条文本gydF4y2Ba

摘要gydF4y2Ba
背景gydF4y2Ba暴露于猪尘会引起严重的气道炎症,导致健康人类受试者鼻腔和支气管肺泡灌洗液中白细胞介素8 (IL-8)水平升高,主要是中性粒细胞水平升高。目前还不清楚猪舍环境中的哪些成分是导致呼吸道反应的原因。本研究的目的是评价和比较猪粉尘成分对人肺泡巨噬细胞和支气管上皮细胞的体外影响。gydF4y2Ba
方法gydF4y2Ba用猪灰、脂多糖(LPS;革兰氏阴性菌中存在)、谷物粉尘(猪饲料成分)和葡聚糖(真菌的结构成分)呈剂量反应(1 ~ 100 μg/ml)。gydF4y2Ba
结果gydF4y2Ba浓度为100 μg/ml的猪粉尘可使NHBE细胞、A549细胞和巨噬细胞的IL-8产量分别增加20倍、28倍和15倍。LPS (100 μg/ml)对三种细胞均有显著的刺激作用,对巨噬细胞的刺激程度与猪灰相同,但对NHBE和A549细胞的刺激作用是猪灰的5-8倍。100 μg/ml颗粒粉对A549细胞无影响,但对NHBE细胞和巨噬细胞有刺激作用。葡聚糖(100 μg/ml)对A549细胞和巨噬细胞有刺激作用,对NHBE细胞无刺激作用。葡聚糖和谷粒粉尘的刺激作用均弱于猪粪和LPS。猪尘溶液中LPS含量为2.16 (0.2)ng/100 μg,谷物粉尘中LPS含量为0.53 (0.04)ng/100 μg。gydF4y2Ba
结论gydF4y2Ba猪尘是支气管上皮细胞和人肺泡巨噬细胞中产生IL-8的强烈刺激物,而LPS在这些细胞中具有不同的效力。gydF4y2Ba
- 气道上皮细胞gydF4y2Ba
- 肺泡巨噬细胞gydF4y2Ba
- 猪尘埃gydF4y2Ba
- 脂多糖gydF4y2Ba
- 引发gydF4y2Ba
数据来自Altmetric.comgydF4y2Ba
白介素8 (IL-8)主要被认为是中性粒细胞的趋化剂和激活剂,可促进与炎症性肺部疾病相关的炎症反应。这种细胞因子已被证明是由呼吸道中存在的多种细胞类型产生的,如人类的肺泡巨噬细胞,gydF4y2Ba1gydF4y2Ba支气管上皮细胞,gydF4y2Ba2gydF4y2Ba,gydF4y2Ba3.gydF4y2Ba成纤维细胞,gydF4y2Ba4gydF4y2Ba内皮细胞,gydF4y2Ba5gydF4y2Ba肥大细胞,gydF4y2Ba6gydF4y2Ba和中性粒细胞。gydF4y2Ba7gydF4y2Ba吸入有机粉尘颗粒后,肺上皮细胞构成重要的第一道防线,肺泡巨噬细胞具有很高的吞噬和杀微生物潜力。gydF4y2Ba8gydF4y2Ba肺泡巨噬细胞可单独清除少量微生物,而大量的接种量可诱导嗜中性粒细胞被巨噬细胞自身产生的趋化物质吸引流入肺泡。gydF4y2Ba
在先前未接触猪粉尘的健康受试者中,急性接触猪粉尘可诱发强烈的气道炎症,其特征是在支气管肺泡灌洗液(BAL)和鼻灌洗液中炎症细胞(IL-1、IL-6、TNF-α和IL-8)的数量增加。gydF4y2Ba9 - 11gydF4y2Ba目前还不清楚猪舍环境中的哪些成分是导致呼吸道反应的原因。猪尘由许多不同的成分组成,包括革兰氏阳性细菌和革兰氏阴性细菌,它们可能导致呼吸道的强烈炎症反应。gydF4y2Ba12gydF4y2Ba脂多糖(LPS)是内毒素的组成部分,存在于革兰氏阴性菌的细胞壁中。吸入LPS可增加健康受试者BAL液中中性粒细胞和淋巴细胞的数量。gydF4y2Ba13gydF4y2Ba体外研究表明,LPS可诱导人肺泡巨噬细胞和支气管上皮细胞分泌IL-8。gydF4y2Ba1gydF4y2Ba,gydF4y2Ba14gydF4y2Ba,gydF4y2Ba15gydF4y2Ba猪粉尘的其他成分,如谷物粉尘(猪饲料成分)和葡聚糖(真菌的主要结构成分之一,存在于大麦中)也可能是重要的。本研究的目的是评价和比较猪粉尘成分对人肺泡巨噬细胞和人支气管上皮细胞的影响。gydF4y2Ba
方法gydF4y2Ba
试剂gydF4y2Ba
脂多糖(LPS BgydF4y2Ba大肠杆菌gydF4y2Ba0111:B4, Difco,底特律,密歇根州,美国)、葡聚糖(β-1,3-葡聚糖,Wako Pure Chemical Industries,日本)和谷物粉尘(粗磨大麦,猪饲料成分,从储存容器中收集)稀释在无血清培养基中,混合并放入超声波浴中10分钟。沉降(距离地面约1.20米)的猪尘来自一个养了700-900头猪的猪圈。将粉尘溶解在培养基中,最终浓度为1mg /ml。将粉尘提取物超声处理10分钟后加入细胞培养。用显色法测定了猪灰和粮灰溶液的内毒素含量gydF4y2Ba鲎变形细胞gydF4y2Ba裂解试验(QCL-1000,内毒素,BioWhittaker,沃克斯维尔,美国,与gydF4y2Ba大肠杆菌gydF4y2Ba0111:标准为B4)。gydF4y2Ba
肺上皮细胞系(a549)gydF4y2Ba
A549人肺上皮细胞(American Type Culture Collection, Rockville, Maryland, USA),最初来源于人肺癌,gydF4y2Ba16gydF4y2Ba是有文化的。共1-2 × 10gydF4y2Ba6gydF4y2Ba将细胞培养至80厘米gydF4y2Ba2gydF4y2Ba塑料瓶(Nunc, Roskilde,丹麦),在中等哈姆氏F-12中培养,含10%胎牛血清(FCS)和100 μg/ml青霉素/链霉素(Penstrep;Biochrom KG, Seromed,柏林,德国)。培养物在37°C的5% CO的潮湿气氛中保存gydF4y2Ba2gydF4y2Ba空气和媒介每隔一天就换一次。在汇合处,将细胞暴露于胰蛋白酶/EDTA溶液(0.05/0.02%无钙和无镁磷酸盐缓冲盐水;Biochrom KG),以5 × 10的浓度补种24孔板gydF4y2Ba4gydF4y2Ba细胞/良好,生长到亚汇合。支原体污染检测(SVA,乌普萨拉,瑞典,每三个月进行一次)为阴性。gydF4y2Ba
正常人支气管上皮细胞gydF4y2Ba
原代培养(Clonetics Corporation, San Diego, California, USA)正常人支气管上皮细胞(NHBE)在支气管上皮细胞生长培养基(BEGM, Clonetics Corporation)中培养。共3.1 × 10gydF4y2Ba5gydF4y2Ba将细胞培养至80厘米gydF4y2Ba2gydF4y2Ba塑料烧瓶,在BEGM培养基中生长。培养物在37°C的5% CO的潮湿气氛中保存gydF4y2Ba2gydF4y2Ba空气和媒介每隔一天就换一次。在汇合处,将细胞暴露于胰蛋白酶/EDTA溶液(0.025/0.01%无钙和无镁磷酸盐缓冲盐水;Clonetics Corporation),以4-7 × 10的浓度在24孔板上复种gydF4y2Ba4gydF4y2Ba细胞/良好,生长到亚汇合。gydF4y2Ba
上皮细胞上清液的制备及il-8的测定gydF4y2Ba
将浓度分别为1、10和100 μg/ml的LPS、葡聚糖、谷物粉尘和猪粉尘分别加入含或不含10% FCS的Ham’s F-12培养基中,在亚汇合时分别加入1 ml的A549细胞,分别培养3个或4个细胞24小时。在BEGM培养基中分别加入25、50、100 μg/ml猪粉、50、100 μg/ml LPS、100 μg/ml谷物粉尘和葡聚糖各1 ml,每3个重复加入NHBE细胞的孔中,孵育24小时。gydF4y2Ba
所有培养上清样品在1000离心gydF4y2BaggydF4y2Ba10分钟以去除细胞碎片和颗粒物质,并储存在-70°C,直到分析。对照培养基用同样的方法从无细胞培养皿中制备。用胰蛋白酶法分离细胞,用血细胞计进行计数。细胞活力通过排除台盼蓝(生理盐水0.4%;塞拉实验室,苏塞克斯,英国)。使用商业ELISA试剂盒(Quantikine, R&D Systems, Europe, Abingdon, UK),用酶联免疫吸附法分析每百万上皮细胞中IL-8的产量。该法的下检出限为31.3 ng/l。对于重复样品,检测内变异系数(CV) <10%,检测间变异系数<20%被接受。28种细胞因子和生长因子未观察到明显的交叉反应或干扰(信息来自R&D Systems)gydF4y2Ba
肺泡巨噬细胞gydF4y2Ba
通过支气管肺泡灌洗(BAL)获得四名健康非吸烟志愿者的肺泡巨噬细胞。使用柔性纤维支气管镜(Olympus Type 4B29)在利多卡因(Xylocaine;Astra Södertälje,瑞典)在预先用药吗啡-东莨菪碱后。将支气管镜插入支气管中部叶,37°C下250 ml无菌生理盐水按5等份(每等份50 ml)灌注。每次灌注后,将液体轻轻吸入,并收集在一个硅化塑料瓶中,冷藏,回收率为65%。细胞在200度时被制成颗粒gydF4y2BaggydF4y2Ba在4°C下放置10分钟。将颗粒置于RPMI 1640培养基中,加入5%热灭活FCS、50 μg/ml庆大霉素和100 μg/ml Penstrep (Biochrom KG)。细胞总数通过血细胞计计数来确定,细胞活力通过排除台盼蓝来确定。每次灌洗得到的细胞总数为15-25 × 10gydF4y2Ba6gydF4y2Ba存活率为>85%。使用流式细胞术评估细胞分化(Epics Profile II, Coulter Electronics Inc, Hialeah, Florida, USA)。样品在Coulter Q-prep (Coulter Electronics Inc)中制备,并与CD14-CD45单克隆抗体(Mo2-RD1/Kc56-FITC, Cytostat/Coulter Clone, Coulter Corp)一起孵育10分钟。共2.5-3 × 10gydF4y2Ba5gydF4y2Ba细胞/孔接种于24孔板中,在37°C, 5% CO下孵育2小时gydF4y2Ba2gydF4y2Ba.用培养基温和清洗孔去除非粘附细胞,细胞在RPMI 1640培养基中与GE和Penstrep孵育过夜。镜检显示粘附步骤后的细胞群中含有>95%的巨噬细胞。gydF4y2Ba
巨噬细胞上清液的制备及细胞上清液中il-8的测定gydF4y2Ba
在无血清培养基中孵育约18小时后,将1 ml浓度为1、10和100 μg/ml的LPS、葡聚糖、谷物粉尘和猪粉尘分别加入含GE和Penstrep的RPMI 1640培养基中,四次重复,孵育8小时。为了研究IL-8的产量随时间的变化,在每个时间点向孔中添加浓度为100 μg/ml的猪粉尘,或仅为培养基作为对照,每四次重复,分别孵育2、4、6、8、10和24小时。收集所有培养上清样品,在1000下离心gydF4y2BaggydF4y2Ba保存10分钟以去除细胞碎片,并在-70°C保存,直到分析。使用商业Quantikine免疫分析法,用ELISA技术分析IL-8的产生,并表达为ng/10gydF4y2Ba6gydF4y2Ba巨噬细胞。用排除台盼蓝的方法测定细胞活力。gydF4y2Ba
统计分析gydF4y2Ba
结果以平均值(SE)表示。各因子分别采用Fisher保护最小显著性差异(PLSD)方差分析(ANOVA)和未配对方差分析进行统计学比较gydF4y2BatgydF4y2Ba测试物质之间。p值<0.05被认为是显著的。gydF4y2Ba
结果gydF4y2Ba
a549上皮细胞产生Il-8gydF4y2Ba
猪尘溶液的内毒素含量为2.16 (0.20)ng/100 μg猪尘和0.53 (0.04)ng/100 μg谷粒。在无血清条件下,未受刺激的A549细胞中IL-8的基线产量为0.71 (0.12)ng/million细胞,而在无血清条件下为0.26 (0.03)ng/million细胞(p = 0.77)。猪灰、LPS和葡聚糖以剂量依赖的方式刺激肺上皮细胞产生IL-8,在最高剂量(100 μg/ml,图)时,猪灰的作用大约是LPS的5倍(p<0.001),是葡聚糖的15倍(p<0.001)gydF4y2Ba1gydF4y2Ba然而,尽管葡聚糖是最弱的刺激,但与对照相比,在最高浓度时,其IL-8的产量几乎翻了一番(90%)。粉尘对IL-8的产生没有显著影响。在血清存在和不存在的情况下,猪灰和LPS都能显著刺激IL-8的产生gydF4y2Ba1gydF4y2BaB).在两种最高浓度下(10 μg/ml, p<0.05;100 μg/ml, p<0.001),而在无血清和有血清的情况下,用猪灰孵育细胞,细胞活性无显著差异。在相同制备的猪灰或测试成分的无细胞制剂中没有IL-8活性。LPS与葡聚糖或颗粒粉尘(10 μg/ml)孵育相比,在相同浓度下仅用一种成分孵育不会增加IL-8的产量(数据未显示)。在与所有浓度的所有药剂孵育后,活性(排除台锥蓝)保持不变(>90%)。gydF4y2Ba

平均(SE) IL-8产量(ng)每百万上皮细胞(A549)。每个数据点基于1-3次重复或四次重复的实验。(A)猪尘(F = 83.2;p<0.001)、脂多糖(LPS) (F = 38.1;p<0.001),葡聚糖(F = 4.9;p<0.01)诱导A549上皮细胞中IL-8的产生显著的剂量依赖性增加。粉尘不影响IL-8的释放(F = 2.3;P = 0.12)。与对照组相比,LPS在10 μg/ml (p<0.01)和100 μg/ml (p<0.001)时都能刺激IL-8的产生。与对照组相比,在最高浓度(100 μg/ml)下,猪粉(p<0.001)和葡聚糖(p<0.01)也能促进IL-8的产生。 (B) Comparison between incubations of swine dust and LPS with or without serum.
nhbe细胞产生Il-8gydF4y2Ba
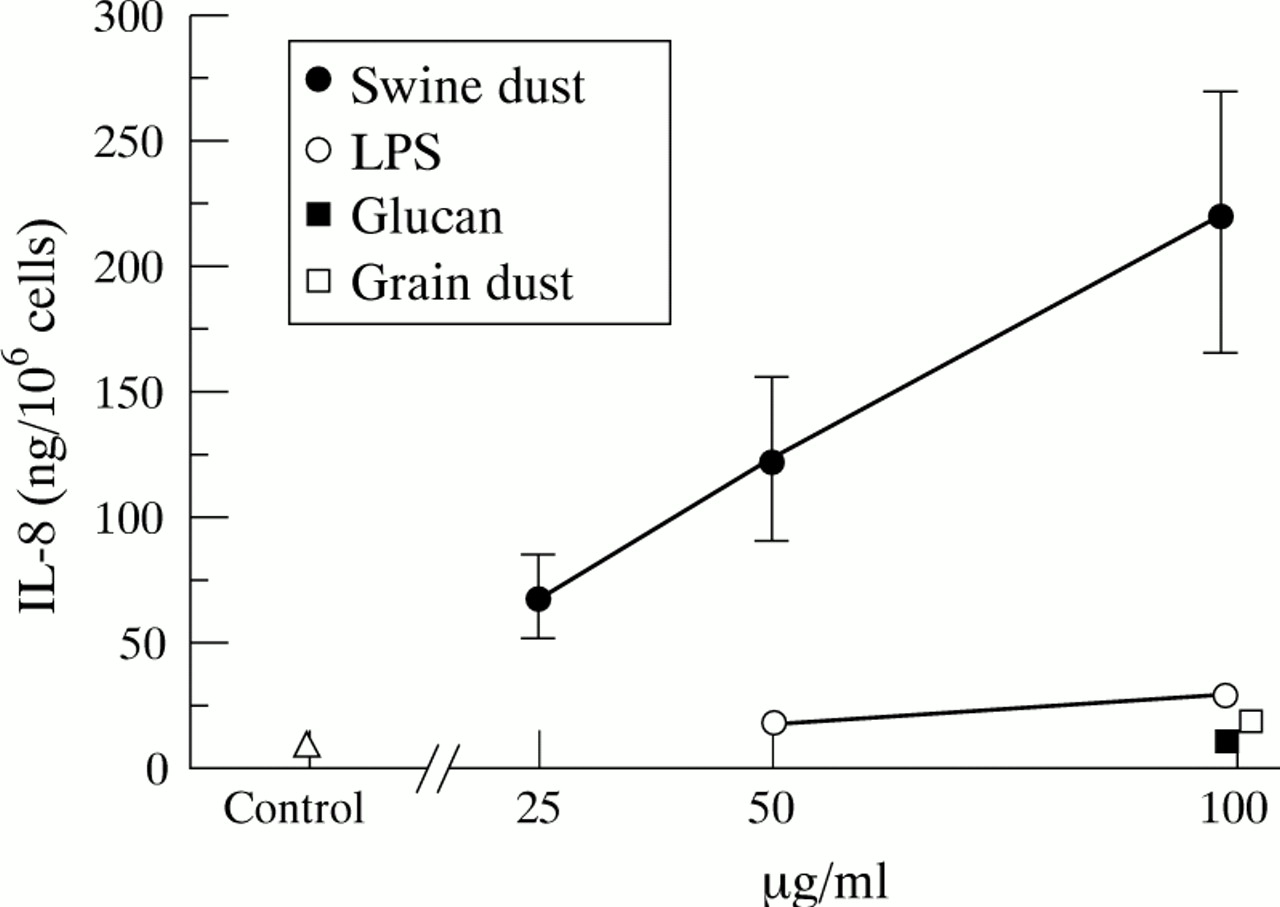
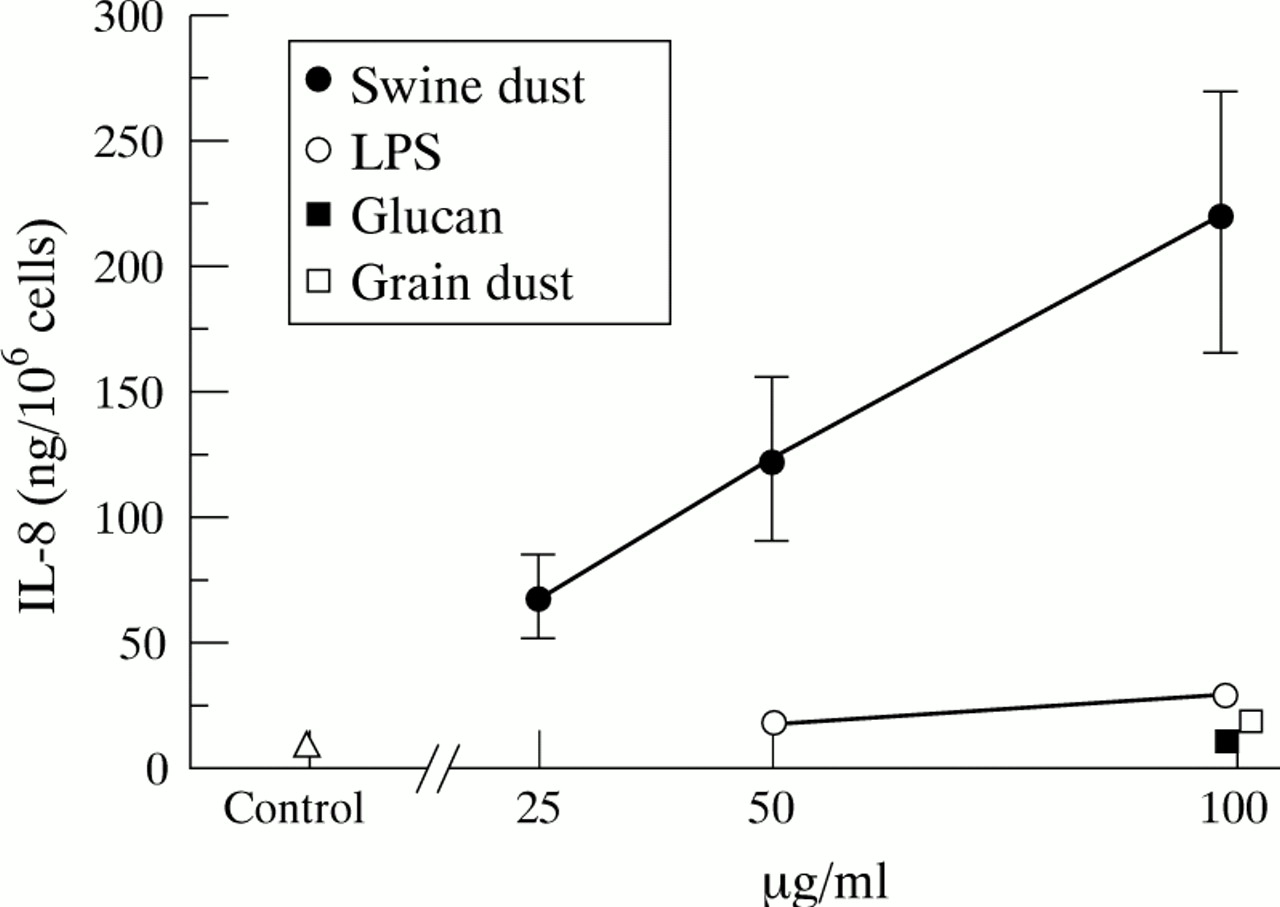
NHBE细胞中IL-8的基线产量为10.8 (1.7)ng/百万细胞。猪尘刺激NHBE细胞以线性剂量依赖方式产生IL-8(图gydF4y2Ba2gydF4y2Ba;50 μg/ml, p<0.05;100 μg/ml, p<0.001)。与对照组相比,最高浓度猪粉使IL-8产量增加20倍,显著高于其他测试物质(p<0.01)。100 μg/ml时,LPS诱导IL-8释放增加170%,颗粒粉尘增加70% (p分别<0.001和<0.05)。在对照组(p<0.001)和用猪粉(100 μg/ml)刺激后,NHBE细胞比A549细胞产生IL-8的能力更高(10-15倍)。p < 0.001)。在任何浓度下,所有试验药物均未改变台锥蓝排除法的细胞活力(活力>90%)。gydF4y2Ba

平均(SE)每百万正常人支气管上皮细胞IL-8产量(ng)。每个数据点都基于三个重复的实验。猪尘(F = 7.9;p<0.001)、脂多糖(LPS) (F = 7.4;p<0.01)和粒尘(F = 5.9;p<0.05)增加IL-8的产生。葡聚糖不影响IL-8的释放(F = 0.1;P = 0.82)。与对照组相比,猪尘在50 μg/ml (p<0.05)和100 μg/ml (p<0.001)时都能促进IL-8的产生。与对照组相比,在最高浓度(100 μg/ml)下,LPS (p<0.001)和粉尘(p<0.05)也能刺激IL-8的产生。gydF4y2Ba
肺泡巨噬细胞产生Il-8gydF4y2Ba
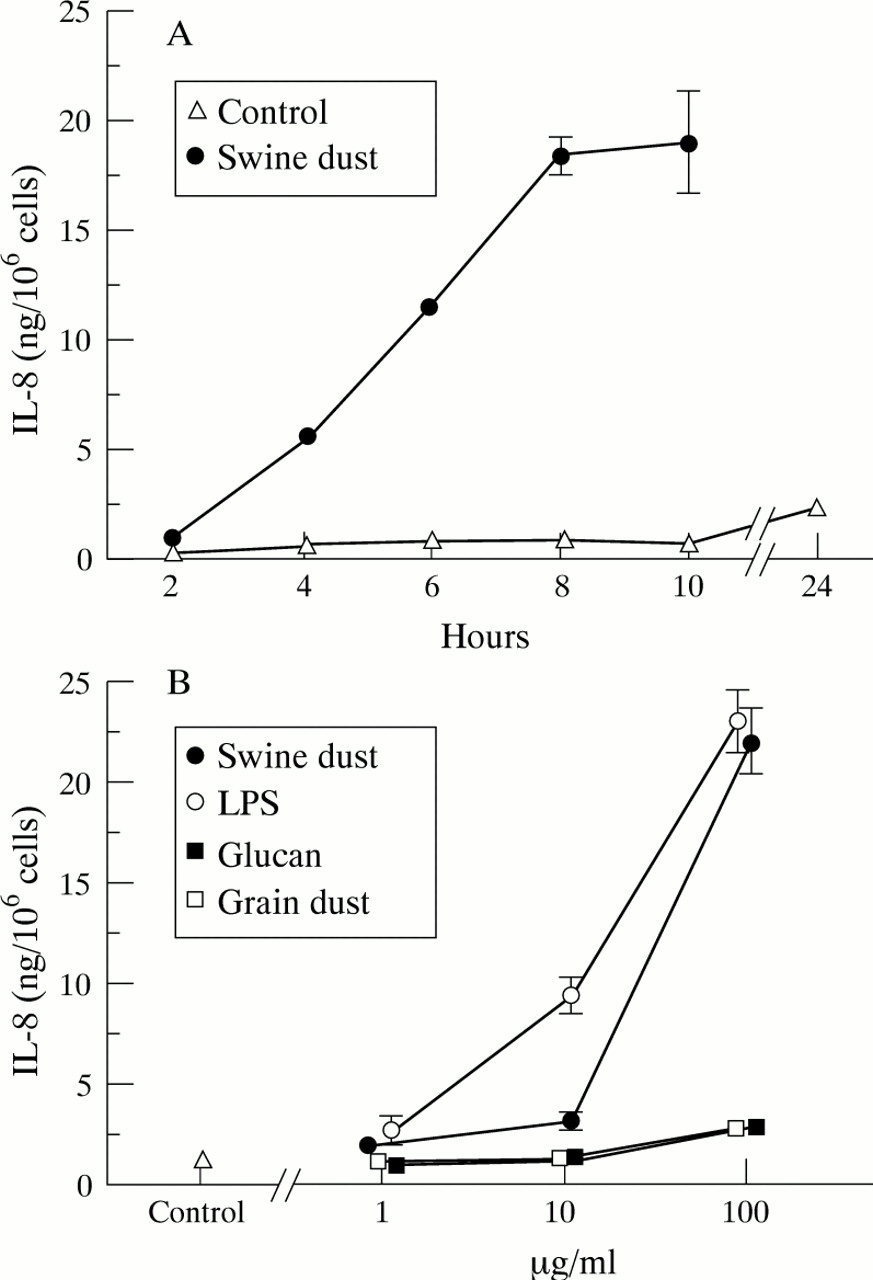
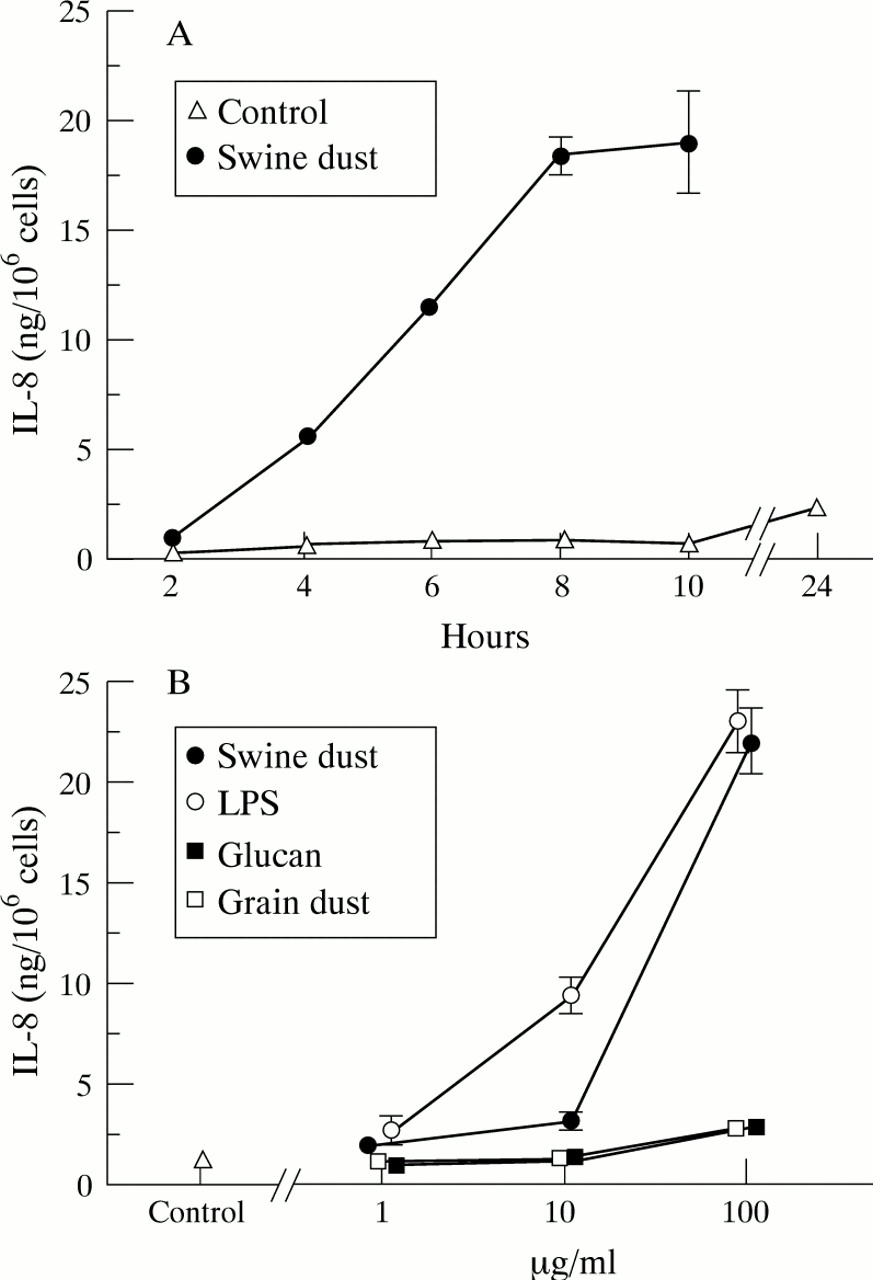
未受刺激巨噬细胞中IL-8的基线产量为1.0 (0.09)ng/百万巨噬细胞。在控制条件下和与猪瘟孵育后,IL-8的产量随着时间的推移而增加,但在所有测试时间点,刺激猪瘟导致IL-8产量显著增加(图2)gydF4y2Ba3.gydF4y2Ba一个;p < 0.001)。猪粉尘、LPS、葡聚糖和谷物粉尘以剂量依赖的方式增加肺泡巨噬细胞中IL-8的产生(图gydF4y2Ba3.gydF4y2BaB).在最高浓度(100 μg/ml)时,猪尘和LPS均能刺激细胞产生更多的IL-8 (p<0.001)。然而,尽管葡聚糖和谷物粉尘的刺激比猪粪和LPS弱得多,但与对照组相比,在最高浓度下,它们的IL-8产量几乎翻了一番(90%)。在与所有浓度的药剂孵育后,活性保持不变(>90%台盼蓝除外)。gydF4y2Ba

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
每百万巨噬细胞平均(SE) IL-8产量(ng)。(A) IL-8随时间的产生(n = 4次观测)。IL-8的产生随着时间的推移而增加,无论是在未受刺激的控制情况下(F = 19.4;p<0.001)和用猪灰刺激后(F = 97.9;p < 0.001)。猪尘刺激24小时后IL-8浓度为113.1 (10.1)ng/million cells,图中未显示。(B)猪尘(F = 131.3;p<0.001)、脂多糖(LPS) (F = 105.4;p<0.001),颗粒粉尘(F = 5.5;p<0.01)和葡聚糖(F = 11.0; p<0.001) stimulated alveolar macrophages to produce IL-8 in a dose-dependent manner. LPS stimulated IL-8 production at both 10 μg/ml (p<0.001) and 100 μg/ml (p<0.001) compared with the control. At the highest concentration tested (100 μg/ml) swine dust (p<0.001), glucan (p<0.001), and grain dust (p<0.01) also stimulated IL-8 production compared with the control (n = 8 observations with exception of swine dust 1 μg/ml where n = 7).
讨论gydF4y2Ba
在目前的研究中,猪尘是肺上皮细胞(NHBE和A549)和肺泡巨噬细胞中产生IL-8的有效刺激物,而NHBE细胞是这方面最有效的IL-8分泌物。两种上皮细胞类型产生IL-8能力的差异可能是由于A549细胞构成了癌细胞系,而NHBE细胞是原代培养的上皮细胞。目前尚不清楚癌细胞系中上皮细胞的功能能力在多大程度上被保存,可以预期NHBE细胞在体内更类似于正常的支气管上皮细胞。然而,A549细胞仍然表现出与NHBE细胞相似的行为方式,表明细胞系细胞具有相当的代表性。NHBE细胞和巨噬细胞孵育时间的差异(分别为24小时和8小时)可以部分解释这两种细胞类型的不同能力。然而,用猪灰刺激巨噬细胞24小时后,IL-8的产量增加了8小时后的5倍以上,但巨噬细胞的产量仍然只有NHBE细胞的一半左右。gydF4y2Ba
猪尘含有含有微生物的粪便物质,主要是革兰氏阴性和革兰氏阳性属的细菌。gydF4y2Ba12gydF4y2Ba,gydF4y2Ba17gydF4y2Ba脂多糖是存在于革兰氏阴性菌壁的内毒素的组成部分。症状、BAL液和血清中的IL-6反应和暴露于猪尘中内毒素含量的支气管反应的增加之间存在相关性。gydF4y2Ba10gydF4y2Ba体外研究LPS诱导支气管上皮细胞释放IL-8的能力产生了相互矛盾的结果。gydF4y2Ba15gydF4y2Ba,gydF4y2Ba18 - 20gydF4y2Ba
LPS对A549和NHBE细胞中IL-8释放的刺激作用弱于猪灰。LPS对上皮细胞的弱作用可以解释为这些细胞上缺乏LPS受体(清道夫受体,CD14和CD18)。gydF4y2Ba21gydF4y2BaCD14识别LPS与血清蛋白脂多糖结合蛋白(LBP)的复合物,并似乎参与了对LPS的摄取和合成反应。CD14以两种形式存在,膜结合(mCD14)或可溶性(sCD14)。gydF4y2Ba22gydF4y2Ba用LPS刺激A549细胞,在有血清(理论上可以同时包含LBP和sCD14)和无血清的情况下孵育,而NHBE细胞在无血清的条件下孵育。然而,在无血清条件下,LPS刺激A549细胞对IL-8产生的刺激程度甚至高于有血清的情况,这表明血清联合LPS可能对IL-8产生抑制作用,而不是刺激作用。gydF4y2Ba
在肺泡巨噬细胞中,LPS和猪尘在最高浓度下产生相同的反应。与上皮细胞不同,肺泡巨噬细胞具有LPS受体(CD14)gydF4y2Ba21gydF4y2Ba这可能解释了巨噬细胞对LPS的反应。而添加到细胞中的猪灰的内毒素含量为2.16 ng/100 μg猪灰。这表明,在最高浓度的猪灰中LPS含量低于使用的最低LPS浓度(1 μg/ml),这表明猪灰中除LPS外的其他成分对IL-8的产生有刺激作用。gydF4y2Ba
早期的研究表明,吸入猪尘后,肽聚糖(革兰氏阳性菌壁的主要成分,在革兰氏阴性菌中也少量存在)与血液和BAL液中中性粒细胞数量增加之间存在相关性。gydF4y2Ba11gydF4y2Ba,gydF4y2Ba23gydF4y2Ba我们还发现革兰氏阴性细菌和革兰氏阳性细菌都是A549细胞中IL-8释放的有效刺激物gydF4y2Ba24gydF4y2Ba还有肺泡巨噬细胞。gydF4y2Ba25gydF4y2Ba革兰氏阳性细菌产物也被证明能刺激上皮细胞中IL-8的产生。gydF4y2Ba24gydF4y2Ba,gydF4y2Ba26gydF4y2Ba革兰氏阳性菌也可能是猪尘的重要组成部分,有助于炎症作用。这些实验有力地支持了一种假设,即革兰氏阴性和革兰氏阳性细菌参与了吸入猪尘后的炎症性气道反应。gydF4y2Ba
谷物粉尘可能与猪粉尘吸入的促炎活性有关。吸入含水颗粒粉尘提取物引起急性炎症反应,中性粒细胞浓度增加,细胞因子(IL-6, IL-1β, IL-8和TNF-α)水平升高。gydF4y2Ba27gydF4y2Ba在本研究中,颗粒粉尘刺激上皮细胞和肺泡巨噬细胞产生IL-8,但以重量计算,其程度远低于猪粉尘。gydF4y2Ba
我们之前发现,吸入猪尘会导致上、下气道灌洗液中IL-1β和TNF-α水平升高。gydF4y2Ba10gydF4y2BaTNF-α似乎启动细胞因子级联。人鼻和支气管上皮细胞在原代培养中表达和分泌IL-8, IL-1和TNF-α增加了这些细胞mRNA的表达和分泌。gydF4y2Ba3.gydF4y2Ba,gydF4y2Ba28gydF4y2Ba因此,IL-8对猪灰的反应可能部分是由TNF-α介导的,尽管它不太可能是唯一的机制。这些发现也表明,粉尘中的因子可能不仅直接与上皮细胞相互作用以增加IL-8的产生,还可能通过其他细胞因子间接作用。gydF4y2Ba
总之,本研究表明,猪尘是肺上皮细胞和肺泡巨噬细胞产生IL-8的有效刺激因子。LPS在测试的两种细胞类型中具有不同的效力,这表明几种细胞类型可能通过不同的机制促进气道中IL-8的产生。猪灰中内毒素含量较低,葡聚糖和谷粒粉尘对IL-8释放影响较弱,提示猪灰中其他成分可能参与气道炎症反应。gydF4y2Ba
致谢gydF4y2Ba
这项研究得到了瑞典工作生活研究委员会(94-1383)、国家工作环境基金(AMFO 90-1680, AMFO 94-0318)、瑞典农民农业研究基金会和瑞典心肺基金会的资助。作者要感谢Siw Siljerud的技术援助。gydF4y2Ba
参考文献gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba