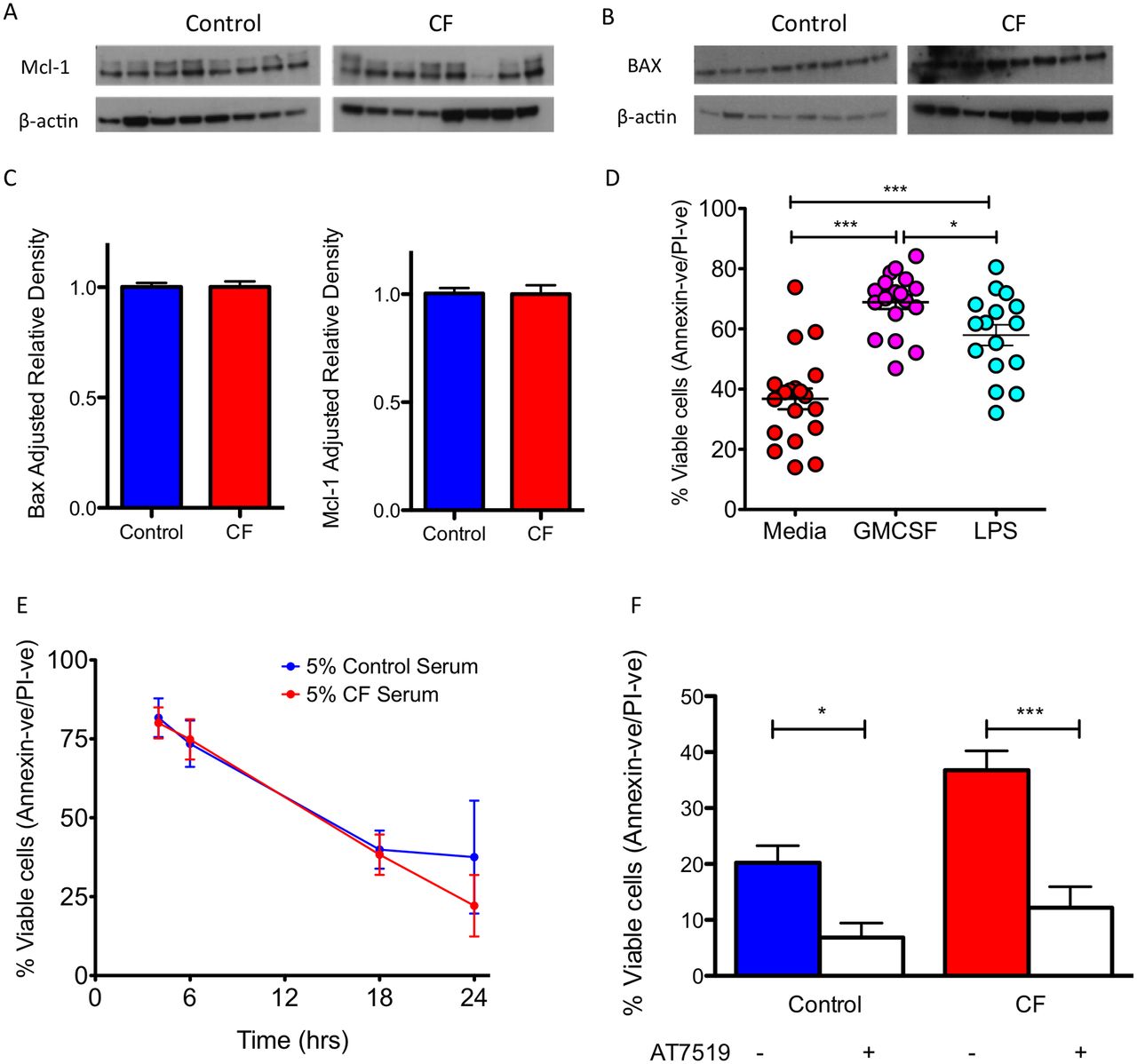
炎症被认为可以通过诱导Bcl-2家族成员,特别是Mcl-1的表达变化来延长中性粒细胞的寿命gydF4y2Ba23gydF4y2Ba和伯灵顿。gydF4y2Ba5gydF4y2Ba使用CF患者的样本,我们发现CF中性粒细胞和健康对照之间Mcl-1和BAX的表达没有差异(gydF4y2Ba图3 a - cgydF4y2Ba),提示炎症诱导的内在凋亡调节蛋白的失调并不是CF中性粒细胞存活时间延长的原因。此外,向CF中性粒细胞中添加促生存刺激GM-CSF或LPS可增强促生存表型,表明激活因子可进一步增强CF中性粒细胞存活的增加(gydF4y2Ba图3 dgydF4y2Ba, p<0.001),而不是潜在的机制。为了评估与炎症环境相关的血清因子是否不会导致细胞凋亡延迟,我们在CF血清存在的情况下培养健康对照中性粒细胞。在健康的中性粒细胞培养物中加入5%的CF血清并不能增加中性粒细胞的存活率(gydF4y2Ba图3 egydF4y2Ba).接下来,我们询问是否可以从治疗上逆转CF中性粒细胞的凋亡延迟。在AT5719的存在下培养CF中性粒细胞,AT5719是一种周期蛋白依赖性激酶(CDK)抑制剂(CDKi),已知可诱导中性粒细胞凋亡,gydF4y2Ba17gydF4y2Ba显著诱导对照组细胞凋亡(p<0.01)和CF中性粒细胞凋亡(p<0.001),有效纠正CF细胞凋亡延迟至对照组水平(gydF4y2Ba图3 fgydF4y2Ba).gydF4y2Ba

CF中性粒细胞寿命延长不是由炎症引起的。新鲜分离的CF中性粒细胞具有正常的基线凋亡信号(A) Mcl-1 (37 kDa抗凋亡蛋白)和(B) BAX (21 kDa促凋亡蛋白)(n=8 CF和8个健康对照,患者详情见在线gydF4y2Ba补充表E3gydF4y2Ba).(C) Mcl-1和BAX密度测定。(D) CF中性粒细胞在促生存刺激GM-CSF (20 ng/mL)和LPS (10 ng/mL)下对延迟凋亡保持敏感性24小时(n=19 CF和20对照组,患者详细信息见在线gydF4y2Ba补充表E1gydF4y2Ba).(E) CF的主要凋亡缺陷不是由于循环的CF血清因子。健康对照中性粒细胞在含5% CF血清的培养基中培养,这并未导致中性粒细胞存活率增加(n=3个独立的健康供体)。(F) CF中的生存缺陷可以通过AT7519(1µM)培养24小时来纠正,以增加中性粒细胞凋亡,并有效地将生存降低到健康对照组水平(n=19 CF和20对照组,患者详细信息见在线gydF4y2Ba补充表E1gydF4y2Ba).数据以平均值±SEM表示。采用未配对t检验(C)分析,采用Newman-Keuls后验(D, F)进行单向方差分析。*p<0.05;* * p < 0.01;* * * p < 0.001。CF,囊性纤维化;GMCSF,粒细胞-巨噬细胞集落刺激因子;有限合伙人,脂多糖;PI,碘化丙啶。gydF4y2Ba
CF中性粒细胞由于其促生存表型,比健康对照组形成更多的netgydF4y2Ba
由于CF中性粒细胞的特征是细胞凋亡减少,我们随后询问它们是否因此更容易发生NETosis。新鲜分离的人CF中性粒细胞对10 nm PMA的反应与健康对照中性粒细胞一样有效地形成NETs (gydF4y2Ba图4一gydF4y2Ba)培养4小时后。通过DNA释放试验评估NET的形成证实了这一点(gydF4y2Ba图4 b, EgydF4y2Ba).然而,在6小时时,CF中性粒细胞的DNA释放与对照中性粒细胞的DNA释放不同,这表明CF中性粒细胞在培养中老化时比非CF对照组形成了更多的net,并且未能参与细胞凋亡(gydF4y2Ba图4 bgydF4y2Ba).因此,我们评估了老年CF中性粒细胞是否比老年非CF对照中性粒细胞形成更多的net。中性粒细胞老化6小时,然后用10 nM PMA刺激诱导NETs。CF中性粒细胞形成的net明显多于相同年龄的非CF对照中性粒细胞(gydF4y2Ba图4 cgydF4y2Ba, p=0.0087,且gydF4y2Ba图4 dgydF4y2Ba, p < 0.001)。然后,我们评估了添加促生存因子来控制中性粒细胞是否会增强NET的形成,或诱导CF中性粒细胞早期凋亡是否会减少NET的形成。在PMA刺激前6小时向非cf对照中性粒细胞中加入GM-CSF可增加NET的形成(gydF4y2Ba图5一个gydF4y2Ba, p < 0.01)。相反,CDKi抑制CF中性粒细胞中NET的形成(gydF4y2Ba图5 a, BgydF4y2Ba, p < 0.001)。gydF4y2Ba

在老化培养条件下,CF中性粒细胞比健康对照组形成更多的net。(A)新分离的CF中性粒细胞与健康对照组形成类似数量的net(11个CF vs 6个对照组,患者详细信息见在线gydF4y2Ba补充表E3gydF4y2Ba),然后用10 nM的豆蔻酸酯(PMA)刺激。(B) DNA释放作为NET产生的替代标记,在pma处理的新鲜分离的中性粒细胞中产生了类似的结果(7个CF vs 6个对照组),尽管我们观察到CF中性粒细胞在稍后时间点的DNA释放无统计学意义上的显著增加。(C)在PMA刺激前培养6小时(10 nM)的中性粒细胞显示CF中性粒细胞与对照组相比增加了NET的产生(n=6 CF vs 6对照组)。(D)通过DNA释放试验证实老年中性粒细胞中NET产量增加(n=6 CF vs 6对照)。(E)在非固定细胞中加入SYTOX绿色后,具有代表性的荧光显微镜显示,与对照组(以及未衰老的CF)相比,中性粒细胞老化后CF中pma诱导的NETs增加。可见弥散形态和扩散形态NETs(比例尺=10µm)。数据以平均值±SEM表示。采用未配对t检验(A, C)分析,采用Bonferroni (D)进行双向方差分析。**p<0.01;* * * p < 0.001。CF,囊性纤维化; NET, neutrophil extracellular trap; OD, optical density.

CF中性粒细胞由于其促生存表型而形成更多的net。(A)健康对照组和CF中性粒细胞在GM-CSF (2.5 ng/mL)±AT7519(1µM)存在下培养6小时,然后用PMA (10 nM)刺激。加入GM-CSF来控制中性粒细胞,使NET的形成增加到CF水平。AT7519逆转了这种作用,提示诱导细胞凋亡阻止了NET的形成(n=6 CF和6健康对照组)。(B)具有代表性的荧光显微图像显示,CF患者培养中SYTOX呈扩散NETs阳性,GM-CSF增强,AT7519抑制(比例线=10µm)。数据以平均值±SEM表示。采用Newman-Keuls后验进行单因素方差分析(A). **p<0.01;* * * p < 0.001。CF,囊性纤维化;GMCSF,粒细胞-巨噬细胞集落刺激因子; NET, neutrophil extracellular trap; PMA, phorbol myristate acetate; AT, AT7519.
NETs刺激巨噬细胞的炎症反应,这在CF中被夸大gydF4y2Ba
我们使用扫描EM证明了一名接受肺移植的CF患者肺中存在NETs (gydF4y2Ba图6gydF4y2Ba)基于形态和特征,类似于以前在EM上对net的描述。gydF4y2Ba9gydF4y2Ba接下来,我们开发了一种技术,使NETs (NETing中性粒细胞)和MDMs以一致的比例共培养,方法是在滚动条件下用100 nM PMA刺激中性粒细胞,清洗,然后将它们添加到MDMs中,在4-6小时后形成具有经典形态的NETs(在线)gydF4y2Ba补充图S3gydF4y2Ba).共培养24小时后收集上清液和RNA (gydF4y2Ba图6 bgydF4y2Ba).在健康志愿者MDMs中加入NETs可诱导产生IL-8和TNF (gydF4y2Ba图6 c, DgydF4y2Ba, p<0.01, p<0.05)。当将net添加到CF MDMs中时,也观察到这种效果(gydF4y2Ba图6 c, DgydF4y2Ba, p<0.001 both)。与健康志愿者相比,CF MDMs中IL-8和TNF对NETs的反应被夸大(gydF4y2Ba图6 c, DgydF4y2Ba, p<0.05, p<0.001)。qRT-PCR证实IL-8和TNF表达增加(未显示),并证明NETs诱导了CCL17的表达,CCL17是巨噬细胞选择性激活的标志,gydF4y2Ba24gydF4y2Ba在CF和控制mdm (gydF4y2Ba图6 egydF4y2Ba, p < 0.001)。值得注意的是,CF MDMs增加了经典巨噬细胞激活标志物CXCL9的基础表达(gydF4y2Ba图6 fgydF4y2Ba, p < 0.001),gydF4y2Ba25gydF4y2Banet进一步增强了这一结果(尽管没有统计学意义)。总的来说,这些数据表明CF MDMs与NETs相互作用具有经典激活和替代激活的表型标记。gydF4y2Ba
补充图s3gydF4y2Ba

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
net刺激巨噬细胞的炎症反应,这在CF中被夸大。(A)移植CF患者的支气管扫描电子显微镜显示与中性粒细胞和细菌相关的特征性net样物质链。(B)新型NET/MDM共培养系统概述。(C) NETs对MDMs具有促炎作用,在共培养24小时后导致IL-8的产生增加,并且这种作用在CF MDMs中更为明显(n=8 CF和7健康MDM供体,患者详细信息见在线gydF4y2Ba补充表E3gydF4y2Ba).(D) NETs诱导MDMs产生TNF。(E) NETs在健康和CF MDMs中诱导CCL17表达。(F) net在健康对照组中诱导CXCL-9表达无显著增加,而CF中CXCL-9在基线时过表达。数据以平均值±SEM表示。采用Newman-Keuls后验(C-F)进行单向方差分析。* * p < 0.01;* * * p < 0.001。趋化因子(C-C motif)配体;CF,囊性纤维化;CXCL,趋化因子(C-X-C motif)配体; IL, interleukin; MDM, monocyte-derived macrophage; NET, neutrophil extracellular trap; PMA, phorbol myristate acetate; TNF, tumour necrosis factor; WT, wild type.
讨论gydF4y2Ba
我们的数据表明,由于延迟凋亡,CF中性粒细胞存活时间更长,这与CFTR功能的缺失有关。此外,细胞凋亡的延迟使得CF中性粒细胞比健康对照中性粒细胞更有效地形成NETs,这可能是一个重要的促炎机制。有报道称CF中中性粒细胞凋亡受损,gydF4y2Ba4 5 26gydF4y2Ba之前的研究表明,这要么是对全身炎症的反应gydF4y2Ba5gydF4y2Ba或者,CF中性粒细胞的特定缺陷。gydF4y2Ba4 26gydF4y2Ba用(CFTR增强剂)ivacaftor治疗的CF患者细胞凋亡延迟的逆转表明中性粒细胞中的CFTR功能与细胞凋亡途径有关。事实上,之前的研究已经证明了用ivacaftor增强CFTR对中性粒细胞的直接影响,gydF4y2Ba27 28gydF4y2Ba这表明CFTR在一些中性粒细胞功能中起着重要作用,与CFTR在生物学上显著表达一致。gydF4y2Ba29gydF4y2Ba我们观察到CFTR无效仔猪的中性粒细胞凋亡受损,这强调了CF中延迟中性粒细胞凋亡的潜在主要性质。CFTR已被证明在两种巨噬细胞的吞噬溶酶体活性中具有功能性作用gydF4y2Ba30.gydF4y2Ba和中性粒细胞,gydF4y2Ba31日32gydF4y2BaCFTR缺失促进巨噬细胞释放炎性细胞因子。gydF4y2Ba14 33gydF4y2BaCFTR还涉及骨髓抑制细胞对T细胞抑制的调节gydF4y2Ba34gydF4y2Ba因此,CFTR在髓系细胞中多样但功能性的作用现在已经确定。gydF4y2Ba
我们的数据表明,CF中性粒细胞的延长生存不能完全用CF的炎症来解释。Bcl-2家族的促凋亡和抗凋亡蛋白BAX和McL-1与炎症诱导的中性粒细胞生存有关。在脓毒症患者的中性粒细胞中观察到Mcl-1过量,导致中性粒细胞存活时间延长,gydF4y2Ba23gydF4y2Ba但我们发现在CF和健康对照中性粒细胞之间没有表达差异。炎症诱导的BAX缺乏被认为是CF和肺炎中延迟中性粒细胞凋亡的原因,gydF4y2Ba5gydF4y2Ba但同样,在CF和健康对照中性粒细胞之间没有表达差异。我们的数据推断,CF中的凋亡延迟是CF中性粒细胞的主要特征,而不仅仅与炎症有关,进一步强调CF血清不能延长健康对照中性粒细胞的生存。将CF中性粒细胞暴露于促生存刺激(如GM-CSF或LPS)可进一步提高生存率,一旦中性粒细胞到达CF肺的炎症环境,这可能尤其相关。CF肺环境已被证明可诱导中性粒细胞行为和表型的变化,gydF4y2Ba35 36gydF4y2Ba因此很可能会强调我们在CF中展示的促生存表型。gydF4y2Ba
我们使用CDKi AT7519将CF中性粒细胞凋亡恢复到健康对照水平,这一策略已被证明可以减少无菌和感染炎症模型中的炎症。gydF4y2Ba17 37gydF4y2Ba细胞凋亡是一种被广泛描述的处理潜在毒性和破坏性中性粒细胞的途径,这些凋亡细胞的吞噬作用驱动炎症的解决。gydF4y2Ba3.gydF4y2Ba因此,针对中性粒细胞凋亡作为CF的抗炎策略是有吸引力的,因为它将提供一种不依赖于患者个体CF基因型的通用治疗。此外,更有效地使CF中性粒细胞参与细胞凋亡可以避免更有毒的细胞处理形式,如NETosis。gydF4y2Ba
我们首次证明CF中性粒细胞比健康对照组形成更多的net。一些证据强调了NETosis在CF中的重要性。自20世纪60年代以来,CF气道中过多的DNA被认为是肺部疾病的潜在因素,尽管DNA以NETs形式的存在是最近的发现。gydF4y2Ba8 9 38 39gydF4y2Ba然而,这些研究未能确定CF中性粒细胞是否比健康对照细胞形成更多的net。我们的数据表明,CF中性粒细胞形成的net比对照组更多,这与CF中性粒细胞凋亡的减少直接相关。这方面的证据是多方面的。首先,当中性粒细胞在PMA刺激前老化时,NET形成的主要差异被观察到,此时控制中性粒细胞虽然存活,但将参与早期凋亡机制。其次,如果非CF中性粒细胞在PMA刺激前在GM-CSF的存在下培养(以延迟凋亡),它们形成与CF中性粒细胞相同数量的net。最后,在PMA刺激前,在CDKi存在下培养CF中性粒细胞6小时,可以将CF中NET的形成水平降低到健康对照组的水平。综上所述,这些数据表明CF中性粒细胞形成了更多的NET,因为它们不太能够参与正常的凋亡过程,但这可以通过CDKi增强凋亡来逆转,这与其他研究表明NET的形成需要几个非凋亡途径的参与一致。gydF4y2Ba40-42gydF4y2Ba这些数据也表明CDKi可能提供了一种可行的治疗策略,可以颠覆CF中的NETosis,并促使细胞凋亡,从而促进炎症的解决。CDKs以多种亚型的形式存在,并在细胞行为中发挥不同的作用,尽管参与调节中性粒细胞凋亡的主要亚型是CDKs 7和CDKs 9。gydF4y2Ba43 44gydF4y2BaCDKi也可能影响其他激酶,例如糖原合成酶激酶3 (GSK-3),特别是GSK-3β亚型。gydF4y2Ba45gydF4y2Ba虽然值得注意的是,GSK-3α是中性粒细胞中发现的主要亚型,gydF4y2Ba46gydF4y2Ba在目前的工作中,这种脱靶效应可能不那么重要。因此,进一步研究CDKs 7和9在CF中性粒细胞凋亡和NETosis中的作用对于我们理解这些过程至关重要。gydF4y2Ba
最近有认识到,粒细胞种群可能显示一定程度的表型异质性,导致低密度中性粒细胞(ldn)的描述。gydF4y2Ba47gydF4y2Ba有趣的是,ldn的一个子类,低密度粒细胞(LDGs)在SLE中可能比“正常”粒细胞形成更多数量的net,gydF4y2Ba48gydF4y2Ba尽管在类风湿关节炎的NETosis研究中有相反的发现。gydF4y2Ba49gydF4y2Ba我们在本研究中没有研究ldn的作用,但是ldn /LDGs对CF中NET形成的贡献值得进一步研究。gydF4y2Ba
我们证明NETs对巨噬细胞是一种强有力的促炎信号。自2004年首次描述NETs杀死细菌的能力以来,gydF4y2Ba6gydF4y2Ba但CF中net存在的潜在炎症后果却没有。我们开发了一种检测方法,以评估NETing中性粒细胞与巨噬细胞以一致的比例培养是否能促进促炎细胞因子的释放(来自巨噬细胞),以及CF巨噬细胞是否更容易受到这种刺激。与之前的数据相反,NETs诱导MDMs释放IL-8和TNF。gydF4y2Ba50gydF4y2Ba然而,我们的数据与最近的发现是一致的,NETs可以启动巨噬细胞进一步释放细胞因子。gydF4y2Ba13gydF4y2Ba我们证明了CF MDMs对NETs的反应增强了细胞因子的释放,与CF巨噬细胞对促炎刺激的高度反应一致。gydF4y2Ba14gydF4y2Ba我们还证明NETs与CF MDMs的相互作用与经典和替代巨噬细胞激活标记物的表达相关,并推测这可能是CF中无法解决(挫败性)炎症的一个促成因素。CF上皮细胞是CF先天免疫系统的核心部分,因此NETs与上皮细胞的相互作用具有重要意义,将有待于未来的研究。gydF4y2Ba
我们承认本研究存在一些局限性。首先,我们使用外周血中性粒细胞研究CF背景下的细胞凋亡和NETosis。气道中性粒细胞在CF中可能具有独特的表型,gydF4y2Ba35 36gydF4y2Ba因此,本文所述过程的进一步研究(在气道中性粒细胞)可能是指示。尽管如此,外周中性粒细胞以前曾被用来评估中性粒细胞的功能,gydF4y2Ba4 27 29 51gydF4y2Ba强调这种方法的有效性。其次,我们从已确诊肺部疾病并因此确诊肺部炎症的CF成人中获得中性粒细胞。因此,我们的数据不能完全排除CF中炎症对中性粒细胞死亡途径的影响,尽管我们的数据表明机制与炎症无关(特别是当我们考虑到来自gydF4y2Ba雌性生殖道gydF4y2Ba-/-gydF4y2Ba小猪)。进一步研究CF患儿的中性粒细胞在重大炎症性肺病发病前后的这些过程将有助于在未来解决这一问题。第三,尽管我们认为CFTR缺乏与延迟凋亡有关,但在G551D突变患者中,可以通过ivacaftor治疗来纠正,CFTR增强和纠正及其对凋亡和NETosis等中性粒细胞过程的影响还需要进一步的研究。新的联合疗法的出现将纠正和增强更常见的CF突变,如F508del,这将在未来成为可能。gydF4y2Ba52gydF4y2Ba允许研究CFTR对中性粒细胞功能的体内和体外校正。此外,我们建议CFTR缺失猪也可以用于调查与CFTR缺乏相关的中性粒细胞功能和死亡,作为我们在本文中的数据和最近在CF猪巨噬细胞中的研究的扩展。gydF4y2Ba33gydF4y2Ba
总之,我们证明CF中性粒细胞具有继发于凋亡减少的延长生存期,这可能是CF中性粒细胞的主要缺陷。CF中性粒细胞增加NET形成是凋亡失败的结果,可以被CDKi药物靶向。NET对巨噬细胞是一种强大的促炎刺激,cf来源的巨噬细胞对NET刺激高度敏感。总之,中性粒细胞凋亡的内在延迟增强了CF中NET的形成,从而导致炎症,这为未来抗炎治疗的发展提供了一个新的靶点。gydF4y2Ba
致谢gydF4y2Ba
我们要感谢苏格兰国家CF服务的护理人员对患者样本收集的帮助。我们也要感谢爱荷华大学中央显微镜研究设施的使用,以及由囊性纤维化基因治疗中心(NIH P30 DK-54759)和囊性纤维化基金会部分支持的电子显微镜的帮助。AT7519是来自Astex制药公司(剑桥,英国)的一份友好的礼物。gydF4y2Ba
参考文献gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
脚注gydF4y2Ba
GH和KHR的贡献相同。gydF4y2Ba
贡献者gydF4y2BaRDG、GH、KHR、MS、CTR、RD、AM、JMF、LP、BNM、GC、CDL和DAD进行了研究。RDG, AGR, DJD, CH, MKBW, PKS, SCD, PBM, DAS和EFM设计了研究。RDG, MKBW, AGR和DJD撰写了手稿。gydF4y2Ba
资金gydF4y2Ba这项工作得到了威康基金会WT093767 (RDG)、WT094415 (CDL)和WT096497 (DAD)的支持;英国医学研究委员会G1002046 (DJD)和MR/K013386/1 (AGR, RD,和CH)。对CF猪的研究得到了NIH的资助:P01 HL-51670, P01 HL-091842和囊性纤维化基金会RDP。接受Ivacaftor治疗的患者的标本采集由Vertex制药公司(Boston, MA, USA)的研究者发起的无限制赠款支持。gydF4y2Ba
相互竞争的利益gydF4y2Ba没有宣布。gydF4y2Ba
伦理批准gydF4y2Ba东苏格兰研究伦理委员会,15/ES/0094;苏格兰西部研究伦理委员会,11/WS/0074;洛锡安研究伦理委员会,08/S1103/38。gydF4y2Ba
出处和同行评审gydF4y2Ba不是委托;外部同行评审。gydF4y2Ba