





摘要gydF4y2Ba
背景gydF4y2Ba肺功能是一种复杂的遗传表型,肥胖是其重要的危险因素之一。然而,关于它们共同的遗传基础的知识有限。大多数肺功能的全基因组关联研究(GWASs)都是基于欧洲人群,限制了在人群之间的普遍性。其他人群缺乏大规模的肺功能GWASs。gydF4y2Ba
方法gydF4y2Ba我们纳入了来自中国嘉orie生物银行(CKB)的100285名受试者。为了确定肺功能的新位点,对1 s的用力呼气量(FEV)进行单性状GWAS分析gydF4y2Ba1gydF4y2Ba)、强迫肺活量(FVC)及FEVgydF4y2Ba1gydF4y2Ba/FVC在CKB。然后,我们进行了肺功能和肥胖特征(身体质量指数(BMI)、BMI调整后的腰臀比和BMI调整后的腰围)之间的全基因组交叉特征分析,以调查CKB中共享的遗传效应。最后,在CKB中研究了肺功能的多基因风险评分(PRSs),并研究了它们与BMI对肺功能的相关性的相互作用。我们还与CKB并行进行了交叉性状分析,使用来自英国生物银行(UKB)的多达457 756名受试者,用于复制和研究祖先特异性效应。gydF4y2Ba
结果gydF4y2Ba我们发现了9个全基因组重要的FEV新位点gydF4y2Ba1gydF4y2Ba,六种为全呼吸性呼吸,三种为全呼吸性呼吸gydF4y2Ba1gydF4y2Ba/FVC在CKB。FEVgydF4y2Ba1gydF4y2Ba在CKB和UKB中,FVC与肥胖性状均呈显著负相关。肺功能与肥胖性状之间共享的遗传位点突出了重要的生物学途径,包括细胞增殖、胚胎、骨骼和组织发育以及基因表达的调控。孟德尔随机化分析显示BMI对FEV有显著的负因果影响gydF4y2Ba1gydF4y2Ba以及CKB和UKB的FVC。在平均随访8年期间,肺功能PRSs显著改善了BMI变化对肺功能变化的影响。gydF4y2Ba
结论gydF4y2Ba这项大规模的肺功能GWAS鉴定了肺功能和肥胖之间的新位点和共同的遗传病因。根据受试者的多基因背景,BMI的变化可能会对肺功能的变化产生不同的影响。这些发现可能会为肥胖和肺功能改善的分子靶向疗法的发展开辟新的途径。gydF4y2Ba
摘要gydF4y2Ba
新的基因座为肺功能的遗传基础提供了更多的见解。了解肺功能和肥胖的共同遗传病因可能为肥胖和肺功能改善的分子靶向治疗开辟新的途径。gydF4y2Bahttp://bit.ly/38oCnezgydF4y2Ba
简介gydF4y2Ba
肺功能受损与慢性阻塞性肺疾病(COPD)等肺部疾病的风险和死亡率有关[gydF4y2Ba1gydF4y2Ba].临床及流行病学研究显示,许多危险因素可影响肺功能[gydF4y2Ba2gydF4y2Ba].在这些风险因素中,肥胖一直是增长最快的公共健康问题之一,在过去30年里,肥胖的患病率几乎增加了两倍[gydF4y2Ba3.gydF4y2Ba].具体而言,根据一项涉及121265名受试者的基于人群的研究,肥胖与肺功能下降的风险约高2倍相关(gydF4y2Ba如。gydF4y2Ba用力呼气量在1秒(FEVgydF4y2Ba1gydF4y2Ba)及强迫肺活量)[gydF4y2Ba4gydF4y2Ba].肥胖还会增加患呼吸道疾病的风险,如哮喘和慢性阻塞性肺病[gydF4y2Ba4gydF4y2Ba,gydF4y2Ba5gydF4y2Ba].然而,这些发现也提出了新的问题,即遗传危险因素是否会导致肺功能下降和肥胖并存。gydF4y2Ba
我们和其他人最近发现了呼吸道疾病的共同基因结构,包括哮喘和慢性阻塞性肺病[gydF4y2Ba6gydF4y2Ba- - - - - -gydF4y2Ba9gydF4y2Ba],表明影响这两种疾病的多效性作用。肺功能和肥胖都是高度遗传的特征,估计遗传率可达70% [gydF4y2Ba10gydF4y2Ba- - - - - -gydF4y2Ba14gydF4y2Ba].肺功能与肥胖之间的负相关表明这些疾病之间潜在的共同遗传风险因素[gydF4y2Ba15gydF4y2Ba].然而,关于肺功能和肥胖的共同遗传基础的知识是有限的。gydF4y2Ba
迄今为止,大多数肺功能全基因组关联研究(GWAS)的参与者都是欧洲血统[gydF4y2Ba13gydF4y2Ba,gydF4y2Ba14gydF4y2Ba,gydF4y2Ba16gydF4y2Ba,gydF4y2Ba17gydF4y2Ba];只有少数研究纳入了少量非欧洲参与者[gydF4y2Ba18gydF4y2Ba,gydF4y2Ba19gydF4y2Ba].因此,基于非欧洲人群的大规模GWASs对于扩展我们对不同人群遗传异质性的理解至关重要[gydF4y2Ba20.gydF4y2Ba,gydF4y2Ba21gydF4y2Ba].此外,了解肺功能与其他复杂特征的共享遗传结构是至关重要的(gydF4y2Ba如。gydF4y2Ba肥胖),这对环境混杂因素有很强的影响[gydF4y2Ba22gydF4y2Ba].因此,在目前的研究中,我们在中国嘉orie生物银行(CKB)和英国生物银行(UKB)进行了大规模的GWAS和跨性状/跨人群分析,以解决三个目标:1)确定包括FEV在内的肺功能性状的新遗传风险位点gydF4y2Ba1gydF4y2Ba、FVC和FEVgydF4y2Ba1gydF4y2Ba/中国人口植被覆盖度;2)研究中国和欧洲人群肺功能性状与肥胖性状(体重指数(BMI)、BMI调整腰臀比(WHRadjBMI)和BMI调整腰围(WCadjBMI)之间的共同遗传效应;3)采用CKB和UKB随访队列,考虑肺功能的多基因背景,研究基线BMI和BMI的纵向变化是否会影响肺功能。gydF4y2Ba
方法gydF4y2Ba
研究设计、设置和参与者gydF4y2Ba
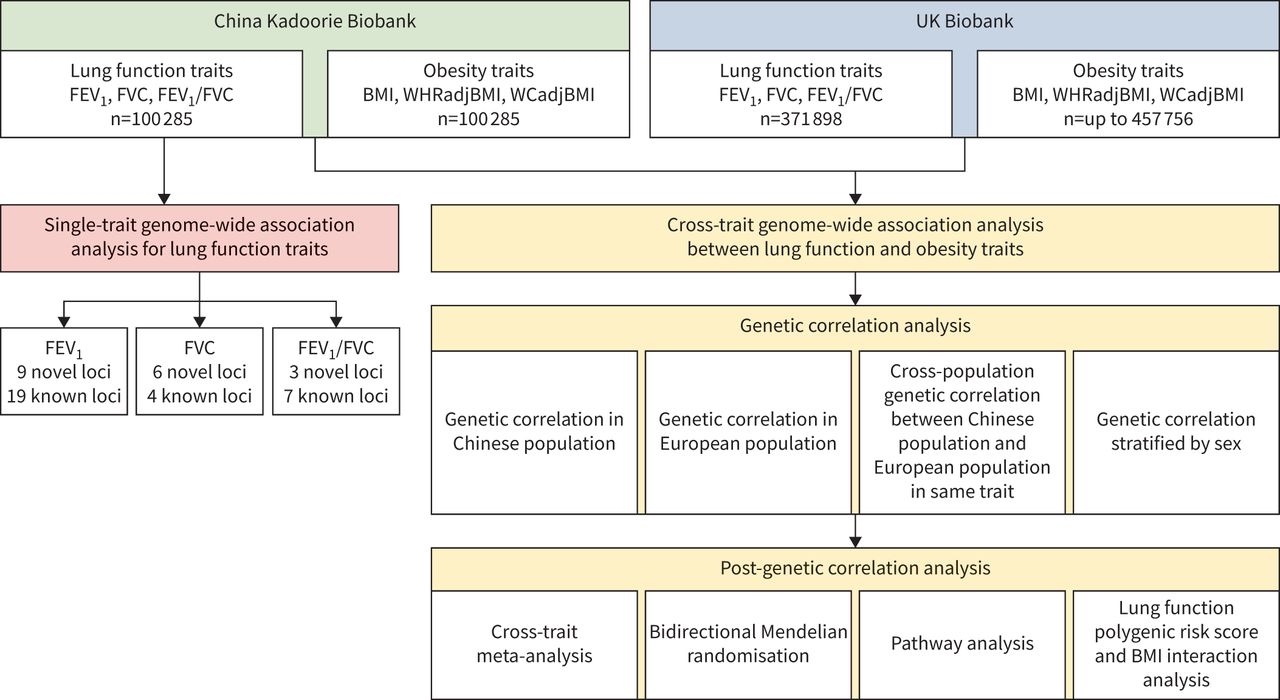
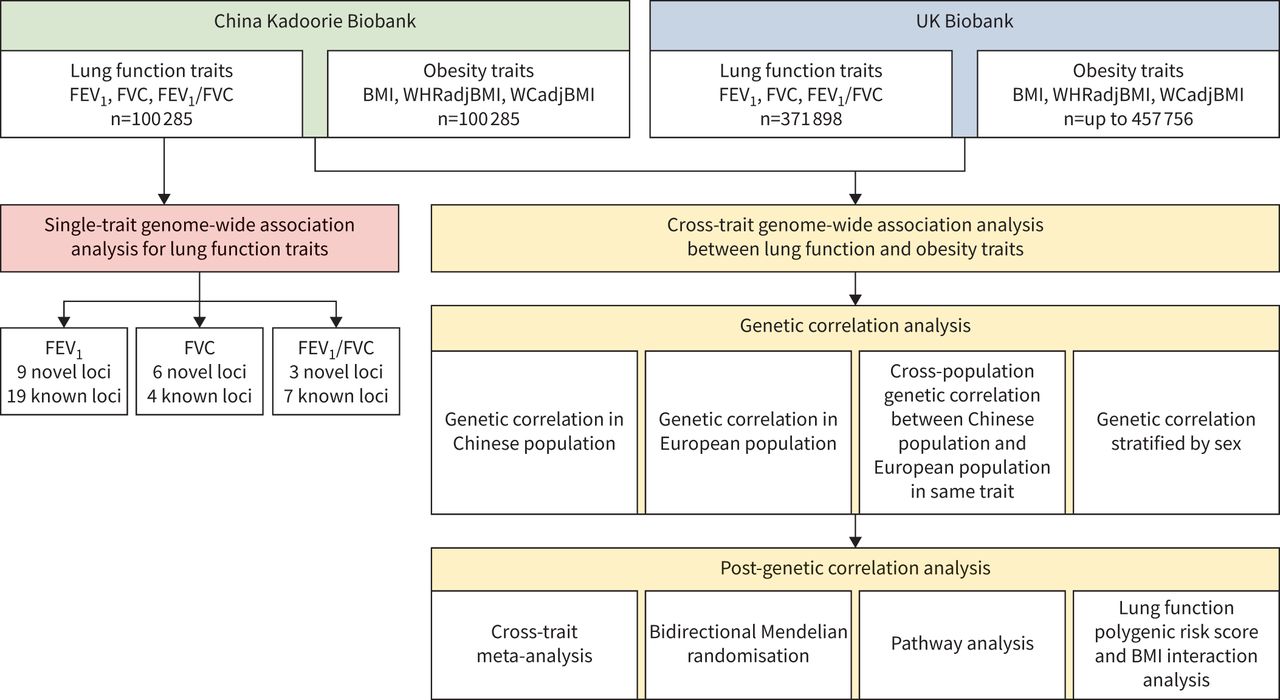
总体的研究设计可以在gydF4y2Ba图1gydF4y2Ba.简而言之,本研究分为两个分析阶段。第一阶段是通过单性状GWAS分析在中国人群中鉴定肺功能的新位点。第二阶段是通过在CKB和UKB中使用交叉性状GWAS分析来调查肺功能和肥胖之间的共同遗传效应。gydF4y2Ba
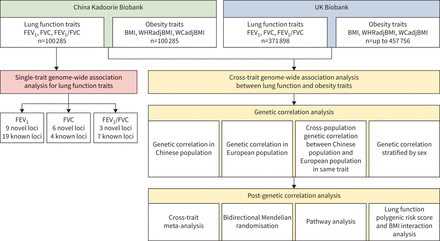
总体研究设计。FEVgydF4y2Ba1gydF4y2Ba:用力呼气量1 s;FVC:强迫肺活量;BMI:身体质量指数;WHRadjBMI: bmi调整后的腰臀比;WCadjBMI: bmi调整腰围。gydF4y2Ba
CKB研究是一项前瞻性队列研究,在中国有超过50万参与者。基建工程的详情已于先前介绍[gydF4y2Ba23gydF4y2Ba].简而言之,CKB在全国10个地区(哈尔滨、青岛、苏州、柳州、海口、河南、甘肃、四川、浙江和湖南)招募了512 715名30-79岁的成年人。所有参与者均给予知情的书面同意。在2004-2008年的基线调查中收集了问卷数据、物理测量数据和血液样本。2008年和2013-2014年进行了两次随访调查,随机选择了约5%的幸存参与者。gydF4y2Ba
UKB的研究已在其他地方详细描述[gydF4y2Ba7gydF4y2Ba,gydF4y2Ba24gydF4y2Ba].简而言之,UKB研究是一项前瞻性队列研究,有超过50万名参与者生活在英国。总共有503325名年龄在40岁至69岁之间的国民健康服务注册参与者从920万份邮寄邀请中被招募。通过问卷调查和人体测量评估收集基线数据(2004-2008年)。在UKB中,我们限制了欧洲血统的受试者。所有详细的基因分型、质量控制和归责程序均在UKB网站(gydF4y2Bahttp://biobank.ctsu.ox.ac.ukgydF4y2Ba).所有参与者均向UKB提供知情同意。gydF4y2Ba
肺功能与肥胖特征的确定gydF4y2Ba
FEVgydF4y2Ba1gydF4y2Ba、FVC和FEVgydF4y2Ba1gydF4y2Ba/FVC根据年龄、年龄平方、性别、身高、吸烟状况(曾经吸烟)进行调整gydF4y2Ba与gydF4y2Ba)和评估中心的线性回归模型[gydF4y2Ba14gydF4y2Ba].所得到的残差是逆正态变换[gydF4y2Ba14gydF4y2Ba].gydF4y2Ba
BMI(测量或自我报告体重(单位:公斤/身高,单位:平方米)在线性回归模型中根据年龄、年龄平方、性别和评估中心进行了调整[gydF4y2Ba14gydF4y2Ba].还测量了CKB和UKB参与者的腰围和臀围。腰臀比和腰围在线性回归模型中根据年龄、年龄平方、BMI、性别和评估中心进行调整。所得到的残差是反正态变换。详细的物理测量程序可参阅gydF4y2Ba补充材料gydF4y2Ba,以前的CKB研究[gydF4y2Ba25gydF4y2Ba]及UKB网站[gydF4y2Ba26gydF4y2Ba].gydF4y2Ba
CKB基因分型程序gydF4y2Ba
CKB进行了三个阶段的基因分型。牛津大学临床试验服务部门和流行病学研究部门(牛津,英国)与北京基因组研究所(中国深圳)和Affymetrix(现为美国加州圣克拉拉的赛默飞世尔科学公司)合作开发了一个定制设计的生物样本库阵列,为中国人口提供优化的全基因组覆盖。该700K单核苷酸多态性(SNP)阵列用于在第一阶段对约32 000名CKB参与者进行基因分型。在第二和第三阶段,对原始阵列进行了修订和更新,覆盖了~ 803K个SNPs,用于对~ 69000名参与者进行基因分型。gydF4y2Ba
变量与呼叫率>0.98,板块效应p>10gydF4y2Ba−6gydF4y2Ba,批量效果p>gydF4y2Ba−6gydF4y2Ba, HWE平衡(HWE)偏差p>gydF4y2Ba−6gydF4y2Ba(结合10个区域的10自由度卡方检验)和1000个基因组的小等位基因频率(MAF)差异东亚频率<0.2,结果在两个阵列版本中均存在532 415个双等位基因变体。对每条染色体的符合基因型进行SHAPEIT分期。然后,基于从1000个基因组III期获得的单倍型,用IMPUTE 4对每个5 mb间隔进行imputation。gydF4y2Ba
GWAS分析gydF4y2Ba
我们选择了不偏离HWE (p>1×10gydF4y2Ba−12gydF4y2Ba),每个变体缺失率<10%,每个样本缺失率<10%,MAF >1%, imputation质量评分(INFO) >0.8。详细的数据汇总,质量控制和imputation信息可以在gydF4y2Ba补充材料gydF4y2Ba.在CKB的100 285个样本和UKB的457 756个样本中进行了基因型-表型关联检测。对于肺功能和肥胖性状,我们进行了线性混合模型(LMM)关联分析,并对基因分型序列、CKB中的10个祖先主成分和UKB中的30个祖先主成分进行了调整,以评估在加性遗传模型下,性状的z-评分与估算基因型剂量之间的相关性,使用BOLT-LMM版本2.3 [gydF4y2Ba27gydF4y2Ba].在关联分析后,我们应用PLINK聚块函数来确定相互独立的顶部位点。具体来说,p<1×10的变量gydF4y2Ba−5gydF4y2Ba, rgydF4y2Ba2gydF4y2Ba>0.2和距离峰值<500 kb被分配到峰值的簇中。每个簇内的基因通过基因区域和簇区域的重叠来识别。新位点在两个层面上定义:聚集和变异(gydF4y2Ba补充材料gydF4y2Ba).简而言之,如果该独立簇区域与GWAS目录(检索日期:2020年4月3日)中同一性状的任何位点都没有重叠,我们将其定义为新位点。如果在聚块区域和GWAS目录之间有重叠,我们进一步检查前哨变异是否新颖,这是由低连锁不平衡(LD) r定义的gydF4y2Ba2gydF4y2Ba前哨变异与GWAS目录中簇区域内的任何变异之间<0.2。gydF4y2Ba
跨性状遗传相关gydF4y2Ba
我们使用跨性状LD评分回归(LDSC)来估计两个性状因果效应之间的遗传相关性(范围从−1到1),基于每个性状的GWAS的汇总统计[gydF4y2Ba28gydF4y2Ba].我们指定LDSC来估计回归截距,以解释不同性状之间的共享主题。gydF4y2Ba29gydF4y2Ba].我们应用Bonferroni校正(p<0.05/9)来解释LDSC分析中的多次测试。gydF4y2Ba
性别遗传相关性gydF4y2Ba
先前的研究表明,肺功能与肥胖之间的关系在男性和女性中可能有所不同[gydF4y2Ba30.gydF4y2Ba,gydF4y2Ba31gydF4y2Ba].因此,我们分别评估了男性和女性肺功能与肥胖之间的遗传相关性。gydF4y2Ba
跨群体遗传相关gydF4y2Ba
为了评估中国和欧洲人群之间的遗传异质性,我们还利用S-LDXR估计了肺功能和肥胖性状的全基因组跨人群遗传相关性[gydF4y2Ba32gydF4y2Ba]使用基线- ld - x模型注释。我们应用Bonferroni校正(p<0.05/6)来解释S-LDXR分析中的多重检验。gydF4y2Ba
Cross-trait荟萃分析gydF4y2Ba
交叉表型关联(CPASSOC)结合了GWAS汇总统计的效应估计和标准误差,以检验SNP与这两个性状之间的关联假设[gydF4y2Ba33gydF4y2Ba].本研究使用了异构版本的CPASSOC(表)。gydF4y2Ba
sheet是一种基于固定效应模型的交叉表型元分析方法。当跨研究存在异质效应时,它会更强大,这在测试多种表型时很常见[gydF4y2Ba34gydF4y2Ba].sheet使用特征的样本量作为权重,而不是使用效果标准误差。它还可以解释由于不同研究或队列内部或之间的重叠或相关主题而导致的效应相关性。gydF4y2Ba
过度代表富集分析gydF4y2Ba
为了了解肺功能和肥胖之间共享的生物学途径,我们从肺功能和肥胖的聚块过程中提取了基因,并使用WebGestalt工具[gydF4y2Ba35gydF4y2Ba]以评估基因本体论(GO)生物通路中已识别基因的富集程度。如果它们在肺功能和肥胖方面都显著增强,我们认为它们是共享的生物途径。使用错误发现率(FDR)方法来校正多次测试。gydF4y2Ba
孟德尔随机分析gydF4y2Ba
我们应用了广义摘要基于数据的孟德尔随机化(GSMR) [gydF4y2Ba36gydF4y2Ba]在默认设置下,从GWAS汇总统计数据推断BMI和肺功能特征之间的假定因果关系。为避免MR分析中受试者重叠,我们使用了Biobank Japan的BMI GWAS (n=158 284) [gydF4y2Ba37gydF4y2Ba]和CKB的肺功能GWAS,我们使用GIANT BMI GWAS (ngydF4y2Ba马克斯gydF4y2Ba=322 154) [gydF4y2Ba12gydF4y2Ba]和UKB的肺功能GWAS。关于Biobank Japan和GIANT GWAS数据的更详细描述可以在gydF4y2Ba补充材料gydF4y2Ba.由于GSMR要求至少10个ld独立仪器,p<5×10gydF4y2Ba−8gydF4y2Ba,我们将分析限制在满足这一标准的特征上。在运行GSMR之前,我们删除了链模糊、INFO <0.9和在人类白细胞抗原(gydF4y2BaHLAgydF4y2Ba)区域(chr6:25-34M)。我们应用Bonferroni校正(0.05/6)来解释MR分析中的性状对数量。gydF4y2Ba
肺功能多基因风险评分与BMI相互作用(PRSgydF4y2Ba肺功能gydF4y2Ba×BMI)分析gydF4y2Ba
我们使用LDpred构建了三个肺功能性状的多基因风险评分(PRSs) [gydF4y2Ba38gydF4y2Ba].有关PRS建设的详情,请参阅gydF4y2Ba补充材料gydF4y2Ba.我们还利用S报告的279个snp的权重构建了另外3个肺功能PRSsgydF4y2BahrinegydF4y2Baet al。gydF4y2Ba[gydF4y2Ba14gydF4y2Ba] (CKB数据中只有275个snp可用)。探讨肺功能PRSs与基线BMI或其纵向变化(ngydF4y2BaCKBgydF4y2Ba=21 791和ngydF4y2BaUKBgydF4y2Ba=12 019),我们拟合了两个线性回归模型来测试PRSgydF4y2Ba肺功能gydF4y2Ba×BMI的影响:gydF4y2Ba
基线模型:gydF4y2Ba 改变模型:gydF4y2Ba
改变模型:gydF4y2Ba 其中基线(gydF4y2BatgydF4y2Ba0)为2004-2010年,随访(gydF4y2BatgydF4y2Ba1)为2012-2014年;其他协变量包括CKB的主成分1-10和UKB的主成分1-30、年龄、年龄平方、性别、站立高度、吸烟状况(曾经/从未)、基因分型组和评估中心。对于基线模型,我们以正常BMI和十分位数2-9组为参考;对于变化模型,我们设定BMI稳定,以十分位数2-9组为参考。gydF4y2Ba
其中基线(gydF4y2BatgydF4y2Ba0)为2004-2010年,随访(gydF4y2BatgydF4y2Ba1)为2012-2014年;其他协变量包括CKB的主成分1-10和UKB的主成分1-30、年龄、年龄平方、性别、站立高度、吸烟状况(曾经/从未)、基因分型组和评估中心。对于基线模型,我们以正常BMI和十分位数2-9组为参考;对于变化模型,我们设定BMI稳定,以十分位数2-9组为参考。gydF4y2Ba
结果gydF4y2Ba
基于GWAS和snp的遗传力gydF4y2Ba
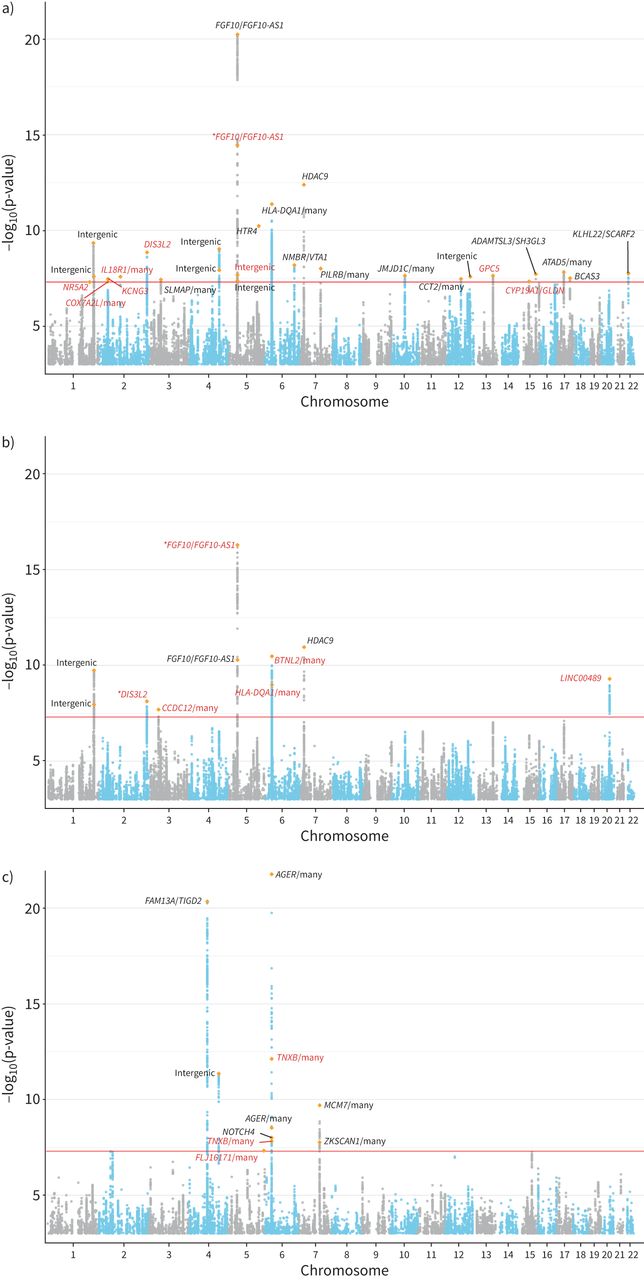
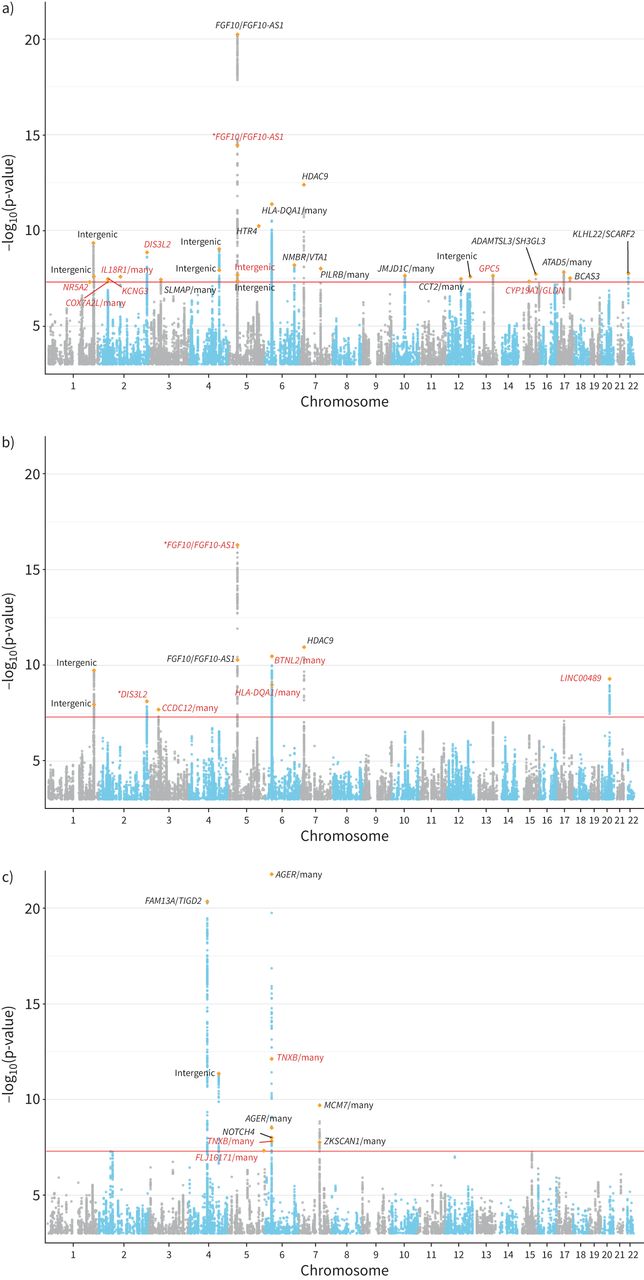
CKB和UKB队列的基线人口学特征总结于gydF4y2Ba补充表S1gydF4y2Ba.所有性状的GWAS结果均未显示因种群分层而导致的膨胀(gydF4y2Ba补充图S1和S2gydF4y2Ba).在CKB中,LDSC对观察尺度上基于snp的遗传力的估计为(均值±gydF4y2BasegydF4y2BaFEV为13.07±0.88%gydF4y2Ba1gydF4y2BaFVC为11.12±0.82%,FEV为5.12±0.67%gydF4y2Ba1gydF4y2BaBMI为21.77±1.23%,WHRadjBMI为8.72±0.83%,WCadjBMI为10.65±0.95%。我们确定了28个全基因组显著性(p<5×10gydF4y2Ba−8gydF4y2Ba) FEV基因座gydF4y2Ba1gydF4y2BaFVC为10,FEV为10gydF4y2Ba1gydF4y2Ba/ FVC (gydF4y2Ba图2gydF4y2Ba).将FEV与GWAS目录结果进行比较gydF4y2Ba1gydF4y2Ba、FVC和FEVgydF4y2Ba1gydF4y2Ba/ FVC (gydF4y2Ba补充表S2-S4gydF4y2Ba),我们共确定了18个肺功能性状的新位点(gydF4y2Ba表1gydF4y2Ba).我们进一步对最近发表的大规模肺功能GWAS (gydF4y2Ba补充表S5gydF4y2Ba) SgydF4y2BahrinegydF4y2Ba等gydF4y2Ba.[gydF4y2Ba14gydF4y2Ba].在S中共有11个位点可用gydF4y2BahrinegydF4y2Ba等gydF4y2Ba.[gydF4y2Ba14gydF4y2Ba)数据。其中4项显著(p<0.05/13),且效应量方向一致。未复制的位点可能是由于CKB和S之间不同的效应等位基因频率gydF4y2BahrinegydF4y2Ba等gydF4y2Ba.[gydF4y2Ba14gydF4y2Ba] (gydF4y2Ba补充表S5gydF4y2Ba).例如,CKB中前哨SNP rs1861229的MAF为0.52,而S中为0.17gydF4y2BahrinegydF4y2Ba等gydF4y2Ba.[gydF4y2Ba14gydF4y2Ba)的研究。在这些先前报道的位点中,显示全基因组与SgydF4y2BahrinegydF4y2Ba等gydF4y2Ba.[gydF4y2Ba14gydF4y2Ba],gydF4y2Ba蒸机gydF4y2Ba,gydF4y2BaAP4M1gydF4y2Ba,gydF4y2BaDIS3L2gydF4y2Ba,gydF4y2BaFAM13AgydF4y2Ba,gydF4y2BaFGF10gydF4y2Ba,gydF4y2BaHLA-DQA1gydF4y2Ba而且gydF4y2BaHTR4gydF4y2Ba是在肺功能中起重要作用的显著基因。就新基因座而言,我们确定了gydF4y2BaGPC5gydF4y2Ba作为FEV的新基因gydF4y2Ba1gydF4y2Ba(sentinel SNP rs528366: p=2.30×10gydF4y2Ba−8gydF4y2Ba).此外,我们发现20q11.23是FVC的一个新区域(前哨SNP rs6063386: p=5.00×10gydF4y2Ba−10gydF4y2Ba).前哨SNP被映射到一个长基因间非蛋白质编码RNA (lncRNA)上,gydF4y2BaLINC00489gydF4y2Ba.在FEV的新位点中gydF4y2Ba1gydF4y2Ba/FVC, 2个位于6p21.33区域,前哨SNP被映射到gydF4y2BaTNXBgydF4y2Ba众所周知,它与肺功能特征(FEVgydF4y2Ba1gydF4y2Ba和FEVgydF4y2Ba1gydF4y2Ba/FVC)及慢性阻塞性肺病[gydF4y2Ba39gydF4y2Ba,gydF4y2Ba40gydF4y2Ba].关于三个肺功能性状的全基因组显著位点的详细汇总统计信息可以在gydF4y2Ba补充表S6-S8gydF4y2Ba.gydF4y2Ba
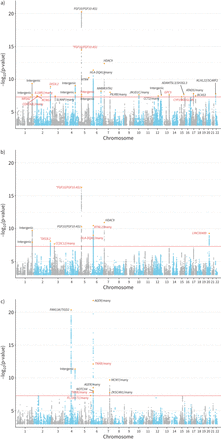
曼哈顿图用于全基因组关联分析100285名中国受试者在中国嘉orie生物银行队列的三个肺功能特征:a) 1秒用力呼气量(FEV)gydF4y2Ba1gydF4y2Ba), b)强迫肺活量(FVC)和c) FEVgydF4y2Ba1gydF4y2Ba/ FVC。的gydF4y2BaxgydF4y2Ba-轴表示基因组位置(染色体1-22);的gydF4y2BaygydF4y2Ba-axis表示-loggydF4y2Ba10gydF4y2Ba(p-value)关联测试,并从-log开始gydF4y2Ba10gydF4y2Ba(假定值)= 3。每个独立簇中最重要的新变体用橙色菱形符号突出显示。黑色的基因是以前报道过的,红色的基因是新的。某些基因上的星号表示一种新的变异。全基因组显著性水平(p=5×10gydF4y2Ba−8gydF4y2Ba)以红线表示。gydF4y2Ba
18个与1 s用力呼气量(FEV)相关的新位点gydF4y2Ba1gydF4y2Ba)、强迫肺活量(FVC)及FEVgydF4y2Ba1gydF4y2Ba/FVC在中国嘉道理生物库gydF4y2Ba
在UKB中,LDSC对观察尺度上基于snp的遗传力的估计为(均值±gydF4y2BasegydF4y2BaFEV为20.13±0.76%gydF4y2Ba1gydF4y2BaFVC为20.26±0.74%,FEV为23.95±1.43%gydF4y2Ba1gydF4y2BaBMI为27.41±1.07%,WHRadjBMI为13.88±0.94%,WCadjBMI为16.58±0.83%。单性状GWAS结果与SgydF4y2BahrinegydF4y2Ba等gydF4y2Ba.[gydF4y2Ba14gydF4y2Ba)的研究。gydF4y2Ba
肺功能与肥胖性状的遗传相关性gydF4y2Ba
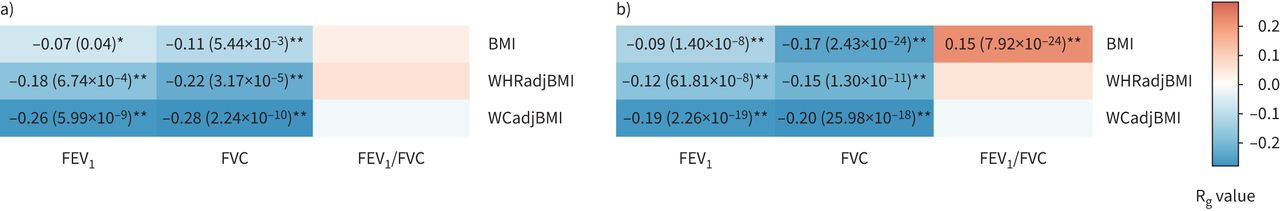
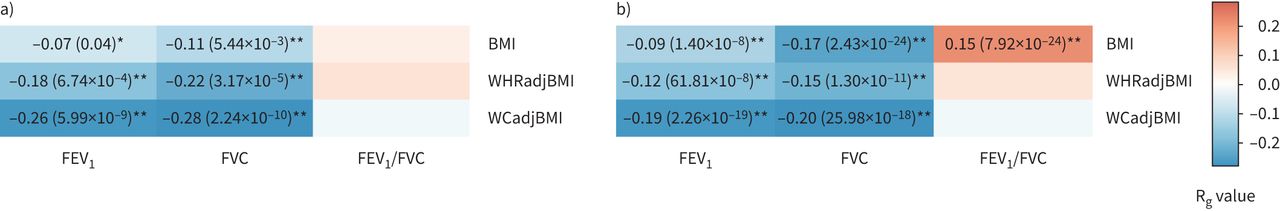
我们研究了CKB和UKB患者肺功能与肥胖特征之间的遗传相关性。如gydF4y2Ba图3gydF4y2Ba,我们发现CKB的两个肺功能性状与肥胖性状存在显著的遗传负相关(gydF4y2Ba如。gydF4y2BaRgydF4y2BaggydF4y2Ba=−0.26,p=5.99×10gydF4y2Ba−9gydF4y2Ba对于FEVgydF4y2Ba1gydF4y2Ba-WCadjBMI RgydF4y2BaggydF4y2Ba=−0.11,p=5.44×10gydF4y2Ba−3gydF4y2BaFVC-BMI和RgydF4y2BaggydF4y2Ba=−0.28,p=2.24×10gydF4y2Ba−10gydF4y2BaFVC-WCadjBMI)。我们发现肺功能与中枢性肥胖特征之间的遗传相关性通常比BMI更强。UKB的遗传相关性在大多数性状对中也与CKB的遗传相关性一致,尽管有一对性状对(gydF4y2Ba如。gydF4y2BaRgydF4y2BaggydF4y2Ba= 0.15, p = 7.92×10gydF4y2Ba−24gydF4y2Ba对于FEVgydF4y2Ba1gydF4y2Ba/ FVC-BMI在UKB中,但在CKB中不显著)(gydF4y2Ba图3gydF4y2Ba).性别特异性分析发现,女性肺功能与肥胖特征之间的遗传相关性强于男性(gydF4y2Ba补充表S9和S10gydF4y2Ba).此外,跨群体遗传相关分析表明,其中一个性状具有估计的跨群体RgydF4y2BaggydF4y2Ba显著<1 (RgydF4y2Bagcross-populationgydF4y2Ba= 0.86, p = 3.76×10gydF4y2Ba−6gydF4y2Ba的体重指数)(gydF4y2Ba补充表S11gydF4y2Ba).gydF4y2Ba
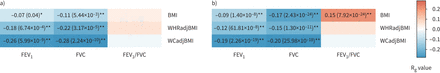
a)中国嘉orie生物银行(CKB)和b)英国生物银行(UKB)队列中三个肺功能特征和三个肥胖特征之间的全基因组遗传相关性。FEVgydF4y2Ba1gydF4y2Ba:用力呼气量1 s;FVC:强迫肺活量;BMI:身体质量指数;WHRadjBMI: bmi调整后的腰臀比;WCadjBMI: bmi调整腰围。每个盒子的颜色随遗传相关性的大小(RgydF4y2BaggydF4y2Ba).*:具有名义显著遗传相关的性状对(p<0.05);**:经多重检验校正后遗传相关性显著的性状对(p<0.05/9)。没有标记的框是遗传相关性不显著的性状对。gydF4y2Ba
Cross-trait荟萃分析gydF4y2Ba
对于Bonferroni校正后显示出显著遗传相关性的性状对(我们还包括BMI-FEV)gydF4y2Ba1gydF4y2Ba尽管p=0.038),我们应用CPASSOC进行全基因组跨性状元分析,以识别每个性状对之间共享的遗传变异(pgydF4y2Ba元gydF4y2Ba< 5×10gydF4y2Ba−8gydF4y2Ba;单一性状p < 1×10gydF4y2Ba−5gydF4y2Ba).共有CKB中的6个性状对和UKB中的7个性状对被纳入跨性状元分析。在CKB中,修剪后,我们发现了7个与BMI和FEV显著相关的位点gydF4y2Ba1gydF4y2BaWHRadjBMI和FEV共5个位点gydF4y2Ba1gydF4y2BaWCadjBMI和FEV共7个位点gydF4y2Ba1gydF4y2BaBMI和FVC共4个位点,WHRadjBMI和FVC共1个位点,WCadjBMI和FVC共1个位点。在这些基因座中,我们强调了三个共享基因座,因为它们是多对肺功能和肥胖特征的共享基因座。第一个轨迹是gydF4y2BaDIS3L2gydF4y2Ba2q37.1 (BMI-FEVgydF4y2Ba1gydF4y2Ba, WCadjBMI-FEVgydF4y2Ba1gydF4y2Ba、BMI-FVC和WCadjBMI-FVC)。第二个轨迹是gydF4y2BaHLA-DQA1gydF4y2Ba(BMI-FEVgydF4y2Ba1gydF4y2Ba和BMI-FVC)。第三个位点由几个前哨snp组成,均在12p13.2内定位,基因包括gydF4y2BaATXN2gydF4y2Ba而且gydF4y2BaACAD10gydF4y2Ba(gydF4y2Ba表2gydF4y2Ba).在CKB中确定的25个共享位点中,其中5个也在UKB中的相同特征对中确定(gydF4y2Ba表2gydF4y2Ba而且gydF4y2Ba补充表S12-S18gydF4y2Ba).gydF4y2Ba
25个肺功能(1 s用力呼气量(FEV))间共享基因位点gydF4y2Ba1gydF4y2Ba)、用力肺活量(FVC))和肥胖(身体质量指数(BMI)、BMI调整腰臀比(WHRadjBMI)和BMI调整腰围(WCadjBMI))特征gydF4y2Ba
路径分析gydF4y2Ba
为了从生物学角度深入了解共享基因,我们评估了GO生物过程类别中每个特征的独立位点的富集情况,以及肺功能和肥胖特征之间确定的共享基因集,并观察到UKB结果中许多显著富集情况(FDR:两种特征的q<0.05) (gydF4y2Ba补充表S19gydF4y2Ba).与共享位点的基因功能一致,氧化石墨烯生物过程强调了几种常见的肺功能和肥胖特征的生物学途径,如细胞增殖、胚胎、骨骼和组织发育以及基因表达的调控。然而,我们在CKB结果中没有观察到任何显著的富集。gydF4y2Ba
孟德尔随机gydF4y2Ba
我们应用GSMR在BMI和肺功能性状之间进行因果推断。在东亚人群分析中,我们观察到BMI(每标准差)对FEV有显著的负因果效应gydF4y2Ba1gydF4y2Ba(bgydF4y2BaxygydF4y2Ba=−0.08,p=2.46×10gydF4y2Ba−4gydF4y2Ba)和植被覆盖度(b .gydF4y2BaxygydF4y2Ba=−0.11,p=3.44×10gydF4y2Ba−7gydF4y2Ba) (gydF4y2Ba表3gydF4y2Ba).我们没有观察到BMI对FEV有显著的因果关系gydF4y2Ba1gydF4y2Ba/ FVC。在相反的方向,我们观察到一个小的量级或不显著的因果效应。gydF4y2Ba
估计身体质量指数(BMI)和肺功能特征(1 s用力呼气量(FEV))的因果效应大小gydF4y2Ba1gydF4y2Ba)、强迫肺活量(FVC)及FEVgydF4y2Ba1gydF4y2Ba/ FVC)gydF4y2Ba
PRSgydF4y2Ba肺功能gydF4y2Ba×体重指数分析gydF4y2Ba
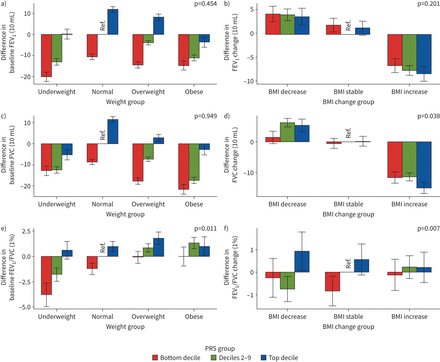
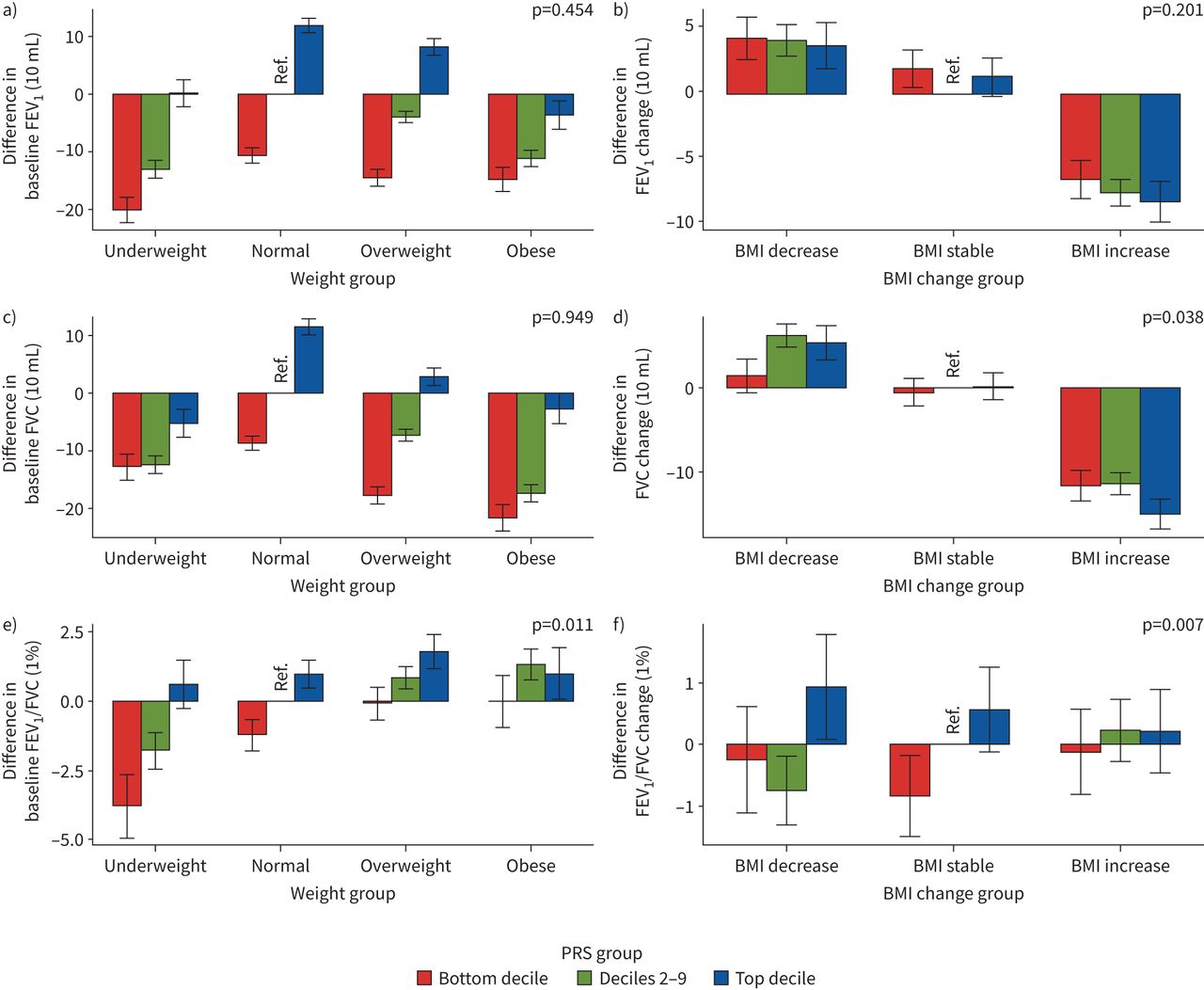
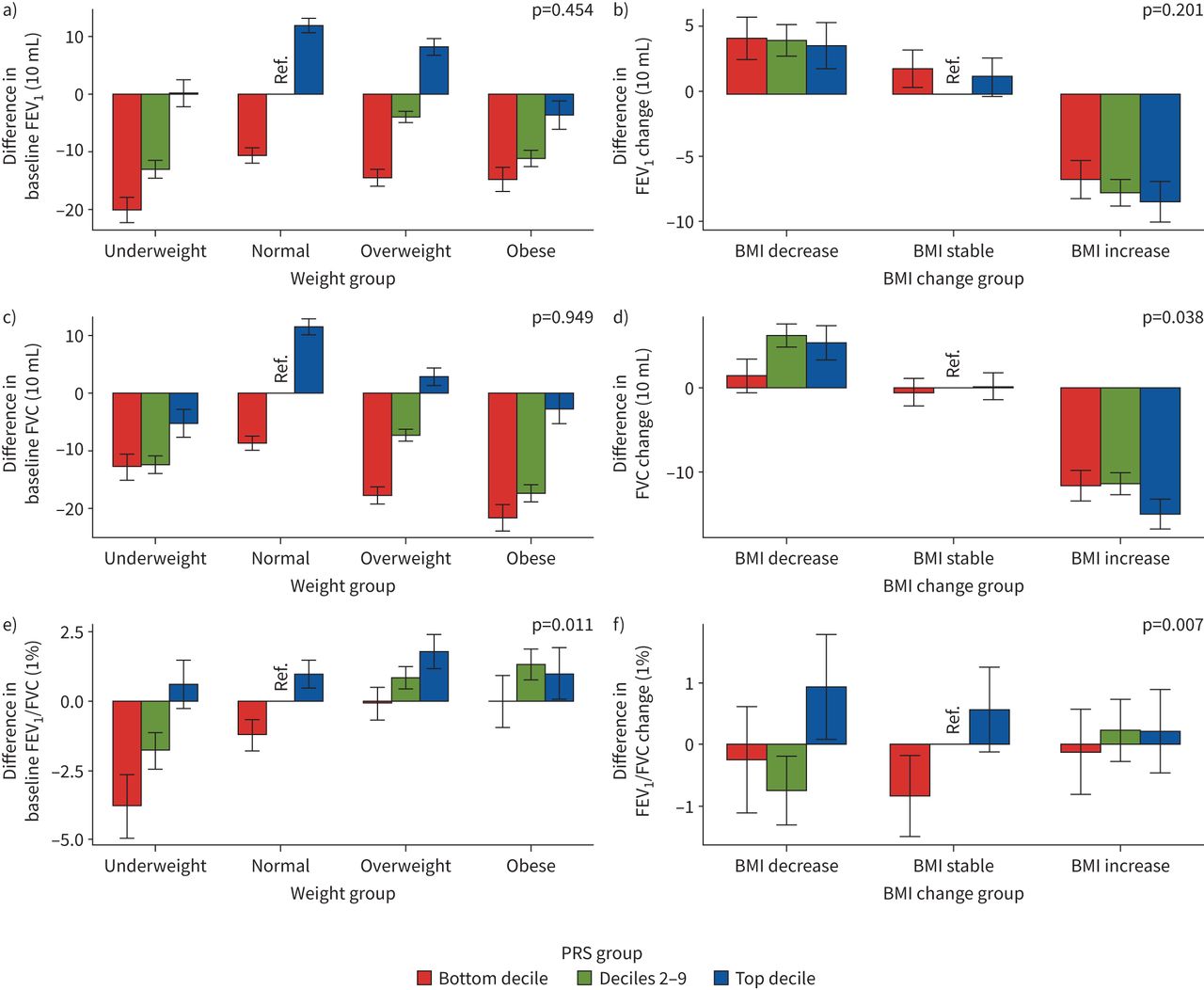
我们从LDpred中为每个肺功能特征构建了7个PRS模型,并选择了具有最高鉴别性能的PRS模型(RgydF4y2Ba2gydF4y2Ba)进行互动分析(gydF4y2Ba补充表S20gydF4y2Ba).我们发现FEV的基线BMI和PRS之间存在显著的相互作用gydF4y2Ba1gydF4y2Ba/FVC对FEVgydF4y2Ba1gydF4y2Ba/FVC (p=0.011) (gydF4y2Ba图4a和补充表S21gydF4y2Ba).一般而言,与参照组相比,PRS底部十分位数加体重不足组的FEV最低gydF4y2Ba1gydF4y2Ba.对于FVC,最大的减少是在最低的PRS十分位数加上肥胖组。对于FEVgydF4y2Ba1gydF4y2Ba/FVC,我们观察到PRS底部十分位数加体重不足组的FEV最低gydF4y2Ba1gydF4y2Ba/FVC,超重和肥胖组增加。在变化模型中,我们发现植被覆盖度的BMI变化与植被覆盖度的PRS变化存在显著的交互作用(p=0.038), fevs的BMI变化与植被覆盖度的PRS变化存在显著的交互作用gydF4y2Ba1gydF4y2Ba/FVC (p=0.007)对FEV变化的影响gydF4y2Ba1gydF4y2Ba/ FVC (gydF4y2Ba图4b和补充表S21gydF4y2Ba).总体而言,与参照组相比,BMI增加组FEV降低gydF4y2Ba1gydF4y2Ba植被覆盖度,且在PRS的前十分位数影响最大。对于UKB,我们在基线模型中没有发现显著的相互作用效应。然而,我们发现植被覆盖度BMI和PRS的变化对植被覆盖度变化(p=0.068)之间存在边缘性显著交互作用(gydF4y2Ba补充表S22和补充图S3gydF4y2Ba).最后,我们利用S中275个snp的权重构建了额外的肺功能PRSsgydF4y2BahrinegydF4y2Ba等gydF4y2Ba.[gydF4y2Ba14gydF4y2Ba]的研究,并没有发现任何显著的PRSgydF4y2Ba肺功能gydF4y2Ba×BMI效应(gydF4y2Ba补充表S23和补充图S4gydF4y2Ba).gydF4y2Ba
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
中国嘉多利生物库三种肺功能多基因风险评分(PRSs)分布与体重指数(BMI)的关系:a、c、e)基线模型和b、d、f)变化模型:a、b) 1 s用力呼气量(FEV)gydF4y2Ba1gydF4y2Ba), c, d)强迫肺活量(FVC)和e, f) FEVgydF4y2Ba1gydF4y2Ba/ FVC。对于基线模型,我们以正常BMI和十分位数2-9组为参考;对于变化模型,我们设定BMI稳定,以十分位数2-9组为参考。对于基线模型,使用gydF4y2BaxgydF4y2Ba-轴表示不同BMI类别,定义如下:体重不足BMI <18.5 kg·mgydF4y2Ba−2gydF4y2Ba,正常BMI 18.5 ~ 24.9 kg·mgydF4y2Ba−2gydF4y2Ba,超重BMI 25.0 ~ 29.9 kg·mgydF4y2Ba−2gydF4y2Ba肥胖BMI≥30.0 kg·mgydF4y2Ba−2gydF4y2Ba.的gydF4y2BaygydF4y2Ba-axis表示各组肺功能测量值与参考组的差值。对于变更模型,使用gydF4y2BaxgydF4y2Ba-轴表示不同BMI变化类别:BMI下降定义为BMIgydF4y2BatgydF4y2Ba1gydF4y2Ba−BMIgydF4y2BatgydF4y2Ba0gydF4y2Ba≤−1 kg·mgydF4y2Ba−2gydF4y2Ba, BMI稳定定义为- 1 kg·mgydF4y2Ba−2gydF4y2Ba< BMIgydF4y2BatgydF4y2Ba1gydF4y2Ba−BMIgydF4y2BatgydF4y2Ba0gydF4y2Ba≤1公斤·mgydF4y2Ba−2gydF4y2BaBMI增加定义为BMIgydF4y2BatgydF4y2Ba1gydF4y2Ba−BMIgydF4y2BatgydF4y2Ba0gydF4y2Ba> 1公斤·mgydF4y2Ba−2gydF4y2Ba.的gydF4y2BaygydF4y2Ba-axis表示肺功能测量值变化(肺功能gydF4y2BatgydF4y2Ba1gydF4y2Ba−肺功能gydF4y2BatgydF4y2Ba0gydF4y2Ba)为每个组别及参考组别。PRS组定义为:底部十分位数,十分位数2-9和顶部十分位数。每个图上的p值代表肺功能和基线BMI或BMI变化相互作用p值,来自基线或变化模型。gydF4y2Ba
讨论gydF4y2Ba
据我们所知,目前的研究是中国人群肺功能的最大GWAS。我们发现肺功能与肥胖特征之间有很强的遗传相关性和共享的遗传位点。我们在UKB中复制了这些中国的发现,并确定了特定人群的遗传效应。我们还发现了肺功能和肥胖特征之间的共同生物学途径,如细胞增殖、胚胎、骨骼和组织发育以及基因表达的调节。gydF4y2Ba
在本研究中,我们发现了9个新的FEV基因座gydF4y2Ba1gydF4y2Ba,六种为全呼吸性呼吸,三种为全呼吸性呼吸gydF4y2Ba1gydF4y2Ba/ FVC。其中,我们强调了一个与FEV相关的新基因gydF4y2Ba1gydF4y2Ba,gydF4y2BaGPC5gydF4y2Baq31.3 13日。gydF4y2BaGPC5gydF4y2Ba是草甘膦基因家族的一员。迄今为止的证据表明,glyypicans的主要功能是调节骨形态发生蛋白、Wnt、Hedgehog和成纤维细胞生长因子的信号通路[gydF4y2Ba41gydF4y2Ba],它们参与调节肺功能[gydF4y2Ba42gydF4y2Ba],肺纤维化[gydF4y2Ba43gydF4y2Ba]和慢性阻塞性肺病的病理生物学[gydF4y2Ba44gydF4y2Ba].gydF4y2BaGPC5gydF4y2Ba也被发现会增加从不吸烟的人患肺癌的风险[gydF4y2Ba45gydF4y2Ba].对于FVC,我们还发现了一个新的独立区域,20q11.23,在这里前哨SNP被映射到lncRNA,gydF4y2BaLINC00489gydF4y2Ba虽然该区域的功能还有待进一步研究。对于FEVgydF4y2Ba1gydF4y2Ba/FVC时,我们注意到几个独立的基因座位于6p21区域内,该区域以其与肺功能特征(FEV)相关而闻名gydF4y2Ba1gydF4y2Ba和FEVgydF4y2Ba1gydF4y2Ba/FVC)及慢性阻塞性肺病[gydF4y2Ba39gydF4y2Ba,gydF4y2Ba40gydF4y2Ba].这个区域含有基因,如gydF4y2Ba蒸机gydF4y2Ba,gydF4y2BaATF6BgydF4y2Ba,gydF4y2Ba新的高度gydF4y2Ba而且gydF4y2BaTNXBgydF4y2Ba,其中gydF4y2Ba蒸机gydF4y2Ba已被报道在肺功能中发挥潜在的功能作用[gydF4y2Ba46gydF4y2Ba].AGER蛋白是一种晚期糖基化终产物(RAGE)的受体,是免疫球蛋白超家族的一种多配体受体,可与与稳态、发育、炎症、糖尿病和神经退行性变有关的不同分子相互作用[gydF4y2Ba46gydF4y2Ba].RAGE信号取决于细胞类型和环境。暴怒的表达在接触香烟烟雾后增加,并部分负责诱导促炎信号通路(gydF4y2Ba如。gydF4y2BaNF -κB) [gydF4y2Ba47gydF4y2Ba].此外,我们还发现了两个新的基因位点gydF4y2BaDIS3L2gydF4y2Ba而且gydF4y2BaFGF10gydF4y2Ba/gydF4y2BaFGF10-AS1gydF4y2Ba)在两种FEV中均显著gydF4y2Ba1gydF4y2Ba和FVC,表明其在不同肺功能性状间的多效性。gydF4y2Ba
我们的LDSC分析显示,在中国和欧洲人群中,肺功能和肥胖特征之间存在很强的全基因组遗传相关性。我们观察到肥胖性状与FEV有很强的遗传负相关gydF4y2Ba1gydF4y2Ba但肥胖特征与FEV之间的遗传相关性不显著gydF4y2Ba1gydF4y2Ba/植被覆盖度。除BMI-FEV外,欧洲人群与中国人群的遗传相关结果基本一致gydF4y2Ba1gydF4y2Ba/FVC,在欧洲人群中也非常显著(RgydF4y2BaggydF4y2Ba= 0.15, p = 7.92×10gydF4y2Ba−24gydF4y2Ba).先前的研究表明FEVgydF4y2Ba1gydF4y2Ba和植被覆盖度在肥胖的情况下会减少[gydF4y2Ba48gydF4y2Ba],但FEVgydF4y2Ba1gydF4y2Ba/植被覆盖度比率通常不受影响[gydF4y2Ba49gydF4y2Ba].有几种生物学机制可以解释肺功能损害和肥胖之间的关系。首先,肥胖的机械效应导致气道狭窄和闭合,增加呼吸系统阻力。与健康体重者相比,肥胖患者气道狭窄与气道关闭和气道高反应性相关[gydF4y2Ba50gydF4y2Ba].气道狭窄及封闭导致空气滞留及通气不均匀[gydF4y2Ba51gydF4y2Ba].此外,我们发现肺功能与中枢性肥胖之间的遗传相关性比与全身性肥胖之间的遗传相关性更强。与不考虑脂肪分布的全身性肥胖相比,腹部和胸部脂肪更容易对肺功能损害起作用。这是因为它们在强制吸气时对横膈膜和胸壁扩张有直接的机械作用[gydF4y2Ba52gydF4y2Ba,gydF4y2Ba53gydF4y2Ba],是限制性肺部疾病的典型症状[gydF4y2Ba54gydF4y2Ba].gydF4y2Ba
跨性状荟萃分析发现肺功能和肥胖性状之间具有显著的独立位点。在中国人群中,基因座gydF4y2BaDIS3L2gydF4y2Ba在2q37.1发现多种肺功能和肥胖特征(BMI-FEV)之间共享gydF4y2Ba1gydF4y2Ba, WCadjBMI-FEVgydF4y2Ba1gydF4y2Ba中国人群的BMI-FVC和WCadjBMI-FVC;欧洲人群中的WC-FVC)。之前的一项研究发现gydF4y2BaDIS3L2gydF4y2Ba成为导致过度生长综合症的基因(gydF4y2Ba如。gydF4y2Ba帕尔曼综合症[gydF4y2Ba55gydF4y2Ba]),表明其在调节细胞生长和分裂中起着关键作用。这一功能也与通路分析的结果一致,共享基因主要富集在与细胞增殖和胚胎、骨骼和组织发育相关的通路中。值得注意的是,这些共享的途径显示了生长对肺功能和肥胖的重要作用,并且部分不同于最近的肺功能GWAS研究中确定的途径[gydF4y2Ba14gydF4y2Ba].不出所料,我们也发现了许多基因座gydF4y2BaHLAgydF4y2Ba这两个群体的肥胖特征和肺功能所共有的区域。gydF4y2BaHLAgydF4y2Ba是一种对多种复杂疾病具有丰富多效性的基因复合体[gydF4y2Ba6gydF4y2Ba,gydF4y2Ba7gydF4y2Ba,gydF4y2Ba9gydF4y2Ba,gydF4y2Ba56gydF4y2Ba]尤其参与免疫相关过程[gydF4y2Ba57gydF4y2Ba].在欧洲人群中,我们还确定了肺功能和肥胖特征之间的许多共享位点。然而,我们发现大多数共享位点在两个种群之间是不同的。gydF4y2Ba
尽管流行病学研究证实了肺功能与肥胖之间的关系[gydF4y2Ba5gydF4y2Ba,gydF4y2Ba15gydF4y2Ba,gydF4y2Ba31gydF4y2Ba,gydF4y2Ba58gydF4y2Ba,但目前尚不清楚肥胖是肺功能的一个驱动因素,还是其存在的共病。目前研究中的MR估计表明BMI对FEV有负因果影响gydF4y2Ba1gydF4y2BaFVC与FEV有正向因果关系gydF4y2Ba1gydF4y2Ba/ FVC。这些估计值提供了证据,证明BMI可能降低东亚和欧洲人群的肺功能,尽管BMI与FEV之间存在因果关系gydF4y2Ba1gydF4y2Ba/FVC仍然可以是双向的。BMI与肺功能性状的因果关系结果与W一致gydF4y2BaielschergydF4y2Ba等gydF4y2Ba.[gydF4y2Ba59gydF4y2Ba],显示出BMI与FEV的负因果关系gydF4y2Ba1gydF4y2BaFVC和BMI与FEV呈正相关gydF4y2Ba1gydF4y2Ba/ FVC。gydF4y2Ba
在这项研究中,我们发现了中国和欧洲人群遗传异质性的证据。在全基因组水平上,跨群体遗传相关分析表明BMI具有估计的跨群体RgydF4y2BaggydF4y2Ba统计学意义<1,表明中国和欧洲人群BMI基因调控存在异质性。在变异水平上,跨性状元分析显示,中国和欧洲群体中大多数共享变异是不同的,这可能是由于两个群体不同的遗传背景和样本量,以及gene×environment相互作用[gydF4y2Ba32gydF4y2Ba].gydF4y2Ba
最近的几项肺功能PRS研究集中在肺功能PRS与COPD风险之间的关系,以及与吸烟状况的相互作用[gydF4y2Ba14gydF4y2Ba,gydF4y2Ba60gydF4y2Ba].我们的研究调查了PRSgydF4y2Ba肺功能gydF4y2Ba×BMI在横断面和纵向设置。我们的相互作用分析显示,保持正常的BMI可以改善肺功能,并且这种有益效果在肺功能遗传特征高的受试者中更为显著。结果提示BMI可能是肺功能遗传影响的中介因素,这与MR结果表明BMI是肺功能的因果危险因素一致。我们还观察到,在BMI变化方面,最高PRS组表现出最有益的影响。这些发现对肺功能改善具有重要意义,因为它们可以在使用PRSs进行更精确的风险分层的基础上,在肺功能降低之前,为有风险的个体提供潜在的BMI干预。gydF4y2Ba
我们也承认当前研究的局限性。首先,CKB GWAS的样本量只有UKB的四分之一。这导致了一些在两个队列之间可能无法直接比较的发现。二是FVC和FEVgydF4y2Ba1gydF4y2Ba海口和青岛地区(n=14 000)的植被覆盖度测量结果与长江沿岸其他8个地区相比可能存在偏差。因此,我们进一步进行了去除这两个区域的敏感性分析。敏感度分析(gydF4y2Ba补充图S5和S6gydF4y2Ba)表示FVC和FEV的效应大小gydF4y2Ba1gydF4y2Ba/FVC新位点与初步分析高度一致(使用完整的GWAS队列),尽管由于功率较低,在去除两个区域后p值略有增加。此外,遗传相关敏感性分析结果与初步分析结果一致(gydF4y2Ba补充表S24gydF4y2Ba).CKB FEVgydF4y2Ba1gydF4y2Ba/FVC PRS也在最近的一项研究中使用,与其他独立的祖先群体相比,结果一致[gydF4y2Ba14gydF4y2Ba].第三,我们选择使用UKB中有两个时间点的肺功能肺活量测量(数据字段3062和3063),因此不能使用最清洁的肺功能肺活量测量(数据字段20 150和20 151)。然而,我们的GWAS结果与SgydF4y2BahrinegydF4y2Baet al。gydF4y2Ba[gydF4y2Ba14gydF4y2Ba这项研究使用了最清洁的肺功能肺活量测量方法。第四,尽管PRS的使用允许研究人员有效地捕获遗传效应的有用部分,但PRSgydF4y2Ba肺功能gydF4y2Ba×BMI分析仍然容易受到混淆或偏差的影响,因为PRS模型中的SNP标记物与BMI的因果变异之间存在LD [gydF4y2Ba61gydF4y2Ba,gydF4y2Ba62gydF4y2Ba].然而,我们的敏感性分析表明,这可能对PRS没有什么影响gydF4y2Ba肺功能gydF4y2Ba×BMI分析和调整身高不太可能在BMI和肺功能之间引入碰撞偏差(gydF4y2Ba补充表S25-S27gydF4y2Ba).的LD模式gydF4y2BaHLAgydF4y2Ba区域是高度复杂的,我们报告的信号可能是标记的因果变量,而不是真正的因果变量,由于归责数据的限制。建议测序数据以确定因果变异gydF4y2BaHLAgydF4y2Ba地区。gydF4y2Ba
总之,本研究是首次对中国人群肺功能进行大规模GWAS分析。我们的研究通过利用大规模的中国和欧洲遗传队列,扩展了肺功能特征遗传景观的现有知识。我们应用单性状和交叉性状分析,确定了中国人群肺功能性状的新位点,肺功能和肥胖性状之间的共同遗传效应,中国和欧洲人群的遗传异质性,以及PRSgydF4y2Ba肺功能gydF4y2Ba×BMI的影响。这些新发现为中国人群肺功能的遗传基础以及肺功能与肥胖之间的遗传共享提供了更多的知识,这将促进后续的转化、临床和公共卫生研究。gydF4y2Ba
补充材料gydF4y2Ba
补充材料gydF4y2Ba
请注意:gydF4y2Ba补充材料不是编辑部编辑的,上传时是作者提供的。gydF4y2Ba
补充附录及图表gydF4y2Baerj - 00199 - 2021。补充gydF4y2Ba
补充表gydF4y2Baerj - 00199 - 2021。表gydF4y2Ba
可共享的PDFgydF4y2Ba
确认gydF4y2Ba
本研究使用中国嘉orie生物银行(CKB)和英国生物银行(UKB)(申请号16549和45052)进行。我们非常感谢参与这项研究的参与者,以及CKB和UKB的研究团队成员。我们感谢GIANT和Biobank Japan财团提供的全基因组关联研究汇总统计数据。我们也感谢Zachary Schwartz (Harvard T.H. Chan公共卫生学院,波士顿,马萨诸塞州,美国)的语言编辑协助。gydF4y2Ba
脚注gydF4y2Ba
这篇文章有补充资料可从gydF4y2Bawww.qdcxjkg.comgydF4y2Ba
本文有编辑评论:gydF4y2Bahttps://doi.org/10.1183/13993003.01615-2021gydF4y2Ba
作者贡献:朱哲、余成、梁良是本研究的设计者。所有作者都获得或解释了数据。朱震、李俊、马波和史宏进行了统计分析。李磊、曹玮、余成、吕杰、郭英等获得资助。所有作者审阅并批准了手稿。gydF4y2Ba
利益冲突:Z. Zhu没有什么可透露的。gydF4y2Ba
利益冲突:李j没有什么可透露的。gydF4y2Ba
利益冲突:J. Si没有什么可透露的。gydF4y2Ba
利益冲突:马云没有什么可透露的。gydF4y2Ba
利益冲突:H. Shi没有什么可透露的。gydF4y2Ba
利益冲突:吕j没有什么可透露的。gydF4y2Ba
利益冲突:曹文伟没有什么可透露的。gydF4y2Ba
利益冲突:郭宇没有什么可披露的。gydF4y2Ba
利益冲突:I.Y.米尔伍德没有什么可透露的。gydF4y2Ba
利益冲突:r。g。沃尔特斯没什么可透露的。gydF4y2Ba
利益冲突:林凯没有什么可透露的。gydF4y2Ba
利益冲突:L. Yang没有什么可透露的。gydF4y2Ba
利益冲突:陈颖没有什么可透露的。gydF4y2Ba
利益冲突:杜h没有什么可透露的。gydF4y2Ba
利益冲突:B. Yu没有什么可透露的。gydF4y2Ba
利益冲突:长谷川在提交的工作之外报告了来自NIH和诺华的资助。gydF4y2Ba
利益冲突:小C.A.卡马戈没什么可透露的。gydF4y2Ba
利益冲突:M.F. Moffatt没有什么可透露的。gydF4y2Ba
利益冲突:W.O.C.库克森没有什么可透露的。gydF4y2Ba
利益冲突:陈杰没有什么可透露的。gydF4y2Ba
利益冲突:Z. Chen没有什么可透露的。gydF4y2Ba
利益冲突:L. Li没有什么可透露的。gydF4y2Ba
利益冲突:C. Yu没有什么可透露的。gydF4y2Ba
利益冲突:L. Liang没有什么可透露的。gydF4y2Ba
支持声明:本工作得到国家重点研发计划项目(2016YFC1303904、2016YFC0900500、2016YFC0900501、2016YFC0900504)和国家自然科学基金项目(81941018、91846303、91843302)的支持。中国嘉道理生物库基线调查及首次重新调查获嘉道理慈善基金会拨款支持。长期随访由英国惠康信托基金(212946/Z/18/Z, 202922/Z/16/Z, 104085/Z/14/Z和088158/Z/09/Z),中国国家自然科学基金(81390540,81390541和81390544)和中国科技部(2011BAI09B01)资助。资助者在研究设计、数据收集、数据分析和解释、报告的撰写或文章发表的决定中没有任何作用。本文的资助信息已存入gydF4y2Ba交叉参考基金注册gydF4y2Ba.gydF4y2Ba
- 收到了gydF4y2Ba2020年10月19日。gydF4y2Ba
- 接受gydF4y2Ba2021年3月2日。gydF4y2Ba
- 版权所有©作者2021。gydF4y2Ba
本版本根据知识共享署名非商业许可4.0的条款发布。为商业复制权利和权限联系gydF4y2Ba权限在}{ersnet.orggydF4y2Ba
参考文献gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba










