


摘要gydF4y2Ba
已知p38丝裂原活化蛋白激酶(MAPK)信号在慢性阻塞性肺疾病(COPD)巨噬细胞中增加。我们研究了p38 MAPK抑制剂的作用gydF4y2BaNgydF4y2Ba含氰基的- - - - - -gydF4y2BaNgydF4y2Ba”——(2 - {[8 - (2,6-difluorophenyl) 4 (4-fluoro-2-methylphenyl) 7-oxo-7 8-dihydropyrido [2, 3 -gydF4y2BadgydF4y2Ba[-嘧啶-2-基]氨基}乙基)胍(SB706504)和地塞米松对COPD巨噬细胞炎症基因表达和蛋白分泌的影响。我们还研究了SB706504联合地塞米松治疗的效果。用地塞米松和/或SB706504培养脂多糖(LPS)刺激的单核细胞来源的巨噬细胞(MDMs)和肺泡巨噬细胞(AMs)。MDMs用于基因阵列和蛋白质研究,而AMs用于肿瘤坏死因子(TNF) α蛋白的产生。SB706504在COPD MDMs中引起一系列细胞因子和趋化因子的转录抑制。与单独使用SB706504(-2.04)或地塞米松(-3.39)相比,SB706504联合使用地塞米松对基因表达的抑制(-8.90)更大。23个基因对两种药物的作用不敏感,包括白细胞介素(IL)-1β、IL-18和趋化因子(CC基序)配体(CCL) 5。此外,4号染色体趋化因子簇成员CXCL1、CXCL2、CXCL3和CXCL8均具有糖皮质激素抗性。SB706504显著抑制lps刺激的COPD和吸烟者am的TNFα产生,与地塞米松联合治疗时,抑制作用接近最大。我们得出结论,SB706504靶向炎性巨噬细胞基因的一个子集,当与地塞米松一起使用时,可有效抑制这些基因。 SB706504 and dexamethasone had no effect on the transcription of a subset of LPS-regulated genes, including IL-1β, IL-18, and CCL5, which are all known to be involved in the pathogenesis of COPD.
慢性阻塞性肺疾病(COPD)的特征是进行性气流阻塞和气道炎症(gydF4y2Ba巴恩斯,2003gydF4y2Ba).肺泡巨噬细胞(AMs)被认为通过分泌促炎细胞因子和趋化因子在疾病发病过程中发挥核心作用(gydF4y2Ba巴恩斯,2003gydF4y2Ba).糖皮质激素(GCs)是COPD中应用最广泛的抗炎药物。GC通过与糖皮质激素受体(GR)形成复合物,抑制核因子(NF)-κB等转录因子的功能,从而抑制炎症基因的转录,这一过程被称为转抑制(gydF4y2BaGlass and Ogawa, 2006gydF4y2Ba).然而,GC在COPD患者中的临床获益并不大(gydF4y2Ba索里亚诺等人,2007gydF4y2Ba),据报道,慢性阻塞性肺疾病AMs对细胞因子产生的抑制具有gc抗性(gydF4y2BaCulpitt et al., 2003gydF4y2Ba;gydF4y2BaCosio et al., 2004gydF4y2Ba).慢性阻塞性肺病患者需要其他抗炎疗法(gydF4y2Ba巴恩斯,2006gydF4y2Ba).gydF4y2Ba
细胞外刺激如toll样受体(TLR) 4配体、脂多糖(LPS)和细胞因子激活p38丝裂原活化蛋白激酶(MAPK)细胞内信号传导。该信号通路上调促炎细胞因子和趋化因子的产生(gydF4y2BaZarubin and Han, 2005gydF4y2Ba)通过激活转录因子如NFκB和激活转录因子2或改变染色质结构,使NFκB与炎症基因的启动子区域结合(gydF4y2BaSaccani et al., 2002gydF4y2Ba).p38 MAPK也可能通过mRNA稳定发挥转录后作用(gydF4y2BaWinzen et al., 1999gydF4y2Ba)或蛋白质翻译水平(gydF4y2Ba牛顿和霍尔顿,2003gydF4y2Ba;gydF4y2BaBrook et al., 2006gydF4y2Ba).活化p38 MAPK在COPD患者肺巨噬细胞中的表达升高(gydF4y2BaRenda et al., 2008gydF4y2Ba),提示该信号通路与COPD的病理生理有关。因此,p38 MAPK抑制剂正处于临床开发阶段,用于治疗COPD (gydF4y2Ba巴恩斯,2006gydF4y2Ba).在THP-1细胞和健康人AMs中,p38 MAPK抑制剂SB203580和2,1(1,3-二羟基丙基-2-基)-4-(4-氟苯基)-5-(2-苯氧嘧啶-4-基)咪唑抑制lps刺激的细胞因子蛋白水平,但对细胞因子mRNA水平影响很小或没有影响(gydF4y2BaBirrell et al., 2006gydF4y2Ba).这表明这些p38 MAPK抑制剂在巨噬细胞中作用于蛋白质翻译水平而不是基因转录水平。gydF4y2Ba
GCs通过增加MAPK磷酸酶-1基因表达抑制p38 MAPK活性(gydF4y2BaLasa等人,2002gydF4y2Ba).此外,p38 MAPK抑制剂可增强GC (gydF4y2BaIrusen et al., 2002gydF4y2Ba)通过改变GR的磷酸化(gydF4y2BaSzatmáry et al., 2004gydF4y2Ba).这些潜在的协同作用为这两类药物一起使用以最大限度地发挥抗炎作用提供了良好的理论依据。gydF4y2Ba
在对照组和肺气肿患者的AMs中,p38 MAPK抑制剂SB239063和SD-282显著抑制lps诱导的肿瘤坏死因子(TNF)-α蛋白水平,但对白细胞介素(IL)-8(也称为CXCL8)和粒细胞巨噬细胞集落刺激因子(GM-CSF)的影响很小,表明p38 MAPK抑制剂的作用因炎症基因而异(gydF4y2BaSmith et al., 2006gydF4y2Ba).同样,配体激活的GR并不针对所有炎症基因,而是抑制具有GR依赖性转录激活的炎症基因亚群的活性(gydF4y2BaGlass and Ogawa, 2006gydF4y2Ba).这种现象是细胞类型特有的,并取决于所使用的刺激类型(gydF4y2BaOgawa et al., 2005gydF4y2Ba).评估和比较GC和p38 MAPK抑制剂对COPD巨噬细胞炎症基因的敏感性具有重要意义。gydF4y2Ba
p38 MAPK抑制剂gydF4y2BaNgydF4y2Ba含氰基的- - - - - -gydF4y2BaNgydF4y2Ba”——(2 - {[8 - (2,6-difluorophenyl) 4 (4-fluoro-2-methylphenyl) 7-oxo-7 8-dihydropyrido - [2, 3gydF4y2BadgydF4y2Ba嘧啶-2-基氨基乙基胍,也称为PCG,对α和β p38 MAPK亚型具有高特异性,对其他激酶的活性较低(gydF4y2BaTudhope et al., 2008gydF4y2Ba).这种新型的MAPK抑制剂已被证明可以减少lps刺激的COPD巨噬细胞的TNFα、GM-CSF和IL-6的产生,而对IL-8的产生影响较小,这支持了先前的数据,即一些炎症细胞因子对p38 MAPK抑制剂不太敏感(gydF4y2BaSmith et al., 2006gydF4y2Ba).我们希望进一步扩展这一观察,以分析一系列炎症基因,以鉴定p38 mapk敏感和-不敏感基因。此外,我们有兴趣了解这种药物在基因转录水平上的作用,以及当与糖皮质激素一起使用时的作用。gydF4y2Ba
本文报道GC地塞米松和SB706504对COPD患者巨噬细胞炎症基因LPS激活的影响。我们使用来自COPD患者的单核细胞源性巨噬细胞(MDMs)进行基因阵列研究,目的如下:1)确定SB706504是否抑制炎症基因的转录,2)鉴定lps激活的炎症基因对SB706504或地塞米松不敏感。我们还研究了SB706504与地塞米松联合治疗的效果。定量PCR和蛋白分析证实了基因阵列的发现。最后,我们利用lps刺激COPD患者和健康吸烟者的AMs进行实验,评估SB706504和地塞米松联合治疗的效果。gydF4y2Ba
材料与方法gydF4y2Ba
病人。gydF4y2Ba根据现行指南诊断为慢性阻塞性肺病的6名受试者(gydF4y2Ba黄金,2008gydF4y2Ba)提供血液样本进行MDM细胞培养和转录组学分析。这些患者在空气中的血氧饱和度均为0.93%,且未使用辅助氧疗。使用TLR2和4激动剂LPS和特异性TLR4激动剂超纯(UP)-LPS (gydF4y2BaWakelin et al., 2006gydF4y2Ba)的数据来源于11例诊断为慢性阻塞性肺病的患者和16例肺功能正常的吸烟者,这些患者因怀疑或确诊肺癌而接受手术切除。前吸烟者的定义是戒烟至少1年。本研究中所有患者的人口统计资料见表gydF4y2Ba1gydF4y2Ba和gydF4y2Ba2gydF4y2Ba当地研究伦理委员会批准了本研究,并获得了每位患者的书面知情同意书。gydF4y2Ba
MDM研究中使用的患者人口统计数据结果以均数(sd)表示。gydF4y2Ba
在AM研究中使用的患者的人口统计学结果表示为平均值(SD)。gydF4y2Ba
MDM文化gydF4y2Ba
如前所述,从全血中分离外周血单个核细胞(gydF4y2BaStengel et al., 1998gydF4y2Ba;gydF4y2BaKent et al., 2008gydF4y2Ba).这些培养物先前已被证明完全由CD68+巨噬细胞组成(gydF4y2BaStengel et al., 1998gydF4y2Ba),我们使用流式细胞术在三名受试者的MDMs中获得了类似的数据(数据未显示)。在加入药物(1ml /孔)、100 nM地塞米松(Sigma-Aldrich, St. Louis, MO)和/或之前储存在DMSO中的3 μM SB706504 (GlaxoSmithKline赠送)之前,从MDMs中去除培养基。这个浓度是根据以前的数据选择的(gydF4y2BaTudhope et al., 2008gydF4y2Ba),表明该化合物对p38的活化作用在10 μM时达到最大。将细胞与药物预孵育30 min后,用对照或LPS (1 μg/ml,gydF4y2Ba产败沙门氏菌gydF4y2Ba;Sigma-Aldrich)媒体。在对照培养基中以1:1265 μl的比例加标DMSO以模拟抑制剂的最高浓度。细胞在37°C和5% CO下保存gydF4y2Ba2gydF4y2Ba.gydF4y2Ba
我的文化gydF4y2Ba
取离肿瘤较远的肺组织,灌注无菌0.15 M NaCl分离巨噬细胞。取回的液体离心10分钟,400gydF4y2BaggydF4y2Ba室温),在RPMI 1640培养基中重悬。细胞悬液在Ficoll梯度上悬浮,离心30min, 400gydF4y2BaggydF4y2Ba4°C),台盼蓝排斥法计数细胞。然后将细胞离心,以1 × 10的浓度重悬gydF4y2Ba6gydF4y2BaRPMI 1640培养基中添加10%胎牛血清(Invitrogen, Paisley,英国),1%青霉素/链霉素(Sigma Chemical, Poole, Dorset,英国)和1%gydF4y2BalgydF4y2Ba谷氨酰胺(表达载体)。在96孔板中,0.1 × 10gydF4y2Ba6gydF4y2Ba细胞/孔播种,放置于37℃,5% COgydF4y2Ba2gydF4y2Ba至少18小时,以使AMs粘附在板上。非贴壁细胞在使用前用补充的RPMI 1640培养基洗涤去除。免疫组织化学实验显示,这些细胞有>90%的CD68表达(方法详情见补充数据)。去除培养基后,加入200 μl添加100 nM地塞米松和/或3 μM SB706504的RPMI 1640培养基。AMs与药物预孵育30 min后,与对照、LPS (1 μg/ml,gydF4y2Ba产败沙门氏菌gydF4y2Ba;Sigma-Aldrich)或UP-LPS (1 μg/ml;InvivoGen,圣地亚哥,加州)媒体。对照培养基以1:1265 μl的比例加标DMSO,模拟药物的最高浓度。在37°C和5% CO的条件下保持gydF4y2Ba2gydF4y2Ba.gydF4y2Ba
转录组分析gydF4y2Ba
RNA分离和芯片分析。gydF4y2Ba细胞在TRIzol (Invitrogen)中裂解,RNA分离、定量和质量评估以及Affymetrix微阵列(Affymetrix, Santa Clara, CA)分析的方案如前面所述(gydF4y2BaKent et al., 2008gydF4y2Ba).gydF4y2Ba
定量PCR。gydF4y2Ba用250-ng的总RNA分两份进行qPCR。根据制造商的说明,使用ABI高容量cDNA存档试剂盒(应用生物系统公司,福斯特城,CA)将RNA转化为cDNA。6 μ g cDNA与ABI 2x TaqMan PCR混合物在10 μl反应中反应,其中0.4 μl合适的正向和反向引物(10 μl)和0.2 μl TaqMan探针(5 μl) (Genosys, Cambridge, UK)。引物和TaqMan探针的序列见补充表1和2。在ABI 7900序列检测系统(Applied Biosystems)上进行热循环,温度为:50°C 10分钟,95°C 10分钟,92°C 15秒,60°C 1分钟,循环45次。家政基因,β-肌动蛋白,甘油醛-3-磷酸脱氢酶和亲环蛋白,被用来评估任何负荷影响。定量相对于标准曲线(基因组DNA)根据制造商的说明。管家基因的表达在所有样本中都是一致的(数据未显示)。gydF4y2Ba
上清蛋白分析gydF4y2Ba
TNFα和IL-8酶联免疫吸附试验。gydF4y2Ba根据制造商的说明书(R&D Systems Europe Ltd, Abingdon, Oxfordshire, UK),对细胞培养上清进行TNFα和IL-8的夹心酶联免疫吸附试验(elisa)。上清液用RPMI 1640稀释,以适应ELISA分析。gydF4y2Ba
Luminex分析。gydF4y2Ba补充资料中提供了微球抗体偶联和Luminex上清蛋白分析IL-6、IL-10、干扰素γ诱导蛋白10和tnf - α的方法。该程序的范围为2.4至10,000 pg/ml;因此,在RPMI 1640中适当稀释样品。所有低于检测下限的数据点的值为下限的一半(gydF4y2BaLaan et al., 2002gydF4y2Ba).gydF4y2Ba
统计分析gydF4y2Ba
使用SIMCA-P+软件(Umetrics, Windsor, UK)通过主成分分析评估基因阵列数据质量的质量控制指标的同质性。基因表达的全局分析最初是通过使用Rosetta Resolver (Rosetta Inpharmatics LLC, Seattle, WA)规范化探针强度数据进行的。gydF4y2Ba翁等,2006gydF4y2Ba),然后加载到SIMCA-P+中,通过基因表达主成分分析来直观评估关键趋势。使用SAS软件(SAS Institute, Cary, NC)对强度进行混合模型方差分析。实验条件作为一个固定因素,供体作为一个随机因素。保留探针集以供进一步分析gydF4y2BapgydF4y2Ba至少3例患者< 0.05。采用邓尼特事后检验将结果与对照组进行比较。所有带有gydF4y2BapgydF4y2Ba≤0.01,2倍变化为显著。探针集基因名称从gydF4y2Bahttp://www.affymetrix.comgydF4y2Ba.gydF4y2Ba
细胞因子和趋化因子基因鉴定采用独创性途径分析(Ingenuity Pathways Analysis, IPA);智慧系统,gydF4y2Bahttp://www.ingenuity.comgydF4y2Ba).IPA还用于鉴定相互作用的基因网络(gydF4y2BaJuric et al., 2007gydF4y2Ba;gydF4y2BaLi et al., 2007gydF4y2Ba;gydF4y2BaSavli et al., 2008gydF4y2Ba).该程序还包括不在上传的基因列表中,但与上传的基因高度相关的基因或复合物。每个基因或复合体被归类为一个节点,相互作用的节点由直接关系(需要直接身体接触)或间接关系(不需要直接身体接触)定义。同时利用国家生物技术信息中心数据库对Man数据库中的基因功能和在线孟德尔遗传进行人工检索(gydF4y2Bahttp://www.ncbi.nlm.nih.gov/sites/entrez?db=omimgydF4y2Ba)用于评估基因染色体位点。使用Heatmap Builder 1.0版本生成热图(Euan Ashley博士,斯坦福大学,Palo Alto, CA)。gydF4y2Ba
RNA和蛋白质数据采用非参数方差分析(Friedman's检验与Dunn's事后分析)进行分析,随后采用Wilcoxon配对对符号秩单尾检验来检验处理之间的差异。采用Mann-Whitney检验评估两组(COPD患者和吸烟者)之间的差异。采用Spearman秩相关系数评估阵列与qPCR数据之间的关系。使用GraphPad InStat软件(GraphPad Software Inc., San Diego, CA)进行测试。gydF4y2Ba

lps刺激的COPD MDMs中对地塞米松、SB706504或联合治疗敏感的炎症介质mRNA水平的热图比较(gydF4y2BangydF4y2Ba= 6)。通过芯片检测mRNA水平。显著调节(至少2倍的变化)gydF4y2BapgydF4y2Ba< 0.01)的炎症基因被加载到Heatmap Builder Version 1.0中,折线图显示了每种情况下的平均折叠变化。gydF4y2Ba
结果gydF4y2Ba
MDM细胞培养gydF4y2Ba
基因表达芯片。gydF4y2Ba58种炎症介质在至少一种实验条件下(单独LPS或地塞米松和/或SB706504治疗)显著调节,>2和a的倍数变化gydF4y2BapgydF4y2Ba经IPA和手工基因交叉比对鉴定,均< 0.01gydF4y2Ba3.gydF4y2Ba,gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba).23个基因被LPS显著上调,而被地塞米松、SB706504和/或联合治疗抑制(gydF4y2Ba表3gydF4y2Ba).其中四个基因CXCL1、2、3和IL-8对地塞米松治疗有耐药性,但对联合治疗敏感(gydF4y2Ba表3gydF4y2Ba).在线Mendelian Inheritance in Man数据库对染色体位点的搜索也发现这些基因占据了相同的位点,4q12-q13。热图gydF4y2Ba图1gydF4y2Ba地塞米松与SB706504联合用药对这23个基因的抑制程度最大(-8.90),大于地塞米松单用(-3.39)或SB706504单用(-2.04)。gydF4y2Ba
地塞米松和sb706504反应性lps诱导的COPD MDMs炎症介质探针组gydF4y2Ba
地塞米松和sb706504不敏感lps诱导的COPD MDMs炎症介质探针组gydF4y2Ba
地塞米松和sb706504反应性炎症介质探针设置在COPD MDMs中,LPS没有影响或抑制表达gydF4y2Ba

基因网络通过独创性路径分析(独创性系统),gydF4y2Bahttp://www.ingenuity.comgydF4y2Ba),来源于lps刺激的COPD MDMs中地塞米松(A)-和SB706504 (B)-不敏感的炎症介质基因。每个基因或复合体被归类为一个节点;v形淋巴结定义炎症介质,圆形淋巴结定义复合物或分子群。图中所示的基团/配合物定义如下:IL1 (IL18、IL1β、IL1F5/6/7/8/10、IL1RN)、p38 MAPK (p38 MAPKα/β/δ/γ)、NFκB (NFκB、NFκB-RelA、Rel/RelA/RelB)、NFκB-RelA (NFκB + RelA或p50/52 + p65)、AP-1 (Jun, Fos)。紫色阴影的节点来自原始上传的数据集,由于分子之间的高水平连接,未着色的节点被IPA纳入网络。交互节点由直接关系(需要直接物理接触)(闭合箭头)或间接关系(不需要直接物理接触)(虚线箭头)定义。箭头方向给出了相互作用的方向,即A“作用于”B。gydF4y2Ba
另外23个基因被LPS诱导,但对任何药物治疗都不敏感(gydF4y2Ba表4gydF4y2Ba).10个基因未被LPS诱导,但对地塞米松和/或SB706504敏感,另外2个基因被LPS抑制,并被地塞米松和/或SB706504诱导(gydF4y2Ba表5gydF4y2Ba).gydF4y2Ba
IPA分析应用于27个地塞米松不敏感基因,以评分为中心的方法创建一个相关基因网络(gydF4y2Ba图2一个gydF4y2Ba).关联程度最高的基因是IL-8、IL-1β、IL-18和CCL5。与gc不敏感基因相关的信号通路有p38 MAPK、NFκB和AP-1。对30个sb706504不敏感基因(gydF4y2Ba图2 bgydF4y2Ba);IL-8、IL-1β和IL-18再次与其他药物不敏感基因高度相关,而涉及的信号通路如预期的p38 MAPK、NFκB,以及较小程度上的PI3激酶和Janus酪氨酸激酶/转录信号的信号转换器和激活器。gydF4y2Ba
qPCR。gydF4y2Ba在lps刺激的MDMs中,地塞米松或SB706504单独治疗可显著降低IL-1β、IL-6、GM-CSF、TNFα和IL-8 mRNA水平(gydF4y2Ba图3gydF4y2Ba).与SB706504相比,地塞米松对IL-1β、GM-CSF和TNFα mRNA水平的抑制作用更大。与地塞米松或SB706504单独治疗相比,地塞米松和SB706504联合治疗导致所有5个基因的mRNA水平进一步显著降低。地塞米松引起的IL-1β、IL-6、GM-CSF、TNFα和IL-8 mRNA表达水平的百分比下降见gydF4y2Ba表6gydF4y2Ba.这证实了微阵列发现IL-8和IL-1β是最不敏感的gc基因。qPCR与阵列变化的相关性非常密切,且具有统计学意义(gydF4y2BargydF4y2Ba= 0.91,gydF4y2BapgydF4y2Ba< 0.0001)。gydF4y2Ba
qPCR检测LPS刺激的copd MDMs中地塞米松和SB706504对炎症介质mRNA的抑制作用gydF4y2BapgydF4y2Ba< 0.05。gydF4y2Ba

地塞米松、SB706504及共治疗对lps刺激COPD MDMs细胞因子mRNA水平的影响(TaqMan qPCR)gydF4y2BangydF4y2Ba= 6)。数据以拷贝数中位数表示,有四分位间距和极差。*,gydF4y2BapgydF4y2Ba< 0.05;**gydF4y2BapgydF4y2Ba< 0.01;***,gydF4y2BapgydF4y2Ba< 0.001。gydF4y2Ba
浮在表面的蛋白质。gydF4y2Ba在lps刺激的MDMs中,单独使用地塞米松或SB706504治疗可显著降低IL-6、IL-10、干扰素γ诱导蛋白10和TNFα蛋白水平。gydF4y2Ba图4gydF4y2Ba).两种药物在ELISA中IL-8的测定值均有数值上的降低,但对于SB706504,这种差异没有达到统计学意义(gydF4y2Ba图5gydF4y2Ba).与地塞米松相比,SB706504对IL-10产生的抑制作用更大,但对其他细胞因子的抑制作用没有差异。与单独使用地塞米松或SB706504治疗相比,地塞米松和SB706504联合治疗导致所有这些细胞因子的蛋白水平进一步显著降低。gydF4y2Ba

比较地塞米松、SB706504和Luminex共治疗对lps刺激的COPD MDMs中细胞因子水平的抑制作用(gydF4y2BangydF4y2Ba= 6)。数据以中位数表示,有四分位间距和极差。*,gydF4y2BapgydF4y2Ba< 0.05;**gydF4y2BapgydF4y2Ba< 0.01;***,gydF4y2BapgydF4y2Ba< 0.001。gydF4y2Ba
AM细胞培养gydF4y2Ba
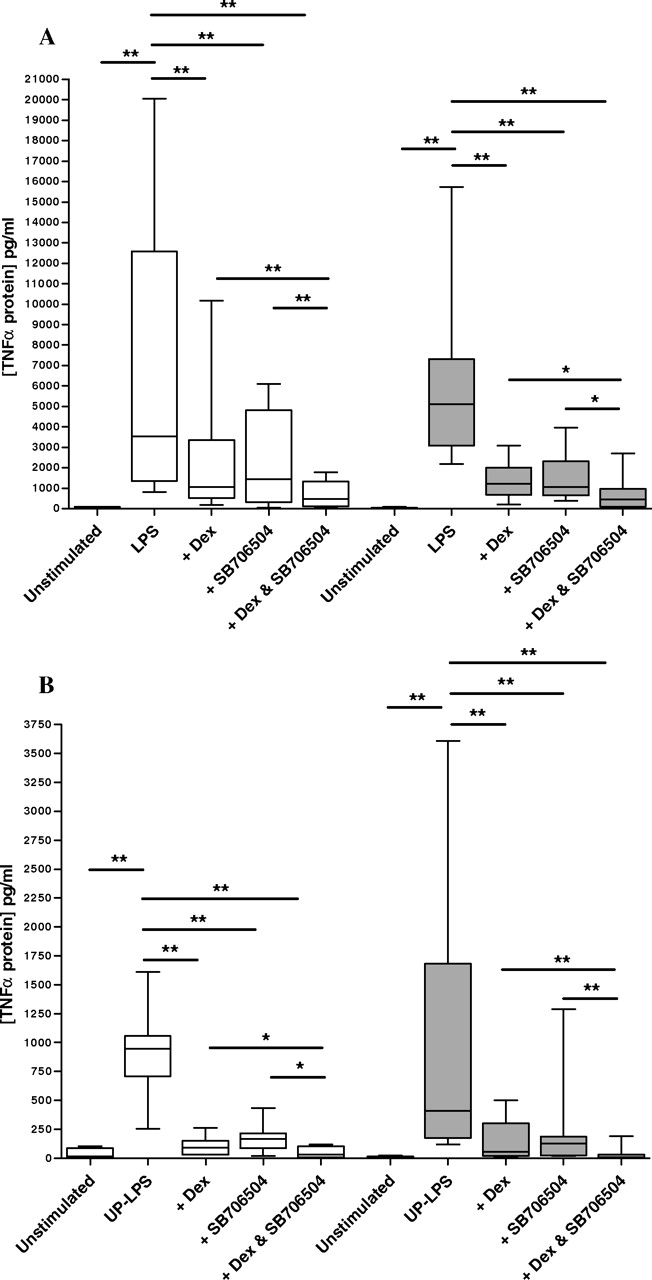
用LPS刺激10例正常肺功能吸烟者和9例COPD患者的AMs。单独使用SB706504和单独使用地塞米松可显著降低COPD患者的TNFα水平(中位抑制率为67.4%;gydF4y2BapgydF4y2Ba= 0.0098和77.1%;gydF4y2BapgydF4y2Ba= 0.002)。单独使用SB706504和单独使用地塞米松可显著降低吸烟者的TNFα生成(中位抑制率为76.1%;gydF4y2BapgydF4y2Ba= 0.001和78.9%;gydF4y2BapgydF4y2Ba= 0.002) (gydF4y2Ba图6gydF4y2Ba).地塞米松在COPD患者和吸烟者中的作用相似,而SB706504对吸烟者的抑制作用比COPD患者大(76.1 vs 67.4%)。gydF4y2BapgydF4y2Ba= 0.047)。联合治疗引起最大程度的TNFα抑制,中位抑制为91.1 (gydF4y2BapgydF4y2Ba= 0.002)和91.8% (gydF4y2BapgydF4y2Ba= 0.001)。gydF4y2Ba
观察到地塞米松和SB706504联合使用TLR2和4激动剂LPS的效果大于单独使用TLR4激动剂LPS的效果,我们随后在7名COPD患者和7名吸烟者中使用特异性TLR4激动剂UP-LPS评估了相同的现象(gydF4y2Ba图6gydF4y2Ba).单用地塞米松和单用SB706504对慢性阻塞性肺病患者TNFα产生的降低作用相似(中位抑制率为88.0%;gydF4y2BapgydF4y2Ba= 0.008和83.7%;gydF4y2BapgydF4y2Ba= 0.008)和吸烟者(中位抑制率为86.1%,gydF4y2BapgydF4y2Ba= 0.008和76.9%;gydF4y2BapgydF4y2Ba= 0.008)。当两种药物一起使用时,细胞因子产生的抑制接近最大;COPD患者的中位抑制率为94.6% (gydF4y2BapgydF4y2Ba= 0.008)和95.6% (gydF4y2BapgydF4y2Ba= 0.008)。这些药物的效果在两组之间没有差异(gydF4y2BapgydF4y2Ba> 0.05为所有比较)。gydF4y2Ba

地塞米松、SB706504和共治疗对未刺激和lps刺激的COPD MDMs中IL-8蛋白水平的抑制作用(gydF4y2BangydF4y2Ba= 6)。数据以中位数表示,有四分位间距和极差。*,gydF4y2BapgydF4y2Ba< 0.05;**gydF4y2BapgydF4y2Ba< 0.01;***,gydF4y2BapgydF4y2Ba< 0.001。gydF4y2Ba
讨论gydF4y2Ba
SB706504显著抑制COPD MDMs中一系列炎症基因的转录。在AMs和MDMs中联合使用SB706504和地塞米松可最大限度地抑制细胞因子的产生,强调这可能是COPD患者有效的抗炎策略。然而,在COPD MDMs中,27个被LPS激活的炎症基因对地塞米松的作用不敏感。此外,其中23个基因对地塞米松和SB706504不敏感,提示COPD巨噬细胞中存在“耐药”炎症基因亚群。gydF4y2Ba
单用SB706504(-2.04)和单用地塞米松(-3.39)对基因表达的褶皱变化抑制的数值平均值小于两药合用时的褶皱变化抑制数值平均值(-8.90)。这提示了一种协同相互作用,例如GC上调丝裂原活化的蛋白激酶磷酸酶(gydF4y2BaLasa等人,2002gydF4y2Ba)或p38 MAPK抑制GR (gydF4y2BaIrusen et al., 2002gydF4y2Ba;gydF4y2BaSzatmáry et al., 2004gydF4y2Ba).需要使用COPD巨噬细胞进行进一步的研究来解决这些潜在的机制。gydF4y2Ba
我们通过微阵列分析鉴定出4个抗gc基因,它们位于4号染色体上的相同位点(q12-q13);CXCL1 (GROα)、CXCL2 (GROβ)、CXCL3 (GROγ)和IL-8。此外,对于其中的三个基因,使用SB706504时也没有明显的抑制作用,但两种药物同时使用时也有明显的抑制作用。这表明这些药物之间存在协同作用或通过抑制p38 MAPK“解锁”GC不敏感。这些基因在单个染色体上的接近性提高了该位点染色质重塑的可能性。Saccani等人(gydF4y2Ba2002gydF4y2Ba)先前的研究表明,LPS刺激树突状细胞导致IL-6、IL-8、12p40和单核细胞趋化蛋白(CCL2)启动子区域的p38依赖性组蛋白3磷酸化,尽管TNFα和巨噬细胞炎症蛋白(CCL3)启动子区域没有磷酸化。由于染色质的解绕,这种磷酸化导致转录潜能增加。同样,在目前的研究中,SB706504可能抑制lps诱导的染色质结构变化,这可能增加了GC效应。在COPD肺泡巨噬细胞中,氧化应激引起的组蛋白去乙酰化酶活性降低被认为导致染色质重塑与GC敏感性降低相关(gydF4y2BaCosio et al., 2004gydF4y2Ba).我们的研究涉及TLR刺激而不是氧化应激,但表明位于4号染色体的四种趋化因子降低了GC敏感性,并且在COPD巨噬细胞中研究该基因的染色质结构是有必要的。gydF4y2Ba
先前COPD am的数据表明IL-8具有gc抗性(gydF4y2BaCulpitt et al., 2003gydF4y2Ba;gydF4y2BaCosio et al., 2004gydF4y2Ba).这些先前的研究进一步支持了我们的发现,即巨噬细胞IL-8的产生对GC不敏感。这可能具有相当大的治疗意义,因为IL-8似乎在COPD病理生理中起关键作用;在慢性阻塞性肺病患者的肺中,这种中性粒细胞趋化剂的水平升高(gydF4y2Ba基廷等人,1996gydF4y2Ba),并与疾病进展速度有关(gydF4y2BaWilkinson et al., 2003gydF4y2Ba).同样,先前的数据表明,与其他细胞因子相比,巨噬细胞产生IL-8对p38抑制剂的影响不太敏感(gydF4y2BaSmith et al., 2006gydF4y2Ba;gydF4y2BaTudhope et al., 2008gydF4y2Ba).我们也观察到了这种现象。gydF4y2Ba

ELISA检测地塞米松、SB706504和共治疗对lps刺激型COPD患者TNFα蛋白抑制作用的比较(清条、gydF4y2BangydF4y2Ba= 9)和HS(灰色条,gydF4y2BangydF4y2Ba= 10) (A)和up - lps刺激的COPD(清条,gydF4y2BangydF4y2Ba= 7)和HS(灰色条,gydF4y2BangydF4y2Ba= 7) AMs (B)。数据以IQR和极差的中位数表示。*,gydF4y2BapgydF4y2Ba< 0.05;**gydF4y2BapgydF4y2Ba< 0.01;***,gydF4y2BapgydF4y2Ba< 0.001。gydF4y2Ba
趋化因子CXCL1、2和3分别构成GRO家族的三个成员,GROα、GROβ和GROγ。CXCL1在肺组织中的表达升高(gydF4y2BaTomaki等人,2007gydF4y2Ba)及诱导痰(gydF4y2Ba特拉维斯等人,2002gydF4y2Ba),与对照组相比。GRO家族成员引起中性粒细胞趋化;因此,我们发现巨噬细胞衍生的GRO产生对GC不敏感,这可能与疾病病理生理学有关。gydF4y2Ba
我们确定了23个炎症基因,这些基因在MDMs中被LPS上调,但对所使用的任何治疗都不敏感。其中包括细胞因子IL-1β、IL-15和IL-18以及趋化因子CCL1、3、4、5、18、20和23以及CXCL5、11和13。IL-1β和IL-18序列同源,是IL-1家族的成员。IL-1β基因在COPD肺组织中的表达升高(gydF4y2BaTomaki等人,2007gydF4y2Ba),而也有证据表明IL-18在COPD中过表达(gydF4y2BaImaoka et al., 2008gydF4y2Ba).其他与COPD发病有关的gc不敏感的炎症基因包括CXCL11 (gydF4y2BaCosta et al., 2008gydF4y2Ba), ccl5 (gydF4y2BaCosta et al., 2008gydF4y2Ba), ccl3 (gydF4y2BaDi Stefano et al., 1998gydF4y2Ba)及CCL20 (gydF4y2BaDemedts et al., 2007gydF4y2Ba)在慢性阻塞性肺病患者的气道中。gydF4y2Ba
我们决定通过qPCR评估IL-1β、IL-6、GM-CSF、TNFα和IL-8 mRNA水平。使用的标准gydF4y2BapgydF4y2BaqPCR分析< 0.05,我们发现这些基因均被两种药物显著抑制,但IL-1β和IL-8是最不敏感的基因。这证实了IL-1β和IL-8都具有gc抗性的阵列分析。值得注意的是,我们使用的标准是gydF4y2BapgydF4y2Ba< 0.01和>2的倍数变化来定义显著变化,这是阵列研究中的常见做法。gydF4y2Ba
IPA分析用于生成gc不敏感基因之间可能关系的网络,同样的方法用于p38 MAPK抑制剂不敏感基因。这是利用已知的蛋白质和复合物之间的机制关系来创建假设基因网络的公认方法(gydF4y2BaJuric et al., 2007gydF4y2Ba;gydF4y2BaLi et al., 2007gydF4y2Ba;gydF4y2BaSavli et al., 2008gydF4y2Ba).IPA网络允许“高度连接”的基因被识别,因此,更有可能参与炎症过程。这种方法是“假设生成”,我们观察到IL-1β, IL-8, IL-18和CCL5在GC抗性中起主要作用。因为所有这些炎症介质的水平在COPD患者中都升高了(gydF4y2Ba基廷等人,1996gydF4y2Ba;gydF4y2BaTomaki等人,2007gydF4y2Ba;gydF4y2BaCosta et al., 2008gydF4y2Ba;gydF4y2BaImaoka et al., 2008gydF4y2Ba),那么这些细胞因子和趋化因子可能被认为是COPD患者gc抵抗性炎症的关键罪魁祸首。gydF4y2Ba
IL-1β、IL-8和IL-18也参与了对SB706504的抗性。这些基因可能受相同的转录因子调控,可能是两种IPA网络提示的NFκBas。需要强调的是,这些网络并不能证明这一机制,但提供了IL-1β、IL-8和IL-18受NFκB信号调控的假设,而NFκB信号在COPD巨噬细胞中对GC和p38 MAPK抑制剂均具有耐药性。gydF4y2Ba
我们的基因阵列研究使用MDMs作为AMs的替代品。在研究p38 MAPK抑制剂的药理作用方面,MDMs和AMs有相似之处(gydF4y2BaTudhope et al., 2008gydF4y2Ba).因此,我们假设我们在MDMs中观察到的GC和p38 MAPK抑制剂抵抗炎症基因也可能对这些药物在AMs中的作用产生抗性。我们使用了一组重度COPD患者进行基因阵列研究。研究同样的结果是否适用于轻度COPD患者和对照组,研究我们的任何发现是否依赖于COPD的存在或严重程度,当然是有意义的。gydF4y2Ba
p38 MAPK抑制剂可能在转录、转录后、翻译或翻译后水平起作用(gydF4y2BaWang等,1999gydF4y2Ba;gydF4y2BaWinzen et al., 1999gydF4y2Ba;gydF4y2BaBrook et al., 2006gydF4y2Ba).在COPD患者的MDMs中,基因阵列和qPCR显示SB706504抑制一系列炎症基因的转录。在抑制转录物丰度方面,SB706504不如地塞米松有效,这在qPCR数据中尤为明显。然而,SB706504和单独地塞米松对蛋白水平的影响大致相似。这表明SB706504具有显著的转录后效应,影响分泌炎性蛋白的水平。同样,Birrell等人(gydF4y2Ba2006gydF4y2Ba)报道SB239063和2,1(1,3-二羟基丙基-2-基)-4-(4-氟苯基)-5-(2-苯氧嘧啶-4-基)咪唑对AM IL-6产生的抑制作用主要是在蛋白质产生水平而不是mRNA表达水平(gydF4y2BaBirrell et al., 2006gydF4y2Ba).p38 MAPK抑制剂对转录的影响可能具有浓度依赖性,值得进一步探讨。gydF4y2Ba
在COPD患者和吸烟者中,SB706504显著抑制lps诱导的TNFα产生,尽管吸烟者的抑制作用略高于COPD患者(76.1和67.4%)。gydF4y2BapgydF4y2Ba= 0.047)。SB706504也被发现在TLR4特异性激动剂UP-LPS刺激下显著抑制TNFα的产生,与吸烟者相比,在COPD患者中具有相似的抑制作用。所使用的样本量在统计学上没有明确的支持,无法对COPD患者和吸烟者之间的药物效应进行敏感的比较。已知COPD am对GC的敏感性低于对照组(gydF4y2BaCulpitt et al., 2003gydF4y2Ba);也许一个更大的研究可以阐明当使用p38 MAPK抑制剂时的任何这种差异。gydF4y2Ba
综上所述,我们已经证明新型p38 MAPK抑制剂SB706504对COPD巨噬细胞产生炎症介质具有显著的抑制作用。重要的是,我们提供的证据表明,将这种p38 MAPK抑制剂与GC一起使用可以增强抗炎作用。然而,我们也证明了巨噬细胞炎症基因的一个亚群对这些抗炎药物的作用具有抗性,其中一些已知与COPD的病理生理有关。可能需要针对这些基因采取不同的抗炎干预措施。gydF4y2Ba
致谢gydF4y2Ba
Laurie Scott、Alistair Maclaren、Siamah Kidwai、Paul Murdoch、Ted Cook、Vicky Bousgouni、Susan Boyce和Tom Southworth参与了这项研究的运行和提供建议。gydF4y2Ba
脚注gydF4y2Ba
本研究得到了英国生物技术与生物科学研究委员会和葛兰素史克公司的支持[Grant BBS/S/N/2004/11516]。gydF4y2Ba
文章,出版日期和引用信息可在gydF4y2Bahttp://jpet.aspetjournals.orggydF4y2Ba.gydF4y2Ba
doi: 10.1124 / jpet.108.142950。gydF4y2Ba
缩写:gydF4y2Ba慢性阻塞性肺疾病;AM,肺泡巨噬细胞;GC,糖皮质激素;GR:糖皮质激素受体;NF:核因子;toll样受体;有限合伙人,脂多糖;MAPK,丝裂原活化蛋白激酶;肿瘤坏死因子;IL,白介素; GM-CSF, granulocyte macrophage colony-stimulating factor; MDM, monocyte-derived macrophage; PCR, polymerase chain reaction; UP, ultrapure; DMSO, dimethyl sulfoxide; qPCR, quantitative PCR; ELISA, enzyme-linked immunosorbent assay; IPA, ingenuity pathways analysis; FEV, forced expiratory volume; FVC, forced vital capacity; CCL, chemokine (CC motif) ligand; SB706504,NgydF4y2Ba含氰基的- - - - - -gydF4y2BaNgydF4y2Ba”——(2 - {[8 - (2,6-difluorophenyl) 4 (4-fluoro-2-methylphenyl) 7-oxo-7 8-dihydropyrido [2, 3 -gydF4y2BadgydF4y2Ba] pyrimidin-2-yl]氨基乙基)胍;}SB239063,gydF4y2Ba反式gydF4y2Ba(1) - 4-hydroxycyclohexyl 4 - (4-fluorophenyl) 5 - (2-methoxypyridimidin-4-yl)咪唑;SD-282,吲哚-5-羧酰胺(atp竞争性p38激酶抑制剂)。gydF4y2Ba
↵gydF4y2Ba
 本文的在线版本(可在gydF4y2Bahttp://jpet.aspetjournals.orggydF4y2Ba)包含补充材料。gydF4y2Ba
本文的在线版本(可在gydF4y2Bahttp://jpet.aspetjournals.orggydF4y2Ba)包含补充材料。gydF4y2Ba-
- 收到了gydF4y2Ba2008年7月9日。gydF4y2Ba
- 接受gydF4y2Ba2008年10月21日。gydF4y2Ba

- 美国药理学和实验治疗学会gydF4y2Ba
参考文献gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
本期gydF4y2Ba

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
地塞米松和p38丝裂原活化蛋白激酶抑制剂抑制脂多糖刺激的慢性阻塞性肺疾病巨噬细胞炎症基因表达gydF4y2BaNgydF4y2Ba含氰基的- - - - - -gydF4y2BaNgydF4y2Ba”——(2 - {[8 - (2,6-difluorophenyl) 4 (4-fluoro-2-methylphenyl) 7-oxo-7 8-dihydropyrido [2, 3 -gydF4y2BadgydF4y2Ba[嘧啶-2-基]氨基}乙基)胍(SB706504)gydF4y2Ba







