

中性粒细胞炎症细胞外陷阱及其影响:概述
 比达尔Delgado-Rizo,
比达尔Delgado-Rizo, 马可·a·Martinez-Guzman,
马可·a·Martinez-Guzman, 莉莉安娜Iniguez-Gutierrez,
莉莉安娜Iniguez-Gutierrez, Alejandra Garcia-Orozco,
Alejandra Garcia-Orozco, Anabell Alvarado-Navarro和
Anabell Alvarado-Navarro和 玛丽Fafutis-Morris*
玛丽Fafutis-Morris*- 大学生理学、瓜达拉哈拉、瓜达拉哈拉、墨西哥
除了物理障碍,中性粒细胞被认为是免疫的第一道防线的一部分。他们可以在血液中,寿命为6 - 8 h,在组织中,在那里他们可以持续7天。中性粒细胞的机制,利用宿主防御的吞噬作用,脱粒,细胞因子的生产,,最近,中性粒细胞胞外陷阱(净)生产。网发布的DNA结构由于染色质decondensation和传播,他们因此占领三到五倍的体积浓缩的染色质。一些蛋白质坚持网,包括组蛋白和30多个组件主要和次要的颗粒,其中组件与杀菌活动如弹性蛋白酶、髓过氧化物酶、组织蛋白酶G,乳铁蛋白,pentraxin 3,白明胶酶,蛋白酶3 LL37, peptidoglycan-binding蛋白质,和其他与杀菌活动能够摧毁毒性因子。三个模型NETosis日期。(一)自杀NETosis,持续时间为2 - 4 h,是最好的描述模型。(b)与核DNA释放NETosis至关重要,中性粒细胞释放网没有表现出失去核和质膜内5-60最小,它独立于活性氧(ROS)和英国皇家空军/默克/ ERK通路。(c)发布的最终类型是至关重要的NETosis线粒体DNA依赖ROS和与gm - csf和脂多糖刺激后产生的。最近的研究显示中性粒细胞作为更复杂的免疫细胞能够精确控制颗粒酶释放的离子通量和可以释放免疫调节细胞因子和趋化因子与免疫系统的各个组件进行交互。因此,他们可以扮演一个关键的角色在自身免疫和autoinflammatory和代谢疾病。综述,我们打算显示中性粒细胞所扮演的两个角色:作为第一道防线对微生物和导致各种疾病的发病机制,如自身免疫性、autoinflammatory和代谢疾病。
定义、机制和功能
除了物理障碍,中性粒细胞被认为是免疫的第一道防线的一部分。他们可以在血液中找到,在那里他们有寿命的6 - 8 h,在组织,在那里他们可以持续7天(1)。他们是第一个细胞免疫系统迁移到一个站点的炎症,他们扮演着重要的角色在消灭病原体和细胞因子的生产(2)。
宿主防御的机制,中性粒细胞进行吞噬作用,脱粒,细胞因子的生产,,最近,中性粒细胞胞外陷阱(网)的生产(3)。
中性粒细胞胞外陷阱被发现由武井等。4)在1996年作为一个与细胞凋亡和坏死细胞死亡的途径不同。他们正在调查的关系中性粒细胞和嗜中性粒细胞激活使用phorbol-12-myristate-13-acetate死亡(PMA);他们观察形态学变化截然不同,那些发生在细胞凋亡和坏死,导致他们表明另一种细胞死亡通路可能发生。首先,他们描述了融合的multilobulated核中性粒细胞和减少染色质结构紧凑。核膜然后分解时胞质细胞器保持不变。3 h后,细胞外膜是中断机制依赖于活性氧(ROS) (4)。最后,在2004年,Brinkmann et al。(3)进一步详细的这个过程,并命名为NETosis(3)。
中性粒细胞胞外陷阱是DNA结构由于染色质decondensation发布和传播,因此他们占领三到五倍的体积浓缩的染色质。一些蛋白质坚持网,包括组蛋白和30多个组件的主要和次要颗粒(5),其中组件与弹性蛋白酶等杀菌活性,髓过氧化物酶、组织蛋白酶G (CG),乳铁蛋白,pentraxin 3,白明胶酶,蛋白酶3 (PR3) LL37, peptidoglycan-binding蛋白质,和其他与杀菌活动能够摧毁毒性因素(6- - - - - -9)。
值得一提的是,核内染色质组蛋白具有内在的抗菌活性。DNA作为螯合剂对阳离子由于其磷酸二酯骨架,使其外部和内部膜的破坏能力铜绿假单胞菌(10,11)。观察组蛋白的抗菌效果在1950年代由詹姆斯•Hisch H2A已被建议作为一种最有效的抗菌药物,特别反对大肠杆菌、弗氏志贺菌、志贺氏杆菌sonnei、肠炎沙门氏菌、鼠伤寒沙门氏菌、肺炎克雷伯菌、铜绿假单胞菌、白色葡萄球菌,金黄色葡萄球菌。此外,重组H4具有抗菌活性金黄色葡萄球菌和丙酸菌属(12)。组蛋白的抗菌效果一直观察不仅对细菌寄生虫。王等人发现H2A、H2B降低了复制的利什曼虫种虫害promastigotes 50% (13)。
三个模型NETosis日期。自杀NETosis,持续时间为2 - 4 h,是最好的描述模型(14),尽管它的分子过程不是完全理解(15)。逐步,它开始于中性粒细胞的激活,通过刺激的识别,导致他们包和激活NADPH氧化酶(NOX)复杂通过蛋白激酶C (PKC) / Raf默克/ ERK,以及增加胞质Ca+ +;这些阳离子作为肽基精氨酸酶脱氨酶的辅因子4 (PAD4),核酶,促进组蛋白的脱氨基作用,因此修改染色质的氨基酸允许decondensation促进正电荷的损失所需的组蛋白与DNA之间的相互作用(16,17)。
活性氧像第二信使自杀NETosis通过促进核膜逐渐分离和丧失,分解成小个体的囊泡。随后,染色质分散在细胞质中,与细胞质蛋白质和颗粒混合毒素。净的形成依赖于弹性蛋白酶和髓过氧物酶运输从颗粒核(18)。最后,染色质在细胞外释放通过膜孔和细胞裂解。自杀NETosis取决于ROS的组蛋白citrullination PAD4,它允许染色质decondensation (16,17,19),最后释放DNA细胞外陷阱(ETs)(图1)(20.,21)。
图1
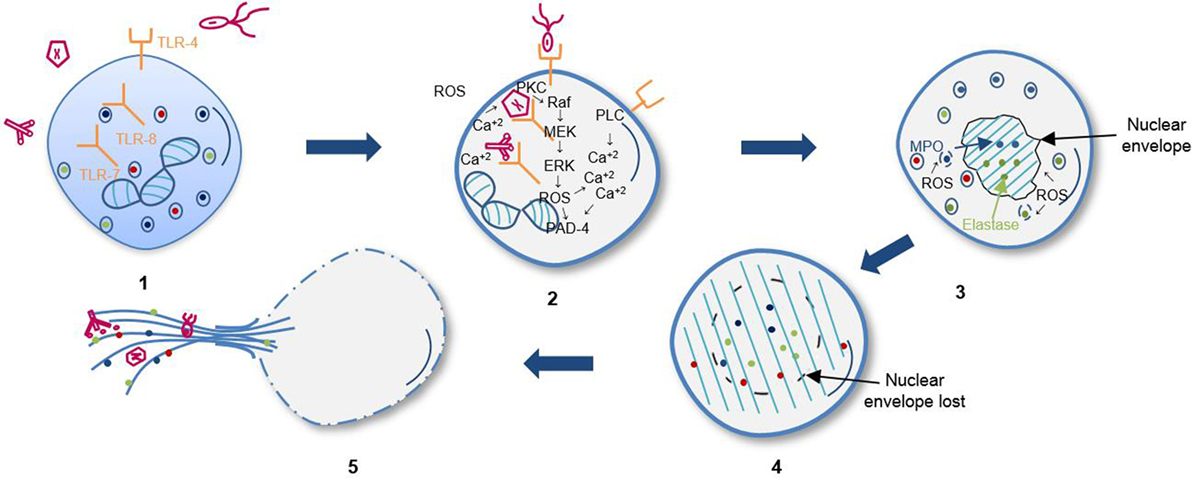
图1所示。自杀NETosis顺序步骤。(1)通过受体识别的刺激。(2)激活Raf MEK / ERK激酶途径和增加胞质钙导致gp91phox磷酸化激活的氧化酶NADPH复杂和随后的活性氧(ROS)的生产。(3)弹性蛋白酶和髓过氧化酶(MPO)易位的核嗜苯胺蓝的颗粒由活性氧和其他未知因素。Decondensation染色质和细胞核的小叶形状的损失。(4)核和颗粒膜,协会decondensed染色质细胞质组件。(5)的血浆的DNA细胞外膜和释放的陷阱。
在NETosis至关重要,中性粒细胞释放网没有表现出失去核和质膜内5-60 min,和它发生独立的ROS和皇家空军/默克/ ERK通路。这个过程包括释放核DNA通过三个形态变化:(a)核膜增长和囊泡释放,核decondensation (b)和(c)核膜破坏(14,22- - - - - -24)。这种类型的NETosis触发通过toll样受体识别的刺激(通常)和补体C3蛋白质受体(25- - - - - -27)。此外,互动在血小板糖蛋白Ibβ2整合素(CD18)中性粒细胞激活ERK可能诱发网络形成,PI3K和src激酶(图2)(28)。原子核释放后,这些中性粒细胞仍然能够吞噬病原体,它们的寿命不受DNA影响损失(24)。
图2
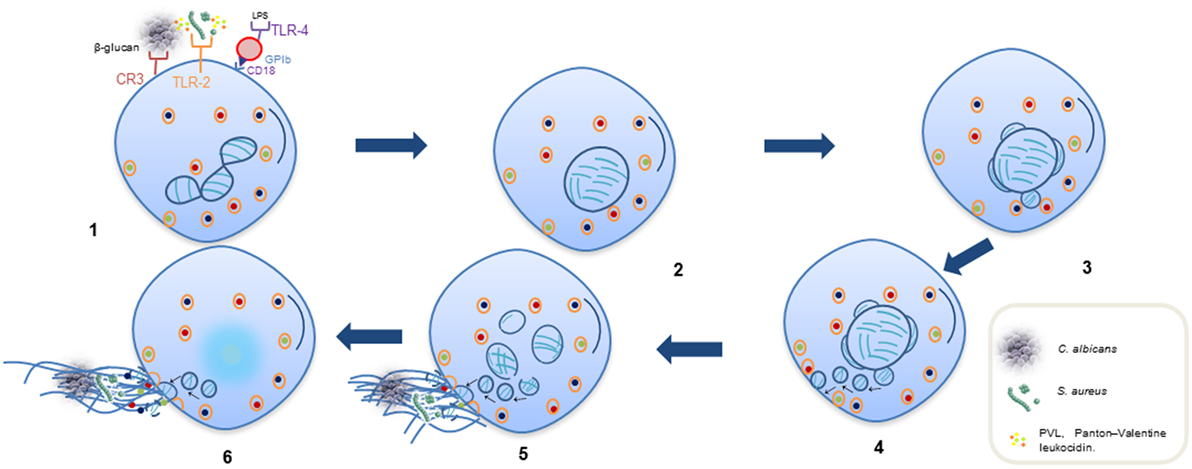
图2。顺序步骤NETosis至关重要。(1)通过受体识别的刺激。(2)损失的小叶和多核核的形状。(3,4)外部和内部核分离膜和崭露头角的囊泡。(5)囊泡在细胞质中含有DNA纤维形式的珍珠串,接近密集的细胞质颗粒向完整的原生质的膜。(6)释放DNA细胞外陷阱释放通过细胞表面的一个小区域;一些细胞质颗粒也原生质的膜融合并释放到细胞外空间与DNA。
最后,Yousefi描述了另一种类型的重要NETosis依赖ROS,线粒体DNA的释放,而不是核DNA;这个过程导致净形成80%的15分钟内中性粒细胞通过识别C5a或脂多糖(LPS) (29日)。
本文将重点讨论网络参与微生物感染、自身免疫、代谢紊乱。净形成的机制的详细描述超出了这个工作的范围,但可以发现其他地方(额外的评论30.)。然而,重要的是要强调高机动组框1 (HMGB1)蛋白表达血小板主要内源性网络形成的诱导物,不仅在感染过程,而且在无菌炎症(31日,32)。
网和病原体
有几种炎症过程引发的细菌,病毒,寄生虫和真菌。由于细菌及其代谢产物刺激净第一次描述了形成机制的启动和逃税网由病原体已经深入研究(表1)。
表1
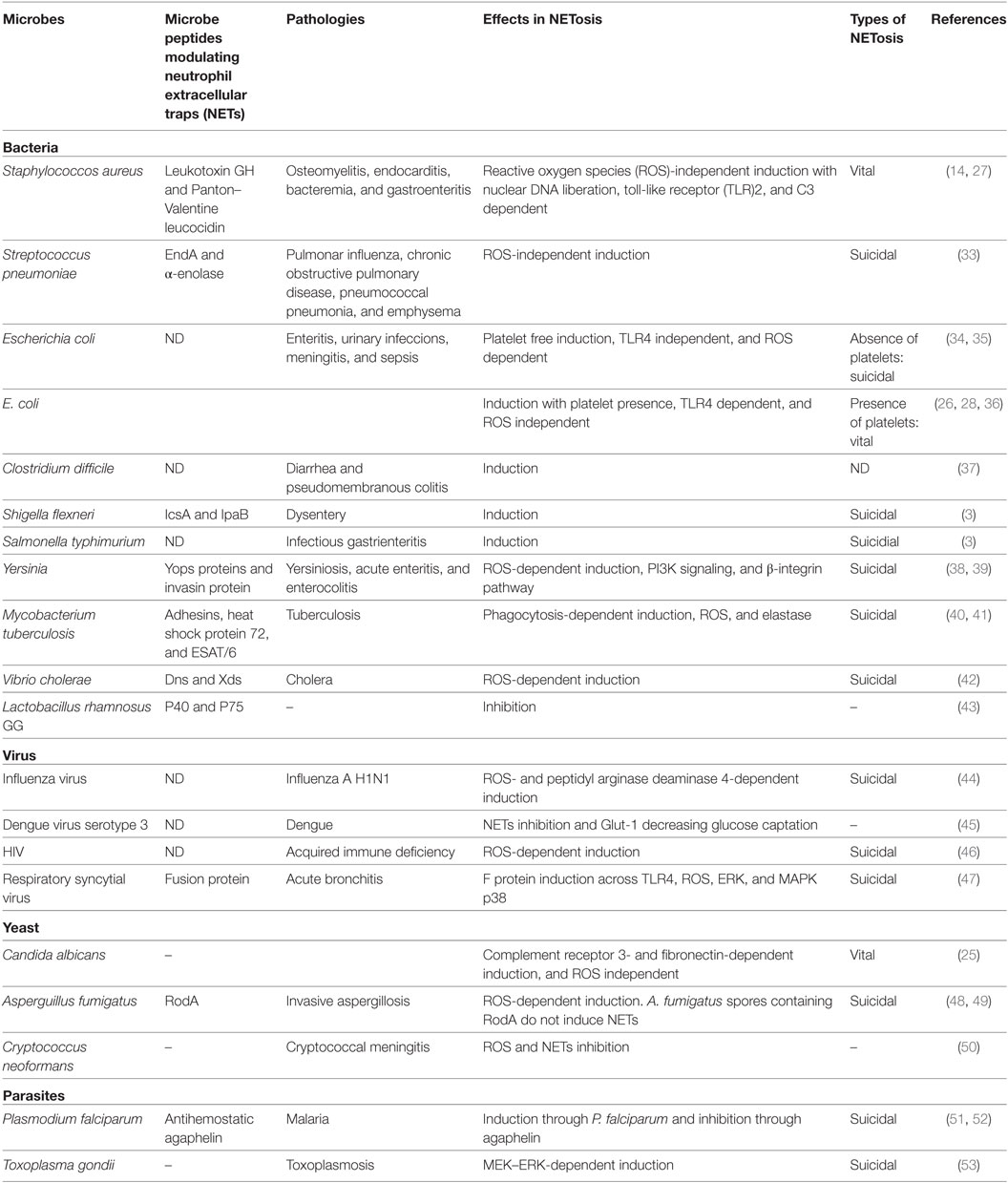
表1。微生物诱导NETosis。
细菌
葡萄球菌葡萄球菌
金黄色葡萄球菌是一种革兰氏阳性细菌,主要居住在潮湿的前鼻腔鳞状上皮(54),被称为“超级细菌”因其逃避免疫系统的能力,抵抗抗生素治疗(55)。它会导致病态如骨髓炎,心内膜炎,菌血症,肠胃炎,严重的炎症反应相关(56)。
2004年在经典化验由Brinkmann金黄色葡萄球菌第一次被用来刺激释放NETosis (3)。2010年,当诱导网的形成和研究其分子机制,Pilscek等人发现金黄色葡萄球菌导致明显更快的ROS NETosis,是独立的,他们叫“重要NETosis”(14)。
金黄色葡萄球菌分泌多种毒力因子,使其逃避宿主的免疫系统,包括金黄色葡萄leucocidin (PVL),白细胞毒素GH (LukGH),白细胞毒素,gamma-hemolysin,氨基端ArgD肽,其中LukGH PVL促进网通过氧化pathway-independent机制(14,57,58)。
细菌入侵促进净形成固定的病原体,阻碍他们的传播。这种先天免疫反应是通过巨噬细胞吞噬作用和细胞因子生产的支持。Thammavongsa等人报道金黄色葡萄球菌可能有细胞毒性影响巨噬细胞通过这些蚊帐,自净孵化与核酸酶和腺苷合成酶源自这种细菌促进脱氧腺苷的形成,能够诱导细胞死亡(59)。
链球菌肺炎
链球菌引起的肺炎革兰氏阳性细菌,通常可以找到人类呼吸道(60),在净感应已经建立(和它的作用3)。
人们已经发现,在肺部感染,类似金黄色葡萄球菌,肺炎链球菌能够产生毒力因素(恩达),降低DNA甚至让它逃离网内的细菌被发现后,促进从上呼吸道细菌传播到肺部,然后血液中(61年- - - - - -63年)。
中性粒细胞胞外陷阱释放也被卷入的开发和并发症由于继发感染呼吸道疾病如慢性阻塞性肺疾病、肺炎、肺气肿(64年- - - - - -66年)。中性粒细胞,当过度招募了肺组织感染,扰乱微循环,促使更多的NETosis肺肺泡。此外,肺功能障碍患者显示更高水平的细胞外比轻度肺部疾病患者DNA,表明网参与气流阻塞,使慢性炎症反应(67年,68年)。
埃希氏杆菌属杆菌
大肠杆菌是一种革兰氏阴性细菌在人体胃肠道出生时(69年),它是主机微生物群中最丰富的兼性厌氧菌。已经与病态如肠炎、泌尿系统感染、脑膜炎、败血症(70年)。
最后,坎巴人等人发现网显著诱导中性粒细胞刺激时引起感染性休克患者的血清大肠杆菌,可能通过激活通常为C3补体受体或C5a (34)。
据报道在几个研究败血症是加重网及其组件(71年- - - - - -73年),因为他们与dna降解以及抗生素治疗变弱组织损伤(74年)。
中性粒细胞能够辨别不同病原体和病毒的有限合伙人之间为了诱发网络形成和释放组织因子(TF)的thrombogen涉及系统性炎症反应介导的激活凝血系统,描述感染性流程(36,75年)。
大肠杆菌应变阿/博士一直在研究小儿腹泻,它已经被证明可以促进上皮细胞的几个信号通路的激活,尤其是参与肠道屏障功能和结构损伤(76年)。关于这个过程之间的关系和渔网,Marin-Esteban等人表明,这些结构能够捕获、固定,消除细菌。然而,当中性粒细胞与上皮细胞系cocultured CaCo-2 / TC7和细菌,前者生产网和破坏上皮细胞。出于这个原因,他们建议网可能参与肠道上皮细胞损伤以及其他肠道炎症性疾病35)。
抗菌肽的浓度(AMP) LL37尿路细胞无法消除引起的感染大肠杆菌。然而,LL37来自招募中性粒细胞明显减少细菌殖民化(77年)。因为LL37与网(3),促进他们形成和稳定性(78年),似乎它扮演相关角色消灭病原体的配合网,虽然这还没有被证明。
梭状芽胞杆菌难相处的
艰难梭状芽胞杆菌导致人类,腹泻和pseudomembranous结肠炎通常由于滥用抗生素(79年),严重损害宿主居民微生物群,导致失调(80年)。
艰难梭状芽胞杆菌人类微生物群是一个正常的组成部分,与其他居民及其对营养物质的竞争物种通常可以防止过度繁殖(81年)。然而,当微生物群改变,营养供应增加以及减少继发性胆酸的生产,它允许梭状芽孢杆菌殖民的肠道(82年,83年)。
艰难梭状芽胞杆菌能够把通过肠毒素引起的损失的能力紧密连接。当梭状芽孢杆菌接触细胞内脏相关的淋巴组织,它可以促进生产的促炎细胞因子和趋化因子如白细胞介素1β(IL-1β),引发,CXCL5促进中性粒细胞的招聘(84年,85年)。
中性粒细胞不仅减少微生物毒素分泌安培和弹性蛋白酶的功能也产生网,这可能采取行动弥补受伤的部位有效阻碍肠道上皮细胞梭状芽孢杆菌传播(37)。
志贺氏杆菌flexneri
弗氏志贺菌是一种革兰氏阴性细菌致肠病的通常被摄入受污染的食物和饮料,其感染可能会导致痢疾的主机。志贺氏杆菌通过M细胞可以遍历肠道内腔;一旦到了那儿,它感染上皮细胞和可能传播水平。作为响应,核因子k B (NF-κB)在感染细胞被激活,产生吸引的中性粒细胞迁移引发感染的组织,在那里neutrophil-derived弹性蛋白酶降解微生物毒力因素(86年,87年)。
Brinkmann等人发现美国flexneri被困在网在体外和测试的能力DNA-associated弹性蛋白酶降解毒性因素IcsA和计划(3)。
佩尔多莫坚称等人表明,志贺氏杆菌发病机理,在体外中性粒细胞轮回是必需的志贺氏杆菌入侵与强烈的中性粒细胞浸润区(88年)。
沙门氏菌沙门氏菌感染
沙门氏菌是属兼性厌氧细菌细胞内。尽管许多物种可以发现在肠道微生物群,这个属美国沙门氏菌感染是一种传染性胃肠炎的主要原因。殖民肠道后,它可以进入肠上皮细胞,M细胞,树突状细胞(dc),,最后,在巨噬细胞到达黏膜下层。美国沙门氏菌感染时间内复制,它可以表达一些毒性因素:丰富,鞭毛菌毛,T3SS (89年)。它也能够使用超氧化物歧化酶称为SodCl抵消在白细胞的吞噬体(ROS的活动90年,91年)。
鼠伤寒沙门氏菌已被证明刺激网Brinkmann et al。他们还表明,它实际上是被困和消除组件的网,包括颗粒蛋白H2A组蛋白(3)。
鼠疫
鼠疫enterocolitica因果代理yersiniosis,急性肠炎和小肠结肠炎。它侵入上皮细胞并把派尔集合淋巴结补丁和影响紧密连接通过减少occludin claudins 5和8 (92年)。所有三个致病性的物种鼠疫,即y伪,y enterocolitica,鼠疫耶尔森氏菌属,主要的目标是把他们的效应蛋白(称为Yops)为中性粒细胞,巨噬细胞和DCs。此外,鼠疫耶尔森氏菌属抑制中性粒细胞的氧化破裂为了促进自己的细胞内生存使用Yops蛋白(93年)。
2015年,Mollerherm等人证明,血清型0:3,0:8,0:8y enterocolitica引起网在体外在1 h的孵化。然而,净感应减少随着培养时间的增加,表明网可能会退化的影响+ +- - - Mg+ +端依赖核酸酶(38)。
鼠疫的伪雇佣了一个专门的叫做侵袭素的蛋白质来破坏肠道上皮细胞。侵袭素是一种高蛋白质介导的粘合剂y伪绑定到β1 M细胞的整合蛋白;然而,这种绑定诱导活性氧产量和净形成(39)。
分枝杆菌肺结核
结核分枝杆菌是一种专性需氧芽孢杆菌导致肺结核。这是一个最成功的胞内病原体关于其逃避免疫系统的策略。它主要感染呼吸系统,但它也可能影响其他器官。细胞被膜含有丰富,相比其他致病细菌,它不会产生毒素。它使用吞噬细胞复制和传播在宿主生物体的一种方式。
它已经表明,不同基因型诱导网与中性粒细胞cocultured时。尽管网有效陷阱和阻碍分枝杆菌的传播,NET-derived组件无法杀死他们(40)。
的免疫逃避机制结核分枝杆菌利用增加细胞凋亡的中性粒细胞,以阻止他们创造肉芽肿,这是由免疫细胞结构针对原发感染,含有芽孢杆菌(94年)。巨噬细胞凋亡中性粒细胞的efferocytosis导致免疫反应对促炎。热休克蛋白72被发现在凋亡和坏死细胞(95年)和DNA内的网,它也需要消除结核分枝杆菌(41)。
弧菌霍乱
霍乱弧菌是革兰氏阴性细菌广泛认可因为O1和O139霍乱流行病引起的血清型。这种细菌可以发现在人类胃肠道和水生环境中;认识提高,感染通常遵循摄入受污染的海鲜和水。当霍乱弧菌到达肠道,它分泌霍乱毒素以及丰富的血凝素,蛋白酶和溶血素。最后,霍乱弧菌诱导细胞因子的生产以及中性粒细胞招募肠道(96年)。
据报道,霍乱弧菌诱发网在在体外与中性粒细胞。然而,霍乱弧菌分泌核酸酶Dns和Xds逃避机制,允许它逃离网,从而允许它继续感染过程。因此,网没有显示包含感染的保护机制霍乱弧菌(42)。
乳酸菌喂食
乳杆菌被认为是一个重要的益生菌的微生物群。它能够坚持肠道上皮细胞,抵抗胃酸和胆汁(97年)。
革兰氏阳性细菌,主要研究了恢复肠道屏障的能力,因为它减少了上皮损伤引起的溃疡性结肠炎(UC)和克罗恩病(CD) (98年)。
乳杆菌GG调节肠道菌群通过刺激免疫系统的反应,通常在免疫细胞(99年)。
它已被证明喂食GG抑制引起的净形成微生物(金黄色葡萄球菌和大肠杆菌)或化学物质(佛波醇12-myristate 13-acetate,更好地称为PMA),可能由于未知其抗氧化活性和分泌蛋白(43)。
病毒
流感
甲型流感病毒是造成超过5000万人死亡,1918年,最近,2009年全球大流行造成18000人死亡。流感病理特点是过度的中性粒细胞招募到肺部,这是由CXCR2促成的。流感模拟依赖PAD4网(One hundred.)。α-Defensin-1与网有关抑制病毒复制的封锁PKC途径。
另一个组件的杀菌网刺激LL37甲型流感病毒,这被证明能增加净产量以应对这种病原体在体外(44)。此外,arginine-rich H3和H4组蛋白对病毒聚合和中和很重要。孵化的甲型流感病毒H4被证明导致显著降低病毒复制在上皮细胞;相比之下,H4是灭活与爆发的流感病毒H1N1孵化时,这可能会强调它的重要性在回应这个病原体(101年)。
过多的中性粒细胞浸润的缺点包括损伤组织由安培和广泛的肺泡毛细血管网(67年,68年)。
登革热
登革病毒(DENV)是一种单链RNA病毒属于黄病毒科。感染登革热的血清型(1 - 4)的影响,从轻微发热严重的登革热,原名登革出血热(45)。登革热感染的发生率增加了近年来(102年);因此,有必要了解用于对抗这种病原体的宿主防御机制。
中性粒细胞胞外陷阱能够抑制由捕获病毒感染在他们的结构。然而,它已经证明,而不是刺激网,DENV-2抑制它们在体外。Moreno-Altamirano等人发现一个降低80% PMA-stimulated-NET由中性粒细胞与DENV-1前孵化后形成。这种抑制作用是由Glut-1-mediated葡萄糖吸收的干扰(45),净释放代谢要求(103年)。
人类免疫缺陷病毒1
1人类免疫缺陷病毒(hiv - 1)是一种免疫系统病毒和取向。目前有超过3500万人感染,约200万每年获得感染(104年)。它已经表明,CD4 CCR5和趋化因子受体CXCR4,充当病毒受体条目,它允许不仅感染的CD4 T细胞抗原递呈细胞如巨噬细胞和DCs。然而,大多数血清血浆来源于激活T细胞,病毒复制是快速和高效的(105年)。
中性粒细胞识别HIV-1-derived核酸通过TLR7和TLR8。之后,他们释放活性氧为了引起网络的形成。这些结构陷阱,可能包含,并消除艾滋病毒通过髓过氧物酶的作用和α-defensins,一直认为这两种抗病毒活动。艾滋病毒,作为逃避的一种机制,促进il - 10由DCs生产,从而抑制ROS和净释放(46)。
呼吸道合胞体病毒
呼吸道合胞病毒(RSV)是在一岁的婴儿住院的主要原因(106年);因此,它代表了最重要的一个儿科感染。RSV引起急性支气管炎,粘膜和粘膜下水肿,腔的阻塞和细胞碎片形成的上皮细胞、巨噬细胞、纤维蛋白和粘蛋白。RSV感染DCs和减少他们的激活T细胞抗原递呈能力(107年)。
生成网在体外,RSV可能刺激中性粒细胞,反过来,这些都是样本所示支气管肺泡灌洗液的严重疾病患者下呼吸道RSV所致。净形成预防RSV传播,但似乎无法杀死病毒(108年)。此外,RSV F蛋白也能够引起网通过TLR4。尽管有这些网作为病毒宿主,它们的存在可能加剧炎症症状,促进腔的闭塞与结构组成的粘液和DNA (47)。
真菌
假丝酵母白色的
白色念珠菌通常是发现殖民粘膜、皮肤和口腔健康个体,导致免疫功能不全的疾病只有主题,如胰腺炎或肾功能不全患者,患者在抗生素治疗或中央静脉导管,和胃肠道手术后的病人。白念珠菌形态变化从酵母到菌丝生产几个毒力因素如Als3和Ssa11侵袭素,将自己变成一个入侵的病原体。上皮细胞和巨噬细胞识别白念珠菌响应通过释放趋化因子,吸引中性粒细胞(109年,110年)。中性粒细胞能够陷阱和消除白念珠菌在它的酵母或菌丝的形式通过释放网(111年)。为了实现快速NETosis响应,β-glucan必须被补体受体3在菌丝,纤连蛋白,细胞外基质的组成部分,必须存在。这些元素是同型的细胞聚合所需支持的网,但独立的ROS生产(25)。
曲霉属真菌来自烟
来自烟曲霉属真菌在健康受试者人类微生物群的一部分。在免疫抑制的个体,它负责侵袭性曲霉菌感染,导致霉菌的感染,慢性肉芽肿性疾病患者的患病率和死亡率(112年)。通过吸入孢子发生感染,而不是被免疫系统细胞,在呼吸道和改变他们的居住形态从酵母到菌丝,肺部感染,导致肺炎和其它器官的感染。类似于白念珠菌它产生侵袭素,让它坚持宿主细胞(113年,114年)。
释放引起的网答:来自烟体外需要激活的氮氧化物(48)。此外,p46−−/老鼠不能形成网(115年)。尽管网捕获和消除的是必要的答:来自烟菌丝,孢子引起的这些都不是由于的存在又在孢子细胞壁(49)。
隐球菌spp。
新型隐球菌是一种机会性致病酵母。孢子是吸入后引起感染和进入肺泡空间,他们潜伏到免疫失衡发生和导致隐球菌脑膜炎(116年)。
新型隐球菌具有荚膜多糖,赋予它能够调节宿主免疫系统。特别是,它能够调节净产量。中性粒细胞孵化与菌株的胶囊包含glucuronoxylomannan (GXM)和galactoxylomannan没有有效的活性氧或生产商网,即使PMA刺激。中性粒细胞与菌株没有荚膜GXM孵化时,网有效地产生;然而,ROS没有观察到。因此,荚膜GXM提高毒力调节电阻网。最后,NET-associated安培的微生物活动,如弹性蛋白酶、髓过氧化物酶、胶原酶,和组蛋白,需要杀死病原体(50)。
寄生虫
疟原虫恶性疟原虫
恶性疟原虫疟疾的因果代理,也称为疟疾。它严重影响5岁以下儿童,代表90%的死亡相关疾病(117年)。疟疾是一种由蚊子传播的血液疾病。疟原虫种虫害感染红细胞,导致炎性细胞因子的生产;这些抑制红细胞生成,导致贫血。入侵是由蛋白质感染红细胞,促进血管内皮粘附在组织和器官和诱导炎症反应和凝固(118年,119年)。
这种感染过程导致血管损伤,损伤内皮细胞,激活血小板,单核细胞和中性粒细胞。中性粒细胞释放网,这些结构可以在儿童感染的血液循环恶性疟原虫坚持红细胞和寄生虫。此外,α-dsDNA抗体被发现在这些患者中,可能参与病理学的发展,加重免疫反应和自身免疫过程(51)。另一方面,受感染蚊子的腺体产生antihemostatic agaphelin,能够抑制中性粒细胞趋化性,封锁血小板聚集由组织蛋白酶/弹性蛋白酶,并减弱neutrophil-induced凝血(52)。
弓形虫刚
刚地弓形虫的因果代理弓形体病,感染了全世界三分之一的人口由于摄入受污染的食物。
感染弓形虫诱导中性粒细胞招募受感染的网站(120年)。因此,鼻内感染的小鼠模型中,中性粒细胞减少传播的病原体在网捕获并杀死它,从而证明主动入侵不是净形成所必需的。这个观察随后人类所示,进一步表明净形成是依赖于mek erk (53)。
网和自身免疫
中性粒细胞和网在宿主体内平衡扮演双重角色。他们都保护宿主免受传染病;然而,他们也引起病理改变,自身免疫性和autoinflammatory一样的疾病。
牛皮癣
牛皮癣是一种慢性炎症性疾病,影响皮肤和特点是一个复杂的免疫反应,从细胞、分子和血管组件参与炎症过程的延续。迄今为止,辅助T 1 (Th1)和Th17淋巴细胞被认为是唯一的监管细胞免疫反应在银屑病病变(121年,122年)。然而,林等人表明IL-17十分由中性粒细胞和肥大细胞,这两个在银屑病斑块的细胞浸润(123年)。最初,干扰素α(IFN-α)和肿瘤坏死因子α(TNF-α)分泌血浆树突细胞(髓)被激活通过通常的识别LL37-nucleic酸复合物释放受损角质细胞分泌细胞因子。最初,pDCs激活通过通常的识别LL37-nucleic酸复合物释放的角化细胞受损,因此,pDCs释放IFN-α和TNF-α(124年)。这些细胞因子可能对DCs和巨噬细胞的激活和其生产IL-23 IL-1β,进而诱发的激活和生产IL-17, TNF-α,CXCL2, chymase,肥大细胞类胰蛋白酶的肥大细胞胞外陷阱(MCET)形成(125年)。此外,这些介质促进中性粒细胞迁移向表皮,他们可能变得激活IL-23 IL-1β和生产网。在一起,中性粒细胞和肥大细胞分泌IL-17和其他促炎介质增强中性粒细胞迁移,这将有助于形成Munro脓肿(126年)。
另一方面,IL-17角质细胞增加的表达等LL37 defensins beta-defensin 2 (HBD-2) S100A7, S100A8和S100A9 (127年,128年),调节细胞渗透MCET和网络的形成。在这种背景下,ET-derived DNA-LL37复合物通常生成。这些配合物可能导致pDC IFN-α激活和随之而来的生产,进一步促进银屑病NETosis和炎症病变即使没有感染(124年)(图3)。
图3
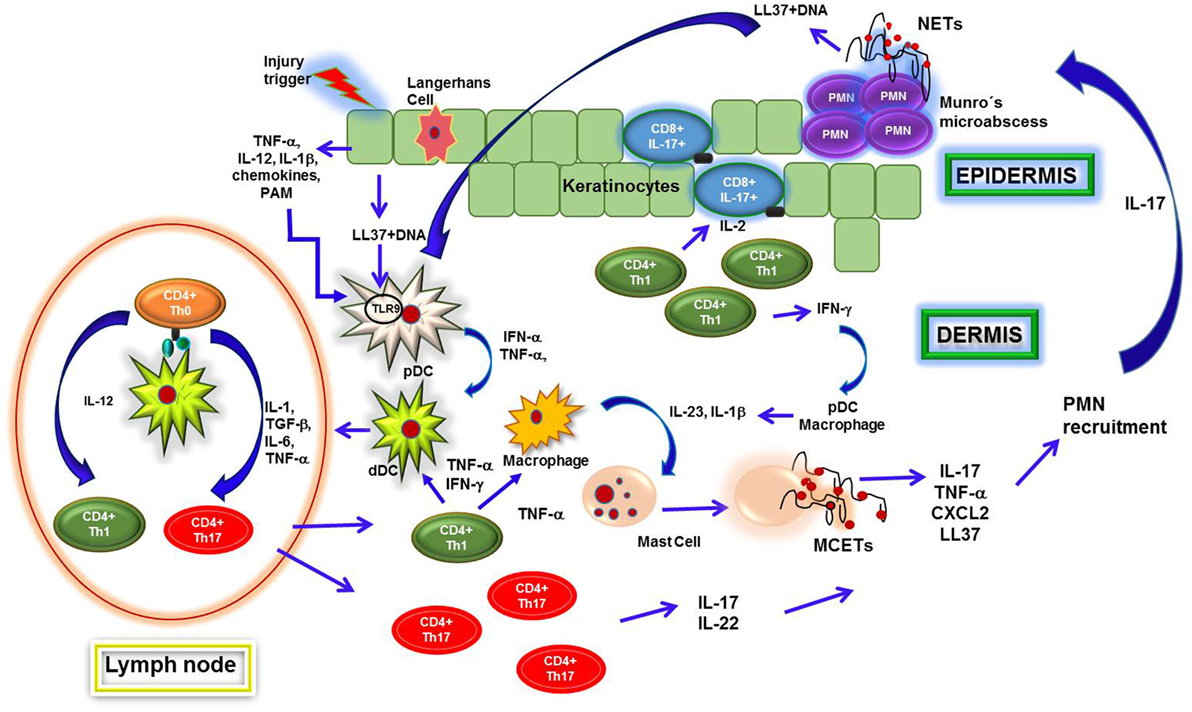
图3。中性粒细胞胞外陷阱(网)在银屑病斑块形成。银屑病病变患者是在皮肤损伤后,诱导角质细胞释放促炎细胞因子和抗菌肽。这样的分子,如抗菌肽LL37,形成复合物在绑定到DNA激活血浆树突细胞(髓)通过toll样受体相互作用9 (TLR9识别),导致他们分泌干扰素α(IFN-α)和肿瘤坏死因子α(TNF-α)。这些细胞因子调节真皮树突状细胞(ddc)激活,这区域淋巴节点迁移到现在的自身抗原天真的CD4 T细胞(Th0)。随后,T细胞分化成辅助T 1 (Th1)或Th17和迁移到真皮,分泌2,IFN-γ,TNF-α,il - 22生成时,和IL-17导致招聘和激活的巨噬细胞,如ddc,肥大细胞。这些细胞合成IL-23和白介素- 1β(IL-1β),从而诱导肥大细胞释放细胞外陷阱(MCETs)。因此,肥大细胞释放细胞内的内容以及IL-17和其他细胞因子,诱导中性粒细胞浸润到表皮,Munro形成的微小脓肿。中性粒细胞遇到高浓度IL-23和IL-1β在这个微环境,这使他们容易发布网。通过网形成,他们也分泌细胞内容包括IL-17,从而放大炎症过程和增加细胞招聘和角化细胞激活。最后,网LL37-DNA复合体的重要来源和IL-17,这两种激活pDCs和角化细胞,分别产生IFN-α和LL37。 Th1 cells induce activation of T CD8 IL-17-producing cells in epidermis and pDCs within dermis, perpetuating the inflammatory environment in psoriatic lesions.
系统性红斑狼疮
系统性红斑狼疮(SLE)是一种自身免疫性疾病,其特征是免疫复合物和高水平的IFN-α诱导的自身抗原由于未能消除产品来自凋亡或坏死细胞(129年,130年)。失去宽容对自体抗原导致autoreactive B细胞的活化和核酸和安培的自身抗体的生产,由浸润中性粒细胞释放,接受NETosis系统性红斑狼疮患者的皮肤和肾脏。生成的免疫复合物沉积在不同的组织,导致损伤和炎症反应,引起皮肤病变为主,肾炎和心血管疾病(131年,132年)。此外,调理自身抗原诱导髓分泌IFN-α,也被称为一个“IFN-α签名,”和诱导中性粒细胞形式网(133年,134年)。
系统性红斑狼疮患者,核材料来源于死细胞积累由于故障的排除。这些自体抗原呈现autoreactive在生发中心B细胞由滤泡树突细胞在二级淋巴器官,从而产生自身抗体对细胞组件来自NETosis和细胞凋亡(134年,135年)。生产的免疫复合物激活补体系统,诱发炎症,血管损伤、血栓形成和脑损伤。免疫复合物内化通过髓II型Fcγ受体介导内吞作用;之后,他们把TLR7和TLR9识别核内体,从而导致IFN-α-secreting pDCs和额外的激活形成网(136年- - - - - -138年)。
系统性红斑狼疮患者被发现拥有数字分组人口的中性粒细胞升高血液中被称为低密度粒细胞(爬)。这些都是不成熟的中性粒细胞快速凋亡和释放活性氧在体外,因此作为强有力的净诱导系统性红斑狼疮患者(132年)。通过网络形成,爬下细胞内的内容释放到微环境,包括几个分子如LL37,α-和β-defensins HMGB1;这些分子与核酸和激活pDC虽然TLR9识别刺激,后来引发IFN-α合成。它已经被观察到,在系统性红斑狼疮患者,IFN-α有力NETosis诱导物(139年),连同激活pDC-derived il - 6,促进分化的autoantibody-secreting autoreactive B细胞(140年)。另一个可能的机制自身抗原的释放HMGB1和核酸等凋亡细胞是通过二级坏死,这种现象时观察到凋亡的身体不被吞噬细胞。HMGB1然后用DNA和激活pDCs同事由于TLR9识别的识别和受体高级糖化终端产品(愤怒)pDCs,因此诱导pDC激活(141年)。DNA-HMGB1复合物也可能被通过B细胞receptor-TLR7/9-RAGE autoreactive B细胞;这导致自身抗体的生产。同样,DNA-HMGB1-immunoglobulin可能激活pDCs RAGE-FcR-TLR9相互作用,这将导致IFN-α合成和正反馈循环(图4)(142年)。
图4
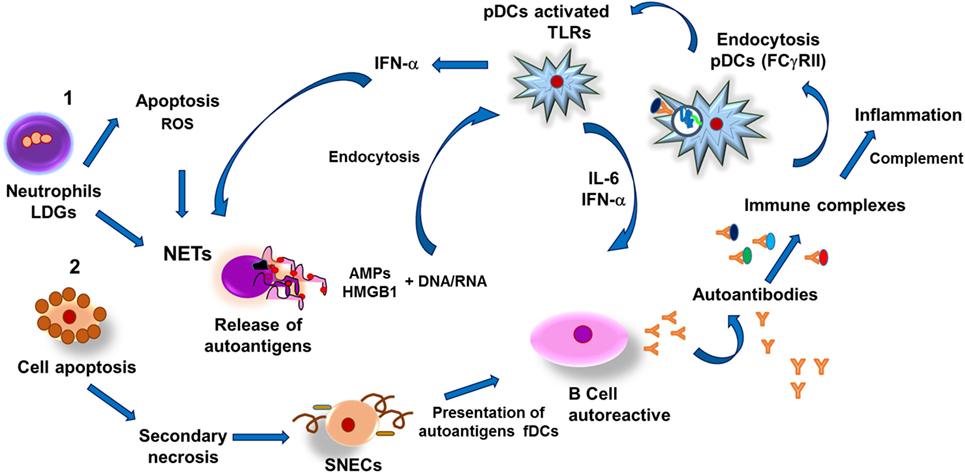
图4。系统性狼疮红斑狼疮的发病机制。(1)低密度粒细胞(爬)进行细胞凋亡和释放活性氧(ROS)和自身抗原,从而刺激中性粒细胞胞外陷阱的形成(网)以及释放抗菌肽(安培)和核酸(DNA, RNA)。核酸和安培形成复合物能够绑定到高机动组框1 (HMGB1)蛋白质,可被toll样受体血浆树突细胞(髓样),反应的合成干扰素α(IFN-α),从而促进网的形成和il - 6。两种细胞因子诱导分化的autoantibody-secreting autoreactive B细胞,导致免疫复合物的形成,激活补体系统,也容易被pDCs通过内化II型Fcγ受体(FcγRII)介导的内吞作用。通常核内体联想到的toll样受体)包含囊泡,从而激活IFN-αpDCs和合成,进一步诱导网和组织的炎症。此外,坏死细胞DNA-HMGB1复合物激活B细胞导致生产自身抗体和免疫复合物的形成,激活pDCs,导致IFN-α合成,从而建立一个积极的反馈。(2)诱导自体抗体生产的另一个途径可以是由自身抗原的释放进行继发性坏死和凋亡细胞产生继发性坏死细胞(SNECs);积累细胞的生发中心的二级淋巴器官促进表示自身抗原的滤泡树突细胞(fDCs) autoreactive B细胞和随后的免疫复合物的形成,导致持续的炎症过程,导致选取病人组织的损伤。
最后,这些产品的发布和持久的存在代表着自体抗原的来源,增强自身免疫和炎症过程,在系统性红斑狼疮导致组织损伤。
类风湿性关节炎
类风湿性关节炎(RA)是一种系统性自身免疫性疾病,其特征是持续的滑膜炎症,导致关节软骨和骨骼损伤(143年)。滑液在RA患者的滑膜腔与中性粒细胞,容易形成网成为渗透;此外,即使是循环中性粒细胞的RA患者更容易刺激NETosis比从健康受试者(144年,145年)。发生在其他自身免疫性疾病,网可以作为细胞外自身抗原的来源;例如,从组蛋白citrullination citrullinated肽产生通过PAD2和PAD4活动在中性粒细胞,可以发现即使在RA患者滑液(144年,146年,147年)。这种citrullinated肽被α-citrullinated肽抗体(acpa),形成免疫复合物,从而诱导净形成,导致中性粒细胞颗粒的释放内容以及关节细胞质自体抗原。他们也可能释放核因子kappa-β配体和受体激活B细胞激活因子,分别激活破骨细胞和B细胞(148年,149年),导致过度的先天和适应性免疫反应的关节和组织损伤。acpa检测血清的RA患者在疾病的早期阶段,在临床症状出现之前,他们因此代表了类风湿性关节炎的早期生物标志物(150年)。Khandpur等人发现,除了自身抗体净感应,IL-17 TNF-α可能也有这种能力,他们发现这些细胞因子在RA患者血清高(144年)。
高机动组框1是另一个自身抗原与RA发病机制有关。它可以发现升高血管翳的RA患者,特别是在软骨间期和地区组织缺氧(151年)。低氧胁迫下,细胞释放HMBG1和诱导炎性分子TNF-α和il - 1的生产表明HMGB1在RA与缺氧和炎症密切相关(152年)。此外,这种蛋白质结合IL-1αIL-1β形成复合物,增强关节的免疫反应,从而引发炎症(141年)。
一些患者可能产生毡状的综合症,严重的风湿性关节炎,体现在患者中性粒细胞减少和脾肿大。后者似乎与T细胞的寡克隆扩张和PAD4的自身抗体(153年)。
1型糖尿病
1型糖尿病(T1DM)是一种自身免疫性疾病,其特征是破坏β胰岛细胞在遗传个体,导致高血糖。的破坏β胰岛细胞也允许表示被autoreactive自身抗原的T细胞,紧随其后的是特定的自身抗体的生产β细胞抗原,包括谷氨酸脱羧酶自身抗体,insulinoma-associated蛋白质2自身抗体,和锌transporter-8自体抗体,用于临床上作为T1DM的预测和诊断,虽然他们不认为致病(154年- - - - - -157年)。
主要与T1DM的个体,浸润的CD8 T细胞CD4 T细胞和B细胞参与的破坏β胰岛细胞通过释放granzymes和穿孔素,FasL通路的激活和促炎细胞因子的生产,即IFN-γ和TNF-α(158年)。先天免疫反应细胞中也扮演着重要的角色在T1DM的发病机理,从巨噬细胞,单核细胞,DCs,可以找到和中性粒细胞浸润中的胰岛细胞,他们合成IFN-α和活性氧,从而促进促炎细胞因子的合成(154年,158年,159年)。
多项研究表明,T1DM和个人发展的危险疾病的患者遭受嗜中性白血球减少症(160年),这可能部分归因于增加NETosis和中性粒细胞浸润在胰腺组织(160年- - - - - -162年)。
中性粒细胞产生超氧化物和细胞因子暴露在高血糖的条件。糖尿病人,TNF-α并激活中性粒细胞升高形成网,随后,释放细胞内的内容,包括中性粒细胞丝氨酸蛋白酶,如中性粒细胞弹性蛋白酶(NE), PR3, CG (161年- - - - - -163年)。T1DM病人血清浓度升高NE和PR3,以及高水平的活动,这些酶(162年)。这些蛋白质是重要的在T1DM病机鉴于其含义的成熟和释放细胞因子IFN-α,IL-1β,地震,以及诱导通常的表达和激活胰岛炎的重要介质和胰岛β细胞的毁坏158年,164年)。他们也倾向于中性粒细胞炎症招聘网站,提供负面反馈和导致自身免疫性糖尿病发病机理(162年)。
小血管血管炎
小血管的血管炎(SVV)是一种全身性疾病,病因不明。SVV病人表现出血管炎症影响小动脉、小静脉,动脉和毛细血管,还可能涉及到在疾病恶化。在这些情况下,坏死性炎症发生在小血管和可能损害任何器官,最主要的是肾脏、肺、皮肤和周围神经(165年)。Antineutrophil细胞质抗体(ANCAs)可以发现在大多数SVV患者(166年)。由于长时间暴露于NETosis时释放的蛋白质,如髓过氧化酶(MPO), PR3,组蛋白,HMGB1, NE, ANCA生产的主要原因,这些蛋白质被认为是促炎介质,激活补体系统,导致内皮损伤。MPO和PR3 ANCAs的主要目标,它已经表明,α-PR3和α-MPO ANCAs诱导NETosis在活动性疾病;此外,高水平的DNA-MPO复合物与疾病相关联活动(167年- - - - - -170年)。ANDA-PR3和ANCA-MPO可能激活中性粒细胞的存在和延续炎症状态通过补体系统激活和嗜中性粒细胞趋化性向受伤的网站(171年)。
感染也可能诱发ANCAs通过分子拟态的生产。此外,微生物可能诱发网络形成,导致自身抗原释放(172年,173年)。然而,由于网发现了SVV患者在缓解期,同样重要的是要考虑到ANCAs的存在也可以帮助去除NET-derived产品和促进宿主体内平衡(174年,175年)。
血栓形成是由于TF的释放,细胞因子,以及其他炎症介质在NETosis发生由于感染、自身免疫性疾病和癌症(75年,176年)。
小血管血管炎患者血液中高浓度的网。爬下已经被提议作为网络的主要来源自发生成网(由于他们的能力177年)。这些蚊帐可能放大炎症过程,导致内皮损伤和激活补体旁路(170年,178年)。另外,组蛋白在网已经证明,导致血栓形成和促进TF生产,进而诱发凝血酶(165年,179年)。在小鼠模型中,没有观察到激活血小板刺激净形成和引发在深静脉血栓形成(180年)。最后,在SVV neutrophil-platelet总量的增加患者的循环已被证明与疾病关联活动,这表明SVV网和凝血酶的形成之间的关系(181年)。
Autoinflammatory疾病和网
痛风
痛风是一种autoinflammatory疾病单钠尿酸盐沉积的特征(MSU)晶体的关节,这吸引了白细胞和形式结构称为痛风石,调解组织损伤。经过吞噬细胞吸收,密歇根州立大学晶体是兴奋剂的免疫反应主要通过NLRP3 inflammasome-mediated IL-1β生产由于渗透压失衡引起的突然增加细胞内加上水侵和随后的钾钠的浓度稀释。此外,MSU-activated中性粒细胞分泌引发,TNF-α,il - 6。这些细胞因子不仅促进中性粒细胞招募还引起网络的形成。特别是,网可以参与痛风石形成,因为他们的组件(图密切相关5)(182年,183年)。
图5
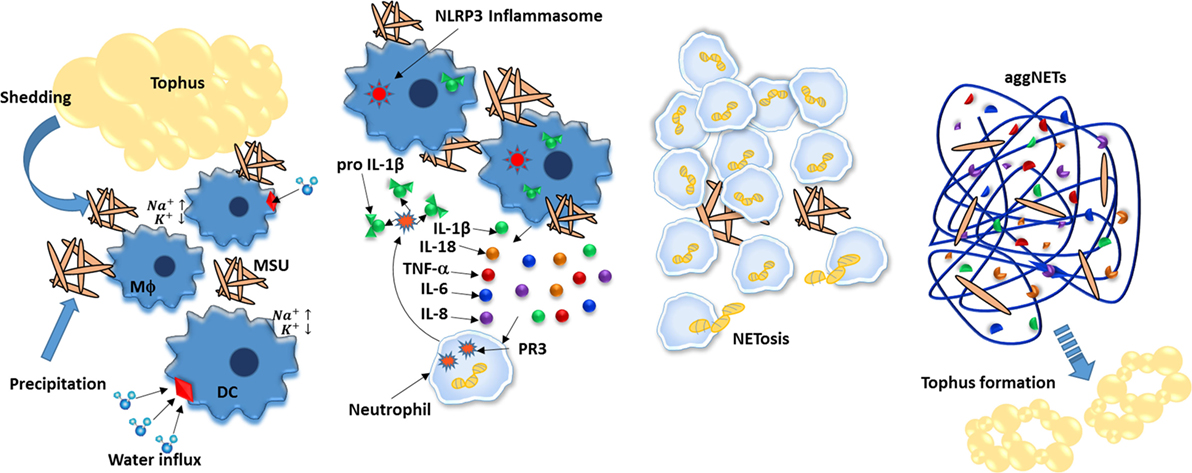
图5。中性粒细胞胞外陷阱(网)aliviate单钠尿酸盐引起的炎症(MSU)晶体。密歇根州立大学晶体是由降水和沉积在关节痛风结节的脱落,进一步通过吞噬细胞,如树突状细胞和巨噬细胞摄取。因此,增加细胞内钠诱发细胞浓度提高水吸收,从而稀释钾inflammasome-activation浓度阈值以下。地震和IL-1β分泌和调解招募中性粒细胞关节发炎;中性粒细胞进一步增加IL-1b水平裂开pro-IL-1b过程中由蛋白酶3 (PR3)。摄取MSU晶体时,中性粒细胞进行NETosis不仅降低MSU还封装了它,有效降低炎症的潜力。最后,聚合网(aggNETs)通过降解进一步阻止炎症原位细胞因子。随着时间的推移,aggNETs可能导致新的痛风石形成。
和其他疾病一样,网促进痛风炎症(已报告184年)。然而,与其他疾病不同,网似乎也发挥着重要的作用在调节炎症过程和停止痛风发作(185年)。最初,NETosis减少中性粒细胞密度,因为它们表明嗜中性粒细胞死亡。第二,DNA网封装MSU水晶和保护他们免受进一步的吞噬作用。最后,NET-derived蛋白酶活性细胞因子和废除其促炎效应(186年)。
克罗恩氏病
克罗恩氏病是一个复杂的系统性疾病,临床表现为胃肠道功能紊乱和回肠和结肠炎症(187年)。尽管炎症CD是一个重要组成部分,细胞成分参与其病理仍有些不清楚。关于中性粒细胞,他们的活动就改变。而chemokine-mediated迁移降低,活性氧产量提高;此外,细菌摄取似乎完全没有改变(188年)。
另一方面,净形成CD还没有被研究过。可以说,从活性氧产量提高,中性粒细胞可能更倾向于形成网络。因此,喂食,一个重要的益生菌与保护和矫正活动对CD,有效地抑制NETosis (43)。
溃疡性结肠炎
类似于CD,加州大学的特点是胃肠道炎症。在一起,CD和加州大学临床实体形式称为炎症性肠病(IBD)。但是,与CD,加州大学主要是限于结肠炎症(189年,190年)。也类似于CD,加州大学临床炎症的细胞成分大多是未知的。意料之中的是,篮网已经观察到加州大学,与炎症相关的蛋白质组学研究191年),不过显然需要更多的细胞和生化研究清楚地了解净参与炎症性肠病。
代谢疾病和网
现在众所周知,代谢疾病相关的慢性低度炎症主要由激活先天免疫系统(192年)。代谢综合征(MS)的代谢紊乱特征(高血糖、高甘油三酯血症、血脂异常和高血压)也与免疫系统的激活(193年)。
多余的热量摄取,增加脂肪堆积,lipotoxicity激活效应分子(细胞因子)的生产,进而促进慢性低度炎症状态,诱发许多成熟的招聘和激活免疫细胞(包括肥大细胞、巨噬细胞、DCs和中性粒细胞)特别是在代谢组织和脂肪组织(194年)。
2型糖尿病
糖尿病(DM)的特点是慢性炎症,包括体液因素和不同类型的白细胞,包括单核和多形核白细胞。众所周知,糖尿病人,中性粒细胞计数和功能障碍的增加吞噬活动(195年)。
已经观察到糖尿病微环境可以支持NETosis,如糖尿病条件(高血糖)、中性粒细胞生成氧化应激和产生细胞因子il - 6和TNF-α等,这使中性粒细胞产生ETs(图6)(196年,197年)。然而,假设连接NETosis放松管制和高血糖,氧化应激,炎症和疾病并发症的仍有待进一步证实(198年)。
图6
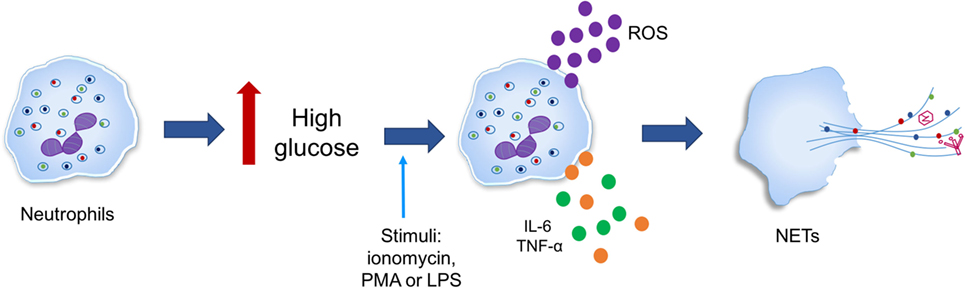
图6。高葡萄糖质数接受NETosis中性粒细胞。中性粒细胞在炎症刺激[ionomycin phorbol-12-myristate-13-acetate (PMA),或脂多糖(LPS)]产生氧化应激除了产生细胞因子白介素和肿瘤坏死因子α等(TNF-α)在2型糖尿病引发的高葡萄糖。
在人类和小鼠模型的研究表明高血糖容易使中性粒细胞释放网无论糖尿病类型与ionomycin刺激时,PMA或有限合伙人。此外,蛋白质的表达PAD4被发现在中性粒细胞增加四倍糖尿病患者与健康对照组相比,表明增加PAD4染色质decondensation可能忙。然而,目前尚不清楚是否高葡萄糖浓度上调的蛋白表达PAD4转录或翻译后水平(197年)。
在2型糖尿病患者的血清,NET-related生物标志物(弹性蛋白酶,mono -和oligonucleosomes dsDNA)增加相比,非糖尿病者;此外,这些生物标志物积极与糖化血红蛋白(HbA1c)水平。在这些患者中,dsDNA也一直与il - 6的浓度有关,这可能表明一个角色的NETosis高血糖和炎症之间的相互作用以及在炎症的后果(198年)。
Joshi等人研究了高血糖的条件是否能调节净释放。他们发现中性粒细胞暴露于高葡萄糖浓度和中性粒细胞分离出糖尿病患者改变潜在的净释放暴露在LPS刺激(196年)。因此,他们推测,在场的慢性炎性疾病期间高血糖促进本构网的形成,然而弱刺激反应。Fadini等人发现相似的结果当他们分析了网络一代通路在糖尿病溃疡患者中性粒细胞分离,即。,NOX-dependent NOX-independent通路。在这个模型中,中性粒细胞显示增强自发诱导NETosis NETosis但妥协能力(199年)。
另一方面,中性粒细胞从非糖尿病个体暴露在高葡萄糖浓度(25毫米)已被证明是更容易自发和PMA-induced NETosis相比,那些暴露于低葡萄糖浓度(5毫米)和甘露醇(25毫米)。non-energetic糖并不影响NETosis以来,这一现象可以解释为通过增加糖酵解增强活性氧的生产(198年)。
总之,这些数据表明,净形成增强高血糖的糖尿病类型和来源的条件独立。
重要的是要注意,在糖尿病患者中,伤口愈合慢代表的主要并发症之一。愈合是一个过程,包括内皮细胞、成纤维细胞、白细胞、血小板、角化细胞。由于炎症愈合过程的一个典型特征,中性粒细胞的第一个细胞招募了受伤的网站,他们也作为抗菌细胞。然而,一些研究报告,NETosis-inducing的过度激活中性粒细胞可能导致伤口愈合不良(199年- - - - - -202年)。
它已经表明,PAD4-deficient老鼠(包括糖尿病和非糖尿病)拥有更快的伤口愈合和re-epithelization过程比野生型(WT)同行,独立于伤口感染。这表明NETosis可能会阻碍伤口愈合通过限制角化细胞迁移,因此,足够的re-epithelization。因此,网是一个公认的治疗目标的一个主要禁用糖尿病并发症(197年)。
最近,它已被证明,过度的NET-related蛋白质与伤口愈合变化、低分辨率有关。此外,东北,低聚糖和单核小体,中性粒细胞gelatinase-associated lipocalin,和PR3增加活检与愈合不良预后相比在缓解或完全愈合(199年)。
这些研究显示中性粒细胞之间的联系,在糖尿病炎症和组织损伤。然而,需要更多的调查研究以完全理解NETosis背后的机制和葡萄糖代谢。
肥胖和女士
过多的肥胖是脂肪组织产生的后果损失的能量摄入和支出之间的平衡(203年)。肥胖的发展意味着复杂的相互作用的遗传和环境因素也经常与其他慢性并发症(高血糖、血脂异常、高甘油三酯血症和高血压)。患者三种标准可能是临床诊断为女士提供根据世界卫生组织。女士会增加患代谢疾病的风险,如2型糖尿病和心血管疾病(194年)。
肥胖和低度的慢性炎症有关在白色脂肪组织(192年)。TNF-α等发病脂肪细胞能够分泌il - 6,和引发,而由于其促炎属性,外围中性粒细胞与增强的相关活动,如生产超氧化物自由基和净形成(图7)。然而,炎症在肥胖的影响及其与NETosis不清楚(204年- - - - - -206年)。由于脂肪组织促进潜在neutrophil-activating促炎的环境中,存在一个需要研究是否肥胖的增加可能导致NETosis。此外,增强葡萄糖代谢可能导致mitochondrial-derived ROS的增量。这种现象也观察到在肥胖,引发炎症通路的激活(192年)。
图7
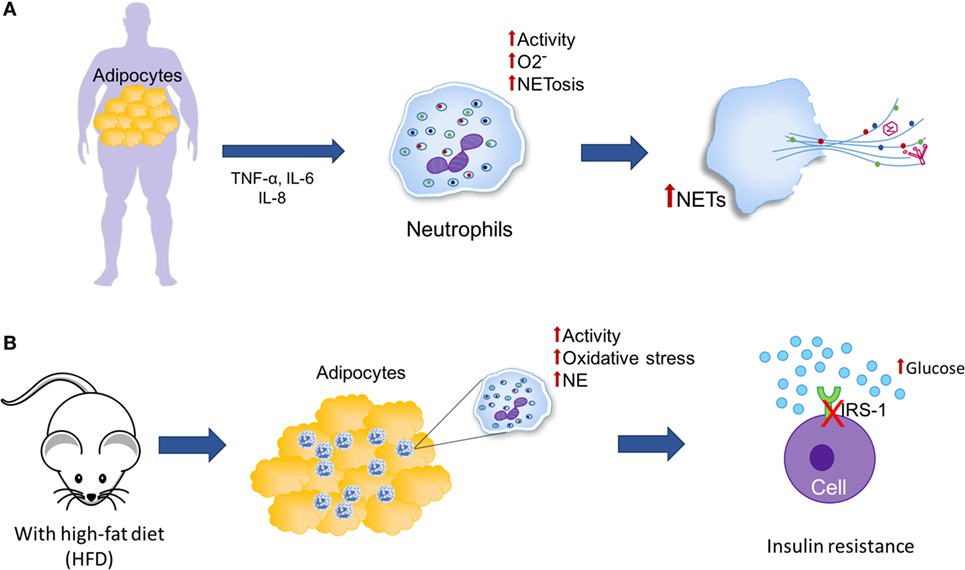
图7。中性粒细胞胞外陷阱(网)和脂肪组织。(一)肥胖的特点是脂肪组织的增加和慢性低度炎症发病中脂肪细胞分泌,如肿瘤坏死因子α(TNF-α)、il - 6,引发,与周边活动增加中性粒细胞(一代超氧化物和感应NETosis)。(B)在小鼠高脂饮食(HFD),中性粒细胞增加招聘观察脂肪组织和嗜中性粒细胞弹性蛋白酶(NE)活动,因此,有可能是中性粒细胞可以促进胰岛素抵抗通过胰岛素受体底物的降解(IRS-1)。
高脂肪饮食喂养老鼠(HFD)诱导中性粒细胞招募脂肪组织(图7B)。因此,它是可能的,中性粒细胞可以导致肥胖的炎症级联反应(207年)。因此,当老鼠HFD感染了流感病毒,病毒滴度增加三倍相比,感染低脂饮食的老鼠(最晚完成日期);H2O2HFD老鼠的浓度也相对较高,表明氧化应激增加肺部。这些老鼠的中性粒细胞相比更倾向于自发的净形成中性粒细胞来自最小的致命剂量的小鼠。最后,作者得出结论,因为促炎细胞因子水平升高,氧化应激引起的这些细胞因子,肥胖与更高的肺病毒滴度可以代表净形成更强的刺激,这表明在病态肥胖患者,肺网可以形成在显著水平应对流感感染,从而加重肺损伤,进一步导致influenza-provoked肺炎并发症(208年)。
Talukdar等人报道,中性粒细胞来自老鼠HFD拥有更高的细粒度的内容和他们的不活动显著高于一个最小的致命剂量的老鼠,这可能通过胰岛素受体底物的降解促进胰岛素抵抗(图7B)。他们也观察到胰岛素信号和降低高葡萄糖生产在人类肝细胞和小鼠脂肪细胞。此外,他们表明NE-knockout老鼠比WT老鼠对胰岛素更敏感,表明东北的消融会导致更高的肝胰岛素敏感性和减少促炎基因的表达(207年)。
氧化低密度脂蛋白(oxLDL)是一个复杂的混合物组成的低密度脂蛋白氧化生物活性元素内在促炎的活动能够刺激ROS生产和改善人类中性粒细胞的脱粒能力。此外,oxLDL能够诱导ROS-dependent NETosis在人类中性粒细胞剂量和时间的方式。有趣的是,TLR2 / TLR6似乎形成所需oxLDL-induced NETosis PKC-IRAK-MAPK通路,这表明NETosis是一个多因子的过程,不仅需要呼吸爆发还几个信号事件的顺序激活依赖于净电感的本质。总之,炎性环境特点是oxLDL和促炎细胞因子(TNF, IL-1β引发)可能被视为一个潜在的电感的网没有微生物的刺激,进一步加重全身炎症反应综合征、动脉粥样硬化和其他无菌炎症条件(209年)。
观点和结论
积累的数据在传染性中性粒细胞的作用,自身免疫,autoinflammatory和代谢疾病通过网络结构表明,它们构成小说的预后和代表候选人适合作为治疗靶点阻断或本地中和净信号的形成。
作者的贡献
VD-R网和LI-G关注微生物疾病相关。AA-N修订自身免疫性疾病和网之间的互动。MM-G写网和autoinflammatory障碍之间的关系。AG-O MF-M讨论了代谢紊乱和蚊帐。所有作者的贡献同样发展的这篇评论。
利益冲突声明
作者声明,这项研究是在没有进行任何商业或财务关系可能被视为一个潜在的利益冲突。
引用
3所示。Brinkmann V, Reichard U, Goosmann C, B的过错,Uhlemann Y,维斯DS, et al .中性粒细胞胞外陷阱杀死细菌。科学(2004)303:1532-5。doi: 10.1126 / science.1092385
4所示。武井H,荒木,渡边H, Ichinose,有些f .快速杀死人类中性粒细胞的有效活化剂佛波醇12-myristate 13-acetate (PMA)伴随着变化不同于典型的细胞凋亡或坏死。J Leukoc(1996)59:229-40。
6。曹JH,弗雷泽IP, Fukase K, Kusumoto年代,藤本Y,斯塔尔GL,等。人类肽聚糖识别蛋白S neutrophil-mediated先天免疫的效应。血(2005)106:2551-8。doi: 10.1182 / - 2005 - 02 - 0530血
7所示。Lominadze G,鲍威尔DW, Luerman GC,链接AJ,病房RA,麦克莱什KR。蛋白质组学分析人类嗜中性粒细胞的颗粒。摩尔细胞蛋白质组学(2005)4:1503-21。doi: 10.1074 / mcp.M500143-MCP200
9。城市CF, Ermert D,施密德M, Abu-Abed U, Goosmann C, Nacken W, et al .中性粒细胞胞外陷阱包含calprotectin,胞质蛋白复合物参与宿主防御白色念珠菌。公共科学图书馆Pathog(2009)5:e1000639。doi: 10.1371 / journal.ppat.1000639
10。霍尔沃森TW,威尔顿M, Poon KK,佩特里B, Lewenza s DNA是中性粒细胞胞外的抗菌成分陷阱。公共科学图书馆Pathog(2015)11:e1004593。doi: 10.1371 / journal.ppat.1004593
11。马尔卡希H, Charron-Mazenod L, Lewenza s细胞外DNA螯合物阳离子和诱发抗生素耐药性铜绿假单胞菌生物膜。公共科学图书馆Pathog(2008)4:e1000213。doi: 10.1371 / journal.ppat.1000213
12。李DY,黄厘米,Nakatsuji T, Thiboutot D,康SA Monestier M, et al .组蛋白H4是人力架构的抗菌作用的重要组成部分。J投资北京医学(2009)129:2489 - 96。doi: 10.1038 / jid.2009.106
13。陈王Y, Y,鑫L,贝弗利SM, Carlsen ED,波波夫V, et al。微分杀菌剂的人类蛋白质组蛋白H2A、H2B的影响利什曼虫promastigotes和无鞭毛体。感染Immun(2011)79:1124-33。doi: 10.1128 / IAI.00658-10
14。Pilsczek FH,盐沼D, Poon KK, Fahey C, Yipp BG, Sibley CD,等。一个新颖的机制快速核中性粒细胞胞外陷阱形成的反应金黄色葡萄球菌。J Immunol(2015)185:7413-25。doi: 10.4049 / jimmunol.1000675
15。Al-Khafaji AB Tohme年代,Yazdani HO米勒D,黄H, Tsung a .过氧化物引发嗜中性粒细胞胞外陷阱形成的地和NOX-dependent机制。摩尔地中海(2016)22:621-31。doi: 10.2119 / molmed.2016.00054
16。刘易斯高清,Liddle J,库特我,阿特金森SJ,巴克博士伯灵顿BD, et al。抑制PAD4活动足以扰乱老鼠和人类网络的形成。Nat化学杂志(2015)11:189 - 91。doi: 10.1038 / nchembio.1735
17所示。先生李,李米,林德伯格,Kennett MJ,熊N,王y PAD4是至关重要的抗菌先天免疫介导的中性粒细胞胞外陷阱。J Exp地中海(2010)207:1853 - 62。doi: 10.1084 / jem.20100239
18岁。麦茨勒Papayannopoulos V, KD, Hakkim Zychlinsky A .中性粒细胞弹性蛋白酶和髓过氧物酶调节中性粒细胞胞外陷阱的形成。J细胞(2010)191:677 - 91。doi: 10.1083 / jcb.201006052
19所示。Neeli我汗SN·拉迪奇m .组蛋白deimination作为响应在中性粒细胞炎症刺激。J Immunol(2008)180:1895 - 902。doi: 10.4049 / jimmunol.180.3.1895
20.Brinkmann V, Zychlinsky a有益自杀:为什么中性粒细胞死网。Nat Microbiol牧师(2007)5:577 - 82。doi: 10.1038 / nrmicro1710
22。Branitzki-Heinemann K, Mollerherm H, Vollger L, Husein DM,德布尔N, Blodkamp年代,et al .中性粒细胞胞外陷阱的形成在低氧水平。前面Immunol(2016)7:518。doi: 10.3389 / fimmu.2016.00518
23。Douda DN,汗马,Grasemann H, Palaniyar n SK3通道和线粒体ROS调解NADPH oxidase-independent NETosis引起钙流入。《美国国家科学院刊年代(2015)112:2817-22。doi: 10.1073 / pnas.1414055112
25。伯德,O ' brien XM,约翰逊厘米,拉维妮LM, Reichner JS。快速中性粒细胞胞外陷阱形成的一个细胞外依赖机制以应对白色念珠菌。J Immunol(2013)190:4136-48。doi: 10.4049 / jimmunol.1202671
26岁。克拉克SR,马AC, Tavener SA麦当劳B, Goodarzi Z,凯利MM, et al .血小板TLR4激活中性粒细胞胞外陷阱诱捕细菌感染性的血。Nat地中海(2007)13:463-9。doi: 10.1038 / nm1565
27。Yipp BG,佩特里B,盐沼D, Jenne CN,斯科特•BNV Zbytnuik LD, et al。动态NETosis是由生活在人类和小鼠细菌性脓肿和中性粒细胞严重感染的革兰氏阳性。Nat地中海(2012)18:1386 - 93。doi: 10.1038 / nm.2847
28。Carestia,考夫曼T, Rivadeneyra L, Landoni VI, Pozner RG, Negrotto年代,et al .介质和分子途径参与中性粒细胞胞外陷阱形成的监管由激活血小板。J Leukoc(2016)99:153 - 62。doi: 10.1189 / jlb.3a0415 - 161 r
29。Yousefi年代,Mihalache C, E,科兹洛夫斯基和施密德我,西蒙。可行的中性粒细胞释放线粒体DNA形成中性粒细胞胞外陷阱。细胞死亡是不同的(2009)16:1438-44。doi: 10.1038 / cdd.2009.96
30.Agraz-Cibrian JM,吉拉尔多DM,玛丽调频,Urcuqui-Inchima美国理解网的分子机制及其在抗病毒先天免疫中的作用。病毒Res(2017)228:124-33。doi: 10.1016 / j.virusres.2016.11.033
31日。Maugeri N,坎帕纳L, Gavina M, Covino C, De Metrio M, Panciroli C, et al .激活血小板高机动组框1到中性粒细胞,诱导自噬的挤压和促进中性粒细胞胞外陷阱。J Thromb Haemost(2014)12:2074 - 88。doi: 10.1111 / jth.12710
32。鲜明的K,腓立比V,施托克豪森年代,会先J,安东内利,米勒M,等。二硫化HMGB1来自小鼠血小板坐标静脉血栓形成。血(2016)128:2435-49。doi: 10.1182 / - 2016 - 04 - 710632血
33。Mori Y,山口M, Terao Y,岩漠年代,Ooshima T,川端康成,等人α-Enolase肺炎链球菌引起的中性粒细胞胞外陷阱的形成。J临床生物化学(2012)287:10472 - 81。doi: 10.1074 / jbc.M111.280321
34。坎巴人K, Mitroulis我Apostolidou E, A,吉伦特Chrysanthopoulou, Pneumatikos我,等。自噬调节的交付形成血栓的组织因子在人类中性粒细胞胞外陷阱败血症。《公共科学图书馆•综合》(2012)7:e45427。doi: 10.1371 / journal.pone.0045427
35。Marin-Esteban V, Turbica我,杜福尔G, Semiramoth N, Gleizes,峡谷R, et al。阿/博士广泛地坚持大肠杆菌应变C1845诱导中性粒细胞胞外陷阱,杀死细菌和人类enterocyte-like细胞损伤。感染Immun(2012)80:1891-9。doi: 10.1128 / IAI.00050-12
36。Pieterse E,洛特N, Yanginlar C, Hilbrands磅,van der Vlag j .中性粒细胞区分不同的细菌脂多糖来源和选择性地释放嗜中性粒细胞胞外陷阱。前面Immunol(2016)7:484。doi: 10.3389 / fimmu.2016.00484
38。Mollerherm H,诺伊曼,Schilcher K, Blodkamp年代,Zeitouni NE, Dersch P, et al。鼠疫enterocolitica介导的中性粒细胞胞外退化陷阱(网)。《列托人(2015)362:fnv192。doi: 10.1093 / femsle / fnv192
39岁。城市CF Gillenius E。胶蛋白侵袭素鼠疫的伪通过beta1整合蛋白诱导中性粒细胞胞外陷阱。微生物感染(2015)17:327-36。doi: 10.1016 / j.micinf.2014.12.014
40。Ramos-Kichik V, Mondragon-Flores R, Mondragon-Castelan M, Gonzalez-Pozos年代,Muniz-Hernandez年代,Rojas-Espinosa O, et al .中性粒细胞胞外陷阱所诱导结核分枝杆菌。肺结核(2009)89:29-37。doi: 10.1016 / j.tube.2008.09.009
41岁。Braian C, Hogea V, Stendahl O。结核分枝杆菌全身的嗜中性粒细胞胞外陷阱激活人类巨噬细胞。J先天Immun(2013)5:591 - 602。doi: 10.1159 / 000348676
42。9月,Hosseinzadeh, Gorkiewicz G, Lichtenegger年代,罗伊,莱特纳,博士等。霍乱弧菌躲避中性粒细胞胞外陷阱由两个细胞外核酸酶的活性。公共科学图书馆Pathog(2013)9:e1003614。doi: 10.1371 / journal.ppat.1003614
43。疯人L,洛伦兹RJ,集团Glogauer M,谢尔曼点。益生菌乳杆菌抑制中性粒细胞胞外陷阱的形成。J Immunol(2014)192:1870-7。doi: 10.4049 / jimmunol.1302286
44岁。特里帕西年代,胆量,金正日EJ,白色先生,鹿角KL。LL-37调节人类嗜中性粒细胞对甲型流感病毒的反应。J Leukoc(2014)96:931-8。doi: 10.1189 / jlb.4a1113 - 604 rr
45岁。Moreno-Altamirano MM, Rodriguez-Espinosa O, Rojas-Espinosa O, Pliego-Rivero B, Sanchez-Garcia陆地。登革病毒serotype-2干扰的形成中性粒细胞胞外陷阱。Intervirology(2015)58:250-9。doi: 10.1159 / 000440723
46岁。Saitoh T, Komano J, Saitoh Y,三泽T, Takahama M, Kozaki T, et al .中性粒细胞胞外陷阱调解人类免疫缺陷virus-1宿主防御反应。细胞宿主细菌(2012)12:109-16。doi: 10.1016 / j.chom.2012.05.015
47岁。丰沙尔GA, Jaeger N, Czepielewski RS,马查多,女士Muraro SP,斯坦RT, et al .呼吸道合胞体病毒融合蛋白促进TLR-4-dependent中性粒细胞胞外陷阱形成的人类中性粒细胞。《公共科学图书馆•综合》(2015)10:e0124082。doi: 10.1371 / journal.pone.0124082
48。比安奇M, Hakkim Brinkmann V,赛勒U,塞格尔RA, Zychlinsky,等。基因疗法恢复网络形成的CGD控制曲霉病。血(2009)114:2619-22。doi: 10.1182 / - 2009 - 05 - 221606血
49。布鲁斯,Kniemeyer O, Hasenberg M, Aimanianda V,尼采,Thywissen,等。生产的细胞外陷阱来自烟曲霉属真菌体外感染肺组织是依赖于入侵的中性粒细胞和hydrophobin又影响。公共科学图书馆Pathog(2010)6:e1000873。doi: 10.1371 / journal.ppat.1000873
50。罗查JD、Nascimento MT Decote-Ricardo D, Corte-Real年代,Morrot, Heise N, et al。荚膜多糖新型隐球菌调节生产中性粒细胞胞外陷阱(网)由人类中性粒细胞。Sci代表(2015)5:8008。doi: 10.1038 / srep08008
51。贝克VS, Imade通用电气、Molta NB Tawde P, Pam SD, Obadofin MO, et al . Cytokine-associated中性粒细胞胞外陷阱和抗核抗体恶性疟原虫六岁以下儿童感染。颧骨J(2008)说。doi: 10.1186 / 1475-2875-7-41
52岁。Waisberg M, Molina-Cruz Mizurini DM,基拉N,公元前苏萨马D, et al。恶性疟原虫感染诱发蚊子唾液蛋白的表达(Agaphelin)目标中性粒细胞功能和抑制血栓形成而不削弱止血。公共科学图书馆Pathog(2014)10:e1004338。doi: 10.1371 / journal.ppat.1004338
53岁。林Abi阿卜杜拉DS, C,球CJ,先生王,Duhamel通用电气、Denkers)等等。刚地弓形虫触发释放人类和小鼠中性粒细胞胞外陷阱。感染Immun(2012)80:768 - 77。doi: 10.1128 / IAI.05730-11
54。孔雀SJ de Silva我,罗伊FD。是什么决定了鼻运输金黄色葡萄球菌吗?趋势Microbiol(2001)9:605-10。doi: 10.1016 / s0966 - 842 x (01) 02254 - 5
57。冈萨雷斯DJ, Corriden R, Akong-Moore K,奥尔森J, Dorrestein PC, Nizet v . n端ArgD肽从古典文学金黄色葡萄球菌agr系统细胞毒性和促炎的活动。化学生物(2014)21:1457 - 62。doi: 10.1016 / j.chembiol.2014.09.015
58岁。小林Malachowa N, SD,弗里德曼B, Dorward DW, DeLeo FR。金黄色葡萄球菌白细胞毒素GH促进中性粒细胞胞外陷阱的形成。J Immunol(2014)191:6022-9。doi: 10.4049 / jimmunol.1301821
59。Thammavongsa V, Missiakas DM, Schneewind O。金黄色葡萄球菌降低中性粒细胞胞外陷阱,促进免疫细胞死亡。科学(2014)342:863-6。doi: 10.1126 / science.1242255
60。Kadioglu Weiser约,佩顿JC,安德鲁PW。的作用链球菌引起的肺炎主机呼吸道毒性因素殖民和疾病。Nat Microbiol牧师(2008)6:288 - 301。doi: 10.1038 / nrmicro1871
61年。拜特K, Wartha F, Albiger B, Normark年代,Zychlinsky, Henriques-Normark B一个核酸内切酶允许链球菌引起的肺炎中性粒细胞胞外的逃离陷阱。咕咕叫杂志(2006)16:401-7。doi: 10.1016 / j.cub.2006.01.056
62年。王焦Moorthy, Rai P, H, S,谭KB,秦L, et al .胶囊的致命的肺炎链球菌血清型增强中性粒细胞胞外陷阱的形成在体内肺炎的发病机理。Oncotarget(2016)7:19327-40。doi: 10.18632 / oncotarget.8451
63年。朱L,旷Z,威尔逊英航,刘GW。Competence-independent肺炎球菌的活性安达(纠正)介导DNA和细胞外网和退化对毒力很重要。《公共科学图书馆•综合》(2013)8:e70363。doi: 10.1371 / journal.pone.0070363
64年。Russo DM,低音出售JIF Gabelloni ML,小盖夫纳,佐丹奴M, Catalano M, et al .细胞外DNA:促炎的主要组成部分铜绿假单胞菌生物膜。J Immunol(2010)184:6386 - 95。doi: 10.4049 / jimmunol.0901640
65年。Moorthy, Narasaraju T, Rai P, Perumalsamy R,谭KB,王年代,et al .体内和体外研究中性粒细胞胞外陷阱的角色在初选后继发肺炎球菌肺炎肺感染流感。前面Immunol(2013)4:56。doi: 10.3389 / fimmu.2013.00056
66年。Kret我年轻的RL,马尔科姆KC,卡塞雷斯SM, Poch KR,尼科尔斯DP, et al。(净)介导的中性粒细胞胞外陷阱杀害铜绿假单胞菌:获得性耐药的证据在CF气道内,独立于雌性生殖道。《公共科学图书馆•综合》(2011)6:e23637。doi: 10.1371 / journal.pone.0023637
67年。Hoskins Dworski R,西蒙•胡,Yousefi嗜酸性粒细胞和嗜中性粒细胞胞外DNA陷阱在人类过敏哮喘气道。J过敏Immunol(2011)127:1260-6。doi: 10.1016 / j.jaci.2010.12.1103
68年。皮德森F, Marwitz年代,Holz O,柯尔斯顿,Bahmer T, Waschki B, et al。中性粒细胞胞外陷阱形成和细胞外DNA稳定的慢性阻塞性肺病患者的痰。和地中海(2015)109:1360-2。doi: 10.1016 / j.rmed.2015.08.008
71年。松原Daigo K,山口N,河村建夫T, K,江年代,大桥R, et al。蛋白质组学概要文件循环pentraxin 3 (PTX3)复杂的脓毒症与azurocidin 1演示了互动和中性粒细胞胞外的其他成分陷阱。摩尔细胞蛋白质组学(2012)11:M111.015073。doi: 10.1074 / mcp.M111.015073
72年。斯利EJ、马Matthay Wolters PJ。拐点在脓毒症生物学:从当地防御系统器官损伤。是杂志肺细胞摩尔杂志吗303年(2012年):l355 - 63。doi: 10.1152 / ajplung.00069.2012
73年。田中K,小池百合子Y,志村T, Okigami M, Ide年代,Toiyama Y, et al。体内的嗜中性粒细胞胞外各器官的陷阱小鼠脓毒症模型。《公共科学图书馆•综合》(2014)9:e111888。doi: 10.1371 / journal.pone.0111888
74年。Czaikoski PG,莫塔JM, Nascimento, Sonego F, Castanheira阵线,梅洛的PH值,et al .中性粒细胞胞外陷阱在实验和临床脓毒症诱导器官损伤。《公共科学图书馆•综合》(2016)11:e0148142。doi: 10.1371 / journal.pone.0148142
75年。Fuchs助教,布里尔,Duerschmied D, Schatzberg D, Monestier M,迈尔斯小DD, et al .细胞外DNA陷阱促进血栓形成。《美国国家科学院刊年代(2010)107:15880-5。doi: 10.1073 / pnas.1005743107
76年。Servin。发病机理的阿发/博士广泛地坚持大肠杆菌。中国Microbiol牧师(2005)18:264 - 92。doi: 10.1128 / cmr.18.2.264 - 292.2005
77年。伯格曼Chromek M, Slamova Z, P, Kovacs L, Podracka L, Ehren我,et al。抗菌肽抗菌肽对入侵保护泌尿道细菌感染。Nat地中海(2006)12:636-41。doi: 10.1038 / nm1407
78年。诺伊曼A Berends等观点,Molhoek EM,盖洛RL, Meerloo T, ET al。抗菌肽LL-37促进中性粒细胞胞外陷阱的形成。物化学J(2014)464:3-11。doi: 10.1042 / BJ20140778
79年。Garey千瓦,Sethi年代,Yadav Y,杜邦霍奇金淋巴瘤。荟萃分析来评估复发的危险因素艰难梭状芽胞杆菌感染。J Hosp感染(2008)70:298 - 304。doi: 10.1016 / j.jhin.2008.08.012
81年。版英格兰M, Straub写M, Durre P。艰难梭状芽胞杆菌是一个自养细菌病原体。《公共科学图书馆•综合》(2013)8:e62157。doi: 10.1371 / journal.pone.0062157
82年。Sorg JA Sonenshein。抑制的起始艰难梭状芽胞杆菌孢子萌发使用类似物的鹅去氧胆酸,胆汁酸。J Bacteriol(2010)192:4983 - 90。doi: 10.1128 / JB.00610-10
83年。Weingarden AR, Dosaπ,DeWinter E,引导CJ,肖尼西可,约翰逊JR, et al .结肠胆汁酸成分粪便微生物群移植后的变化足以控制艰难梭状芽胞杆菌发芽和成长。《公共科学图书馆•综合》(2016)11:e0147210。doi: 10.1371 / journal.pone.0147210
84年。特纳Nusrat,冯Eichel-Streiber C, JR, Verkade P,杰,马达腊镇Parkos CA。艰难梭状芽胞杆菌毒素破坏上皮屏障功能通过改变膜microdomain紧密连接蛋白的定位。感染Immun(2001)69:1329-36。doi: 10.1128 / iai.69.3.1329 - 1336.2001
85年。Zemljic M, Rupnik M,斯卡帕M, Anderluh G,巴鲁G, Castagliuolo即重复的领域艰难梭状芽胞杆菌毒素B展品对人类肠道上皮细胞细胞毒性影响和减少上皮屏障功能。厌氧生物(2010)16:527-32。doi: 10.1016 / j.anaerobe.2010.06.010
86年。Eilers B, Mayer-Scholl,沃克T,唐C, Weinrauch Y, Zychlinsky A .中性粒细胞抗菌蛋白增强弗氏志贺菌粘附和入侵。细胞Microbiol(2010)12:1134-43。doi: 10.1111 / j.1462-5822.2010.01459.x
87年。Phalipon, Sansonetti PJ。志贺氏杆菌的方法操纵宿主肠道先天和适应性免疫系统:一个工具盒为生存吗?Immunol细胞生物(2007)85:119-29。doi: 10.1038 / sj.icb7100025
88年。佩尔多莫坚称JJ, Gounon P Sansonetti PJ。多形核白细胞轮回促进结肠上皮细胞单层的入侵弗氏志贺菌。中国投资(1994)93:633-43。doi: 10.1172 / JCI117015
90年。伊巴拉JA Steele-Mortimer O。沙门氏菌——终极内幕。沙门氏菌毒力因子,调节细胞内生存。细胞Microbiol(2009)11:1579 - 86。doi: 10.1111 / j.1462-5822.2009.01368.x
91年。Pacello F,塞西P, Ammendola年代,Pasquali P, Chiancone E, a Battistoni周质的铜、锌超氧化物歧化酶和细胞质Dps赞成保护沙门氏菌血清从细胞外活性氧型沙门氏菌感染。Biochim Biophys学报(2008)1780:226-32。doi: 10.1016 / j.bbagen.2007.12.001
92年。郝林NA,弗洛姆,Kikhney J,李,如果电动机,Schulzke JD, et al。鼠疫enterocolitica影响结肠肠屏障功能。J感染说(2016)213:1157 - 62。doi: 10.1093 / infdis / jiv571
93年。转轮杰,Seo KS, intuition杰,Cundiff是的,Minnich SA Bohach GA,等。抗中性粒细胞鼠疫YopJ / P-induced免受ROS-mediated细胞死亡的凋亡和III型分泌系统。《公共科学图书馆•综合》(2010)5:e9279。doi: 10.1371 / journal.pone.0009279
94年。吉尔·O,迪亚兹,Vilaplana C, Tapia G,迪亚兹J, M堡et al .肉芽肿封装是一个关键因素包含minipigs肺结核感染。《公共科学图书馆•综合》(2010)5:e10030。doi: 10.1371 / journal.pone.0010030
95年。佩尔松丫,Blomgran-Julinder R,拉赫曼年代,郑L, Stendahl O。结核分枝杆菌全身的凋亡中性粒细胞巨噬细胞通过释放引发炎性反应的热休克蛋白72,代理协同作用的细菌。微生物感染(2008)10:233-40。doi: 10.1016 / j.micinf.2007.11.007
97年。Doron年代,Snydman博士,Gorbach SL。乳酸菌GG:细菌学和临床应用。杂志中国北部是(2005)34:483 - 98。doi: 10.1016 / j.gtc.2005.05.011
One hundred.缝边器,Teijaro JR Arandjelovic年代,Mowen KA。PAD4-mediated中性粒细胞胞外陷阱形成感染流感免疫力不是必需的。《公共科学图书馆•综合》(2011)6:e22043。doi: 10.1371 / journal.pone.0022043
101年。Hoeksema M,特里帕西年代,白人M, L,陶本伯杰J, van Eijk M, et al . Arginine-rich组蛋白有很强的A型流感病毒的抗病毒活性。先天Immun(2015)21:736-45。doi: 10.1177 / 1753425915593794
102年。古兹曼毫克,霍氏某人Artsob H, Buchy P,法勒J, Gubler DJ, et al。登革热:持续的全球威胁。Nat Microbiol牧师(2010)8:S7-16。doi: 10.1038 / nrmicro2460
103年。Rodriguez-Espinosa O, Rojas-Espinosa O, Moreno-Altamirano MM, Lopez-Villegas光电,Sanchez-Garcia陆地。中性粒细胞胞外代谢需求陷阱的形成。免疫学(2015)145:213-24。doi: 10.1111 / imm.12437
104年。卡明斯基R,陈Y,费舍尔T, Tedaldi E,那不勒斯,张Y, et al。消除hiv - 1从人类T-lymphoid细胞基因组CRISPR / Cas9基因编辑。Sci代表(2016)6:22555。doi: 10.1038 / srep22555
106年。Ogra PL呼吸道合胞体病毒:病毒、疾病和免疫应答。Paediatr和牧师(2004)5:S119-26。doi: 10.1016 / s1526 - 0542 (04) 90023 - 1
107年。常Borchers, C,格什温我,格什温LJ。呼吸道合胞体病毒——全面审查。中国过敏Immunol牧师(2013)45:331 - 79。doi: 10.1007 / s12016 - 013 - 8368 - 9
108年。Cortjens B, de Boer橙汁,德容R, Antonis房颤,Sabogal皮尼罗y,附近地区R, et al。中性粒细胞胞外陷阱引起气道阻塞在呼吸道合胞病毒疾病。中草药(2016)238:401-11。doi: 10.1002 / path.4660
110年。Netea MG, Joosten洛杉矶,范德梅尔JW, Kullberg BJ, van de Veerdonk FL。免疫防御假丝酵母真菌感染。Nat Immunol牧师(2015)15:630-42。doi: 10.1038 / nri3897
111年。城市CF Reichard U, V Brinkmann Zychlinsky a .中性粒细胞胞外陷阱捕捉并杀死白色念珠菌酵母和菌丝的形态。细胞Microbiol(2006)8:668 - 76。doi: 10.1111 / j.1462-5822.2005.00659.x
114年。李·乔丹,刘H,巴克BM, Snarr BD, Gravelat FN,艾尔·阿卜杜拉问,et Al。真菌胞外galactosaminogalactan介导的毒性增强抗中性粒细胞胞外陷阱。公共科学图书馆Pathog(2015)11:e1005187。doi: 10.1371 / journal.ppat.1005187
115年。罗姆M,格林MJ, D 'Auria AC, Almyroudis NG,西格尔BH、城市CF。NADPH氧化酶促进中性粒细胞胞外陷阱形成肺曲霉病。感染Immun(2014)82:1766 - 77。doi: 10.1128 / IAI.00096-14
116年。Bose我,瑞茜AJ, Ory JJ, Janbon G,林根TL。酵母掩护下:胶囊新型隐球菌。Eukaryot细胞(2003)2:655 - 63。doi: 10.1128 / ec.2.4.655 - 663.2003
117年。陈JA, Fowkes陆地,Beeson詹。表面抗原恶性疟原虫来华的红细胞免疫目标和候选疟疾疫苗。细胞摩尔生命科学(2014)71:3633-57。doi: 10.1007 / s00018 - 014 - 1614 - 3
118年。自助餐PA, Safeukui我Deplaine G, Brousse V, V Prendki,尹浩然,Thellier的发病机制恶性疟原虫在人类疟疾:从脾生理学的见解。血(2011)117:381 - 92。doi: 10.1182 / - 2010 - 04 - 202911血
121年。Girolomoni G Mrowietz U,保罗·c .牛皮癣:针对interleukin-17理由。Br北京医学(2012)167:717-24。doi: 10.1111 / j.1365-2133.2012.11099.x
123年。林,鲁宾CJ, Khandpur R,王司法院,Riblett M, Yalavarthi年代,et al。肥大细胞和中性粒细胞释放IL-17通过细胞外陷阱形成牛皮癣。J Immunol(2011)187:490 - 500。doi: 10.4049 / jimmunol.1100123
124年。朗德R,格雷戈里奥J, Facchinetti V, Chatterjee B,王YH,家的B, et al。血浆树突细胞感觉self-DNA加上抗菌肽。自然(2007)449:564-9。doi: 10.1038 / nature06116
125年。比德尔曼T, Kneilling M, Mailhammer R,麦尔K,砂光机,Kollias G, et al。肥大细胞控制中性粒细胞招募在T细胞介导的dth反应通过肿瘤坏死因子和巨噬细胞炎性蛋白2。J Exp地中海(2000)192:1441-52。doi: 10.1084 / jem.192.10.1441
126年。黄李L, L, Vergis AL, H, Bajwa, Narayan V, et AL . IL-17由中性粒细胞调节IFN-gamma-mediated中性粒细胞迁移在小鼠肾脏缺血再灌注损伤。中国投资(2010)120:331-42。doi: 10.1172 / JCI38702
127年。梁SC,谭XY, Luxenberg DP,卡里姆R, Dunussi-Joannopoulos K,柯林斯M, et al .白介素(IL) -22和IL-17 coexpressed Th17细胞和合作提高抗菌肽的表达。J Exp地中海(2006)203:2271-9。doi: 10.1084 / jem.20061308
128年。斯佩里奇M, Koglin年代,Kim SM Morizane年代,Besch R,普林茨JC, et al . IL-17A提高维生素D3-induced抗菌肽抗菌肽的表达在人类角质细胞。J Immunol(2008)181:8504-12。doi: 10.4049 / jimmunol.181.12.8504
129年。Schulze穆尼奥斯勒,Janko C, C, Schorn C, sart K, Schett G, et al .自身免疫和慢性inflammation-two clearance-related步骤系统性红斑狼疮的发病机理。Autoimmun牧师(2010)10:38-42。doi: 10.1016 / j.autrev.2010.08.015
130年。赫曼Perniok A、F·韦德金德,M,斑点C,施耐德M高水平的循环早期apoptic外周血单核细胞在系统性红斑狼疮。红斑狼疮(1998)7:113-8。doi: 10.1191 / 096120398678919804
131年。朗德R, Ganguly D, Facchinetti V, Frasca L,康拉德C,格雷戈里奥J, et al .中性粒细胞激活血浆树突细胞通过释放self-DNA-peptide复合物在系统性红斑狼疮。Sci Transl地中海(2011)3:73ra19。doi: 10.1126 / scitranslmed.3001180
132年。史密斯CK,卡普兰乔丹。中性粒细胞的作用在系统性红斑狼疮的发病机制。当今Rheumatol(2015)27:448-53。doi: 10.1097 / BOR.0000000000000197
133年。布兰科P, Palucka AK,吉尔M,帕斯卡V, Banchereau IFN-alpha j .诱导树突状细胞分化的系统性红斑狼疮。科学(2001)294:1540-3。doi: 10.1126 / science.1064890
134年。赫曼Mahajan A, M,穆尼奥斯勒。间隙不足和细胞死亡通路:系统性红斑狼疮的发病机制的模型。前面Immunol(2016)35。doi: 10.3389 / fimmu.2016.00035
135年。鲍曼,Kolowos W,沃尔再保险公司经理B, Gaipl U, Neuhuber西城,et al。受损的凋亡细胞易染体巨噬细胞在生发中心的系统性红斑狼疮患者。关节炎感冒(2002)46:191 - 201。doi: 10.1002 / 1529 - 0131 (200201) 46:1 < 191:: AID-ART10027 > 3.0.CO; 2 k
136年。Banchereau J,帕斯卡v . I型干扰素在系统性红斑狼疮等自身免疫性疾病。免疫力(2006)25:383 - 92。doi: 10.1016 / j.immuni.2006.08.010
137年。茧丝U, Magnusson M, Eloranta马丁,Perers, Alm问,Ronnblom l . FcγRIIa表达对自然IFN-α-producing细胞(血浆树突细胞)和所需IFN-α生产引起的凋亡细胞结合狼疮免疫球蛋白。J Immunol(2003)171:3296 - 302。doi: 10.4049 / jimmunol.171.6.3296
138年。哈亚希意味着TK,莱兹E, F, Murali先生Golenbock DT,光泽广告。人类狼疮autoantibody-DNA复合物激活CD32 DCs通过合作和TLR9识别。中国投资(2005)115:407-17。doi: 10.1172 / JCI23025
139年。Garcia-Romo GS Caielli年代,织女星B, Connolly J, Allantaz F,徐Z, et al .网中性粒细胞是I型干扰素生产的主要诱发儿童系统性红斑狼疮。Sci Transl地中海(2011)3:73ra20。doi: 10.1126 / scitranslmed.3001201
140年。Dubois B, C Massacrier Vanbervliet B,菲也特J, Briere F, Banchereau J, et al .关键作用il - 12的树突cell-induced天真的B淋巴细胞的分化。J Immunol(1998)161:2223-31。
141年。哈里斯他Andersson U, Pisetsky DS。HMGB1:多功能alarmin推动自身免疫和炎性疾病。Nat Rheumatol牧师(2012)8:195 - 202。doi: 10.1038 / nrrheum.2011.222
142年。Abdulahad哒,Westra J,林堡PC, Kallenberg CG,除m . HMGB1在系统性红斑狼疮:它在皮肤病变发展中的作用。Autoimmun牧师(2010)9:661-5。doi: 10.1016 / j.autrev.2010.05.015
143年。Schellekens GA,维瑟H,德容英航,van den Hoogen FH,阴霾JM, Breedveld FC,等。类风湿性关节炎的诊断特性识别循环citrullinated肽的抗体。关节炎感冒(2000)43:155 - 63。doi: 10.1002 / 1529 - 0131 (200001) 43:1 < 155:: AID-ANR20 > 3.0.CO; 2 - 3
144年。Khandpur R, Carmona-Rivera C, Vivekanandan-Giri Gizinski, Yalavarthi年代,骑士JS, et al。网的citrullinated自身抗原,刺激类风湿性关节炎的炎症反应。Sci Transl地中海(2013)5:178ra40。doi: 10.1126 / scitranslmed.3005580
145年。Lee CY Raza K, Scheel-Toellner D, D抗起球,科诺SJ,法尔恰尼F, et al。滑液白细胞凋亡抑制早期类风湿性关节炎患者。关节炎Res其他(2006)8:R120。doi: 10.1186 / ar2009
146年。Foulquier C, Sebbag M, Clavel C, Chapuy-Regaud年代,艾尔Badine R, Mechin MC,等。2型(ipad 2)和肽基精氨酸deiminase PAD-4但不是PAD-1,附加三,PAD-6表达在类风湿性关节炎滑膜组织炎症紧密结合。关节炎感冒(2007)56:3541-53。doi: 10.1002 / art.22983
147年。琼斯我,堤道CP, Knuckley B, Slack-Noyes杰,汤普森公关。蛋白质精氨酸deiminase 4 (PAD4):当前的理解和未来的治疗潜力。当今药物越是加大重击(2009)12:616-27。
148年。黄Assi路,SH,路德维希·拉扎·K,戈登C,尹浩然,鲑鱼。肿瘤坏死因子α激活释放的B淋巴细胞刺激中性粒细胞浸润风湿性关节。关节炎感冒(2007)56:1776 - 86。doi: 10.1002 / art.22697
149年。Chakravarti Raquil MA, Tessier P Poubelle PE。表面RANKL toll样受体4-stimulated人类中性粒细胞激活监测骨吸收。血(2009)114:1633-44。doi: 10.1182 / - 2008 - 09 - 178301血
150年。张X,山田R,铃木,泽田师傅T,吉野年代,Tokuhiro年代,et al .本地化peptidylarginine deiminase 4 (PADI4)和citrullinated蛋白质类风湿性关节炎滑膜组织。风湿病学(2005)44:40-50。doi: 10.1093 /风湿病学/ keh414
151年。田中岩漠T, Torikai M, Kuwazuru, M, Horai N,福田T, et al .细胞外高机动组框染色体蛋白1是一个耦合的缺氧和炎症因子关节炎。关节炎感冒(2008)58:2675 - 85。doi: 10.1002 / art.23729
152年。Evankovich J,曹西南,张R,红衣主教J, Dhupar R, L,等。高机动组框1释放肝细胞在缺血和再灌注损伤是由组蛋白脱乙酰酶活性下降。J临床生物化学(2010)285:39888 - 97。doi: 10.1074 / jbc.M110.128348
153年。已经受理N,阿帕德海耶J, Neeli我,汗,Pattanaik D,迈尔斯L, et al .毡状的综合征自身抗体结合deiminated染色质组蛋白和中性粒细胞胞外陷阱。关节炎感冒(2012)64:982 - 92。doi: 10.1002 / art.33432
154年。阿特金森,Eisenbarth GS,米歇尔啊。1型糖尿病。《柳叶刀》(2014)383:69 - 82。doi: 10.1016 / s0140 - 6736 (13) 60591 - 7
155年。Gan MJ, Albanese-O奥尼尔,Haller乔丹。1型糖尿病:当前的概念在流行病学、病理生理学、临床护理和研究。咕咕叫Probl Pediatr Adolesc卫生保健(2012)42:269 - 91。doi: 10.1016 / j.cppeds.2012.07.002
157年。范美女TL, Coppieters KT,冯Herrath毫克。1型糖尿病:病因学、免疫学和治疗策略。杂志牧师(2011)91:79 - 118。doi: 10.1152 / physrev.00003.2010
158年。帕吉特勒,Broniowska KA,汉森PA, Corbett杰,谢霆锋嗯。活性氧的作用,促炎细胞因子在1型糖尿病的发病机理。安N Y科学(2013)1281:16-35。doi: 10.1111 / j.1749-6632.2012.06826.x
159年。Foulis AK, Farquharson妈,微薄的a .免疫反应性的α干扰素分泌β细胞的1型糖尿病。《柳叶刀》(1987)2:1423-7。doi: 10.1016 / s0140 - 6736 (87) 91128 - 7
160年。Harsunen MH粉扑R D 'Orlando O, Giannopoulou E, Lachmann L, Beyerlein, et al .降低血液白细胞和中性粒细胞数量在1型糖尿病的发病机制。霍恩金属底座Res(2013)45:467 - 70。doi: 10.1055 / s - 0032 - 1331226
161年。山谷,Giamporcaro通用,Scavini M, Stabilini,格罗根P, Bianconi E, et al。减少循环中性粒细胞之前和伴随1型糖尿病。糖尿病(2013)62:2072-7。doi: 10.2337 / db12 - 1345
162年。肖王Y, Y,钟L D,张J,你Y, et al。增加中性粒细胞弹性蛋白酶和蛋白酶3和增强NETosis与β细胞自身免疫密切相关的1型糖尿病患者。糖尿病(2014)63:4239-48。doi: 10.2337 / db14 - 0480
163年。Beaudoin戴安娜J,西摩尼Y,法里奥L, L, Agerberth B, F Barrat, et al .中性粒细胞之间的串扰,B-1a细胞和血浆树突细胞启动自身免疫糖尿病。Nat地中海(2013)19:65 - 73。doi: 10.1038 / nm.3042
164年。格雷科FA, Vendrame F, Spagnuolo我Dotta F .先天免疫和1型糖尿病的发病机制。Semin Immunopathol(2011)33:57 - 66。doi: 10.1007 / s00281 - 010 - 0206 - z
165年。坎巴人K, Chrysanthopoulou Vassilopoulos D, Apostolidou E, Skendros P,吉伦特,等。组织因子表达在中性粒细胞胞外陷阱和中性粒细胞衍生微粒antineutrophil胞质抗体相关血管炎可以促进thromboinflammation和thrombophilic状态与疾病有关。安大黄说(2014)73:1854 - 63。doi: 10.1136 / annrheumdis - 2013 - 203430
167年。Kessenbrock K, Krumbholz M Schonermarck U, W,西城总值Werb Z, et al .网中性粒细胞在自身免疫性小血管血管炎。Nat地中海(2009)15:623-5。doi: 10.1038 / nm.1959
168年。Nakazawa D,师大H Tomaru U,吉田M,西年代,Atsumi T, et al。增强形成和无序的监管在myeloperoxidase-ANCA-associated微观polyangiitis网。J是Soc Nephrol(2014)25:990-7。doi: 10.1681 / ASN.2013060606
169年。Saffarzadeh M, Juenemann C、马Queisser Lochnit G, Barreto G, Galuska SP, et al .中性粒细胞胞外陷阱直接诱导上皮和内皮细胞死亡:组蛋白的主要作用。《公共科学图书馆•综合》(2012)7:e32366。doi: 10.1371 / journal.pone.0032366
171年。施赖伯,小H, Jennette JC,施耐德W,勒夫特FC, Kettritz r . C5a受体介导中性粒细胞激活和ANCA-induced肾小球肾炎。J是Soc Nephrol(2009)20:289 - 98。doi: 10.1681 / ASN.2008050497
172年。实物地租R,报告》,Brandes R, Ziebermayr R,坎宁安D, Alderson CA等。分子拟态pauci-immune焦坏死性肾小球肾炎。Nat地中海(2008)14:1088 - 96。doi: 10.1038 / nm.1874
173年。Pendergraft WF三世,普雷斯顿GA、沙RR Tropsha,卡特连续波小,Jennette JC, et al .自身免疫是由cPR-3(105 - 201),对人类自身抗原的蛋白质互补proteinase-3。Nat地中海(2004)10:72-9。doi: 10.1038 / nm968
175年。索德伯格D, Kurz T, Motamedi Hellmark T,埃里克森P, Segelmark m .水平的提高中性粒细胞胞外陷阱残余小血管血管炎患者的循环,而是一个负相关在缓解anti-neutrophil胞质抗体。风湿病学(2015)54:2085 - 94。doi: 10.1093 /风湿病学/ kev217
177年。格雷森PC, Carmona-Rivera C,徐L, Lim N,高Z,巨人,et AL . Neutrophil-related基因表达和低密度粒细胞ANCA-associated血管炎疾病活动和对治疗的反应。关节炎Rheumatol(2015)67:1922-32。doi: 10.1002 / art.39153
178年。古普塔AK, Joshi MB, Philippova M,白尾海雕P, Hasler P,哈恩年代,et al。激活内皮细胞诱导中性粒细胞胞外陷阱和容易NETosis-mediated细胞死亡。2月列托人(2010)584:3193-7。doi: 10.1016 / j.febslet.2010.06.006
179年。王黄YM,王H, C,陈M,赵MH,促进血凝过快antineutrophil细胞质antibody-associated血管炎C5a-induced组织factor-expressing微粒子和嗜中性粒细胞胞外陷阱。关节炎Rheumatol(2015)67:2780 - 90。doi: 10.1002 / art.39239
180年。冯Bruhl ML,鲜明的K,斯坦哈特,Chandraratne年代,康拉德,我洛伦茨M, et al .单核细胞,中性粒细胞和血小板合作启动和传播在老鼠体内静脉血栓形成。J Exp地中海(2012)209:819-35。doi: 10.1084 / jem.20112322
181年。Surmiak M, Hubalewska-Mazgaj M, Wawrzycka-Adamczyk K, Szczeklik W, Musial J, Brzozowski T, et al。Neutrophil-related和血清生物标志物与polyangiitis支持细胞外陷阱肉芽肿病疾病的机制。中国Exp Rheumatol34 (2016):s98 - 104。
182年。Schauer Maueroder C, D Kienhofer,哈恩J, C,经理B, Schett G, et al。中性粒细胞胞外陷阱编排痛风的局部免疫反应。J摩尔(2015)93:727-34。doi: 10.1007 / s00109 - 015 - 1295 - x
183年。Mitroulis我坎巴人K Ritis K .中性粒细胞IL-1beta,和痛风:有链接吗?Semin Immunopathol(2013)35:501-12。doi: 10.1007 / s00281 - 013 - 0361 - 0
184年。Mitroulis我坎巴人K, Chrysanthopoulou Skendros P, Apostolidou E, Kourtzelis我,et al .中性粒细胞胞外陷阱形成痛风与IL-1beta和autophagy-related相关信号。《公共科学图书馆•综合》(2011)6:e29318。doi: 10.1371 / journal.pone.0029318
185年。Schauer C, C之前,穆尼奥斯勒,赵Y, Kienhofer D,弗雷B, et al。聚合中性粒细胞胞外陷阱限制降低炎症细胞因子和趋化因子。Nat地中海(2014)20:511-7。doi: 10.1038 / nm.3547
186年。Schauer Schett G、C,霍夫曼M,赫尔曼·M·为什么痛风发作停止?痛风的免疫发病机理的路线图。限制型心肌病开放(2015)1:e000046。doi: 10.1136 / rmdopen - 2015 - 000046
188年。Somasundaram R, Nuij VJ, van der Woude CJ, Kuipers EJ, Peppelenbosch MP, Fuhler通用。外围中性粒细胞功能和细胞信号在克罗恩病。《公共科学图书馆•综合》(2013)8:e84521。doi: 10.1371 / journal.pone.0084521
189年。福伊尔斯坦JD, Cheifetz。溃疡性结肠炎:流行病学,诊断和管理。梅奥中国Proc(2014)89:1553 - 63。doi: 10.1016 / j.mayocp.2014.07.002
190年。Kawalec p和炎症性肠病的间接成本:克罗恩病和溃疡性结肠炎。系统回顾。地中海拱形Sci(2016)12:295 - 302。doi: 10.5114 / aoms.2016.59254
191年。Bennike结核病,Carlsen TG, Ellingsen T, Bonderup好,Glerup H, Bogsted M, et al .中性粒细胞胞外陷阱在溃疡性结肠炎:蛋白质组分析肠活检。Inflamm肠说(2015)21:2052 - 67。doi: 10.1097 / MIB.0000000000000460
193年。格雷戈尔MF, Hotamisligil GS。炎症机制肥胖。为Immunol(2011)29:415-45。doi: 10.1146 / annurev - immunol - 031210 - 101322
194年。Boulange CL,七巧板,Chilloux J,尼科尔森JK,杜马斯我。肠道微生物群对炎症的影响,肥胖和代谢疾病。基因组医学(2016)42。doi: 10.1186 / s13073 - 016 - 0303 - 2
196年。Joshi MB,小伙子,Bharath Prasad, Balakrishnan,拉马L, Satyamoorthy k .高葡萄糖调节il - 6通过阻碍嗜中性粒细胞介导免疫内稳态细胞外陷阱的形成。2月列托人(2013)587:2241-6。doi: 10.1016 / j.febslet.2013.05.053
197年。黄SL, Demers M,适应K,勇敢的M,王Y, Goldfine AB, et al .糖尿病质数中性粒细胞进行NETosis,影响伤口愈合。Nat地中海(2015)21:815-9。doi: 10.1038 / nm.3887
198年。Menegazzo L, Ciciliot年代,Poncina N, Mazzucato M, Persano M, Bonora B, et al。NETosis引起高葡萄糖和与2型糖尿病有关。Acta Diabetol(2015)52:497 - 503。doi: 10.1007 / s00592 - 014 - 0676 - x
199年。Fadini GP, Menegazzo L Rigato M, Scattolini V, Poncina N, Bruttocao, et al . NETosis延迟糖尿病伤口愈合在小鼠和人类。糖尿病(2016)65:1061 - 71。doi: 10.2337 / db15 - 0863
203年。谁。肥胖和超重。字幕新闻。(2016)。可以从:http://www.who.int/mediacentre/factsheets/fs311/en/
204年。Brotfain E, N,哈达Shapira Y, Avinoah E,兹特尼克,Raichel L, et al .中性粒细胞功能在病态肥胖受试者。中国Exp Immunol(2015)181:156 - 63。doi: 10.1111 / cei.12631
205年。Keshari RS, Dubey Jyoti A, M, Kothari N,克里米,Bogra J, et al。细胞因子诱导中性粒细胞胞外陷阱形成:含义的炎性疾病状态。《公共科学图书馆•综合》(2012)7:e48111。doi: 10.1371 / journal.pone.0048111
206年。Trellakis年代,Rydleuskaya,费舍尔C, Canbay, Tagay年代,Scherag, et al。低脂联素,高水平的细胞凋亡,增加外周血中性粒细胞活动健康肥胖受试者。ob事实(2012)5:305-18。doi: 10.1159 / 000339452
207年。李Talukdar年代,哦,DY, Bandyopadhyay G, D,徐J,麦克尼利J, et al .中性粒细胞调节胰岛素抵抗在通过弹性蛋白酶分泌高脂肪饮食的老鼠。Nat地中海(2012)18:1407-12。doi: 10.1038 / nm.2885
208年。王Moorthy,谭KB,年代,Narasaraju T, Chow VT。高脂肪饮食对肺中性粒细胞胞外陷阱的形成在BALB / c小鼠流感肺炎。前面Immunol(2016)7:289。doi: 10.3389 / fimmu.2016.00289
关键词:网、传染病、autoinmmune疾病,autoinflammatory疾病,代谢疾病
引用:马Delgado-Rizo V, Martinez-Guzman Iniguez-Gutierrez L, Garcia-Orozco, Alvarado-Navarro和Fafutis-Morris M(2017)中性粒细胞炎症细胞外陷阱及其影响:概述。前面。Immunol。8:81。doi: 10.3389 / fimmu.2017.00081
收到:2016年11月14日;接受:2017年1月17日;
发表:2017年2月06
编辑:
安娜Rubartelli,Azienda Ospedaliera大学联盟圣马蒂诺是意大利版权:©2017 Delgado-Rizo、Martinez-Guzman Iniguez-Gutierrez、Garcia-Orozco Alvarado-Navarro Fafutis-Morris。这是一个开放分布式根据文章知识共享归属许可(CC)。使用、分发或复制在其他论坛是允许的,提供了原始作者(年代)或许可方认为,最初发表在这个期刊引用,按照公认的学术实践。没有使用、分发或复制是不符合这些条件的允许。
*通信:玛丽Fafutis-Morris,mfafutis@gmail.com