
黏膜表面分泌IgA的多面功能
- 瑞士洛桑州立大学医院免疫和过敏科研发实验室
分泌IgA (SIgA)在肠道、呼吸和泌尿生殖黏膜上皮的保护和稳态调节中发挥重要作用,将外界环境与体内隔离开来。SIgA的主要功能被称为免疫排斥,这一过程限制了大量微生物和粘膜抗原进入这些薄而脆弱的粘膜屏障。SIgA已被证明参与避免机会致病菌进入系统隔室传播,并严格控制共生生物与宿主之间存在的必要共生关系。因此,蠕动清除似乎是SIgA在粘膜表面发挥功能的众多机制之一。通过微折叠(M)细胞对抗原-SIgA复合物进行采样,与Peyer 's patch树突状细胞(DC)发生亲密接触,炎症过程的下调,上皮细胞和DC反应性的调节是最近发现的一些过程,这些过程强调了SIgA的贡献。这篇综述旨在介绍,重点在生化水平,SIgA的分子复杂性如何服务于这些多重和非冗余的作用模式。
介绍
分泌IgA (SIgA)是人类和许多其他哺乳动物粘膜表面的主要免疫球蛋白(Ig)。在全球范围内,产生的IgA比所有其他Ig同型的总和还要多。由于其特殊的生物合成途径依赖于固有层浆细胞的产生和覆盖在粘膜表面的上皮细胞介导的多Ig受体(pIgR)分泌,与在循环和组织中发现的IgA抗体相比,SIgA显示出非常不同的分子形式。SIgA在不断变化的环境中工作,其功能是将需要保持无菌的身体内部与富含抗原刺激的外部世界物理隔离,包括空气、液体和食物中的抗原刺激。在胃肠道中,保护宿主的SIgA面临的进一步挑战是区分共生无害的共生细菌和周期性入侵的、可能危及生命的微生物。所涉及的机制的复杂性还远远没有被完全理解。从更全面的免疫监测角度来看,包括SIgA在内的粘膜免疫系统必须不断监测环境,并在对正常微生物群的耐受性和对微生物病原体的免疫之间保持平衡,而系统免疫系统则被设计成对任何外来抗原或微生物产生强烈反应。鉴于肠道和气道黏膜屏障由单层上皮细胞所保证的固有脆弱性质,SIgA在维持稳态方面的贡献似乎是必不可少的。这反映在越来越多的证据表明,从生命早期开始,母乳SIgA在上皮成熟过程中的作用。然而,值得一提的是,由pIgR主动跨上皮转运的聚合IgM(就像聚合IgA一样),以及从血浆进入局部分泌物的IgG,也可以参与保护肠道和呼吸道(1- - - - - -4).
当讨论结构-功能关系时,这将变得很明显,抗体的各种分子形式是高度糖基化的,包括每个组成多肽中的糖源性残基。关于pIgA的糖基化,人类的IgA1和IgA2亚类在每条重链上都有两个保守的n -聚糖位点。此外,IgA2优先在分泌物中发现,在Cα1结构域上有一个或两个额外的n -聚糖。IgA1是唯一一个在铰链区含有o -碳水化合物的亚类。小鼠有一类IgA,在多肽组装和糖基化方面与人类IgA2结构相似。与单体血清IgA相比,在SIgA中发现的其他生化特征包括连接(J)链和分泌成分(SC) (5),是一种由前体pIgR的细胞外部分组成的多肽,可在上皮细胞间运输聚合IgA,这一过程也称为胞吞作用(6)(图1J链与两个IgA单体共价结合后,在黏膜分泌IgA的浆细胞的生物合成过程中触发二聚化(可能,但不常见的是,高量级的低聚化),这些粘膜分泌IgA的浆细胞大量存在于上皮下层的固有层中。J链只有一个n部分,是SIgA中糖基化最少的肽组成部分。在聚合IgA(和IgM五聚体)中加入J链对于膜结合的pIgR或从初乳或重组来源纯化的游离SC选择性识别两种抗体同型至关重要。碳水化合物残基占SC分子质量的20%,并鉴定出7个n -糖基化位点(7).SC在SIgA中的功能是多方面的(见下文),这可能解释了为什么它在确保单胞吞作用后,与来自上皮细胞合成的前体pIgR的聚合IgA一起释放。
图1
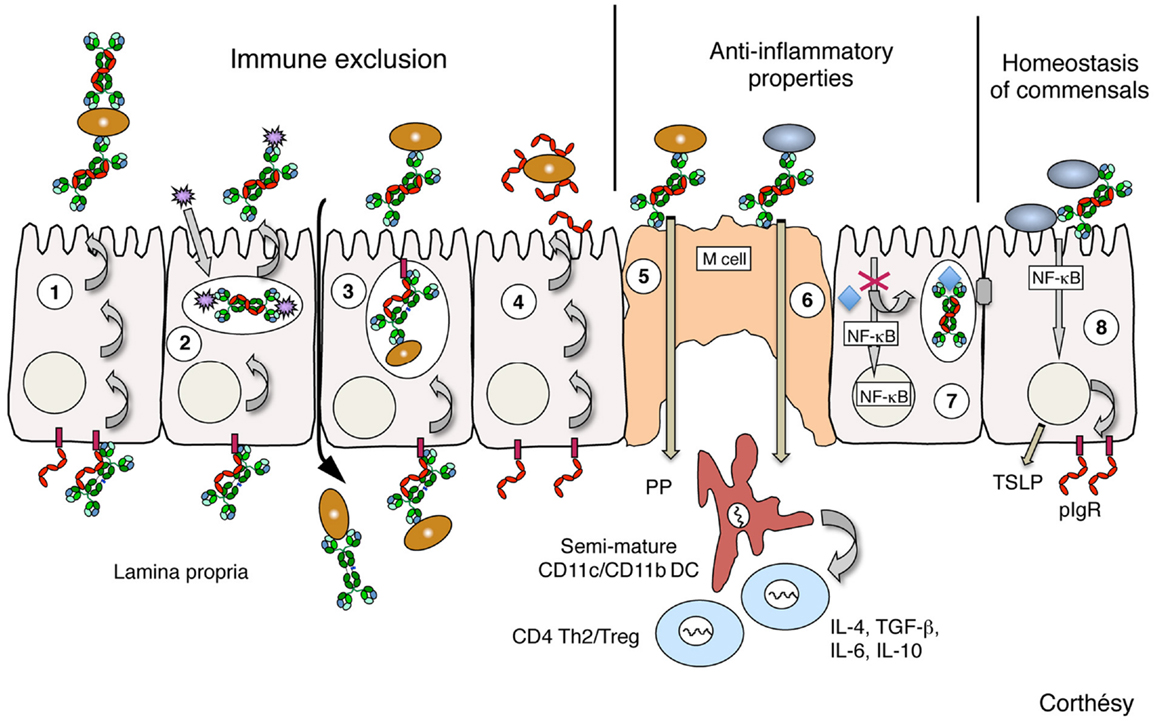
图1所示。所确定的聚合IgA、SIgA或SC可能有助于保护粘膜表面的水平的示意图表示在活的有机体内而且在体外模型.(1)固有层局部浆细胞产生的聚合IgA通过聚合Ig受体(pIgR)跨上皮细胞转运(这一过程称为胞吞作用),并通过与环境抗原(细菌、病毒、毒素等)相互作用,以SIgA的形式在腔内分泌物中释放,进行免疫排斥。(2)聚合IgA在进入pigr介导的分泌物过程中可以在细胞内拦截进入的病毒,并以非毒力免疫复合物的形式将病毒排出体外。(3)聚合IgA可在固有层中和侵入炎症上皮缺口的感染因子;随后pIgR的转运将有利于清除免疫复合物。(4)分泌物释放的游离SC通过其表面丰富的聚糖来中和病原体源性产物,并有助于保护上皮表面;当SC在SIgA中与聚合IgA结合时,这种性质是保守的。(5) M细胞在Peyer 's patches (PP)中采样SIgA,导致抗体特异性靶向上皮下穹丘区树突状细胞(DC)。在与有害抗原的免疫复合物的形式下,向PP中的naïve T细胞呈递和排泄的肠系膜淋巴结(未提取)导致减弱的th2偏置粘膜免疫反应的发生,并伴有炎症回路的猝灭。(6)值得注意的是,在共生菌中也实现了同样的siga介导的逆向转运,导致粘膜免疫系统通过诱导调节性T细胞形成非炎症性、耐受性模式。 (7) Neutralization of Gram-negative bacterial lipopolysaccharide (LPS) in apical recycling endosomes by transcytosing polymeric IgA abrogates NF-κB-mediated activation of pro-inflammatory gene products, thus preserving the epithelial barrier’s integrity. (8) Cross-talk between the probiotic bacteria and the intestinal mucosa is enhanced by SIgA, with various consequences extending from increased expression of epithelial pIgR and tight junction proteins to production of thymic stromal lymphopoietin (TSLP) involved in priming of mucosal DCs. Brown ellipses depict pathogen bacteria; gray ellipses depict commensal bacteria; purple spiky spheres depict virus; polymeric IgA are drawn in green; Free secretory component and polymeric Ig receptor (pIgR) are drawn in red; TSLP, thymic stromal lymphopoietin.
与SIgA功能相关的保护作用机制
免疫排斥是SIgA阻止微生物和毒素附着在粘膜靶上皮细胞上的主要机制,从而防止表面损伤、定植和随后的大规模入侵(8).在肠道环境中,免疫排斥被定义为SIgA通过识别病毒和细菌表面的多个抗原表位以及蛋白质,在肠腔中交联这些不同抗原的能力,从而延迟或取消它们粘附和/或穿透上皮的内在潜力(图)1SIgA对细菌、病毒和寄生粘膜病原体,以及毒素和可能的食物过敏原的这种共识作用模式,已通过动物模型的令人信服的证据得到了定义。在体外模型和人类流行病学研究。
在人类中,IgA已被用于粘膜表面的被动保护或治疗干预(9- - - - - -17),但取得了不同程度的成功,这可能是因为没有使用完整的SIgA分子。在小鼠的肠道中,被动口服特定的IgA抗体也可以防止细菌感染,包括鼠伤寒沙门氏菌(18,19),霍乱弧菌(20.),弗氏志贺菌(21),幽门螺杆菌(22).针对呼吸道合胞病毒的单克隆IgA抗体被动应用于小鼠鼻咽粘膜,随后预防了初始感染和肺炎(23).类似的观察结果表明,被动注入IgA在预防流感病毒感染中的关键作用(24)和呼肠孤病毒(25,26).静脉注射类似的病毒中和剂量的抗流感聚合IgA单抗,但不包括单体IgA,可保护小鼠免受病毒进入鼻腔分泌物而感染(27).通过植入小鼠背部(背包技术)的IgA分泌杂杂瘤克隆产生的抗原特异性IgA抗体被证明能提供有效的保护霍乱弧菌(28)和轮状病毒(29)在pigr介导的转运进入分泌物后。这些单克隆抗体的研究表明,使用IgA作为唯一的免疫剂可以保护免疫幼稚的动物。虽然在这些不同的实验环境中观察到的保护水平一般都很好,但重要的是要记住,在自然条件下,黏膜免疫反应将是多克隆的,因此更有效。在这方面,被动给予富含特异性和非特异性SIgA的初乳已被证明可以预防胃肠道和气道感染(30.,31).为了支持这些强调特定特异性SIgA保护功能的大量研究,无法产生IgA、J链或pIgR的转基因小鼠都表现出对抗粘膜感染因子的能力不足(32- - - - - -36).
利用极化单层生长的上皮细胞系来模拟肠道和气道中的粘膜屏障,已被证明是一种有价值的工具,可以证明IgA/SIgA对病原体和毒素的免疫排斥特性在体外.在这种模型中,抗体通过阻断霍乱毒素的结合起作用(37),梭状芽孢杆菌毒素A (38)及蓖麻毒素(39),从而防止随后的损害,包括液体流失,细胞毒性和中毒暴露的上皮细胞。干扰上皮细胞的附着和阻断HIV从上皮细胞向作为病毒靶点的外周血单个核细胞的传播被证实是HIV gp120特异性IgA的有效作用机制(40).IgA同型抗体在这一过程中的重要意义被进一步证明,从血清阳性患者中分离出的SIgA,而不是IgG,可防止艾滋病毒进入(41,42).沿着同一条线,粘连肠致病性大肠杆菌能靶向上皮细胞的菌株可被SIgA抑制(43).当测试时,聚合IgA在维持细胞完整性方面比特异性的单体IgA或IgG有系统的优势,这表明与聚合抗体相关的最高亲和性在中和过程中很重要,可能是通过有利于凝集(38,44).
这样一个在体外模型进一步显示了其价值,强调了经细胞转运的SIgA在细胞内中和入侵的流感病毒,Sendaï或轮状病毒的能力(45- - - - - -48)(图1在到达根尖表面的过程中,特异性聚合IgA抗体与病毒血凝素、神经氨酸酶或表面病毒蛋白在根尖循环内小体内共定位,从而阻止细胞内复制或组装,最终导致上清液和细胞裂解液中的病毒滴度降低。通过极化上皮细胞单分子层的HIV分离株的顶端到基底外侧转胞作用表明,HIV的传播被针对糖蛋白(gp)41包膜蛋白的聚合IgA阻断,从而排除了病毒向固有层的传播(49).至于其他病毒,细胞内中和发生在顶端循环核内体内,基于siga的免疫复合物选择性地循环到极化单分子层的顶端管腔状表面。细胞内用转胞作用IgA中和针对HIV gp120,但不包括具有相同Fv结构域的IgG,伴随着上皮细胞内病毒复制的抑制(50).有趣的是,中和作用依赖于基底侧血清样室中添加的聚合IgA的浓度,并达到与人体分泌物中SIgA含量相对应的平台,即约100 μg/ml (51).SIgA的排泄功能似乎是促进粘膜表面微生物消除的另一种可能的机制:当可溶性聚合iga基免疫复合物被添加到表达pIgR的极化上皮细胞单层的基底外侧隔室时,复合物被完整地运输到根尖侧(52).进一步证实了固有层中存在的聚合IgA捕获抗原,随后肠隐窝细胞分泌大量基底侧pIgR在活的有机体内(53)(图1,途径3)。
从这些不同的作用模式,我们可以得出结论,多级siga介导的保护发挥互补功能,以(1)在粘膜表面建立屏障,(2)在上皮细胞内消除,或(3)使有害微生物远离身体的内部隔间。虽然没有在这篇综述中讨论,但我们必须记住,涉及系统性IgA和fc α r1的细胞伙伴的各种其他备份机制是可用的,以确保有效的保护对抗无数致病性抗原(54,55).
在人类中,对感染的抵抗力和高特异性SIgA滴度之间的相关性在一些处理免疫的研究中被描述霍乱弧菌感染。lps特异性SIgA的存在(通过ELISA检测粪便中的抗体确定),以及ELISA检测抗体分泌细胞,允许建立SIgA与感染水平降低之间的强关联(56,57).在接种疫苗的小鼠中,也记录了强烈的粘膜IgA反应与对流感病毒的保护之间的相关性(58).患有IgA缺乏症(IgAD)的人出现上呼吸道和胃肠道感染的频率增加(59,60),但后果并不总是深刻的,主要是因为代偿性适应或先天机制,如SIgM取代SIgA (61).此外,由于缺乏IgA很少是绝对的,而且可能伴有其他同型的缺乏,因此评估IgA的真正贡献是复杂的。某些主要组织相容性复合体单倍型的相关性(62)和跨膜激活子和钙调节亲环素配体相互作用子的突变(63)在IgAD患者中可能进一步导致混淆,当涉及到IgA在预防胃肠道疾病中的直接和独特作用时(64).在这方面,iga敲除小鼠似乎在其他同型的表达和免疫反应缺陷方面表现出相同的改变(65,66),因此很难得出明确的结论。
SC在SIgA抗体中功能的多个方面
游离SC由5个类似于ig的结构域折叠成紧密的椭球,内部由几个二硫键桥稳定。总的来说,该分子显示一个j型,所有七个糖基化位点暴露在同一表面,远离聚合IgA的结合位点(67).在SIgA中SC与聚合IgA特异而稳定的相互作用基本涉及到所有结构域,结构域1是聚合IgA的原始锚定位点,结构域2和结构域3是空间约束结构域5,以确保聚合IgA中一个单体的一个Cα2结构域形成高产的二硫键桥(68,69).对人SIgA1和SIgA2亚类的三维分析显示,SC紧密包裹着聚合IgA的Cα2和Cα3结构域,这一特征可能对抗体的显著稳定性至关重要(70).当暴露于聚合IgA的肠道蛋白酶导致快速降解为Fab和F(ab’)2片段时,结合SC存在时,Cα2和Cα3结构域内的切割位点被掩盖,导致酶切延迟近24小时(71,72).交叉保护发生,因为结合SC不受影响,相反,自由SC迅速和完全降解,其程度与对照IgG抗体相似。在与各种大小和性质的抗原结合时,SIgA的稳定性也会增加:等级显示,在与细菌相互作用后,对蛋白酶的最佳保护,然后与病毒相互作用,最后与蛋白质毒素相互作用(73).这些内在的特性使得SIgA能够在肠道中普遍存在的恶劣环境中生存下来,并实现其保护功能。
结合SC在SIgA中的另一个特征是它能够通过7个表面暴露的n -链寡糖赋予抗体Fc片段的亲水性,这些寡糖配备了末端唾液酸残基。据认为,这种模式对与粘液的相互作用很重要,因此,在接近粘膜表面的适当位置放置抗体应该起到保护作用。基于siga的免疫复合物被拴在覆盖上皮细胞的黏液层内,进一步限制了在腔内环境中的扩散,并通过蠕动帮助它们从肠道中清除(74).SIgA在黏液中的锚定可能解释了观察到的黏液外层是结肠中微生物群的优先栖息地(75).然而,在Ussing腔培养的肠上皮细胞(IECs)顶端表面,CD71作为SIgA受体的鉴定(76),以及SIgA通过SC结合共生细菌(77)表明在肠道的这部分发生了更动态的情况。在小鼠肺部感染模型中弗氏志贺菌,粘液介导的SIgA锚定被发现有助于保证细菌的中和,防止进入组织(78);聚合IgA主要存在于鼻腔和支气管的管腔中,对小鼠的保护效果不佳。类似地,去除SIgA分子内SC的碳水化合物链,取消了黏液的锚定和相关的保护功能(79).因此,SC可以被视为SIgA的重要组成部分,因为它确保了持续的稳定性和在粘膜环境中的适当定位,这两个特征有助于抗体的最佳功能。
在SC中,n-聚糖约占蛋白质分子量的20%,并赋予SIgA更多的细菌凝集素样粘连蛋白的结合位点,除了四个Fab结构域(80).例如,SC直接与的表面蛋白相互作用链球菌引起的肺炎,胆碱结合蛋白A (CbpA),一种参与大鼠鼻咽定殖的细菌因子(81).结合依赖于SC结构域3和4的氨基酸序列(82)和CbpA中高度保守的六肽基序(83).有人提出这样的联系将有助于排除与上皮细胞的接触,然而一份使用未包被的报告肺炎链球菌菌株Rx6可促进表达pIgR的Detroit 562细胞的侵袭(84).游离SC在防御粘膜病原体方面的先天性质在病例中得到进一步证明艰难梭状芽胞杆菌毒素A和肠致病性大肠杆菌intimin (85)(图1,通路4);游离SC或非特异性SIgA对细菌产物的中和作用通过与SC表面显示的唾液和半乳糖残基相互作用阻止了目标上皮细胞的感染。游离SC多肽支架的变性使糖部分的构象不再能够与细菌表位相互作用,这表明精细调整的空间分布对复杂碳水化合物的特异性识别很重要。
SC和SIgA的糖基化性质在发挥其保护功能方面的重要性可以进一步说明,例如在唾液寡糖防止上皮粘连的情况下大肠杆菌通过I型菌丛凝集素(86,87).研究了人初乳SIgA中SC抑制细胞黏附的作用幽门螺杆菌对人胃表面黏液细胞的影响(88).SIgA中的碳水化合物侧链是蓖麻毒素的停靠点;人SIgA对蓖麻毒素没有fab依赖的特异性,减少了培养中上皮细胞系顶端表面的附着,并通过SC和IgA重链附着在人肠绒毛的管腔表面(89).虽然不是一个普遍的机制,这些许多例子确定自由和结合SC作为微生物清道夫,有助于抗致病武器库,保护身体上皮表面。
SIgA对感染性细菌适应度的干扰作用
可能通过粘膜微生物的凝集而阻断与上皮细胞的相互作用可能不是SIgA发挥保护功能的唯一机制。最近的证据表明,对细菌的生存能力或致病性有更直接的影响,例如通过扰动生物能量机制,影响运动,破坏细菌进入时的毒力因子(90).例如,在存在亚凝集量的IgA特异性的脂多糖o抗原(Sal4单抗),能力鼠伤寒沙门氏菌侵入上皮细胞的单分子层减少了20倍(91).为了支持这一观察结果,来自同一IgA的Fab片段虽然不能触发凝集,但与整个抗体分子一样有效地阻止了进入。此外,用Sal4处理导致细菌在15分钟内完全瘫痪,同样独立于凝集(91).这些数据与iga介导的对运动和进入的干扰解释了Sal4在的保护功能的观点是一致的沙门氏菌入侵。进一步的研究表明,Sal4处理损害了t3ss介导的转位子的形成,并减弱了标记效应蛋白向靶上皮细胞的传递(92).Sal4 IgA与细胞表面超微结构改变、外膜通透性改变、膜能量部分降低和细胞内ATP水平均被检测到沙门氏菌,这一系列特征可以使这种细菌无毒。这是通过YeaJ触发环二聚鸟苷单磷酸依赖信号通路发生的,YeaJ是一种被提出的内膜定位的双胍酸环化酶和一种已知的纤维素生物合成调节因子。对于细菌而言,由于产生胞外多糖和形成生物膜而导致失去活力(93).从抗体的角度来看,IgA具有转化的能力美国沙门氏菌感染通过对几种代谢途径的直接影响,从侵入性,运动状态到非运动,无毒状态。类似的抑制机制发生在小鼠单克隆IgA (IgAC5)与o -抗原的结合弗氏志贺菌血清型5a (94):细菌进入IECs所必需的T3SS的短暂损伤(45-60分钟)与细菌膜电位的部分降低和细胞内ATP水平的降低相一致。
SIgA在控制上皮转运中的作用
SIgA在粘膜表面的功能的延伸是免疫排斥的重要性,以保护宿主免受来自环境大分子的过度抗原挑战。IgAD患者ige介导的特应性疾病增加了过敏原通过粘膜的渗透和循环免疫复合物的形成(95,96)最初提出SIgA在控制食物抗原的吸收和降低特应性过敏易感性方面有作用。在气道过敏小鼠模型上进行的实验支持这一发现,抗原特异性SIgA抑制与炎症和哮喘相关的特征(97- - - - - -One hundred.).IgA在这一过程中的重要性在肠道中得到了进一步的证明,研究发现,与主动耐受相同蛋白质的小鼠相比,牛乳球蛋白致敏的小鼠在Peyer 's斑块中产生IgA的细胞频率要低得多,而且粪便SIgA也减少了(101).在过敏性哮喘小鼠模型中,唾液抗原特异性SIgA的产生持续增强,其中舌下疫苗接种引发了对后续挑战的保护(102).然而,在耐受成功的动物中,过敏原特异性SIgA并不总是增加,甚至可能在致敏动物中大量存在而不给予保护(103).在缺乏SIgA的pIgR敲除小鼠中可诱导口服耐受性,通过代偿性Treg功能确保对全身超敏反应的保护(104).这一系列在过敏和炎症疾病中相互矛盾的结果增加了关于SIgA在这些有害过程中的保护作用的持续辩论。此外,最近的临床研究还不清楚SIgA对过敏性疾病的重要性。IgAD患者在4岁时食物过敏的风险增加(105),而在另一个队列中,IgAD与食物过敏没有任何相关性(106).需要进一步的研究来阐明SIgA在维持局部耐受性和最终肠屏障完整性方面的重要性。
除了在免疫排斥中发挥重要作用外,与IgM和IgG相比,SIgA表现出选择性粘附在小鼠和人类Peyer 's斑块上的M细胞顶端膜上的显著能力(107,108).随后通过上皮细胞的有限运输导致在M细胞袋和基底层延伸的过程中存在少量的SIgA (109).到目前为止,还没有发现一种M细胞特异性受体确保SIgA的受控反转胞作用,尽管人们可以推测它需要表现出特定的特性(低表达,在共受体存在时的结合活性,识别改变的SIgA分子形式),以避免大量过量的SIgA进入肠腔。在活的有机体内摄取SIgA进入小鼠含有Peyer 's贴片的结扎回肠袢导致特异性靶向,并在上皮下圆顶区被树突状细胞(DC)内化(110).体外只有CD11c+CD11b+从Peyer斑块和引流肠系膜淋巴结分离出的DC表现出选择性结合和内化在活的有机体内情况(111)(图1有趣的是,在黏膜组织中,这种DC是IL-12的不良生产者,但却是分泌IL-10的T细胞的有效诱导剂(112)和从naïve B细胞产生IgA (113).DC- sign最近被确定为小鼠SIgA识别的可能候选人(114),而CD89和CD71(转铁蛋白受体)的存在已在成熟的人类DC (115).为了支持这些互补机制,最近有研究描述了IgA通过抑制IL-12产生来调节DC功能(116).
这些观察结果导致了一个明显的问题,即SIgA分子在M细胞中的转运及其随后与DC的关联的免疫学相关性。在有粘膜辅助霍乱毒素的情况下口服(117),基因工程SIgA携带外来表位弗氏志贺菌入侵素B触发唾液和全身针对细菌抗原的抗体的产生(118).为了进一步评估SIgA重新进入肠道黏膜后的粘膜免疫反应的性质,在没有任何佐剂的情况下,用由小鼠聚合IgA和人SC组成的异源SIgA口服免疫小鼠。工程SIgA触发人sc特异性抗体的产生和混合Th1/Th2型反应,保存或诱导IL-10和TGF-β在MLN中的表达,以及DC沿Peyer 's patch-MLN-脾轴的迁移和成熟(119)(图1通过与霍乱毒素佐剂的人SC进行比较,发现SIgA在有利于保持胃肠道局部稳态的非炎症环境中诱导低程度的激活。中和的弗氏志贺菌导致促炎回路的局部抑制,导致肠道组织损伤,这一特征是由于免疫复合物在恶劣的肠道环境中的稳定性(120)(图1,通路7)。
在以siga为基础的免疫复合物的背景下,一个有趣的可能性是,后者有助于局部免疫调节,或在生命早期,教育粘膜免疫系统朝着耐受的方向发展。为了支持这一点,牛奶抗体,特别是SIgA,可以阻止新生儿对共生细菌的反应(121).在这方面,及时提供一套适合新生儿肠道菌群的母体抗体,主要由母亲移交(至少在“经典的”阴道分娩之后),可以证明这种调节机制是合理的。有理由推测,母乳SIgA抗体通过上皮细胞将相关抗原直接导向DC,并在初始暴露于非自身抗原结构时在防御或耐受方面塑造胃肠免疫系统。基于SIgA重新进入Peyer的补丁的证据,对数据的广泛解释表明,在没有全球感染的情况下,SIgA涂层的中和细菌可以启动整个种群中naïve个体的免疫系统。
SIgA在调节微生物群中的作用
最近,SIgA已被确定为维持由微生物群组成的三位一体、胃肠道内衬的iec和底层粘膜免疫系统之间脆弱平衡的必要伙伴。发生在肠道粘膜表面的稳态控制对于保持数十亿定殖至关重要,第一眼看到潜在有害微生物是为了确保与宿主的最佳共生关系。事实上,任何潜在的功能障碍都可能导致疾病的发展,如炎症性肠病(122),或影响能量的提取过程,以及无法获得的营养物质的消化过程,例如碳水化合物的最终降解。共生细菌与肠道相关淋巴组织的正常发育直接相关,如孤立的淋巴滤泡(123)或分泌正常水平的SIgA (124),特异性未知,称为“天然”SIgA (125).考虑到黏膜表面遇到的抗原种类繁多,IgA的储备似乎被限制在最低限度(126),认为肠分泌物中存在多特异性、低亲和力抗体(127,128).最近的一项研究对这一概念提出了挑战,该研究使用高通量测序来调查IgA库的形成(129).分析超过一百万VH序列显示,IgA库包括高度扩增和低频克隆,这两者都有助于高度多样性,这一现象随着高突变引起的衰老而放大。与小鼠IgA序列相似,人类VH序列也携带大量体细胞高突变(130).这一过程是否依赖于多个Peyer 's斑块生发中心的再利用,如最近发现的(131)需要进一步调查。程序性细胞死亡蛋白1敲除小鼠的Peyer 's patch Treg细胞数量增加,其SIgA的结合能力发生变化,这反过来影响共生细菌的性质(132).人类和小鼠粪便中的共生细菌被SIgA自然包裹的事实强烈表明,这种相互作用对于维持稳态共生定殖是必要的。小鼠表达活化诱导的胞苷脱氨酶低形态(破坏体细胞高突变,但仍支持类开关重组)显示其微生物群组成的变化(133).总之,这表明SIgA利用fab依赖的适应性和甘聚糖介导的先天免疫相互作用来阻止微生物群。
利用游离SC和非特异性SIgA(从杂交瘤细胞系和初乳中纯化)替代天然粘膜抗体,得到了SIgA与肠道寄居菌相互作用的分子基础,即:乳酸菌,双歧杆菌,大肠杆菌,拟杆菌被鉴定为SC携带的许多聚糖残基(77).虽然与革兰氏阳性细菌的相互作用表明了碳水化合物在这一过程中的重要作用,但无论蛋白质伙伴使用何种分子形式,与革兰氏阴性细菌的结合都被保留了下来,这表明涉及不同的结合基序。革兰氏阳性菌与对照IgG之间的低相关性或无相关性证实了SC中糖部分在整个SIgA蛋白选择性结合高度多样化的微生物群中的关键作用。
IECs对共生细菌的识别已被认为在黏膜稳态中发挥基本作用,例如促进细胞因子释放、细胞扩张和增强屏障完整性(134- - - - - -136).此外,由SIgA包被的共生菌株可以增强重建的IEC单层的响应性在体外(137)(图1出乎意料的是,与SIgA的结合增加了IECs顶端表面的细菌锚定,通过增加紧密连接蛋白的磷酸化来促进细胞间接触,从而增强了屏障的完整性。此外,IECs促炎细胞因子/趋化因子的分泌被抑制,而pIgR的表达被促进。由于pIgR参与了从IECs基底外侧到顶端的SIgA转胞作用,因此可以得出结论,与SIgA复合的共生细菌对pIgR的表达产生了正反馈,导致更多的受体可用于活性SIgA转胞作用。这一现象可以解释先前观察到的由共生定殖引起的持续SIgA分泌(138).这有助于进一步确定SIgA在通过结合iec适当中和和适当传感的微妙平衡来保持共生细菌的功能。转铁蛋白受体(CD71)是否能够在IECs的顶端极点与SIgA结合(76)参与调控siga共生细菌复合物的结合仍有待确定。在生命早期,母亲SIgA的作用可能被认为是在限制新生儿肠道主要定植引起的潜在炎症的原始重要性。SIgA的存在有助于对新植入的微生物群的初始感知,并允许免疫系统在非炎症条件下正常发育。这种机制可能与理解炎症性肠病有关,其中,炎症性肠病与对肠道细菌的炎症反应失调有关(139).
虽然上面报道的数据阐明了SIgA在iec对共生体的粘膜监测中的作用,但它们并没有说明与潜在免疫系统伙伴的通信是如何建立的。细菌摄取有限,包括a乳酸菌和一个拟杆菌通过肠Peyer 's斑块中发现的M细胞采样发生,并且与非特异性SIgA相关可促进调节进入(140)(图1在几乎检测不到肠道SIgA的无菌小鼠中观察到的几乎不存在经上皮通道,可以通过给药预先形成的SIgA细菌复合物来补偿。单独给药的共生菌在3小时内被内源性SIgA包被,这强烈表明在稳态条件下随时都能发生结合,从而参与将绝大多数细菌留在肠腔内。SIgA在塑造肠道微生物群落组成中的作用可能源于其抑制某些细菌表位表达的能力(141),因此更倾向于一个物种或属的适应性。选择性siga介导的细菌靶向作用仅限于耐受性CD11c+CD11b+CD8−DC亚群和巨噬细胞位于Peyer斑块的上皮下圆顶区,表明宿主并非不知道其常驻共生细胞(140).在共生细菌被包被后,天然和/或特异性SIgA在很大程度上维持了微生物群的腔室化,同时偶尔允许罕见的易位事件,以控制宿主免疫系统与其共生生物之间的持续对话。在上皮下圆顶区,与局部DC相关的共生细菌不能穿透肠系膜引流淋巴结,导致对黏膜微生物群的免疫诱导受到限制(142,143).在这个阶段,使系统免疫系统对这些生物体相对无知,将允许在败血症的情况下获得足够的刺激。虽然SIgA单独或与蛋白质抗原或细菌联合通过Peyer 's patch M细胞的转运已经很好地建立起来,但其他经上皮途径(例如孤立的淋巴滤泡中的M细胞)是否仍有待确定。144)、固有层DC浮管状树突穿过致密上皮(145)、Peyer’s patch DC在M细胞周围延伸树突(146),或杯状细胞传代(147)可以解释调节肠道免疫反应的细胞的选择性采样和靶向。
结论
在外部世界和身体内部之间的界面的粘膜表面是潜在感染因子、共生细菌和外来蛋白质持续挑战的主要部位。这些脆弱上皮细胞的完整性和选择功能的维持意味着严格控制的内稳态在任何时候都得到保证。因此,根据刺激的性质,必须适当地激活非常不同的免疫调节机制。SIgA是该网络中的一个重要效应因子,在宿主粘膜环境与无害居民或有害病原体/抗原的适当感知之间的基本通信中起着至关重要的作用(图1).为了实现这一要求苛刻的多任务功能,SIgA显示了几种特性,从经典的免疫排斥和微生物群的永久检查,到通过与微生物、包括肠细胞和M细胞在内的上皮细胞以及粘膜相关淋巴组织中的DC的复杂接触进行局部免疫调节。必须强调的是,与SIgA相关的生化特征,如在侵袭性介质中的稳定性,在粘液中的固定,重度糖基化,抗原的fab独立识别,肠上皮的转胞作用和反转胞作用,都有助于使抗体在粘膜表面的特定环境中发挥最佳性能。
利益冲突声明
作者声明,这项研究是在没有任何商业或财务关系的情况下进行的,这些关系可能被解释为潜在的利益冲突。
致谢
Blaise Corthésy实验室由瑞士科学研究基金会资助,资助号为3100-138422。我们深深感谢Gilles Bioley博士对手稿的全面和透彻的阅读。
参考文献
1.Cardinale F, Friman V, Carlsson B, Björkander J, Armenio L, Hanson LA。IgA缺乏症患者对微生物和食物抗原的血清IgM和IgG抗体滴度和亲和力的畸变。扫描J免疫(1992)36: 279 - 83。doi: 10.1111 / j.1365-3083.1992.tb03100.x
2.费雷罗,李志强,李志强。胃内局部免疫球蛋白G抗体可能有助于免疫幽门螺杆菌小鼠感染。胃肠病学(1997)113: 185 - 94。doi: 10.1016 / s0016 - 5085 (97) 70094 - 5
3.Giannasca PJ,张志祥,雷卫东,Boden JA, Giel MA, Monath TP,等。血清抗毒素抗体介导全身和黏膜保护艰难梭状芽胞杆菌仓鼠的疾病。感染Immun(1999)67: 527 - 38。
4.Neal LM, McCarthy EA, Morris CR, Mantis NJ。在没有分泌IgA的情况下,疫苗诱导的蓖麻毒素肠道免疫疫苗(2011)29: 681 - 9。doi: 10.1016 / j.vaccine.2010.11.030
6.Kaetzel CS。聚合免疫球蛋白受体:在粘膜表面连接先天和适应性免疫反应。Immunol牧师(2005)206: 83 - 99。doi: 10.1111 / j.0105-2896.2005.00278.x
7.Hughes GJ, Reason AJ, Savoy L, Jaton J, Frutiger-Hughes S.人分泌成分中的碳水化合物部分。生物化学生物物理学报(1999)1434: 86 - 93。doi: 10.1016 / s0167 - 4838 (99) 00168 - 5
8.Mestecky J, Russell MW, Elson CO.肠道IgA:其在最大粘膜表面防御功能的新观点。肠道(1999)44: 2 - 5。doi: 10.1136 / gut.44.1.2
9.Eibl MM, Wolf HM, Fürnkranz H, Rosenkranz A. IgA-IgG喂养预防低出生体重儿坏死性小肠结肠炎。N英语J医学(1988)319: 1 - 7。doi: 10.1056 / NEJM198807073190101
10.Tjellström B,施当恒L,埃里克森S,马格努松KE。口服免疫球蛋白A补充剂治疗艰难梭状芽胞杆菌肠炎。《柳叶刀》(1993)341: 701 - 2。0140 - 6736 . doi: 10.1016 / (93) 90477 - x
11.Hammarström V, Smith CI, Hammarström L.口服免疫球蛋白治疗空肠弯曲杆菌肠炎。《柳叶刀》(1993)341: 1036。0140 - 6736 . doi: 10.1016 / (93) 91136 - a
12.Giraudi V, Riganti C, Torales MR, Sédola H, Gaddi E.儿童上呼吸道感染:对鼻内给药IgA的反应。儿科耳鼻咽喉(1997)39: 103 - 10。doi: 10.1016 / s0165 - 5876 (96) 01472 - 3
13.Heikkinen T, Ruohola, Ruuskanen O, Waris M, Uhari M, Hammarström L.鼻内注射免疫球蛋白预防儿童鼻炎。儿科感染病学(1998)17: 367 - 72。doi: 10.1097 / 00006454-199805000-00004
14.马江克,Hikmat BY, Wycoff K, Vine ND, Chargelegue D, Yu L,等。重组植物单克隆分泌抗体的特性及其在人体内的预防性免疫治疗。Nat地中海(1998)4: 601 - 6。doi: 10.1038 / nm0598 - 601
16.Zeitlin L, Cone RA, Whaley KJ。利用单克隆抗体预防流行性传染病粘膜传播。新兴感染疾病(1999)5: 54 - 64。doi: 10.3201 / eid0501.990107
17.Corthésy B.被动免疫治疗中的重组分泌免疫球蛋白A:连接免疫学和生物技术。Curr Pharm生物技术公司(2003)4: 51 - 67。doi: 10.2174 / 1389201033378020
19.Iankov ID, Petrov DP, Mladenov IV, Haralambieva IH, Kalev OK, Balabanova MS,等。IgA单克隆抗体对O和H抗原在小鼠鼻内感染模型中的保护作用沙门氏菌血清血清型肠炎。微生物感染(2004)6: 901 - 10。doi: 10.1016 / j.micinf.2004.05.007
20.Apter FM, Michetti P, Winner LS III, Mack JA, Mekalanos JJ, Neutra先生抗脂多糖和抗霍乱毒素免疫球蛋白A (IgA)抗体在预防霍乱中的作用分析霍乱弧菌通过体内单克隆IgA抗体和霍乱毒素。感染Immun(1993)61: 5279 - 85。
21.Phalipon A, Kaufmann M, Michetti P, Cavaillon JM, Huerre M, Sansonetti P,等。单克隆免疫球蛋白A抗体,针对血清型特异性表位弗氏志贺菌脂多糖可预防小鼠实验性志贺氏菌病。J实验医学(1995)182: 769 - 78。doi: 10.1084 / jem.182.3.769
22.Blanchard TG, Czinn SJ, Maurer R, Thomas WD, Soman G, Nedrud JG。脲酶特异性单克隆抗体预防幽门螺杆菌猫属小鼠感染。感染Immun(1995)63: 1394 - 9。
23.韦tzin R, Traina-Dorge V, Soike K, Zhang JY, Mack P, Soman G,等。呼吸道合胞病毒鼻内IgA单克隆抗体保护恒河猴免受上呼吸道和下呼吸道感染。J感染病(1996)174: 256 - 61。doi: 10.1093 / infdis / 174.2.256
25.Silvey KJ, Hutchings AB, Vajdy M, Petzke MM, Neutra先生免疫球蛋白A对呼肠孤病毒进入小鼠Peyer贴片的保护作用。J性研究(2001)75: 10870 - 9。doi: 10.1128 / jvi.75.22.10870 - 10879.2001
26.赫钦斯AB, Helander A, Silvey KJ, Chandran K, Lucas WT, Nibert ML,等。针对呼肠孤病毒1型朗的sigma1外衣壳蛋白的分泌免疫球蛋白A抗体可预防小鼠Peyer 's斑块感染。J性研究(2004)78: 947 - 57。doi: 10.1128 / jvi.78.2.947 - 957.2004
27.Renegar KB, Small PA Jr, Boykins LG, Wright PF. IgA与IgG在小鼠呼吸道流感病毒感染控制中的作用。J Immunol(2004)173: 1978 - 86。
28.Winner L III, Mack J, Weltzin R, Mekalanos JJ, Kraehenbuhl JP, Neutra MR.粘膜免疫分析的新模型:杂交瘤肿瘤肠道分泌特异性单克隆免疫球蛋白A可预防霍乱弧菌感染。感染Immun(1991)59: 977 - 82。
29.Burns JW, Siadat-Pajouh M, Krishnaney AA, Greenberg HB。轮状病毒vp6特异性IgA单克隆抗体缺乏中和活性的保护作用。科学(1996)272: 104 - 7。doi: 10.1126 / science.272.5258.104
30.Wijburg OL, Uren TK, Simpfendorfer K, Johansen FE, Brandtzaeg P, Strugnell RA。先天分泌的抗体可以抵抗天然的抗体鼠伤寒沙门氏菌感染。J实验医学(2006)203: 21 - 6。doi: 10.1084 / jem.20052093
31.阿尔瓦雷斯N,奥特罗O,卡马乔F, Borrero R, Tirado Y, Puig A,等。被动给药纯化的人初乳分泌IgA诱导对结核分枝杆菌在进行性肺部感染小鼠模型中。BMC Immunol(2013)14: S3 doi: 10.1186 / 1471 - 2172 - 14 - s1 - S3
32.Lycke N, Erlandsson L, Ekman L, Schön K, Leanderson T. J链缺失抑制肠道IgA转运,抑制肠道抗毒保护的发展。J Immunol(1999)163: 913 - 9。
33.Schwartz-Cornil I, Benureau Y, Greenberg H, Hendrickson BA, Cohen J.轮状病毒内衣壳蛋白诱导的异源保护需要粘膜免疫球蛋白的转胞作用。J性研究(2002)76: 8110 - 7。doi: 10.1128 / jvi.76.16.8110 - 8117.2002
35.david BJ, Palm JE, Housley MP, Smith JR, Andersen YS, Martin MG,等。多聚免疫球蛋白受体在肠道免疫防御腔内居住的原生动物寄生虫贾第虫属.J Immunol(2006)177: 6281 - 90。
36.Blutt SE, Miller AD, Salmon SL, Metzger DW, Conner ME。IgA对清除和保护轮状病毒感染很重要。粘膜Immunol(2012)5: 712 - 9。doi: 10.1038 / mi.2012.51
37.Apter FM, Lencer WI, Finkelstein RA, Mekalanos JJ, Neutra MR.抗霍乱毒素单克隆免疫球蛋白A抗体可防止毒素诱导的氯化物分泌反应,并在体外阻断毒素与肠上皮细胞的结合。感染Immun(1993)61: 5271 - 8。
38.Stubbe H, Berdoz J, Kraehenbuhl JP, Corthésy B.聚合IgA在预防疾病方面优于携带相同可变结构域的单体IgA和IgG艰难梭状芽胞杆菌对T84单分子层的破坏。J Immunol(2000)164: 1952 - 60。
39.Mantis NJ, McGuinness CR, Sonuyi O, Edwards G, Farrant SA。免疫球蛋白A抗体对抗蓖麻毒素A和B亚单位保护上皮细胞免受蓖麻毒素中毒。感染Immun(2006)74: 3455 - 62。doi: 10.1128 / IAI.02088-05
40.张晓明,朱志强,朱晓明,朱志强,Corthésy B,等。携带b12 V区域的人单克隆IgG和IgA抗体抑制HIV-1传染性和上皮细胞转移J Immunol(2007)179: 3144 - 52。
42.Alfsen A, Iniguez P, bougyon E, Bomsel M.分泌IgA对gp41包膜糖蛋白保守表位的特异性抑制HIV-1上皮转胞作用。J Immunol(2001)166: 6257 - 65。
43.刘志强,李志强,陈志强,陈志强,等。肠致病菌黏附抑制作用的研究进展大肠杆菌通过人初乳和母乳的免疫球蛋白和低聚糖部分转移到HEp-2细胞。J感染病(1991)163: 1247 - 55。doi: 10.1093 / infdis / 163.6.1247
44.Renegar KB, Jackson GD, Mestecky J.单克隆单体IgA、聚合IgA和分泌IgA生物学活性的体外比较。J Immunol(1998)160: 1219 - 23所示。
46.藤冈H,解放者SN,相川M,黄ds,布拉尼克F,卡尔班T,等。特异免疫球蛋白A与仙台病毒蛋白在感染极化上皮细胞中的免疫细胞化学共定位。J实验医学(1998)188: 1223 - 9。doi: 10.1084 / jem.188.7.1223
47.Ruggeri FM, Johansen K, Basile G, Kraehenbuhl JP, Svensson L.抗轮状病毒免疫球蛋白A通过上皮细胞转运后体外中和病毒并保护幼鼠腹泻。J性研究(1998)72: 2708 - 14所示。
48.Corthésy B, Benureau Y, Perrier C, Fourgeux C, Parez N, Greenberg H,等。轮状病毒抗vp6分泌免疫球蛋白A通过细胞内中和作用而不是通过免疫排斥作用发挥保护作用。J性研究(2006)80: 10692 - 9。doi: 10.1128 / JVI.00927-06
49.邦塞尔M,海曼M, Hocini H, Lagaye S, Belec L,杜邦C,等。抗HIV包膜蛋白dIgA或IgM在细胞内中和通过严密上皮屏障的HIV转胞作用。免疫力(1998)9: 277 - 87。doi: 10.1016 / s1074 - 7613 (00) 80610 - x
51.杰克逊,梅斯特基J, Moldoveanu Z,斯皮曼P.收集和处理人体粘膜分泌物。在:Ogra PL, Mestecky J, Lamm ME, Strober W, Bienenstock J, McGhee JR编辑。粘膜免疫.佛罗里达州圣地亚哥:学术出版社(1999)。p . 1567 - 75。
52.Kaetzel CS, Robinson JK, Chintalacharuvu KR, Vaerman JP, Lamm ME。聚合免疫球蛋白受体(分泌成分)介导免疫复合物在上皮细胞间的转运:IgA的局部防御功能。美国国立自然科学研究院(1991)88: 8796 - 800。doi: 10.1073 / pnas.88.19.8796
55.Bakema JE, van Egmond M.人免疫球蛋白A Fc受体Fcα ri:黏膜免疫的多面调节因子。粘膜Immunol(2011)4: 612 - 24。doi: 10.1038 / mi.2011.36
56.Uddin T, Harris JB, Bhuiyan TR, Shirin T, Uddin MI, Khan AI,等。孟加拉国霍乱患者的粘膜免疫反应。临床疫苗免疫(2011)18: 506 - 12所示。doi: 10.1128 / CVI.00481-10
57.李志强,李志强,李志强,等。o -特异性多糖和脂多糖的免疫应答比较霍乱弧菌孟加拉国成年霍乱患者中的O1。临床疫苗免疫(2012)19: 1712 - 21所示。doi: 10.1128 / CVI.00321-12
58.埃里亚松DG, Helgeby A, Schön K,尼格伦C, El-Bakkouri K, Fiers W,等。一种新型无毒的CTA1-DD联合ISCOMS佐剂载体对流感病毒有效的粘膜免疫。疫苗(2011)29: 3951 - 61。doi: 10.1016 / j.vaccine.2011.03.090
59.什卡里姆V, Monselize Y,西格尔N,赞巴尔I,霍夫V,加蒂BZ。以色列儿童的选择性IgA缺乏症。临床免疫学杂志(2010)30.: 761 - 5。doi: 10.1007 / s10875 - 010 - 9438 - x
61.倪建平,刘建平,杨建平,等。口服霍乱疫苗后b细胞在十二指肠黏膜的激活在iga缺乏或没有IgG亚类缺乏的受试者。扫描J免疫(1993)38: 201 - 8。doi: 10.1111 / j.1365-3083.1993.tb01714.x
63.Martinez-Gallo M, Radigan L, Almejún MB, Martínez-Pomar N, Matamoros N, Cunningham-Rundles C. CVID患者和健康杂合子的TACI突变和b细胞功能受损。过敏临床免疫(2013)131: 468 - 76。doi: 10.1016 / j.jaci.2012.10.029
65.Arulanandam BP, Raeder RH, Nedrud JG, Bucher DJ, Le J, Metzger DW。IgA免疫缺陷导致Th细胞启动不足,增加流感病毒感染的易感性。J Immunol(2001)166: 226 - 31所示。
66.杜彻斯,阿敏R, Cogné N, Delpy L, Sirac C, Pascal V,等。α免疫球蛋白链过早替换mu会损害淋巴生成和粘膜归巢,但会促进浆细胞成熟。美国国立自然科学研究院(2010)107: 3064 - 9。doi: 10.1073 / pnas.0912393107
67.Bonner A, Perrier C, Corthésy B, Perkins SJ。人类分泌成分的溶液结构及其对生物功能的影响。生物化学(2007)282: 16969 - 80。doi: 10.1074 / jbc.M701281200
69.Crottet P, Corthésy B.绘制小鼠IgA和携带表位替换的小鼠分泌成分之间的相互作用,揭示了结构域II和III在与IgA共价结合中的作用。生物化学(1999)274: 31456 - 62。doi: 10.1074 / jbc.274.44.31456
70.Bonner A, Almogren A, Furtado PB, Kerr MA, Perkins SJ。非平面分泌IgA2和近平面分泌IgA1溶液结构使它们不同的粘膜免疫反应合理化。生物化学(2009)284: 5077 - 87。doi: 10.1074 / jbc.M807529200
72.Longet S, Miled S, Lötscher M, Miescher SM, Zuercher AW, Corthésy B.人血浆来源的聚合IgA和IgM抗体与重组分泌成分结合产生生物活性的分泌样抗体。生物化学(2013)288: 4085 - 94。doi: 10.1074 / jbc.M112.410811
73.Duc M, Johansen FE, Corthésy B.抗原与分泌免疫球蛋白A的结合导致对肠道蛋白酶的敏感性降低,并增加与细胞Fc受体的结合。生物化学(2010)285: 953 - 60。doi: 10.1074 / jbc.M109.059220
75.约翰逊ME,拉尔森JM,汉森GC。结肠的两层黏液是由MUC2黏液蛋白组织的,而外层是宿主-微生物相互作用的立法者。美国国立自然科学研究院(2011)108: 4659 - 65。doi: 10.1073 / pnas.1006451107
76.Lebreton C, Ménard S, Abed J, Moura IC, Coppo R, Dugave C,等。分泌免疫球蛋白A、CD71和转谷氨酰胺酶-2之间的相互作用影响肠上皮细胞对麦胶蛋白多肽的通透性。胃肠病学(2012)143: 698 - 707。doi: 10.1053 / j.gastro.2012.05.051
77.Mathias A, Corthésy B.杂交瘤和初乳衍生的分泌性免疫球蛋白A识别革兰氏阳性肠道细菌是由碳水化合物介导的。生物化学(2011)286: 17239 - 47。doi: 10.1074 / jbc.M110.209015
78.Phalipon A, Cardona, Kraehenbuhl JP, Edelman L, Sansonetti PJ,和Corthésy B.分泌成分:体内分泌iga介导的免疫排斥的新作用。免疫力(2002)17: 107 - 15所示。doi: 10.1016 / s1074 - 7613 (02) 00341 - 2
79.Murthy AK, Chagenty BK, Troutman T, Guentzel MN,俞俊杰,Ali SK,等。含甘露糖的非特异性人分泌免疫球蛋白A低聚糖对霍乱弧菌生物膜形成的抑制作用。《公共科学图书馆•综合》(2011)6: e16847。doi: 10.1371 / journal.pone.0016847
80.Royle L, Roos A, Harvey DJ, Wormald MR, van Gijlswijk-Janssen D, el-Redwan RM,等。分泌IgA N-和o -聚糖提供先天免疫系统和适应性免疫系统之间的联系。生物化学(2003)278: 20140 - 53。doi: 10.1074 / jbc.M301436200
81.Hammerschmidt S, Talay SR, Brandtzaeg P, Chhatwal GS。SpsA,一种新型肺炎球菌表面蛋白,特异性结合分泌免疫球蛋白a和分泌成分。摩尔Microbiol(1997)25: 1113 - 24。doi: 10.1046 / j.1365-2958.1997.5391899.x
82.吕玲,林梅,李华,Corthésy B,张俊荣。人聚合免疫球蛋白受体结合链球菌引起的肺炎通过域3和域4。生物化学(2003)278: 48178 - 87。doi: 10.1074 / jbc.M306906200
83.罗R, Mann B, Lewis WS, Rowe A, Heath R, Stewart ML,等。肺炎链球菌主要黏附蛋白胆碱结合蛋白A的溶液结构。EMBO J(2005)24: 34-43。
84.张俊杰,莫斯托夫·柯,米兰姆,米奈野,石田S,大华基M,等。聚合免疫球蛋白受体使肺炎球菌在人鼻咽上皮细胞中易位。细胞(2000)102: 827 - 37。doi: 10.1016 / s0092 - 8674 (00) 00071 - 4
85.Perrier C, Sprenger N, Corthésy B.分泌成分上的聚糖参与对粘膜病原体的先天保护。生物化学(2006)281: 14280 - 7。doi: 10.1074 / jbc.M512958200
87.施罗腾H, Stapper C, Plogmann R, Köhler H, Hackre J, Hanisch FG。分泌免疫球蛋白A对s -绒毛不依赖fab的抗粘连作用大肠杆菌是由唾液寡糖介导的。感染Immun(1998)66: 3971 - 3。
88.Borén T, Falk P, Roth KA, Larson G, Normark s幽门螺杆菌由血型抗原介导的人胃上皮细胞。科学(1993)262: 1892 - 5。doi: 10.1126 / science.8018146
90.Mantis NJ, Forbes SJ。分泌IgA:在上皮边界阻止微生物病原体。Immunol投资(2010)39: 383 - 406。doi: 10.3109 / 08820131003622635
91.福布斯SJ, Eschmann M,螳螂NJ。抑制沙门氏菌血清抗脂多糖单克隆免疫球蛋白a抗体对血清型鼠伤寒的运动性和进入上皮细胞的影响。感染Immun(2008)76: 4137 - 44。doi: 10.1128 / IAI.00416-08
92.福布斯SJ, Martinelli D, Hsieh C, Ault JG, Marko M, Mannella CA,等。保护性单克隆IgA与O抗原的结合沙门氏菌血清血清型鼠伤寒影响3型分泌和外膜完整性。感染Immun(2012)80: 2454 - 63。doi: 10.1128 / IAI.00018-12
93.Amarasinghe JJ, D 'Hondt RE, Waters CM, Mantis NJ。暴露的沙门氏菌血清型沙门氏菌感染通过双胍酸环化酶依赖通路触发胞外多糖的产生。感染Immun(2013)81: 653 - 64。doi: 10.1128 / IAI.00813-12
94.Forbes SJ, Bumpus T, McCarthy EA, Corthésy B, Mantis NJ。瞬态抑制弗氏志贺菌由保护性o抗原特异性单克隆IgA分泌3型。MBio(2011)2: e00042-11。doi: 10.1128 / mBio.00042-11
95.斯托克斯CR,泰勒B,特纳MW。屋尘和草花粉过敏与特异性IgA抗体缺乏的关系。《柳叶刀》(1974)2: 485 - 8。doi: 10.1016 / s0140 - 6736 (74) 92014 - 5
96.Cunningham-Rundles C, Brandeis WE, Good RA, Day NK。牛奶沉淀,循环免疫复合物和IgA缺乏症。Adv Exp医学生物学(1978)107: 523 - 30。doi: 10.1007 / 978 - 1 - 4684 - 3369 - 2 - _59
97.Schwarze J, Cieslewicz G, Joetham A,孙LK,孙WN, Chang TW,等。抗原特异性免疫球蛋白A可防止致敏小鼠气道挑战后气道反应性增加和肺嗜酸性粒细胞增多。J呼吸急救医疗吗(1998)158: 519 - 25所示。doi: 10.1164 / ajrccm.158.2.9801014
98.Pilette C, Nouri-Aria KT, Jacobson MR, Wolcock LK, Detry B, Walker SM,等。草花粉免疫治疗诱导与粘膜TGF-β表达相关的过敏原特异性IgA2抗体反应。J Immunol(2007)178: 4658 - 66。
99.Hajek AR, Lindley AR, Favoreto S, Carter R, Schleimer RP, Kuperman DA。12/15脂氧合酶缺乏保护小鼠免受过敏性气道炎症,增加分泌IgA水平。过敏临床免疫(2008)122: 633 - 9。doi: 10.1016 / j.jaci.2008.06.021
One hundred.Smits HH, Gloudemans AK, van Nimwegen M, Willart MA, Soullié T, Muskens F,等。霍乱毒素B通过诱导分泌IgA抑制过敏性炎症。粘膜Immunol(2009)2: 331 - 9。doi: 10.1038 / mi.2009.16
101.Frossard CP, Hauser C, Eigenmann PA。在食物过敏小鼠模型中,肠道中抗原特异性分泌IgA抗体减少。过敏临床免疫(2004)114: 377 - 82。doi: 10.1016 / j.jaci.2004.03.040
102.图尔多,埃罗什,贝约特,穆苏,H,贝特贝德,诺尼,等。舌下向量化重组Bet v 1a对桦木花粉过敏性哮喘小鼠模型的疗效。疫苗(2013)31: 2628 - 37。doi:10.1016/ j.b ox .2013.03.041 pii: S0264-410X(13)00378-2,
103.Perrier C, Thierry A-C, Mercenier A, Corthésy B.致敏和耐受小鼠口服挑战时过敏原特异性抗体和细胞因子反应,肥大细胞反应性和肠通透性。临床Exp过敏(2009)40: 153 - 62。doi: 10.1111 / j.1365-2222.2009.03329.x
104.Karlsson MR, Johansen FE, Kahu H, Macpherson AJ, Brandtzaeg P.无分泌免疫系统的过敏和口服耐受。过敏(2010)65: 561 - 70。doi: 10.1111 / j.1398-9995.2009.02225.x
105.Janzi M, Kull I, Sjöberg R, Wan J, Melén E, Bayat N,等。早期生活中的选择性IgA缺乏:与儿童时期的感染和过敏性疾病有关。中国Immunol(2009)133: 78 - 85。doi: 10.1016 / j.clim.2009.05.014
106.Aghamohammadi A, Cheraghi T, Gharagozlou M, Movahedi M, Rezaei N, Yeganeh M,等。IgA缺乏:临床和免疫表型的相关性。临床免疫学杂志(2009)29: 130 - 6。doi: 10.1007 / s10875 - 008 - 9229 - 9
107.罗伊MJ, Varvayanis M.肠相关淋巴组织中圆顶上皮的发育:IgA与M细胞的关联。细胞组织测定(1987)248: 645 - 51。doi: 10.1007 / BF00216495
108.Mantis NJ,张MC, Chintalacharuvu KR, Rey J, Corthésy B, Neutra MR. IgA对小鼠Peyer 's patch M细胞的选择性粘附:一种新型IgA受体的证据。J Immunol(2002)169: 1844 - 51。
109.Weltzin R, Lucia-Jandris P, Michetti P, Fields BN, Kraehenbuhl JP, Neutra MR.肠道M细胞对免疫球蛋白的结合和经上皮转运:使用抗肠道病毒蛋白的单克隆IgA抗体进行演示。细胞生物学(1989)108: 1673 - 85。doi: 10.1083 / jcb.108.5.1673
110.Rey J, Garin N, Spertini F, Corthésy B.分泌IgA在肠M细胞转运后靶向Peyer 's patch树突和T细胞。J Immunol(2004)172: 3026 - 33所示。
112.Chirdo FG, Millington OR, Beacock-Sharp H, Mowat AM。肠固有层中的免疫调节树突状细胞。欧元J免疫(2005)35: 1831 - 40。doi: 10.1002 / eji.200425882
114.鲍曼J,帕克CG,螳螂新泽西州。DC-SIGN对分泌IgA的识别:肠内免疫监测的意义Immunol列托人(2010)131: 59 - 66。doi: 10.1016 / j.imlet.2010.03.005
115.Pasquier B, Lepelletier Y, Baude C, Hermine O, Monteiro RC。树突状细胞成熟过程中IgA受体(CD89和CD71)的差异表达和功能白细胞生物学(2004)76: 1134 - 41。doi: 10.1189 / jlb.0204101
116.Lecocq M, Detry B, Guisset A, Pilette C. fc α ri介导的人单核细胞和树突状细胞IL-12产生和IFN-γ启动的抑制J Immunol(2013)190: 2362 - 71。doi: 10.4049 / jimmunol.1201128
118.Corthésy B,考夫曼M, Phalipon A, Peitsch M, Neutra MR, Kraehenbuhl JP。通过口服途径插入重组分泌性免疫球蛋白A的病原体特异性表位具有免疫原性。生物化学(1996)271: 33670 - 7。
120.鲍丽尔,陈志强,陈志强,陈志强,等。秘书长伊加斡旋的中和弗氏志贺菌通过降低炎症回路防止肠道组织破坏。J Immunol(2009)183: 5879 - 85。doi: 10.4049 / jimmunol.0901838
122.Nell S, Suerbaum S, Josenhans C.微生物群对IBD发病机制的影响:小鼠感染模型的经验教训。Nat Rev微生物(2010)8: 564 - 77。doi: 10.1038 / nrmicro2403
124.铃木K,米克B, Doi Y,村松M,千叶T,本条T,等。缺iga肠道中节段丝状细菌的异常扩增。美国国立自然科学研究院(2004)101: 1981 - 6。doi: 10.1073 / pnas.0307317101
125.全CP, Berneman A, Pires R, Avrameas S, Bouvet JP。天然多反应性分泌免疫球蛋白A自身抗体可能成为人类感染的屏障。感染Immun(1997)65: 3997 - 4004。
126.Stoel M, Jiang HQ, van Diemen CC, Bun JC, Dammers PM, Thurnheer MC,等。B-1和B-2细胞来源的肠浆母细胞中限制性IgA库。J Immunol(2005)174: 1046 - 54。
129.Lindner C, Wahl B, Föhse L, Suerbaum S, Macpherson AJ, Prinz I,等。年龄、微生物群和T细胞在肠道中形成不同的个体IgA库。J实验医学(2012)209: 365 - 77。doi: 10.1084 / jem.20111980
130.杨晓东,李志强,李志强,等。产生iga的浆细胞起源于生发中心,是由人类b细胞受体参与诱导的。胃肠病学(2011)140: 947 - 56。doi: 10.1053 / j.gastro.2010.12.005
131.Bergqvist P, Stensson A, Hazanov L, Holmberg A, Mattsson J, Mehr R,等。多个Peyer 's斑块生发中心的再利用导致高度同步的寡克隆和亲和成熟的肠道IgA反应。粘膜Immunol(2013)6: 122 - 35。doi: 10.1038 / mi.2012.56
132.川本生,陈德华,马鲁也,铃木科,Doi Y, Tsutsuim Y,等。抑制性受体PD-1调节IgA的选择和肠道中的细菌组成。科学(2012)336: 485 - 9。doi: 10.1126 / science.1217718
133.Wei M, Shinkura R, Doy Y, Mayura M, Fagarasan S, Honjo t携带Aicda基因敲入突变导致体细胞高突变缺陷的小鼠肠道稳态受损,粘膜防御受损。Nat Immunol(2011)12: 264 - 70。doi: 10.1038 / ni.1991
134.根岸H, Miki S, Sarashina H, Taguchi-Atarashi N,中岛A, Matsuki K,等。IRF3对肠道稳态和微生物介导的Tslp基因诱导的重要贡献。美国国立自然科学研究院(2012)109: 21016 - 21所示。doi: 10.1073 / pnas.1219482110
136.于立龙,王景泰,魏顺生,倪永华。宿主-微生物的相互作用及肠上皮屏障功能的调节:从生理学到病理学。世界胃肠病生理学杂志(2012)3.: 27-43。doi: 10.4291 / wjgp.v3.i1.27
137.马志刚,张志刚,张志刚,杨志刚,Corthésy B.肠道Caco-2细胞对益生菌和分泌IgA反应的增强作用。生物化学(2010)285: 33906 - 13所示。doi: 10.1074 / jbc.M110.135111
139.Swidsinski A, Weber J, Loening-Baucke V, Hale LP, Lochs H.炎症性肠病患者黏膜菌群的空间组织和组成。临床微生物学(2005)43: 3380 - 9。doi: 10.1128 / jcm.43.7.3380 - 3389.2005
140.罗尔N, Favre L, Benyacoub J, Corthésy B.分泌性免疫球蛋白A在小鼠Peyer 's patch树突状细胞对共生细菌自然感知中的作用。生物化学(2012)287: 40074 - 82。doi: 10.1074 / jbc.M112.405001
141.Peterson DA, McNulty NP, Guruge JL, Gordon JI。IgA对共生细菌的反应作为肠道稳态的中介。细胞宿主微生物(2007)2: 328 - 39。doi: 10.1016 / j.chom.2007.09.013
143.麦克弗森AJ,斯莱克E,克金MB,麦科伊KD。粘膜防火墙对抗共生肠道微生物。Semin Immunopathol(2009)31: 145 - 9。doi: 10.1007 / s00281 - 009 - 0174 - 3
144.赫伯德,王志强,Förster R,王志强。单性肠淋巴组织动力学与功能研究。暴击Rev免疫(2008)28: 1-13。doi: 10.1615 / CritRevImmunol.v28.i1.10
145.Rescigno M, Urbano M, Valzasina B, Francolini M, Rotta G, Bonasio R,等。树突状细胞表达紧密连接蛋白,并穿透肠道上皮单层以取样细菌。Nat Immunol(2001)2: 361 - 7。doi: 10.1038/86373
146.H, Fallet M, de Bovis B, Méresse S, Gorvel JP。Peyer 's贴片树突细胞通过M细胞特异性的跨细胞孔延伸树突来采样抗原。胃肠病学(2012)142: 592 - 601. - e3。doi: 10.1053 / j.gastro.2011.11.039
关键词:分泌IgA,黏膜稳态,抗体,上皮细胞,感染因子,共生菌
引用:Corthésy B(2013)黏膜表面分泌IgA的多方面功能。前面。Immunol。4: 185。doi: 10.3389 / fimmu.2013.00185
收到:2013年5月21日;待发论文:2013年6月6日;
接受:2013年6月24日;网上发表:2013年7月12日。
编辑:
Nils Yngve Lycke瑞典哥德堡大学教授版权:©2013 Corthésy。这是一篇开放获取的文章,根据创作共用署名许可协议,允许在其他论坛上使用、分发和复制,前提是原作者和来源注明出处,并遵守有关任何第三方图形等的版权声明。
*通信:Blaise Corthésy,研发实验室,免疫和过敏,大学国立医院(CHUV), Rue du Bugnon, 1011洛桑,瑞士电子邮件:blaise.corthesy@chuv.ch