





摘要gydF4y2Ba
呼吸道合胞病毒(RSV)感染婴儿气道中大量中性粒细胞被白细胞介素-8等趋化因子招募,特异性炎症分子可延缓细胞凋亡,增加其寿命。本研究的目的是研究呼吸道合胞病毒毛细支气管炎气道分泌物中是否含有影响中性粒细胞凋亡的因素。gydF4y2Ba
24例RSV细支气管炎患儿(31例婴儿对照和12例成人)的鼻腔灌洗液(NLF)。从健康成年志愿者中分离的中性粒细胞与NLF在Dulbecco修饰Eagle培养基(DMEM)中孵育24 h,并用Hoechst 33342和碘化丙啶活力染料测定细胞凋亡和坏死。使用白三烯- b抑制剂研究延迟中性粒细胞凋亡的假定因素的存在gydF4y2Ba4gydF4y2Ba,脂多糖和IL-8受体CXCR2,以及粒细胞单核细胞集落刺激因子阻断抗体。NLF的特征包括热不稳定性、蛋白质水解、脱氧核糖核酸酶消化和分子过滤试验。gydF4y2Ba
RSV细支气管炎患儿和对照组的NLF明显延迟中性粒细胞凋亡,而健康成人的NLF则没有。这些抑制剂分子都不能阻止这种凋亡延迟,但活性是易热的和> 3kda。gydF4y2Ba
研究表明,婴儿鼻腔灌洗液可明显延缓中性粒细胞凋亡。推测是婴儿呼吸道中性粒细胞存活时间的延长导致了患有呼吸道感染的婴儿呼吸道中性粒细胞的特征性积累。gydF4y2Ba
本研究部分由谢菲尔德医院儿童基金和行动实验室慈善机构提供资金。gydF4y2Ba
呼吸合胞病毒(RSV)负责影响整个人口的呼吸系统疾病的年度流行病gydF4y2Ba1gydF4y2Ba- - - - - -gydF4y2Ba3.gydF4y2Ba尤其是婴儿,他们中的大多数在第一个冬天就被感染了gydF4y2Ba1gydF4y2Ba,gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba.其中,20-30%的发展较低呼吸道症状,0.5-2%患者与RSV支气管炎住院gydF4y2Ba1gydF4y2Ba,gydF4y2Ba2gydF4y2Ba,gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba.越来越多的人认识到RSV是老年人呼吸道疾病的主要原因gydF4y2Ba6gydF4y2Ba,gydF4y2Ba7gydF4y2Ba.一些前瞻性研究也表明住院患者下呼吸道RSV感染gydF4y2Ba8gydF4y2Ba和非生长gydF4y2Ba9gydF4y2Ba婴儿在随后几年中导致过量的较高呼吸道症状。这些症状在生命的第一个十年后逐渐下降,但这种发病率过度的原因仍然不明确gydF4y2Ba10gydF4y2Ba.gydF4y2Ba
关于宿主/病毒相互作用的性质有很多猜测gydF4y2Ba3.gydF4y2Ba,gydF4y2Ba11gydF4y2Ba30多个 多年前,有人提出RSV毛细支气管炎是一种特异性免疫病理学的结果。尽管有大量研究证明了这一点,但尚未发现明显的免疫病理过程。目前的作者已经证明,在婴儿呼吸道合胞病毒感染中,中性粒细胞是上下气道中的主要炎性细胞,在呼吸道合胞病毒感染中占主导地位∼80%的细胞从纤维束中恢复gydF4y2Ba12gydF4y2Ba.这些发现随后被其他人复制gydF4y2Ba13gydF4y2Ba.来自这些婴儿的气道分泌物中的蛋白酶活性增加,中性粒细胞弹性蛋白酶(Ne)核算这些蛋白酶的大部分活动gydF4y2Ba14gydF4y2Ba灌洗液样本中存在降解的纤维连接蛋白gydF4y2Ba15gydF4y2BaNE可能通过直接损伤未成熟气道、诱导气道高反应性以及诱导腺体和杯状细胞增生而在复发性症状的发生中发挥作用gydF4y2Ba16gydF4y2Ba.中性粒细胞在病毒间隙中的作用,如果有的话,尚不清楚。gydF4y2Ba
中性粒细胞在气道内的积聚可受其迁移率和寿命的影响。在这种情况下,白细胞介素(IL)-8是中性粒细胞的主要趋化因子,RSV感染刺激各种细胞释放IL-8gydF4y2Ba体外gydF4y2Ba,包括上皮细胞,巨噬细胞和中性粒细胞gydF4y2Ba17gydF4y2Ba,gydF4y2Ba18gydF4y2Ba.目前的作者报道了急性呼吸道合胞病毒毛细支气管炎婴儿鼻腔灌洗液(NLF)中IL-8的升高gydF4y2Ba19gydF4y2Ba.疾病严重程度与IL-8浓度之间也可能存在相关性gydF4y2Ba20.gydF4y2Ba虽然目前尚不清楚IL-8水平是否反映或有助于疾病严重程度。中性粒细胞寿命通过细胞凋亡来调节gydF4y2Ba21gydF4y2Ba.凋亡的中性粒细胞可被驻留的巨噬细胞吞噬gydF4y2Ba通过gydF4y2BaVITRONECTIN或磷脂酰丝氨酸受体gydF4y2Ba22gydF4y2Ba因此,将有害的中性粒细胞产品的释放限制在气道中,如果中性粒细胞因坏死而死亡。有可能存在于婴儿的气道分泌患有RSV支气管炎的一些因素,例如IL-8,白酮-BgydF4y2Ba4gydF4y2Ba(LTBgydF4y2Ba4gydF4y2Ba)粒细胞-单核细胞集落刺激因子(GM-CSF)抑制中性粒细胞凋亡gydF4y2Ba21gydF4y2Ba,gydF4y2Ba23gydF4y2Ba,延长细胞存活时间。以前一组研究呼吸道合胞病毒毛细支气管炎患儿呼吸道中性粒细胞恢复的研究表明,在这种情况下中性粒细胞凋亡可能加速gydF4y2Ba24gydF4y2Ba.然而,本研究评估了已经驻留在气道中的中性粒细胞凋亡的速率未知持续时间,并将其与循环中性粒细胞的存活率进行了比较。不知道这些中性粒细胞居住多长时间居住在气道中,很难确定它们的存活是否已缩短或延长。gydF4y2Ba
本研究的目的是确定RSV细支气管炎婴儿气道分泌物中释放的因素是否影响中性粒细胞凋亡率。另外还进行了一些研究,试图确定可能导致这些影响的因素。gydF4y2Ba
材料和方法gydF4y2Ba
学科gydF4y2Ba
NLF来自于患有RSV毛细支气管炎(RSV-NLF)住院的婴儿,他们在被招募之前通过鼻咽抽吸物的间接免疫荧光证实RSV呈阳性。NLF也来自于健康婴儿和成人。健康婴儿在常规的“健康婴儿诊所”被招募在采样时没有呼吸道或其他临床疾病史。2-3天后联系父母,以确保这些婴儿在采样时没有感染呼吸道疾病。成年人是没有呼吸道问题史的健康志愿者。gydF4y2Ba
伦理批准(南谢菲尔德伦理委员会,谢菲尔德,英国)和知情同意获得了婴儿的父母和成年志愿者。gydF4y2Ba
鼻灌洗液的收集和加工gydF4y2Ba
根据诺亚所述收集NLFgydF4y2Ba等gydF4y2Ba25gydF4y2Ba,通过介绍5 将1毫升无菌等渗盐水注入一个鼻孔,同时将婴儿头部倾斜至另一侧。生理盐水排入对面的鼻孔,并收集在一个容器中。然后在对侧鼻孔重复该过程。十 将mL NLF汇集在一起,在875×10℃下离心gydF4y2BaggydF4y2Ba五 在4°C下最小,并在室温下等分储存−70°C。婴儿和成人的每个NLF样本的蛋白质浓度均使用敏感的双金鸡纳酸蛋白质测定法(BCA测定法;美国伊利诺伊州皮尔斯市)。gydF4y2Ba
中性粒细胞分离gydF4y2Ba
中嗜中性粒细胞与史密斯描述的健康成人志愿者的周围全血中分离出来gydF4y2Ba等gydF4y2Ba26gydF4y2Ba,分层在组织盖-1077(Sigma Chemical Company Ltd,Poole,UK)和离心。收集粒状细胞并在无内毒素的磷酸盐缓冲盐水(Biowhittaker,MD,USA)中洗涤。然后在氯化铵溶液中裂解红细胞,中性粒细胞以160×离心gydF4y2BaggydF4y2Ba8分钟颗粒。gydF4y2Ba
中性粒细胞制剂的活力常规> 95%,纯度,通过自动止血仪分析评估,均为95%,总收益率不同于0.75-1.5×10之间gydF4y2Ba6gydF4y2Ba·毫升gydF4y2Ba−1gydF4y2Ba为这些捐献者提供血液gydF4y2Ba
中性粒细胞与鼻腔灌洗液的孵育gydF4y2Ba
纯化的中性粒细胞在1×10重悬gydF4y2Ba6gydF4y2Ba·毫升gydF4y2Ba−1gydF4y2Ba在Dulbecco改性Eagle培养基(DMEM;Sigma Chemical Company Ltd),含2 mM谷氨酰胺,100 g·mLgydF4y2Ba−1gydF4y2Ba青霉素50µg·mLgydF4y2Ba−1gydF4y2Ba链霉素(称为DMEM-AP)。将其中一些中性粒细胞重悬于20%的自体等离子体中(在离心时除去)。还将抗生素添加到NLF样品中。gydF4y2Ba
作者播种了1×10gydF4y2Ba5gydF4y2Ba在100μl等分试样(康宁)组织培养皿(康宁Costar Ltd,High-Wycombe,UK)中的井中的嗜中性粒细胞以三份适用于每个样品/处理,加入另外100μL整齐,稀释或处理的NLF并轻轻混合内容。在每个实验中,在200μlDMEM-AP中测量中性粒细胞的活力。将细胞在37℃下在37℃下在孵化器(Sanyo,Osaka,Japan)的潮湿的95%空气中,5%二氧化碳孵育24小时。gydF4y2Ba
中性粒细胞活力评估gydF4y2Ba
在37℃温育24-H后,中性粒细胞用8μmHoechst33342(Sigma Chemical Company Ltd)和5μm碘化物(Molecular Probides,Cambridge Biochemicalics,Cambridge,UK)在37℃下染色15分钟。然后使用荧光倒的徕卡DMIRB显微镜(Leica,Wetzlar,Germany)检查它们而无需进一步操纵。The morphology and characteristics of the viable and apoptotic cells (Hoechst stained) and necrotic cells (propidium iodide stained) are shown in figures 1a and b⇓gydF4y2Ba.这两种荧光染料在紫外线过滤下都是可见的,可以同时计数。每一种治疗方法中,三个井中每个井的五个独立随机网格区域使用Whipple网格进行计数,获得活细胞、凋亡细胞和坏死细胞的发生率。gydF4y2Ba
鼻腔灌洗液中细胞凋亡抑制活性的生化特征gydF4y2Ba
在一些实验中,NLF在加入中性粒细胞之前被处理过。来自RSV组的具有类似高活性的NLF样本也被汇集在一起,以帮助进行生化特征分析。gydF4y2Ba
为了确定影响细胞凋亡的活性是否不稳定,RSV-NLF(n=5)在液氮中冷冻并在五个重复循环中解冻。RSV-NLF(n=5)也被加热至90°C 15分钟 min,冷却,然后用于活性测定。gydF4y2Ba
细菌脱氧核糖核酸(DNA)具有潜在的促炎特性,但据报道,降解宿主DNA具有免疫抑制作用作用gydF4y2Ba27gydF4y2Ba. 为了确定受损呼吸道细胞释放的DNA是否影响中性粒细胞凋亡,将合并的RSV-NLF(n=5)消化2小时 h在37°C下,温度为500 微克·毫升gydF4y2Ba−1gydF4y2Ba脱氧核糖核酸酶(DNAse)I型(西格玛化学有限公司)。在类似条件下使用小牛精子DNA证实了DNA酶的降解活性,并证明在用溴化乙锭染色的1%琼脂糖凝胶上分离后涂片。gydF4y2Ba
RSV-NLF(n=5)在0.5%牛胰蛋白酶(西格玛化学有限公司)中进行有限消化2小时 h,以研究蛋白质在NLF中的作用。胰蛋白酶活性在100℃时停止 微克·毫升gydF4y2Ba−1gydF4y2Ba大豆胰蛋白酶抑制剂(Sigma Chemical Company Ltd)。通过将消化的NLF在10%十二烷基硫酸钠 - 聚丙烯酰胺凝胶电泳(SDS-PAGE)染色用银染色的10%钠的NLF来证实有限的蛋白水溶性。对照包括添加胰蛋白酶和大豆胰蛋白酶抑制剂复合物,但没有NLF。gydF4y2Ba
为了确定NLF是否含有低分子量活性,0.5 对RSV-NLF的mL样品进行离心过滤,使用3 K分子量Centricon截止过滤器(美国马萨诸塞州贝德福德市密理博公司Amicon)。精确测量滤液和滞留物的体积,两者的体积都达到500 µL,使用无菌盐水溶液。五百 µL未过滤NLF也使用无菌生理盐水以等量稀释。滤液、滞留液和未过滤的NLF随后与中性粒细胞一起培养。gydF4y2Ba
LTB的表征gydF4y2Ba4gydF4y2BaNLF样本中的IL-8、GM-CSF和LPS活性gydF4y2Ba
据报道,这些因素延迟了实验模型中的中性粒细胞凋亡gydF4y2Ba21gydF4y2Ba,gydF4y2Ba23gydF4y2Ba并考察了他们对NLF活动的贡献。gydF4y2Ba
白三烯的LTBgydF4y2Ba4gydF4y2Ba抑制剂VML 295 (LY29311;来自英国萨里Vanguard Medica的G. Parker的礼物)溶解在二甲基亚砜(DMSO)中。将VML 295(10µM和100µM)溶解在汇集的和单个NLF样品中,然后加入中性粒细胞。gydF4y2Ba
SB-237844(来自美国普鲁士国王史密斯克林·比查姆S.Sarau的礼物),一种CXCR2 IL-8受体的竞争性拮抗剂,溶解在二甲基亚砜中,并从10-0.05添加到中性粒细胞中 µM为5 添加混合样本和单个样本NLF前的最小值。gydF4y2Ba
在1-100μg·ml下加入脂多糖(LPS)抑制剂多粘剂-BgydF4y2Ba−1gydF4y2Ba在添加合并或单独的NLF样品之前,在中性粒细胞5分钟。通过抑制10μg·mL的抑制来证实了多粘土-B的效果gydF4y2Ba−1gydF4y2Ba埃斯克里希亚洲gydF4y2Ba科利gydF4y2BaLPS(血清型0.26:B6;西格玛化学有限公司),被证明能显著延迟中性粒细胞凋亡率。gydF4y2Ba
阻断细胞因子GM-CSF 10µg·mLgydF4y2Ba−1gydF4y2Ba, 50µg·毫升gydF4y2Ba−1gydF4y2Ba和100µg·毫升gydF4y2Ba−1gydF4y2Ba将阻断的山羊免疫球蛋白(Ig)G抗人GM-CSF(英国剑桥剑桥生化研发系统公司)与混合或单个NLF孵育30天 在37°C下至少稀释一分钟。然后将NLF抗体混合物添加到中性粒细胞中。这些抑制实验的对照包括用于某些化合物的溶剂的等效稀释液和GM-CSF阻断实验中类似稀释液的非免疫山羊IgG。gydF4y2Ba
数据统计分析gydF4y2Ba
组间和组内差异分别采用非参数Mann-Whitney u检验和Kruskal-Wallis非参数检验。用Spearman相关系数检测鼻腔灌洗液中细胞凋亡水平与蛋白含量的关系。差异有显著性(p<0.05)。数据在文本中显示为均值±95%置信限。gydF4y2Ba
后果gydF4y2Ba
学科gydF4y2Ba
招募24名因RSV细支气管炎入院的婴儿,中位年龄为120天(范围33-242),样本在症状出现后的中位3天(范围1-4)获得。对照组样本来自31名婴儿(中位年龄149天(19-433天))和12名成人(中位年龄38岁(21-48天)),没有呼吸道感染的症状证据。33%患有毛细支气管炎的婴儿和27%的对照组婴儿被动地暴露在烟雾中。这些成年人中没有一个是吸烟者。gydF4y2Ba
鼻灌洗液蛋白含量gydF4y2Ba
来自RSV支气管炎的婴儿的NLF样品的中值蛋白质浓度为55.9μg·mLgydF4y2Ba−1gydF4y2Ba(范围7.6-386.5)。婴儿对照的价值为62.3μg·mLgydF4y2Ba−1gydF4y2Ba(19.9–212.0范围),成人对照组的值为19.5 微克·毫升gydF4y2Ba−1gydF4y2Ba(3.1-53.1范围),它们显着降低(P <0.001),而不是两个婴儿组。gydF4y2Ba
鼻灌淫对中性粒细胞生存的影响gydF4y2Ba
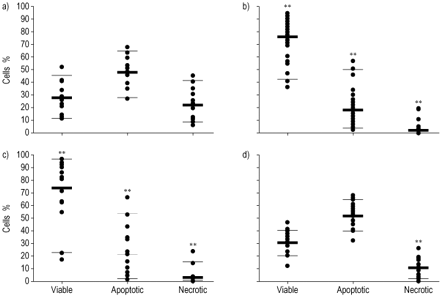
中性粒细胞在培养基中或用RSV-NLF孵育,对照婴儿的NLF和健康成人的NLF,测定24小时后,测定细胞活力后。单独的培养基中活细胞的平均量为27.5%(±3.3),其中49.3%的细胞是凋亡(±3.1)和23.4%是坏死(±3.5),如图2a所示gydF4y2Ba⇓gydF4y2Ba.在添加RSV-NLF后,与单独单独的培养基相比,较为显着的增加至77.5%(±3.3)活细胞,减少至19.5%凋亡(±2.9)和3.0%坏死(±0.9)(图。 2b⇓gydF4y2Ba).这些变化也可以在婴儿对照组NLF中培养的中性粒细胞中看到,与单独培养基相比,其中76.3%(±5.3)细胞存活,20.4%(±3.3)细胞凋亡,3.3%(±1.4)细胞坏死(图2c)gydF4y2Ba⇓gydF4y2Ba).然而,当从成年人中孵育NLF时,该模式与培养基中的细胞相当,细胞可行33.2%(±2.1),54.5%(±2.2)凋亡,小但显着降低至12.1%(±1.6))坏死的细胞(图2DgydF4y2Ba⇓gydF4y2Ba).gydF4y2Ba
凋亡抑制活性的特征gydF4y2Ba
热稳定性gydF4y2Ba
当RSV-NLF样品在90°C下加热5分钟时,观察到抗凋亡活性的显著损失。在本实验中(n=5), RSV-NLF的活细胞数量为85.5%(±10%),加热后显著(p<0.05)为37.0%(±27)。凋亡细胞由24.2%(±16)增加到53.5%(±20),p<0.05;坏死细胞由0.8%(±0.8)增加到9.8%(±9.0),差异无统计学意义。这些水平的生存能力热处理后,可与那些看到的培养基单独。gydF4y2Ba
重复冻融样本不会改变RSV-NLF延迟中性粒细胞凋亡的能力。gydF4y2Ba
DNA酶消化gydF4y2Ba
的治疗汇集(N = 5)RSV-NLF样品用DNAse没有减少抗凋亡作用相比未处理RSV-NLF(91.2%(±7.0)gydF4y2Ba对gydF4y2Ba89.2%(±8.7)活细胞)。gydF4y2Ba
DiafiltrationgydF4y2Ba
将合并的RSV-NLF(n=5)用滤纸进行透析过滤∼3. kDa切断孔膜以确定NLF中是否存在影响细胞凋亡率的低分子量成分,如离子。来自RSV-NLF的滤液没有增加中性粒细胞的活力(6.3%(±6.0)),表明凋亡抑制活性>3 kDa。与单独培养基(17%(±3.6))相比,过滤器上方的RSV-NLF支持中性粒细胞活力的增加(38.8%(±24.1)),但其活性低于未过滤NLF(活力91%(±8.0))。gydF4y2Ba
阻塞LTB的影响gydF4y2Ba4gydF4y2Ba、GM-CSF、LPS和IL-8gydF4y2Ba
在所有这些实验中,通过将这些试剂添加到单独培养基中培养的中性粒细胞中,首先确定了与有效活性一致但毒性最小的浓度范围。gydF4y2Ba
LTB.gydF4y2Ba4gydF4y2Ba抑制剂gydF4y2Ba
LTB对中性粒细胞活力无明显影响gydF4y2Ba4gydF4y2Ba加入抑制剂以合并(n = 8)RSV-NLF,10μm(79.2%(±8.3))或100μm(85.4%(±13.5)),与单独的样品的效果相比(74.3%(±16.5)))。gydF4y2Ba
gm - csf抑制剂gydF4y2Ba
100%时,中性粒细胞的存活率(81.0%(±14.2))无显著增加 微克·毫升gydF4y2Ba−1gydF4y2Ba与单独的RSV-NLF相比,将对GM-CSF的阻断抗体加入到RSV-NLF(n = 8)中(72%(±22.1))。使用其他浓度的抗体没有观察到显着差异。在等效浓度下的非免疫山羊IgG也没有影响RSV-NLF中的活力水平。gydF4y2Ba
多粘菌素gydF4y2Ba
有限合伙人(10µg·毫升gydF4y2Ba−1gydF4y2Ba)显着(P <0.01)与仅在培养基中保持的细胞(33.6(±14.2))增加,增加了中性粒细胞的活力(82.6(±6.6))。金蛋白(10μg·mlgydF4y2Ba−1gydF4y2Ba)降低(p<0.01)中性粒细胞活力(66.7%(±9.4))(n=5),相比于单独RSV-NLF(80.0±(8.6)),但这一变化不显著。在此浓度下,与单独培养的中性粒细胞(42.5(±16.8))相比,多粘菌素添加到单独培养的中性粒细胞中无毒性作用(活力为53%(±19.3))。gydF4y2Ba
CXCR2 (IL-8)受体抑制剂gydF4y2Ba
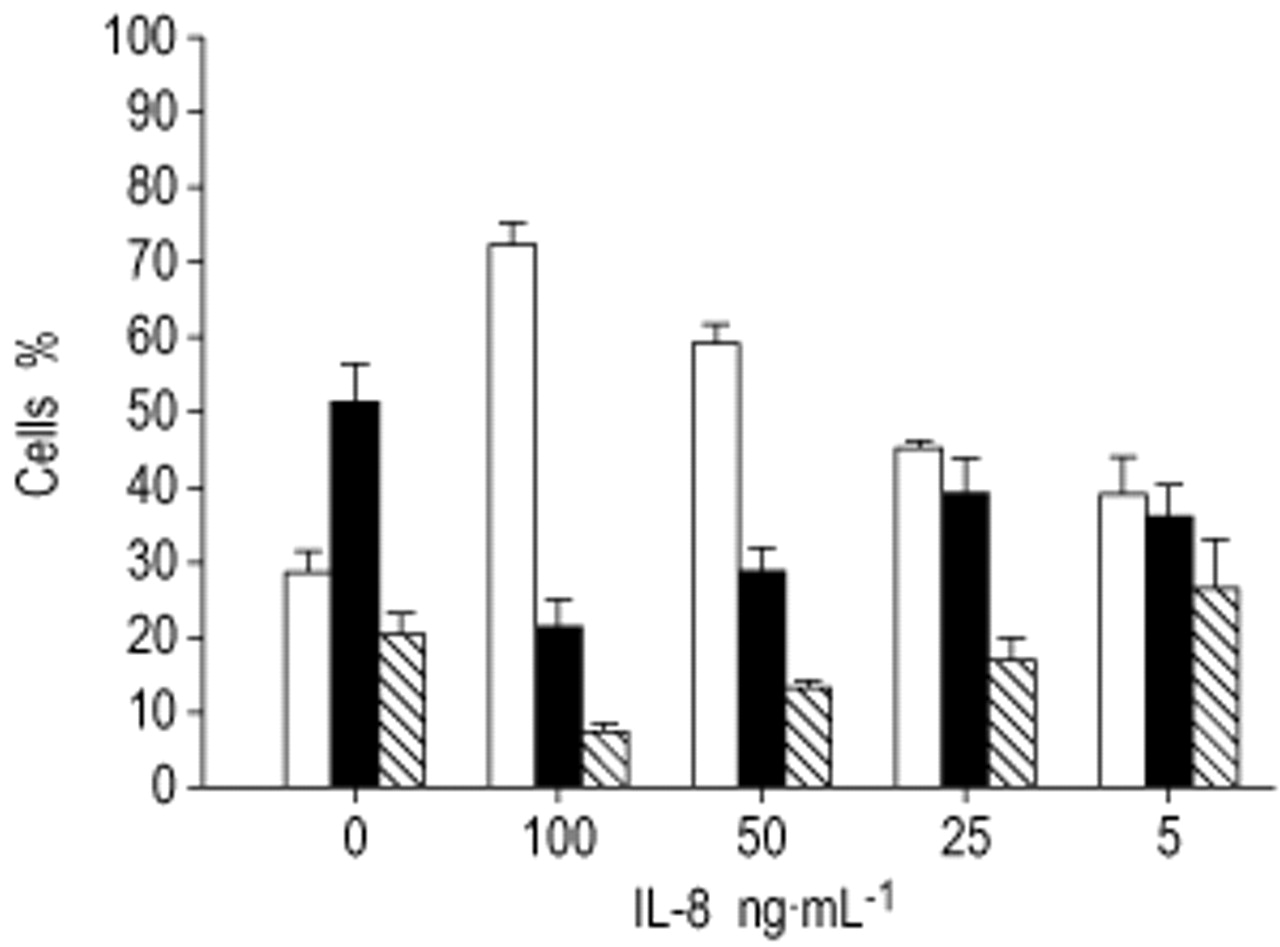
IL-8加入来自5-100ng·ml的中性粒细胞gydF4y2Ba−1gydF4y2Ba随着细胞因子浓度的增加,支持存活率的剂量依赖性增加和细胞死亡的减少(图。 3.gydF4y2Ba⇓gydF4y2Ba).在低(5 ng·ml的情况下gydF4y2Ba−1gydF4y2Ba)或高浓度(100 ng·mLgydF4y2Ba−1gydF4y2Ba), CXCR2抑制剂(从0.05-10 μ M)显示IL-8的作用没有逆转。CXCR2抑制剂对RSV-NLF的活性也没有影响,gydF4y2Ba例如gydF4y2Ba100 µM抑制剂的活性为82%(±8.4)gydF4y2Ba对gydF4y2Ba仅为RSV-NLF 72.1%(±24.8)。gydF4y2Ba
讨论gydF4y2Ba
这些结果表明,与未暴露中性粒细胞和暴露于健康成人NLF的中性粒细胞相比,婴儿气道分泌物显著延缓中性粒细胞凋亡。RSV-NLF的观察效果并非出乎意料,因为在气道分泌物中可能存在许多抑制中性粒细胞凋亡的因素。然而,NLF对明显健康的无症状婴儿的等效效果令人惊讶。婴儿NLF的影响被热变性所否定,不被有限的蛋白质水解所减少,也不是由小分子量成分、DNA或LTB引起的gydF4y2Ba4gydF4y2Ba在纯化的IL-8(>25%)存在下 ng·mLgydF4y2Ba−1gydF4y2Ba)细胞凋亡呈剂量依赖性减少,这与其他报道一致gydF4y2Ba28gydF4y2Ba.这种浓度超过RSV-NLF测量的浓度gydF4y2Ba19gydF4y2Ba但是由于糖酰胺聚糖的烯-8螯合,可以反映细胞表面的局部浓度。然而,CXCR2抑制剂没有阻断IL-8抑制细胞凋亡,并且在NLF中没有影响,尽管它在从CXCR2移位IL-8的浓度范围(50nm-10μm)内进行测试gydF4y2Ba28gydF4y2Ba.这表明il -8介导的凋亡阻断不是通过CXCR2的信号通路介导的,如果这种趋化因子有助于婴儿NLF的凋亡抑制活性,它必须通过CXCR1介导其作用。gydF4y2Ba
而其他研究表明LTB具有抗凋亡作用gydF4y2Ba4gydF4y2Ba、IL-8、GM-CSF和脂多糖gydF4y2Ba21gydF4y2Ba,gydF4y2Ba23gydF4y2Ba,这些药剂已被单独研究gydF4y2Ba体外gydF4y2Ba.这些抑制剂不能显著影响婴儿与NLF孵育的中性粒细胞的延长生存,提示gydF4y2Ba体内gydF4y2Ba这种作用可能是由促炎因子或其他未知因子联合介导的。一个可能影响这些中性粒细胞存活的非特异性因素是NLF的蛋白浓度。虽然成年NLF样本的中位蛋白含量低于两个婴儿组,但在个体水平上存在相当大的重叠,在任何研究组中,都没有观察到蛋白浓度和凋亡水平之间的显著相关性。此外,目前的作者最近开发了一种明确的无血清培养基,以支持中性粒细胞的最佳生存。在这些实验中,我们发现>蛋白(白蛋白)浓度为2 mg·mLgydF4y2Ba−1gydF4y2Ba将生存率提高至10%。婴儿NLF样本的中位蛋白浓度比这个低40倍(约60µg·mL)gydF4y2Ba−1gydF4y2Ba),而婴儿NLF样本将中性粒细胞存活率从~ 27%提高到~ 77%,这表明在婴儿样本中增加的蛋白质不能归因于非特异性影响。gydF4y2Ba
结果表明婴儿患有RSV支气管炎的幼儿系生存期,因此似乎与王的结果相矛盾gydF4y2Ba等gydF4y2Ba24gydF4y2Ba.然而,如果延长寿命最终导致气道内发生细胞凋亡的中性粒细胞比例增加,这两种观察结果可能并不矛盾。王gydF4y2Ba等gydF4y2Ba24gydF4y2Ba提示加速细胞凋亡可以通过限制炎症产物的释放来保护气道。然而,目前的研究可能提出相反的结论,gydF4y2Ba即。gydF4y2Ba嗜中性粒细胞在气道中积聚,解释了婴儿尤其是RSV毛细支气管炎患儿中性粒细胞弹性蛋白酶活性增加的原因gydF4y2Ba14gydF4y2Ba,gydF4y2Ba15gydF4y2Ba,gydF4y2Ba19gydF4y2Ba. 中性粒细胞存活和功能的延长可能有助于RSV感染的症状特征。有趣的是,研究表明,出生后即刻中性粒细胞趋化性相对受损gydF4y2Ba29gydF4y2Ba,可能解释为什么急性支气管炎在生命的第一个月内罕见。gydF4y2Ba
观察到明显健康婴儿的NLF抑制细胞凋亡的程度与RSV-NLF相似,表明婴儿气道中存在与年龄相关的发育变化促进炎症细胞存活。这些细胞的延长存活可以作为一种非特异性防御机制,旨在对抗生命早期经历的频繁呼吸道感染。需要进一步的工作来更详细地探索这种可能性,包括从婴儿和幼儿中获取样本。另一种可能性是,所有健康的对照组婴儿都经历了亚临床感染,尽管可能性很小。随访电话接触证实,当采样时,这些婴儿均未处于明显呼吸系统疾病的临床前阶段。gydF4y2Ba
中性粒细胞性炎症是一种公认的对许多呼吸道病毒的特征性反应gydF4y2Ba30.gydF4y2Ba15-25%的毛细支气管炎是由呼吸道合胞病毒以外的病毒引起的。因此,呼吸道合胞病毒毛细支气管炎患儿呼吸道可能不存在特异性免疫病理,每年冬季婴幼儿毛细支气管炎的高患病率提示对该病毒没有有效的群体免疫。因此,呼吸道合胞病毒每年冬天感染大多数婴儿,其中少数人出现严重的下呼吸道症状,需要住院治疗。gydF4y2Ba
总之,从婴儿(包括患有呼吸道合胞病毒毛细支气管炎的婴儿)中提取的鼻腔灌洗液可显著延缓中性粒细胞凋亡。这可能解释了在呼吸道合胞病毒毛细支气管炎期间这些细胞在呼吸道的特征性积累,也可能促进这些细胞延长寿命,以保护呼吸道免受病原体进入健康婴儿呼吸道的感染。gydF4y2Ba

用荧光脱氧核糖核酸结合染料Hoechst 33342(Sigma Chemical Company Ltd)染色的可行(v),凋亡(a)和坏死(n)供体中性粒细胞。新鲜分离的中性粒细胞A)在3小时后在没有鼻灌注液(NLF),b)在24小时后没有NLF和c)在24小时后,在培养基中,50:50稀释的NLF从患有呼吸合胞增长的婴儿获得的NLF。病毒支气管炎。秤条=25μm。gydF4y2Ba
{kind=link}
{kind=link}
{kind=link}
{kind=link}
在37°C孵育24小时后,测量Dulbecco改良Eagle培养基中活、凋亡和坏死中性粒细胞的百分比。a)中等(n=13)。b)从因呼吸道合胞病毒细支气管炎(RSV-NLF)住院的婴儿获得的鼻腔灌洗液(NLF) (n=25个样本)。c)婴儿对照NLF (n=19个样本)。d)成人对照NLF (n=18个样本)。每个样本数据点都显示了每个数据集的中值(粗条)和95%置信极限(细条)。比较各组(RSV-NLF、婴儿NLF和成人NLF)的活细胞、凋亡细胞和坏死细胞数(活细胞数、凋亡细胞数和坏死细胞数)与单独在培养基中生长的细胞数。**: p<0.01,经Mann-Whitney检验。gydF4y2Ba
{kind=link}

{kind=link}
在白细胞介素(IL)-8浓度增加的情况下,37℃孵育24小时后,测量(n=8) Dulbecco改良Eagle培养基中活(□)、凋亡(▪)和坏死(汉化)中性粒细胞的百分比。Kruskal-Wallis法显示,随着IL-8剂量的增加,各生存指标均有显著差异(p<0.01)。gydF4y2Ba
致谢gydF4y2Ba
作者要感谢G. Parker (Vanguard Medica, Surrey, UK)和E. Lilly提供VML 295 (LY293111),以及S. Sarau (SmithKline Beecham, King of Prussia, USA)提供SB-237844。gydF4y2Ba
脚注gydF4y2Ba
↵gydF4y2Ba编辑评论见第515页。gydF4y2Ba
- 已收到gydF4y2Ba2001年9月7日。gydF4y2Ba
- 接受gydF4y2Ba二○○二年二月二十一日。gydF4y2Ba
- ©ers Journals LtdgydF4y2Ba
参考gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba









